Глоссарий анатомии динозавров
В этом глоссарии разъясняются технические термины, обычно используемые при описании окаменелостей тел динозавров . Помимо терминов, специфичных для динозавров , он охватывает термины более широкого использования, когда они имеют центральное значение в изучении динозавров или когда их обсуждение в контексте динозавров полезно. Глоссарий не охватывает ихнологические и костно-гистологические термины, а также не охватывает измерения.
А
[ редактировать ]- блюдо
- У динозавров вертлужная впадина (множественное число: вертлужная впадина ) или тазовая впадина представляет собой отверстие в тазу, образованное подвздошной , лобковой и седалищной костью , которое видно на боковой и медиальной проекциях. В нем размещается головка бедренной кости , образуя тазобедренный сустав. У большинства четвероногих имеется закрытая вертлужная впадина , у которой лунка полностью заполнена костью , образуя углубление. Динозавры уникальны тем, что имеют перфорированную или открытую вертлужную впадину , где лунка на всю длину представляет собой отверстие без заполнения кости. [1]
- акромион
- Акромион — это костный гребень на внешнем дистальном конце лопатки, который служит для прикрепления ключицы . [2] [3] У нодозаврид развивается выраженная шпора, известная как псевдоакромион вероятно, образует место прикрепления передней мышцы лопатки и, следовательно аналогична акромиону , млекопитающих , которая , . [4]
- приводящая ямка
- Приводящая ямка или меккелевое отверстие у рептилий и динозавров является основным отверстием нижней челюсти , расположенным между областью, несущей зубы , и челюстным сочленением. Он открывается дорсально и ограничен латеральной стенкой надугловой кости , а медиальной – предсуставной ; поскольку последняя обычно намного ниже первой, ямка видна медиально. Дно отверстия образовано угловатой поверхностью и вмещает заднюю часть меккелева хряща. Приводящая ямка и окружающие ее края служат местом прикрепления основных приводящих мышц, закрывающих челюсть; это также позволяет главному нерву, артерии и вене челюсти проникнуть внутрь челюсти. [5]
- воздушные мешки
- У современных птиц легочные воздушные мешки представляют собой тонкостенные полупрозрачные, наполненные воздухом мешки, соединенные с легкими . Вместе с легкими воздушные мешки образуют высокоэффективную дыхательную систему, которая у птиц способна извлекать до 160% больше кислорода, чем это возможно у млекопитающих. Хотя внутри воздушных мешков происходит лишь ограниченный газообмен , они обеспечивают вентиляцию легких. Воздушные мешочки, расположенные как сзади, так и спереди от легкого, обеспечивают постоянный поток воздуха через легкое. Меньшие и слепые наполненные воздухом мешки, простирающиеся от воздушных мешков и легких, известны как пневматические дивертикулы (единственное число: дивертикул ); они могут быть многочисленными и присутствовать на большей части тела. Некоторые пневматические дивертикулы резорбируются и проникают в кости по мере роста животного, создавая ямки (впадины) на поверхности костей, а также внутренние камеры внутри костей - процесс, известный как посткраниальная пневматизация скелета (ПСП). Хотя воздушные мешки не окаменевают, на их присутствие, по крайней мере, у ящеротазовых динозавров, указывают характерные следы пневматизации в костях. [6]
- альвеолы
- Зубные альвеолы (единственное число: альвеолы ) или зубные лунки представляют собой ямки в челюстях , содержащие корни зубов . Внутри альвеол зубы периодически заменяются новыми, растущими под ними; отпадение старой коронки зуба происходит после резорбции корня. Состояние глубоких альвеол известно как текодонт и присутствует во всех архозавриях , включая динозавров. большинство чешуйчатых зубов С другой стороны, демонстрируют состояние плевродонта , когда зубы срастаются с внутренними сторонами костей челюсти, или акродонтный зубной ряд , при котором зубы срастаются с верхней частью костей челюсти без гнезд. Состояние текодонта исторически использовалось для определения клады Thecodontia , которая сейчас считается парафилетической и, следовательно, устаревшей. [7]
- угловой
- Угловая кожная — кость нижней челюсти . При виде сбоку он охватывает большую площадь задне-вентральной области челюсти, располагаясь позади зубной кости и ниже надугловой кости . При медиальной проекции нижней челюсти она видна ниже предсуставной кости . Он образует дно приводящей ямки и поддерживает заднюю часть меккелева хряща . [5]
- анторбитальное окно
- Анторбитальное окно — одно из пяти основных отверстий черепа , расположенное между глазницей и внешней ноздрей . Присутствует у большинства ранних Archosauriformes (включая Archosauria ), примитивно присутствует у динозавров; у ящеротазовых он имеет тенденцию быть большим, но у птицетазовых он редуцирован или полностью закрыт. Анторбитальное окно находится внутри более крупного углубления — анторбитальной ямки . Эта ямка может содержать дополнительные отверстия меньшего размера, а именно верхнечелюстное окно (также: добавочное предглазничное окно ) и промаксиллярное окно . [1]
- суставной
- Суставная — самая задняя кость нижней челюсти . На своей дорсальной поверхности он содержит гленоид — углубление в которое входит нижний конец квадратной кости черепа , , образуя челюстной сустав. Это единственная эндохондральная кость нижней челюсти, образующаяся непосредственно из задней части меккелева хряща . У млекопитающих суставная часть мигрировала в череп, образуя молоточек среднего уха . [5]
- ясно сформулированный
- Сочленение – это любое соединение между костями . Этот термин также используется для описания сохранности образцов: в сочлененном образце отдельные кости слипаются в исходном анатомическом положении. И наоборот, кости разъединенного скелета выходят из своего первоначального анатомического состава. Образец, кости которого разъединены, но находятся в непосредственной близости друг от друга, называется ассоциированным скелетом. Переход от полностью сочлененного скелета к скоплению изолированных и не связанных между собой костей происходит плавно. [8]
- астрагал
- Астрагал (множественное число: астрагал [1] ) — основная кость голеностопного сустава . Он расположен непосредственно под большеберцовой костью и медиальнее пяточной кости , которая находится ниже малоберцовой кости . Таранная кость не вращается относительно пяточной кости или большеберцовой кости; вместо этого у двуногих он часто слит с обоими элементами (см. также: tibiotarsus ). [9]
- атлас
- Атлант – первый (передний) позвонок позвоночного столба . Он принимает затылочный мыщелок основания черепа , образуя таким образом связь между черепом и позвоночником . У динозавров были одиночные мыщелки, обеспечивающие вращательное движение между атласом и черепом. У млекопитающих, напротив, наблюдаются двойные мыщелки, ограничивающие движение одной плоскостью; здесь вращательное движение преимущественно происходит между атлантом и вторым позвонком, аксисом . Атлас динозавров примитивен и состоит из трех отдельных элементов, отражающих базальное состояние у рептилий: атлантальный нейропофиз (множественное число: атлантальные нейропофизы; также: атлантальная нервная дуга ) вверху; [10] промежуточный центр спереди и центр сзади. Дополнительная небольшая кость, проатлас , расположена перед нервной дугой. [11] Центральную часть атласа также называют зубчатым отростком . [12] У динозавров эти три элемента обычно остаются отдельными, но могут сливаться вместе. У цератопсов атлас, а также ось и третий позвонок срослись в единую кость . [11]
- аркто-метатарзальный
- При аркто-метатарзальном состоянии (от лат. arctus – «сжатый») средняя (третья) плюсневая кость стопы защемлена между окружающими плюсневыми костями (второй и четвертой) на своем верхнем конце, исчезая при этом при виде спереди. Это состояние встречается у некоторых производных теропод, включая орнитомимид, тираннозаврид, троодонтид, элмизаврид и авимимид, и предполагалось, что оно действует как амортизатор во время бега, обеспечивая более высокую степень беглости . Первоначально этот признак использовался для определения клады Arctometatarsalia, которая сейчас считается полифилетической . [13] [14]
- ось
- Аксис — второй позвонок позвоночника , следующий за атлантом . Он крупнее последнего, его центр слит с нервной дугой , а нервный отросток хорошо развит. Его небольшой интерцентр, расположенный впереди центра, у теропод обособлен, но у птицетазовых сливается с центром атласа. [11]
- аксония
- Термин «аксония» описывает расположение доминантного (самого большого и выступающего) пальца руки или ноги и чаще всего используется при описании следов. Когда центральный палец (обычно палец III) доминирует, как в случае со стопами большинства двуногих динозавров, кисть или стопа являются мезаксонными . Энтаксоническая рука или стопа имеют доминантный палец посередине (первый или второй палец); это редкое состояние можно обнаружить на стопах зауроподов. И наоборот, эктаксоническая рука/стопа имеет доминантный палец снаружи (часто палец IV). Когда присутствуют два или четыре пальца и ни одна половина руки или ноги не является более доминирующей, чем другая, состояние называется параксоническим . [15] : 47–48


Б
[ редактировать ]- базальная трубка
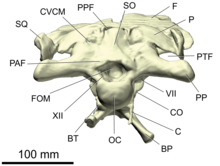
- Базальный бугорок (единственное: базальный бугорок ) или клиновидно-затылочные бугорки. [16] представляют собой пару бугоркообразных расширений на нижней стороне черепной коробки , которые служат местами прикрепления вентральных мышц шеи . Они образованы основной затылочной и базисфеноидной костями и выражены сильно у ящеротазовых и умеренно у птицетазовых. [17] [18]
- базикраний
- Базикраниум — это дно черепной коробки , образованное базиокципитальной , базисфеноидной и парасфеноидальной частями черепа . [19]
- базисфеноид
- Базисфеноидная кость образует переднюю часть дна черепной коробки . Невидимый снаружи в сочлененном черепе , он сращен с основной затылочной костью сзади и с парасфеноидом спереди. Пара характерных отростков, отростков , простирается от его нижней стороны и сочленяется с крыловидными отростками неба базиптеригоидных . [20]
- базиокципитальный
- Основная затылочная кость — непарная кость черепа и одна из четырёх костей, образующих затылочную кость черепной коробки . Он виден на сочлененном черепе, вид сзади. Он образует большую часть затылочного мыщелка , округленного отростка, соединяющего череп с , первым позвонком шеи атлантом . Сверху он слит с парными экзоципитальными костями . Он также образует заднюю часть дна черепной коробки, сливаясь с базисфеноидом спереди. [20]
- кость
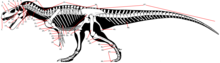
- Кости, помимо зубов , являются наиболее распространенными окаменелостями тела динозавров. Кости, состоящие из гидроксиапатита , твердо минерализованы и поэтому прочны. Мягкие части, напротив, могут сохраняться только как отпечатки, а непосредственно сохраняются только в исключительных случаях. К костным частям относятся череп , осевой скелет (позвоночник и ребра ) и добавочный скелет ( пояса и конечности ). [21] Большинство этих костей парные , с зеркально перевернутым аналогом на другой стороне тела, или непарные , и в этом случае они обычно лежат на средней линии тела и делятся последней на две симметричные половины. [1] Кроме того, кости включают ряд элементов, образованных в коже, таких как гастралии , костные щитки и шипы . [21] Палеонтологи чаще всего изучают морфологию костей, но также их гистология (внутренняя микроструктура вплоть до клеточного уровня) и химический состав дали важную информацию о биологии динозавров. [22] Существует два основных типа костей: Дермальная кость формируется непосредственно в дерме (коже), обычно вырастая из изначально тонких пластинок. Среди прочих большинство костей, образующих наружную поверхность черепа и нижней челюсти, являются кожными костями. Напротив, эндохондральная кость формируется из хрящевого предшественника, который окостеневает (превращается в кость). [1]
- черепная коробка
- Черепная коробка – это часть черепа, в которой находится мозг . В сочленённом черепе он не виден снаружи, за исключением задней части — затылка . Сложная структура, она пронизана многочисленными отверстиями, содержащими кровеносные сосуды и черепно-мозговые нервы. Отдельные кости черепной коробки у взрослых имеют тенденцию полностью срастаться, при этом границы между исходными элементами часто не видны. У динозавров анатомия черепной коробки консервативна, но по этой причине ее можно использовать для вывода о взаимоотношениях группы, когда другие особенности скелета претерпели столь глубокие изменения, что их происхождение уже невозможно проследить. Черепная коробка также может позволить реконструировать мозг и внутреннее ухо с выводами о чувствах и интеллекте. [20]


С
[ редактировать ]- каблук
- Пяточная кость (множественное число: пяточная кость) [1] ) — основная кость голеностопного сустава (в задней части стопы) и вместе с таранной костью образует верхний ряд костей предплюсны. Он расположен латеральнее таранной кости и дистальнее малоберцовой кости . [9]
- голова
- Caputegulae (лат. «черепные плитки») — плоские кости, покрывающие кости черепа анкилозавров. Вместе с рогами пирамидальной формы они образуют орнамент черепа. Придуман Уильямом Т. Блоузом в 2001 году. [23] этот термин может использоваться для элементов, представляющих как кооссифицированные остеодермы, так и черепную скульптуру. Положение головки черепа можно указать с помощью модификаторов; например, носовые головки располагаются на носовых костях. [24]
- кили
- Кили (единственное число: carina ) представляют собой гребни эмали, образующие режущие края на переднем и заднем краях зубов. Обычно они встречаются у хищных динозавров и часто имеют зазубрины . [25] : 41
- запястья
- Кости запястья образуют запястье или запястье , которое соединяет предплечье ( лучевую и локтевую кости ) с пястными костями кисти. [26] Соответствующая часть стопы — предплюсна . У базальных рептилий имеются три ряда запястий. У динозавров запястье часто не полностью окостенело, а количество и идентичность запястных элементов во многих случаях остаются неясными. Особенно хорошо закостенело запястье у базальных птицетазовых Heterodontosaurus , где оно состояло из девяти элементов. Проксимальный ряд состоял из радиальной (ниже лучевой кости), локтевой (ниже локтевой кости) и гороховидной (небольшой элемент ниже локтевой кости и латеральнее локтевой кости). Дистальный ряд состоял из пяти элементов, расположенных над пятью плюсневыми костями, которые обозначаются как дистальные запястные кости 1–5 . Средняя строка представлена одним элементом — Centrale . [27] [28]
- карпометакарпус
- Запястно-пястная кость — костный элемент кисти, состоящий из сросшихся запястных и пястных костей . Эта структура встречается у современных птиц, но присутствовала также у некоторых теропод, тесно связанных с птицами, и развивалась независимо у альваресзаврид . [26]
- каудальный
- Каудальные позвонки (от лат. caudum — хвост), или хвостовые позвонки, — это позвонки, составляющие хвост. [26] У предков динозавров было около 50 хвостовых позвонков, хотя их количество, размеры и форма значительно различались в отдельных группах. Число хвостовых хвостов уменьшалось по эволюционной линии, ведущей к современным птицам; в последнем оставшиеся хвостовые части слиты в пигостиль . Хотя у ранних динозавров они были гибкими, некоторые клады укрепляли свой хвост с помощью окостеневших сухожилий (как у многих птицетазовых) или удлиненных презигапофизов (как у некоторых теропод, таких как дромеозавриды ). Диплодоцидные зауроподы имели удлиненный хлыстовидный хвост, а у некоторых птицетазовых хвосты снабжены булавами, шипами и/или пластинами. [25] : 45
- каудофеморальный
- Мышцы хвостобедренного сустава, или Musculus caudofemoralis , являются основными двигательными мышцами у всех длиннохвостых динозавров. Расположенные в основном в хвосте, они тянут бедренную кость задней конечности назад при сокращении (втягивание бедренной кости/разгибание бедра), обеспечивая тем самым движение. У длиннохвостых динозавров Musculus caudofemoralis исключительно велик и может составлять 58% от общей массы хвоста тираннозавра . У динозавров выражены две части: короткая мышца caudofemoralis возникла на нижнем крае задней части подвздошной кости, а длинная мышца caudofemoralis возникла на передней части хвоста, вентральнее поперечных отростков и под поверхностной гипаксиальной мускулатурой хвоста. . Обе части прикрепляются к четвертому вертелу на задней поверхности бедренной кости. Мышцы хвостобедренного сустава сократились в ходе эволюции теропод и по большей части утрачены у современных птиц. Птицы, в отличие от длиннохвостых динозавров, не полагаются на втягивание бедренной кости для движения, а вместо этого втягивают голень вокруг коленного сустава. [4] [29]
- центроцель
- Центроцель — это кавернозная полость костного мозга внутри центра позвонка. [30] : 31, 47
- центропрезигапофизарная ямка
- Центропрезигапофизарная ямка представляет собой углубление на передней стороне ножок нервных дуг, ограниченное центропрезигапофизарной пластинкой. [31]
- центр
- Центр (множественное число: centra ), также тело или корпус позвонка , представляет собой элемент в форме катушки или цилиндра, который вместе с нервной дугой образует позвонок . [1] У молодых особей и центры, и нервные дуги являются отдельными элементами, и слияние этих элементов является важным критерием определения взрослости. Передняя и задняя поверхности центра образуют сочленение с центрами предыдущего и последующего позвонка. Центры можно классифицировать на основе морфологии суставных поверхностей:

Классификация морфологии центров на основе их суставных концов у динозавров. - амфикоэлозные (существительное: амфикоэлии ) центры сильно вогнутые с обоих концов.
- platycoelous (существительное: platycoely ) слегка вогнутые на обоих концах.
- амфиплатийные (существительное: амфиплатия . Также: ацелозные ) центры плоские с обоих концов.
- процельные (существительное: процели ) центры спереди вогнуты, а сзади выпуклы.
- opisthocoelous (существительное: opisthocoely ) центры сзади вогнуты, а спереди выпуклы.
- гетероцельные (существительное: гетероцелие ) центры имеют седловидную форму на обоих концах.
Амфикоэлия — примитивное состояние четвероногих. У рыб концы центров глубоко выемчаты и соединены через небольшое отверстие — проход для хорды . У рептилий этот тип центров имеется у зародышей, а также у взрослых форм некоторых видов; у большинства видов, включая динозавров, центры более окостенели, а нотохордальное отверстие закрыто, что повышает устойчивость к силам сжатия. Гетероцельные позвонки обеспечивают гибкость, предотвращая вращение. Процельные и опистоцельные центры образуют вогнуто-выпуклые ( шаровидные ) суставы, в которых выпуклый конец, мыщелок , соединяется с вогнутым концом, семядолей (также: семядолей ). Такая конфигурация обеспечивает большую стабильность без ограничения мобильности. У длинных шей и хвостов эта стабилизация работает лучше всего, когда выпуклая часть направлена от тела. Таким образом, у зауроподов позвонки перед крестцом обычно опистоцельные, а позвонки хвоста - процельные. Поскольку позвоночник может иметь разные типы центральной морфологии, могут возникать переходные центры с двумя концами разной формы. [11] [32] [33] [34]
- мозжечок
- Мозжечок («маленький мозг») — это дорсальная часть заднего мозга между стволом мозга и головным мозгом, служащая для контроля равновесия, позы и движения. Эту часть мозга обычно невозможно наблюдать у окаменелостей динозавров, поскольку ее редко можно увидеть на эндокастах ; Исключением является возможная сохранность мозжечковых листков у Конхораптора . [35] [36] [26]
- шейный
- Шейные позвонки, или шейные позвонки, являются шейными позвонками. [26] У большинства динозавров было от 9 до 10 шейных позвонков, хотя в некоторых группах, включая зауроподов, их число было достигнуто как за счет увеличения количества позвонков, так и за счет интеграции спинных позвонков в шею. [25] : 44
- шейное полукольцо
- Шейное полукольцо — поперечно ориентированный воротниковидный ряд остеодерм, защищающий верхнюю сторону шеи у анкилозавров (нижняя сторона шеи остается голой). У особи обычно было два шейных полукольца, переднее из которых было меньше заднего. Шейные полукольца являются синапоморфией Ankylosauria. [37]
- шейное ребро
- Шейные ребра — это кости, которые прикрепляются латеро-вентрально к шейным позвонкам. Они плезиоморфны для амниот (хотя и утрачены у млекопитающих) и включают передний и задний отростки. Некоторые зауроподоморфы, особенно некоторые зауроподы с длинной шеей, такие как Giraffatitan , обладали сверхудлиненными шейными ребрами с задними отростками, перекрывавшими два или три предыдущих позвонка. Гистологический анализ костей показал, что эти удлиненные отростки представляют собой окостеневшие сухожилия , а это означает, что их концы были соединены с мышцами. Большая длина отростков увеличила бы расстояние между телом мышцы и позвонком, которым управляет мышца, что позволило бы мышце располагаться близко к телу, облегчая шею. [38] [39]
- шеврон
- Шевроны, или гемальные дуги , представляют собой кости, прикрепленные к нижней стороне хвостовых позвонков , образующие вентральную поверхность хвоста. Шеврон состоит из левой и правой частей, которые обычно соединены друг с другом V-образно, образуя большое отверстие, которое можно увидеть спереди или сзади. Отверстия нескольких последовательных шевронов образуют гемальный канал хвоста, который защищает нервы и кровеносные сосуды. [26] [25] : 46
- хоаны
- Хоаны (единственное число: хоана), или внутренние ноздри (единственное число: внутренние ноздри ), представляют собой пару отверстий в нёбе, которые продолжаются с внешними ноздрями или ноздрями, образуя носовой ход. Левое и правое отверстия пары разделены по средней линии черепа сошником . У динозавров хоаны обычно очень большие и удлиненные. [25] : 39 [40]
- поясная кость
- Поясная часть (множественное число: cingula ) — это выступ в форме полки, окружающий основание коронки зуба . [41]
- ключицы
- Ключицы, также ключицы (единственное число: clavicula ) или ключицы, представляют собой пару изогнутых изогнутых костей, расположенных над клювовидным отростком плеча. Эти кожные кости прикреплены к акромиальному отростку лопатки и лучше всего видны на виде скелета спереди. Ключицы нечасто встречаются со скелетами динозавров, что, вероятно, связано с их низкой сохранностью . [42] [25] : 49
- коракоид
- Коракоид — парная кость грудного пояса, прикрепляющаяся к нижнему концу лопатки . Плоский элемент, у динозавров обычно имеющий форму от полукруга до квадрата, эндохондрального происхождения. Ее нижний край вместе с лопаткой образует часть суставной кости плечевого сустава. [25] : 47–49 На боковой поверхности клювовидного отростка имеется отверстие — клювовидное отверстие , через которое проходит супракоракоидный нерв. [3]
- венечный отросток
- Венечный отросток представляет собой костный выступ, идущий вверх от верхней поверхности нижней челюсти за зубным рядом. Этот отросток служит местом прикрепления мышц, закрывающих челюсть; более высокий венечный отросток увеличивает плечо рычага этих мышц и, следовательно, силу укуса. [43] [26] ( см. рисунок здесь )
- краниальный кинезис
- Краниальный кинезис — это способность частей черепа двигаться друг против друга в суставах черепа. Хотя краниальный кинез был предложен для ряда нептичьих таксонов динозавров, обзор 2008 года показал, что большинство этих выводов проблематичны. [44] Предполагаемые формы краниального кинезиса, встречающиеся у динозавров, включают:
- стрептостилия : движение квадратной кости вокруг чешуйчатой кости ушного сустава. Квадратная мышца в основном движется параллельно средней линии черепа, хотя могут быть задействованы и поперечные движения. [44]
- прокинез : движение между черепной коробкой и лицевым скелетом вокруг носолобного сустава (сочленения между носовой и лобной костью) или движение между левой и правой носовой частью. Этот тип встречается у современных птиц. [44]
- нейрокинез (также: кинезис базального сустава ): движение черепной коробки против неба вокруг базиптеригоидного сустава (или базального сустава). [44]
- плеврокинез : способность верхней челюсти поворачиваться наружу. Плеврокинез был предложен как новый тип краниального кинеза для орнитопод, позволяющий использовать поперечные движения челюсти при жевании. Эта сложная форма краниального кинеза требует, помимо подвижности верхней челюсти и лицевого скелета, поперечных движений квадратной кости относительно чешуйчатой кости (модифицированная форма стрептостилии), а также кинеза базальных суставов. [44]




Д
[ редактировать ]- поза смерти
- Поза смерти , или опистотоническая поза , — это распространенная поза, наблюдаемая у сочлененных окаменелостей динозавров и птиц, где позвоночник сильно изогнут вверх (перерастянут). [45] Обсуждаются различные возможные причины, в том числе опистотонус (предсмертные муки). [45] и высвобождение межламинарной эластичной связки во время разложения. [46]
- стоматологическая батарея
- — Зубная батарея это тип зубного ряда некоторых травоядных динозавров, в котором отдельные зубы плотно прилегают друг к другу, образуя непрерывную шлифовальную поверхность. У гадрозаврида Edmontosaurus зубная батарея каждой половины верхней и нижней челюстей содержит более 60 рядов зубов, причем каждый ряд включает до трех функциональных зубов, расположенных друг на друге, и до пяти замещающих зубов под этим набором, которые могут прорезаться. из челюстей, когда функциональные зубы изнашивались и выпадали. Самый сложный тип зубного ряда у динозавров, он независимо развился у гадрозаврид, цератопсов и некоторых зауроподов и различается по форме и функциям в этих отдельных кладах. [25] : 185–186 [47]
- зубной ряд
- Зубная кость — основная кость нижней челюсти . Это единственная кость нижней челюсти, несущая зубы, и она расположена впереди всех остальных костей челюсти, за исключением птицетазовых, у которых кончик нижней челюсти образован предзубной челюстью . Передними концами зубные кости левой и правой челюсти соединяются между собой, образуя нижнечелюстной симфиз . [25] : 40
- зубной ряд
- Зубной ряд — это собирательный термин, обозначающий все зубы, имеющиеся в челюстях отдельного динозавра. Зубной ряд может быть гомодонтным , когда имеется только один тип зубов, или гетеродонтным, когда зубы разных типов. [48] : 232–233 Зубы постоянно заменяются в течение жизни. Зубы, которые прорезались и используются в настоящее время, называются функциональными зубами . Для каждого положения зуба обычно в любой момент времени имеется один или два непрорезавшихся замещающих зуба , которые последовательно мигрируют в лунку зуба и заменяют функциональный зуб после его выпадения. У некоторых травоядных видов может быть до шести сменных зубов на каждую позицию. Обмен зубов начался с резорбции корня функционального зуба. [49] Зубы, выпавшие после замены, не имеют корня и называются выпавшими зубами . [41]
- дельтопекторальный гребень
- Дельтопекторальный гребень представляет собой направленный вперед костный выступ на верхней части плечевой кости . Особенно длинный и выступающий дельтопекторальный гребень является синапоморфией динозавров , т. е. признаком, отличающим группу от других групп. У динозавров гребень составляет 30–40% длины плечевой кости. Он обеспечивал поверхности прикрепления мышц плеча и груди (соответственно дельтовидной и грудной мышц), которые при сокращении притягивали руку к туловищу. [25] : 14
- цифры
- Пальцы, или пальцы рук и ног, образуют дистальную часть аутоподия , следующую за пястью кисти и плюсной стопы. Они обозначаются римскими цифрами от I до V, где I обозначает самую внутреннюю цифру, а V - самую внешнюю. Отдельные пальцы имеют одну или несколько фаланг (кости пальцев рук и ног). [1] : 145
- долихолиакальный
- Долихойляк относится к одной из двух основных конфигураций бедренной кости у ранних ящеротазовых динозавров. Долихоподвздошный тип характеризуется большими передними и задними расширениями подвздошной кости, а также удлиненными и узкими лобковыми и седалищными костями. Напротив, брахиподвздошный тип характеризуется короткими расширениями подвздошных костей и более крепкими лобковыми и седалищными костями. Брахиподвздошный тип характерен для базальных зауроподоморфных динозавров триаса, а долихойилакный тип характерен для теропод. Оба термина были введены Эдвином Колбертом в 1964 году. [50] [51]
- спины
- Спинные позвонки, также называемые спинными позвонками или задними позвонками, представляют собой позвонки задней области между шейными отделами шеи и крестцовыми частями бедра. В терминологии Nomina Anatomica Avium (NAA) вместо этого используется термин грудные позвонки . У млекопитающих термин «грудные позвонки» относится только к несущим ребра позвонкам грудной клетки, тогда как позвонки нижней части спины, у которых отсутствуют ребра, называются поясничными позвонками . Это подразделение спинного отдела позвоночника неприменимо у динозавров, а термин NAA «грудные позвонки» относится к спинному отделу позвоночника в целом. [1] : 143 У большинства динозавров было около 15 спинных костей. По сравнению с шейными спинные кости короче, нервные отростки у них длиннее, а поперечные отростки более крепкие и направлены наружу. [25] : 45

И
[ редактировать ]- эктоптеригоидный
- Эктоптеригоид — меньшая кость неба . Парный и дермальный по происхождению, он соединяется со скуловой костью латерально и с крыловидной костью медиально. [25] : 39–40
- эндокаст
- Эндокасты представляют собой заполнения (слепки) нервных полостей, включая черепную коробку и нервный канал позвонков. Таким образом, они могут фиксировать внешние особенности нервных структур, присутствующих в этих полостях, в первую очередь мозга. Однако эндокасты не являются точными копиями нервных структур, поскольку нервные полости обычно содержат дополнительную ткань, которая может скрывать морфологию нервной структуры. Естественные эндокасты образуются, когда нервная полость заполняется осадком, тогда как искусственные эндокасты могут быть изготовлены с использованием отливочного материала. [35] : 192
- эпиюгальный
- Эпижугалия — это кожное окостенение, уникальное для цератопсов. Он закрывает нижний конец обращенного вниз треугольного фланца скуловой кости, типичного для этой группы. У Arrinoceratops и Pentaceratops надскуловая кость образует отчетливый скуловой рог. Кость можно найти у ямацератопса и всех цератопсов, более продвинутых, чем последние. [52] : 500 [53] : 538
- эпокципитальный
- Эпокципитали — кожные окостенения, выстилающие края оборок цератопсов. Эпокципитальные кости у молодых особей представляют собой отдельные кости, но у взрослых они срастаются либо с чешуйчатой , либо с теменной костью , в зависимости от их положения. Эпокципитали чешуйчатой кости также известны как эписквамозные кости , а теменные кости - эпипариетальные кости . Некоторые авторы выступали против использования термина «эпокципитальная», поскольку эти кости не имеют связи с затылочной областью черепа, вместо этого они предпочитали термины «эписквамозная» и «эпипариетальная». Эпокципитали были декоративными, а не функциональными, и сильно различались по форме, образуя у центрозавринов сильно увеличенные шипы. [54] [52] : 502 [53] : 540
- эпипофиз
- Эпипофизы — это костные выросты шейных позвонков, обнаруженные у динозавров и некоторых ископаемых базальных птиц. Эти парные отростки располагаются над постзигапофизами позвонка в задней части нервной дуги . Их морфология разнообразна и варьируется от небольших простых холмообразных возвышений до больших сложных крыловидных выступов. Эпипофизы обеспечивали большие места прикрепления нескольких мышц шеи; Таким образом, крупные эпипофизы указывают на сильную мускулатуру шеи. Наличие эпипофизов считается синапоморфией динозавров. [25] : 14
- эпиоссификации
- Эпиоссификации (также называемые эпиоссификациями) — это дополнительные окостенения, прикрепленные к черепам цератопсов . [54] Они включают в себя:
- Ростральная . , дополнительная кость перед предчелюстной костью верхней челюсти
- кость Эпиназальная , образующая роговую сердцевину рога носа.
- Эпижугалы — пара костей, которые часто образуют выступающие скуловые рога.
- Эпипариетальные — и эписквамозные кости небольшие окостенения, выстилающие края воротника шеи (теменная и чешуйчатая кости соответственно). Эти окостенения также называются эпокципитальными . [54]
- эпиптеригоидный
- Эпиптеригоид — кость парная эндохондральная в задней части неба . Небольшая пластинчатая кость, соединяющаяся с крыловидной костью и черепной коробкой . [25] : 39–40
- экзоципитальный
- Экзозатылочная кость — парная кость, которая вместе с непарными надзатылочной и основной затылочной костью образует затылочную кость в задней части черепа. У взрослых динозавров экзоципитальные кости слиты с опистотическими , образуя структуру, также известную как экзоципитально-опистотические . Ниже экзозатылочные кости слиты с базиокципитальными . [20] [25] : 38
- внешние ноздри
- Наружные ноздри (единственное число: наружные ноздри , также: костные ноздри) . [55] ) представляют собой пару наружных отверстий черепа для ноздрей. Важные ориентиры черепа: они примитивно расположены перед анторбитальным окном, возле кончика рыла. Внешние ноздри иногда называют ноздрями; однако такое использование не рекомендуется, поскольку термин « ноздря» зарезервирован для обозначения мясистого отверстия носового прохода (также называемого «мясистая ноздря»). [55] ), который в некоторых случаях может лежать далеко за пределами внешней ноздри. [1] : 139 Отверстия носового хода в ротовую полость, наоборот, называются внутренними ноздрями , или хоанами. [25] : 39 Часто увеличенная передняя часть полости носа, расположенная непосредственно под краями внешней ноздри, называется преддверием носа . [55]
Ф
[ редактировать ]- бедренная кость
- Бедренная кость (множественное число: femora ) или бедренная кость является проксимальным элементом задней конечности. Его проксимальная головка представляет собой большую головку бедренной кости , направленную медиально и образующую с диафизом угол 90°; головка входит в вертлужную впадину бедра, образуя тазобедренный сустав. [25] : 15 В верхнем отделе бедренной кости имеется ряд вертлугов (отростков прикрепления мышц):
- Малый вертел (также: передний вертел ) расположен на передней поверхности сразу под головкой. Это особенность, уникальная для динозавров и их непосредственных внешних групп: она закрепляет лобково-бедренные мышцы, которые тянут заднюю ногу вперед и внутрь и, следовательно, имеют решающее значение для передвижения. У многих ранних динозавров дополнительно был костный гребень или мышечный рубец, известный как вертельная полка , который простирается от основания переднего вертела к заднелатеральному краю бедренной кости. Малый вертел динозавров не гомологичен малому вертелу млекопитающих. [25] : 19 [4] : 179
- Большой вертел (также: дорсолатеральный вертел ) представляет собой гребень или выпуклость на латеральной поверхности, противолежащую головке бедренной кости и продолжающуюся с ней. [30] : 47 Он обеспечивал крепление как лобково-бедренной, так и седалищно-бедренной мышц. [4] : 181
- Четвертый вертел представляет собой обширный выступ на задней поверхности бедренной кости, направленный назад. У динозавров четвертый вертел отчетливо асимметричен, проксимальная часть более расширена, чем дистальная. Это синапоморфия Dinosauria. Четвертый вертел обеспечивает прикрепление длинной мышцы хвоста бедренной кости, наиболее важной двигательной мышцы, которая при сокращении тянет бедренную кость назад, обеспечивая движение. [25] : 16 [4] : 181–182
- малоберцовая кость
- ( Малоберцовая кость множественное число: fibulae ) или икроножная кость — длинная кость , расположенная латеральнее большеберцовой кости ; вместе две кости образуют голень. Обычно она более тонкая, чем голень, но особенно тонкая у производных теропод и птиц. [1] : 147 [25] : 53
- отверстие
- Отверстие (множественное число: foramina ) — это отверстие в кости для прохождения кровеносных сосудов, нервов, мышц и подобных образований. [26] Питательные отверстия — это отверстия, позволяющие кровеносным сосудам проникать внутрь костей, обеспечивая поступление питательных веществ в кость. [25] : 224
- фронтальный
- кость Лобная представляет собой парную кожную кость крыши черепа , которая лежит между носовой частью спереди и теменной сзади. Лобные кости обычно обширны и образуют верхний край глазницы, а также большую часть крыши черепной коробки . [25] : 38
- фонтаны
- Роднички (также: роднички ) — это промежутки между костями (обычно костями черепа), которые в основном присутствуют у эмбрионов, но часто закрываются по мере роста животного. [56] : 25 У цератопсов есть отверстия между лобной и теменной костями, называемые лобными родничками, которые представляют собой отверстия супракраниальной полости, что является уникальной особенностью этой группы. [52]
- ямка
- Ямка (множественное число: fossae ) — общий термин, обозначающий углубление или бороздку на поверхности кости. [26]

Г
[ редактировать ]
- Гастралия
- Гастралии (единственное число: gastralium ) — это кожные кости, находящиеся в дерме брюшной полости между грудиной и лобком . Гастралии примитивно присутствуют у динозавров и могут быть обнаружены у базальных зауроподоморфов и теропод , но независимо потеряны у зауроподов , птицетазовых и неорнитинов (сегодняшних птиц). Когда они присутствуют, они располагаются в от 8 до 21 ряда, каждый из которых состоит из четырех элементов: правый и левый медиальный гастралий сочленяется на средней линии тела зигзагообразным образом, а латеральный гастралий сочленяется с латеральным концом каждого из них. медиальный гастралий. Первый и последний ряд могут быть слиты в единую дугообразную структуру, называемую шевронным гастралом . Использование термина «брюшное ребро» , который иногда использовался для обозначения гастралии и других структур брюшной полости, неоднозначно, поэтому не рекомендуется. [57]
- гленоид
- Гленоид — это суставная поверхность, образованная клювовидным отростком и лопаткой , которая принимает головку плечевой кости , образуя плечевой сустав . Вогнутая и круглая структура, она окаймлена костным краем и расположена на задней поверхности клювовидного отростка и задне-дистальной поверхности лопатки. [3] [1] : 144
ЧАС
[ редактировать ]- все
- Пятка – это задняя часть стопы . Этот термин также относится к задней части следа, и в этом случае он не обязательно соответствует анатомической пятке, которая у многих динозавров приподнята над землей. [15] : 47
- рог
- Рога представляют собой выросты костей черепа, покрытые ороговевшей оболочкой. Костное ядро рога называется ядром рога . Поскольку ороговевшая оболочка обычно не сохраняется в окаменелостях, ее наличие у ряда динозавров было сделано путем сравнения с современными быками . Другие черепные выросты или складочки могут быть покрыты кожными структурами, как это было предложено, например, для носового выступа пахиринозавра и куполов пахицефалозавров . [58]
- плечевая кость
- ( Плечевая кость множественное число: плечевая кость ) — это единственная кость плеча и обычно самая большая кость передней конечности. [1] : 145 [25] : 49
- гипосфен-гипантральное сочленение
- Гипосфен-гипантральные сочленения представляют собой добавочные суставы, соединяющие соседние позвонки. Они состоят из отростка на задней стороне позвонка — гипосфены , который вписывается в углубление на передней стороне следующего позвонка — гипантрума . Гипосфен-гипантральные сочленения встречаются в спинных позвонках, а иногда также в задних шейных и передних хвостовых позвонках . [59]
- подъязычная мышца
- Подъязычные кости — это кости области горла ниже и позади черепа, которые поддерживают язык. Эти элементы редко встречаются у динозавров, так как они не сочленены с остальным скелетом и поэтому легко теряются. [4] : 155
- хайпекс
- Гипекс (множественное число: гипексы или хайписы ) — это вершина угла, образованного между двумя цифрами , или, другими словами, самая ближайшая точка пространства между двумя цифрами. Гипекс можно идентифицировать только при наличии мягких тканей, поэтому его в основном используют при изучении следов окаменелостей. [15] : 46–47

я
[ редактировать ]- подвздошная кость
- Подвздошная кость (множественное число: ilia ) — верхняя и самая крупная кость бедра. Он состоит из основного тела — лопатки подвздошной кости , медиальная сторона которой прикрепляется к крестцовым позвонкам . Вентрально он образует утолщенный изогнутый гребень, нависающий над вертлужной впадиной , надвертлужный гребень или гребень , распределяющий напряжения, передаваемые через бедренную кость задней конечности. Переднее расширение лопатки подвздошной кости называется предацетабулярным отростком (также: предацетабулярное крыло ), а заднее расширение - поствертлужным отростком (также: поствертлужное крыло ); [60] оба расширения позволяют оптимально разместить начало мышц конечностей. Перед вертлужной впадиной крепкий отросток, лобковая ножка , простирается вентрально и прикрепляется к лобковой кости, тогда как седалищная ножка проходит позади вертлужной впадины и прикрепляется к седалищной кости. [61] : 33 Непосредственно за вертлужной впадиной на седалищной ножке находится противовертел — суставная поверхность, которая вентрально доходит до седалищной кости и сочленяется с большим вертелом бедренной кости. [25] : 19 [62] : 726 На вентральной поверхности постацетабулярного отростка имеется углубление, называемое короткой ямкой , где начинается короткая мышца хвоста бедренной кости. [61] : 21 Латерально короткую ямку граничит с короткой полкой — гребнем , проходящим вдоль вентролатерального края подвздошной кости между седалищной ножкой и задним концом кости. [62] : 733–734
- седалищная кость
- Седалищная кость (множественное число: седалищная кость ) — одна из трех костей, составляющих таз, расположенная ниже подвздошной кости и позади лобковой кости . Это стержневидный элемент с расширенной полкой на проксимальном конце — запирательной пластинкой , контактирующей с подвздошной и лобковой костью (соответствующая запирательная пластинка имеется и в лобковой кости). Как и лобковая кость, левая и правая седалищные кости сращены по всей длине. Расширенный симфиз каудовентрально направленного стержня называется седалищным фартуком . [63] : 139 Дистальный конец седалищной кости часто расширен в седалищный башмак . Вентральная поверхность стержня имела продольный гребень — запирательный отросток . У базальных таксонов запирательный отросток расположен на дистальном конце седалищной кости, у целурозавров - на дистальном конце седалищной кости. [63] : 152
- подвисочное окно
- Подвисочное окно (также: латеральное височное окно ) — это главное отверстие на боковой стороне черепа позади глазницы . [1] : 140 Он ограничен заглазничной костью в верхней передней части, чешуйчатой костью в верхней части спины, квадратно-скуловой костью в нижней части спины и скуловой костью в нижнем переднем углу. [25] : 37
- межключица
- Межключица представляет собой одиночную стержневидную кожную кость, расположенную между ключицами в плечевом поясе. [42] Обычно считается, что межключицы, являющиеся примитивной особенностью четвероногих, отсутствуют у динозавров и утеряны у Dinosauriformes. [64] Однако наличие межключиц у нескольких таксонов динозавров неоднократно предполагалось. [42]
- межзубные пластины
- Межзубные пластинки (также: поперечные перегородки , морщины [30] ) — костные пластинки на язычном (внутреннем) крае зубного ряда, поднимающиеся от альвеолярного края челюсти и заполняющие область между зубами. [65] [66]

Дж
[ редактировать ]- скуловой
- Скуловая кость, или скуловая кость, представляет собой кость черепа, которая определяет нижнюю границу глазницы и соединяется с верхней челюстью спереди и с квадратноскуловой костью сзади. Его задний конец раздваивается в месте сочленения с квадратноскуловой костью, что считается синапоморфией динозавров. [1] : 140 [25] : 12
л
[ редактировать ]- слезный
- Слёзная кость представляет собой распорку в черепе, отделяющую анторбитальное окно от глазницы . Его пронизывает канал слезного протока (слезный проток), соединяющий орбиту с анторбитальным окном. У некоторых видов верхний конец слезной кости имеет треугольный гребень или рог, возвышающийся над крышей черепа . [25] : 37 [17] : 62
- пластинки
- Позвоночные пластинки (единственное число: пластинка ) или костные перегородки (единственное число: osseus septum ) представляют собой гребни на позвонках, которые разделяют пневмокамеры. Они начинаются и заканчиваются на разных отростках и участках позвонка в зависимости от их терминологии и называются по точкам начала и конца. Пластинки делятся на четыре произвольные категории в зависимости от их окончания: диапофизарные пластинки , парапофизарные пластинки , зигапофизарные пластинки и спинномозговые пластинки . Их также можно разделить на субламины , если они разделены по длине, или на составные пластинки, если более одной пластинки образуют гребень. Пример пластинок: [67]
- Спинопрезигапофизарная пластинка (SPRL) : пластинка, простирающаяся от нервного отростка ( спино- ) до презигапофиза ( -презигапофизарная ).
- Преспинальная пластинка (PRSL) : пластинка, простирающаяся вдоль передней поверхности ( пре- ) нервного отростка ( -спинальная ).
- Задняя центродиапофизарная пластинка (PCDL) : пластинка, простирающаяся от заднего конца ( заднего ) центра ( центро- ) до диапофиза ( -диапофизарная ).
- Интрапостзигапофизарная пластинка (TPOL) : пластинка, простирающаяся между ( интра- ) постзигапофизами ( -постзигапофизарная пластинка ).
- Презигопарапофизарная пластинка (ПРПЛ) : пластинка, простирающаяся от презигапофиза ( Презиго- ) до парапофиза ( -парапофизарно ).
- латеросфеноид
- Латеросфеноид (также: плевросфеноид ) — крыловидная кость передней части латеральной стенки черепной коробки . Незаметная снаружи, она расположена кпереди от проотической и сверху соприкасается с теменной , лобной и заглазничной . Латеросфеноид как отдельное окостенение уникально для архозаврообразных; у других рептилий эта часть нейрокраниального хряща не окостеневает. [25] : 39 [17] : 83 [20] [26]
- межламинарная эластичная связка
- ligamentum elasticum interlaminare (иногда сокращается до ligamentum elasticum ) — связка, состоящая из непарных пучков, соединяющих нервные отростки соседних позвонков у динозавров и птиц. [46] Эта связка охватывает шею и хвост, а у современных птиц поддерживает вогнутость шеи. [46] [68] Было высказано предположение, что освобождение этой связки во время распада приводит к характерным позам смерти, при которых позвоночник сильно искривляется. [46]
- конечность
- Конечности вместе с поясами составляют часть добавочного скелета. Передние и задние конечности очень похожи по строению и разделены на три основные части. [1] Верхняя конечность, стилоподий , состоит из плечевой кости (передняя конечность) или бедренной кости (задние конечности), а нижняя конечность, зевгоподий , состоит из локтевой и лучевой кости (передняя конечность) или большеберцовой и малоберцовой костей (задняя конечность). Дистальная часть — аутоподия , включающая руки или ноги. Сама аутоподия подразделяется на три сегмента: запястье (передняя конечность) или предплюсна (задняя конечность) образуют базиподий ; пястная кость (передняя конечность) или плюсневая кость (задняя конечность) метаподия ; и фаланги пальцев — акроподий . [69] : 18 Неправильная терминология была предложена Гюнтером Вагнером и Чи-Хуа Чиу в 2001 году. Здесь аутоподий подразделяется только на две части: мезоподий, включающий узелковые элементы (запястье или предплюсну), и акроподий, который, согласно этому определению, включает все аутоподиальные длинные кости, включая пястные кости (или плюсневые кости) и фаланги. [70]

М
[ редактировать ]- верхняя челюсть
- Верхняя челюсть (множественное число: maxillae ) — это задняя и обычно самая крупная зубная кость верхней челюсти. Спереди она соединяется с предчелюстной костью и часто доходит до границы наружной ноздри. На большей части своей длины верхняя челюсть образует вентральные границы предглазничного отверстия, а также впадину вокруг этого отверстия, предглазничную ямку , которая глубоко утоплена в верхней челюсти. У производных теропод верхняя челюсть может иметь два дополнительных отверстия меньшего размера: промаксиллярное окно и верхнечелюстное окно. На заднем конце верхняя челюсть сочленяется со скуловой и слезной костью. [25] : 37 [1] : 139–140
- нижняя челюсть
- , Нижняя челюсть или нижняя челюсть , является частью черепа ниже черепной коробки . Каждая половина нижней челюсти, называемая нижнечелюстной ветвью (множественное число: нижнечелюстные ветви ; от латинского «ветвь») или полунижнечелюстной ветвью , [71] : 157 соединяется с квадратной костью задней части черепа, образуя челюстной сустав. Спереди обе ветви нижней челюсти прикреплены друг к другу; область прикрепления называется нижнечелюстным симфизом . Нижняя челюсть состоит как из эндохондральных костей, окостеневших от меккелевого хряща , так и из кожных костей. [5] У динозавров только на зубной кости имеются зубы . [25] : 40
- нижнечелюстное окно
- Наружное нижнечелюстное окно представляет собой отверстие в нижней челюсти между зубной , надугольной и угловой костями. Он характерен для архозаврообразных и присутствует у большинства групп динозавров. [25] : 40
- рука
- Рука (множественное число: manus), или рука, включает , пястные кости и фаланги пальцев запястья . [1] : 145 Первая цифра – это поллекс (или большой палец). [72]
- Меккелевская канавка
- Меккелева борозда представляет собой продольный канал внутри нижней челюсти. В нем располагались неоссифицированные остатки меккелева хряща (также: Меккелева хрящ ), от которого окостенели эндохондральные элементы нижней челюсти. Передняя часть меккелевой борозды обнажена на медиальной стороне зубной кости вблизи вентрального края кости, тогда как задняя часть окружена костью с обеих сторон и открывается только дорсально через приводящую ямку . [5] [16] : 44
- мезетмоидный
- Мезетмовидная кость — кость в передней части черепной коробки . Он расположен под лобными костями и впереди клиновидной кости и спереди сочленяется с межглазничной перегородкой . Мезетмоид состоит из горизонтальной дорсальной пластинки, расположенной непосредственно под лобными костями, от которой медиальная перегородка (тонкая костная стенка на средней линии черепа) простирается вентрально, соприкасаясь с базисфеноидом . Как и в случае с сфенетмоидом и орбитосфеноидом , мезетмоид у многих видов не окостенел и поэтому редко сохраняется; однако он обычно встречается у птиц. [73] [20] [25] : 39
- пястная кость
- Пястные кости — длинные кости руки, образующие ладонь. Вместе они называются пястями и расположены между запястьями и фалангами пальцев. Они обозначаются римскими цифрами от I до V, причем I пястная кость является самой внутренней (соединяется с большим пальцем), а пястная V - самой внешней. [1] : 145
- плюсневая кость
- Плюсневые кости – это длинные кости стопы. Вместе называемые плюснами (множественное число: плюсны ), они соединяют предплюсны с пальцами. Как и в случае с пястными костями руки, плюсневые кости обозначаются римскими цифрами от I до V, где I плюсневая кость является самой внутренней (соединяется с большим пальцем ноги), а V плюсневая кость — самой внешней. [1] : 147–148
- Миорхабдои
- Myorhabdoi (также: миорхабдоидные окостенения) — кости, обнаруженные в хвосте пахицефалозавров . Множественные миорхабдои образуют сетчатую структуру, окружающую позвоночный столб (но не соединяющуюся с ним), которую иногда называют каудальной корзиной . Myorhabdoi формируются внутри миосепты (соединительной ткани, разделяющей блоки мышечной ткани, миомеры ) и представляют собой окостеневшие миосептальные сухожилия . Поэтому миорхабдои формируются на периферии мышечной ткани под дермой (а не внутри дермы, как в случае остеодерм ). Они также отличаются от окостеневших сухожилий , которые придавали жесткость хвосту у других птицетазовых динозавров, хотя, возможно, они имели схожие функции. Myorhabdoi известны прежде всего по костистым рыбам ; пахицефалозавры - единственные известные четвероногие , у которых эти структуры окостенеют. Сочлененная хвостовая корзина известна у Homalocephale . [74]

Н
[ редактировать ]- носовой
- Парная носовая кость — самая передняя кость крыши черепа и самая крупная кость верхней поверхности морды. Он расположен между предчелюстной костью спереди и лобной частью сзади. [25] : 38 [1] : 141
- нервная дуга
- Нервная дуга — это дугообразный верхний компонент позвонка , расположенный над позвоночным каналом и центром . У взрослых динозавров центр и нервная дуга обычно сливаются вместе, закрывая нейроцентральный шов , который находится между компонентами у молодых особей. Нервная дуга может содержать ряд костных отростков, таких как нервные отростки , эпипофизы , поперечные отростки (которые соединяются с ребрами и зигапофизы (которые сочленяются с соседними позвонками). У динозавров сложные нервные дуги, часто украшенные системой пластинки , ямки и/или плевроцели , образующие воздушные мешочки, расположенные вдоль позвонков.
- нервный отдел позвоночника
- Нервные отростки (также: остистые отростки ) — это отростки, поднимающиеся вверх от верхней поверхности нервной дуги позвонка . Отростки единичные, они образуют один ряд вдоль средней линии позвоночника, причем их кончики обычно являются высшими точками отдельных позвонков. Нервные отростки обеспечивают поверхности прикрепления мышц и связок, расположенных на вершине позвоночника. [75] : 8 У некоторых видов они могут быть сильно удлинены, образуя парус, или глубоко раздвоены, так что их верхние части образуют двойной ряд. [76] [77]
- нейроцентральный шов
- Нейроцентральный шов — это разделение между центром и нервной дугой каждого позвонка . Молодые динозавры обычно имеют «открытые» (несросшиеся) нейроцентральные швы, которые по мере достижения взрослой жизни становятся «закрытыми» (сросшимися).
- затылочный гребень
- Поперечный затылочный гребень представляет собой гребень в задней части черепа, встречающийся у многих теропод. Ориентированная поперечно (перпендикулярно средней линии черепа), она образует границу между крышей черепа и затылочной областью. В основном он образован задним краем теменных костей с участием чешуйчатых костей . Гребень фиксирует мускулатуру шеи, а его задняя поверхность может содержать ямки для прикрепления выйной связки . Поперечный затылочный гребень не следует путать с сагиттальным затылочным гребнем , срединным гребнем на надзатылочной кости . [78] : 82 [79] : 117 [80] : 167
- выйная связка
- Затылочная связка — это эластичная связка , которая тянется от задней части черепа к задней части шеи над позвоночником. [81] : 354
ТО
[ редактировать ]- затылок
- Затылок – это область на задней стороне черепа. [26] Он состоит из четырех костей, окружающих большое затылочное отверстие , проход спинного мозга, который открывается в полость головного мозга: надзатылочную сверху, парные экзозатылочные кости по бокам и основную затылочную кости снизу. Непосредственно под большим затылочным отверстием находится сферический бугорковидный отросток — затылочный мыщелок , который сочленяется с атлантом — первым позвонком шеи. Затылочный мыщелок в основном образован основной затылочной мышцей с меньшим вкладом экзозатылочной кости на верхних краях. [20] [25] : 38
- опистотический
- Опистоз — парная кость задней части черепной коробки . У взрослых динозавров он всегда слит с парными экзоципитальными костями , образуя структуру, известную как экзоципитально-опистотические. Вместе с экзоципитальными отростками опистотики образуют пару крупных крыловидных отростков, отходящих назад и в стороны от черепной коробки, — парокципитальные отростки . Эти отростки соприкасаются с теменной , чешуйчатой и квадратной костями . [20] [25] : 38
- орбитосфеноид
- Орбитосфеноид (устарело: пресфеноид ) — кость передней части черепной коробки , лежащая в основе переднего мозга. кость Парная , левая и правая половины окружают канал второго черепного нерва (зрительного нерва). У динозавров он окостеневает лишь изредка. [73] [20] [25] : 39
- остеодерма
- Остеодермы – это кости, образующиеся в дерме кожи. У некоторых видов они могут образовывать пластинчатые или шипообразные структуры, особенно у панцирных анкилозавров. Маленькие и неровные остеодермы размером менее 1 см (или 5–15 мм). [82] ) в диаметре известны как косточки . [83] [84]
- окостеневшие сухожилия
- Окостеневшие сухожилия представляют собой костные стержни, удерживающие и придающие жесткость позвоночнику. [1] : 148 на верхних сторонах позвонков считают синапоморфией птицетазовых Их наличие . [85]
- слуховая капсула
- Отическая капсула — это утолщенная боковая область черепной коробки , в которой находится внутреннее ухо. Он состоит из двух оссификаций: проотического спереди и опистотического сзади. Между обеими костями лежит овальное окно — большое отверстие, в которое входит стремечко . [17]

П
[ редактировать ]- подушечка
- Подушечка — это мясистая припухлость на нижней стороне руки или ноги, которую обычно можно увидеть на следах. В зависимости от части руки или стопы, которую они поддерживают, их называют подушечками для ладоней , подушечками для подошв или подушечками для пальцев . [15] : 47 Цифра может иметь отдельные подушечки фаланг (также: цифровые узлы [86] : 112–116 ), которые могут соответствовать отдельным фалангам (костям пальцев рук и ног). [87] У динозавров подушечки фаланг обычно имеют суставное расположение: подушечка охватывает сустав между двумя фалангами. У людей, с другой стороны, наблюдается мезартральное расположение, при котором переход от одной подушечки к другой происходит под суставом. Подушечка, охватывающая сустав между пястной (или плюсневой) и первой фалангой, называется плюснефаланговой подушечкой . [86] : 112–116
- нёбо
- Нёбо — это часть черепа между левой и правой верхними челюстями, образующая нёбо. Он состоит из одной непарной кости — сошника и пяти парных костей ( небной , крыловидной , эктоптеригоидной , эпиптеригоидной и квадратной ). Небо имеет три пары отверстий, а именно хоаны или внутренние ноздри, подглазничные отверстия между небными костями и эктоптеригоидами и подвисочные отверстия позади эктоптеригоидов, которые продолжаются с подвисочными отверстиями . [25] : 39–40
- небный
- небо — парная кожная кость неба Небное . Он контактирует с сошником и крыловидным отростком медиально, а также с верхней и скуловой костью сбоку и образует задний край хоан, а также передний край подглазничных окон . [25] : 39–40
- пальпебральный
- Пальпебральный (также: супраорбитальный , адлакримальный [88] ) представляет собой небольшую кость, прикрепленную к верхнему краю глазницы ( глазнице) или образующую ее части. Формируясь на верхнем веке, он первоначально происходит из остеодермы . По обе стороны черепа может быть одно или несколько глазных яблок. Наличие пальпебралий является синапоморфией птицетазовых, хотя пальпебралы также эволюционировали и в других кладах рептилий, включая крокодилиформных . [89]
- парапофиз
- Парапофизы — отростки в центрах шейных и спинных позвонков, сочленяющиеся головками ребер с . Обычно они расположены близко к переднему краю боковой поверхности центра и хорошо отделены от диапофизов на поперечных отростках нервных дуг, к которым примыкает вторая головка ребра, бугорок . Однако постепенно положение парапофиза смещается на поперечный отросток вдоль позвоночника от шейных к задним спинным позвонкам, где он может образовывать единую структуру с диапофизом, получающим одноглавое ребро. [75] : 8 [11] : 225
- парасфеноид
- Парасфеноид — единственная кожная кость черепной коробки . Он составляет часть нижней части черепной коробки, расположен перед базисфеноидом и простирается вперед под глазами в виде удлиненного отростка. Этот отросток поддерживал мембрану, которая простиралась дорсально до крыши черепа , разделяя правое и левое глазные яблоки. Эта мембрана может быть окостеневшей, тогда ее называют межглазничной перегородкой . [25] : 39
- теменный
- Парная теменная кость — самая задняя кость крыши черепа . Кожная кость, расположена позади лобных костей и покрывает черепную коробку . [25] : 38 [1] : 141 [20]
- теменное окно
- Теменные окна представляют собой пару оконных отверстий, обычно встречающихся в воротниках шеи цератопсов . У хасмозавра и пентацератопса эти отверстия были особенно большими. [90]
- грудной пояс
- Грудной пояс, или плечевой пояс, представляет собой совокупность костей добавочного скелета , соединяющегося с передними конечностями. В строгом смысле грудной пояс включает лопатку , клювовидный отросток и ключицу , а грудину по-разному относят либо к грудному поясу, либо вместе с ребрами к осевому скелету. В отличие от тазового пояса, который жестко закреплен на позвоночнике, грудной пояс не имеет такого костного соединения, а подвешивается за счет мышц. [3]
- таз
- Таз, также известный как тазовый пояс, — это собирательный термин для костей бедра, которые соединяют позвоночный столб с задними конечностями. Таз включает подвздошную , лобковую и седалищную кости . Его внутренняя поверхность имеет прочное соединение с крестцовыми ребрами крестцовых позвонков , которые могут срастаться в крестец. [3]
- пес
- Pes ( множественное число: pedes ), или стопа, состоит из предплюсны , плюсневых костей и педальных фаланг . [1] Первый палец стопы — это большой палец стопы , короткий и не касающийся земли у большинства двуногих динозавров, но увеличенный и перевернутый (направленный назад, а не вперед) у птиц. [2] [91]
- фаланги
- Фаланги (единственное число: фаланга ) — это кости пальцев рук и ног. Последняя фаланга пальца называется когтевой и образует костное ядро рогового копыта или когтя. [1] : 145
- шишковидное отверстие
- Шишковидное отверстие (также: лобно-теменное отверстие ; теменное отверстие ; постфронтальное отверстие ; межлобное отверстие ) — непарное отверстие на средней линии крыши черепа , присутствующее у некоторых динозавров, но часто закрытое у взрослых. Его положение является переменным; он может располагаться между лобными и теменными костями или на стыке лобных и теменных костей. Это отверстие обнажало теменный орган и, таким образом, служило для фоторецепции . [92] [93] [94]
- плевроцель
- Плевроцели — это отверстия на боковых поверхностях позвонков, которые ведут во внутренние камеры в центре и/или нервной дуге позвонка. Плевроцель может представлять собой одну полость или комплекс более мелких взаимосвязанных полостей. [1] : 143–144
- постчерепной отдел
- Посткраниум («за черепом»; множественное число: посткраниа ) — часть скелета, следующая за черепом. Он подразделяется на осевой скелет , включающий все позвонки и ребра, и добавочный скелет , включающий пояса и конечности. [1] Кроме того, отдельные экземпляры, найденные без черепа, часто называют посткраниями.
- посторбитальный
- Заглазничная кость — парная кость на боковой стороне черепа, образующая большую часть заднего края глазницы . У цератопсов он видоизменён и образует выступающие рога над глазами. [25] : 38
- посттеменное отверстие
- Посттеменное отверстие (также: посттеменной родничок , посттеменное окно ) — непарное отверстие, расположенное по средней линии черепа между теменной и надзатылочной костью , встречающееся у некоторых зауроподоморфов . Как и в случае с шишковидным отверстием , оно, возможно, обнажало теменный орган и, таким образом, служило для фоторецепции . [95] [96] [92]
- поствисочное окно
- Поствисочное окно (альтернативное написание: post-temporal fenestra) — парное отверстие, расположенное ниже заднего края теменной части и крыши черепа над околозатылочным отростком черепной коробки . Возможно, он представляет собой выход затылочной ветви глазной артерии . [97] : 30
- предсуставной
- Предсуставная кость — кость заднего отдела нижней челюсти . Обнажаясь на внутренней стороне последней, она располагается впереди суставной и над угловой . Он образует внутренний край приводящей ямки . [5]
- префронтальный
- Префронтальная кость представляет собой меньшую по размеру кость на боковом крае крыши черепа между лобной , слезной и носовой костью . [25] : 38
- предродный
- Преддентарий — непарная кость, находящаяся перед зубными рядами нижней челюсти, образующая нижний клюв у птицетазовых . [1] : 142 Его наличие считается птицетазовой синапоморфией . [98]
- предчелюстная кость
- Парная . предчелюстная кость (множественное число: предчелюстные кости ) — это передняя кость верхней челюсти, образующая кончик морды В нем находится передняя часть верхнего зубного ряда, продолжающая верхнюю часть . идущую сзади [25] : 36
- проатлас
- Проатлан – это небольшая парная кость, расположенная между дугой атланта нервной (первым позвонком) и основанием черепа. Элементы каждой пары имеют треугольную форму, широкий конец сочленяется с нервной дугой атланта, а острый конец — с внешней частью черепа. Учитывая небольшой размер, проатлас легко теряется во время окаменения, сбора или консервации, поэтому его редко находят. [11]
- проотический
- Проотик — парная кость, образующая части боковой поверхности черепной коробки . Вместе с опистотом , расположенным позади него, проотик образует отическую капсулу . [17] Проотик содержит отверстия для тройничного и лицевого нервов и изнутри выкопан пазухами. Его внешняя поверхность обеспечивала шероховатую поверхность крепления мышц, открывающих челюсть. [25] : 39
- крыловидный отросток
- Крыловидный отросток кожная парная . кость неба — Большой и сложный, он сочленяется как с черепной коробкой , так и с другими костями неба. В задней части левая и правая крыловидные кости заключают в себя отверстие — межкрыловидную пустоту . [25] : 39–40
- лобок
- Лобковая кость (множественное число: pubes ) — одна из трех костей, составляющих таз, расположенная вентральнее подвздошной кости и впереди седалищной кости . У ящериц лобковое стержне примитивно направлено вперед ( пролобковое состояние). У птицетазовых первоначальный лобковый стержень направлен назад ( опистолобковое состояние); вместо этого имеется хорошо развитая передняя ветвь или препубис , которая у производных форм может быть больше, чем направленное назад стержне. [99] Левая и правая лобковые кости обычно сращены по всей длине, при этом дистальная часть расширена, образуя лобковый фартук . [63] : 152 У некоторых неотероподов дистальный отдел шва не является непрерывным, а образует фенестру, лобковое отверстие , которое видно с вентральной стороны. [63] : 141 В проксимальной части лобковой кости имеется отверстие — запирательное отверстие , расположенное вблизи вертлужной впадины и видимое при проекции сбоку. Запирательное отверстие обеспечивает прохождение запирательного нерва. У теропод запирательное отверстие не полностью окружено костью, так как вентральный край отсутствует; это состояние называется запирательной выемкой . [63] : 143
- пигостиль
- Пигостиль — это костная структура, состоящая из сросшихся задних хвостовых позвонков. Он встречается у пигостильских птиц , где он служит для закрепления длинных рулевых перьев. [26]

вопрос
[ редактировать ]- квадратный
- Квадратная кость — это большая парная эндохондральная кость в задней части черепа. Столбчатая структура, ее нижний конец сочленяется с нижней челюстью, образуя челюстной сустав, а верхний конец достигает чешуйчатой кости . Квадратная часть нёба . [25] : 39–40
- квадратноскуловой
- Квадратоскуловая кость — это кость в щечной области черепа, которая видна при виде сбоку и образует нижний задний угол черепа . [25] : 37–38
Р
[ редактировать ]- радиус
- Лучевая кость (множественное число: radii ) — меньшая и передняя из двух костей предплечья. [1] : 145
- рамфотека
- Рамфотека (множественное число: рамфотека ) или клюв представляет собой роговое ( ороговевшее ) покрытие на кончиках челюстей, обычно встречающееся у птицетазовых, орнитомимозавров и птиц. [26] [21] Поскольку кератин редко окаменевает, клювы сохраняются только в виде отпечатков. О наличии клювов часто можно судить по шероховатой текстуре поверхности кончиков челюстей, которые сильно васкуляризированы (содержат многочисленные кровеносные сосуды). [2]
- ребра
- Спинные ребра или ребра (единственное число: Costa ; этот латинский термин используется реже, чем английский термин «ребра»). [1] ) прикрепляются к обеим сторонам спинных позвонков и обеспечивают защиту органов, расположенных в туловище. Ребра двуглавые (двуглавые): [100] Дорсальная головка, бугорок (множественное число: туберкула ), сочленяется с поперечными отростками нервной дуги , тогда как вентральная головка, головка (множественное число: capitula ), сочленяется с центром . Ребра также присутствуют на шее ( шейные ребра ) и крестце ( крестцовые ребра ). [1]
- ростральный
- Ростральная кость — новая кость, обнаруженная у цератопсов . Непарный элемент, он покрывает переднюю часть предчелюстных костей на кончике рыла. [25] : 36 В качестве прилагательного термин «ростральный» относится к трибуне . [81] : 357 Ростральные и другие дополнительные окостенения, прикрепленные к черепам цератопсов, в совокупности называются эпиоссификациями . [54]
- трибуна
- Рострум ) (множественное число: ростра — это мордовая часть черепа. [81] : 357

С
[ редактировать ]- сакральные символы
- Крестцовые позвонки, или крестцовые позвонки, позвоночного столба расположены между спинными и хвостовыми позвонками. Они часто сливаются вместе в структуру, известную как крестец (множественное число: крестец ). [1] : 143 Иногда один или несколько спинных позвонков срастаются с крестцом в структуру, называемую синсакрумом . [48] : 368
- плыть
- Паруса (также: паруса нервного позвоночника ) представляют собой структуры, возникающие в результате удлинения позвоночных нервных отростков , обычно в спинных, крестцовых и хвостовых позвонках. Они различаются по размеру и форме: от неглубоких гребней до высоких, заметных структур. [76] [101] В некоторых случаях, например, у Concavenator , нервные отростки образуют «горб». [102] или, как, например, в ( волнообразный ) Ихтиовенаторе, синусоидальный парус . [103] Это состояние наиболее выражено у спинозавридных теропод . [101] но встречается также у кархародонтозаврид , метриакантозаврид , [76] [104] и несколько динозавров -орнитоподов и зауроподов . [105] [106] [107]
- лопатка
- Лопатка (множественное число: scapulae ), или лопатка, представляет собой эндохондральную кость и самый крупный элемент грудного пояса. У взрослых лопатка обычно срастается с клювовидным отростком , образуя лопаточно-коракоидный отросток . [3]
- склеротическое кольцо
- Склеротическое кольцо (также: склеральное кольцо ) — кольцо мелких пластинчатых костей, расположенных на глазу внутри склеры и вокруг зрачка . Отдельные пластинчатые кости называются склеральными косточками . [108] [109] [25] : 84
- вторичное небо
- Вторичное небо представляет собой крышеобразную структуру, отделяющую носовые дыхательные пути от рта. Изначально у рептилий хоаны открываются в ротовую полость в передней части рта, что позволяет пищевым продуктам блокировать дыхательные пути во время кормления. Вторичное небо, которое обычно встречается у млекопитающих, смещает хоаны назад, позволяя одновременно есть и дышать. [81] В нескольких кладах, включая анкилозавров , [110] спинозавриды , [111] и манирапториформные целурозавры, [78] вторичное небо было костным и называлось костным вторичным небом . Напротив, было сделано предположение о мягком вторичном небе для некоторых других таксонов, таких как Plateosaurus и Stegosauria, на основании выступов и гребней небных костей. [112] [113] Костное вторичное небо обычно образовано полкообразными расширениями предчелюстных костей и верхних челюстей , которые встречаются с сошником на средней линии черепа. У анкилозаврид развилась новая дополнительная часть вторичного неба, которая простирается вперед от крыловидной кости и небной кости в задней части неба и получила название каудовентральной небной полки (соответственно, передняя часть, образованная предчелюстными и верхнечелюстными костями, также называется ростродорсальная небная полка ). [110]
- зубцы
- Зубцы — это небольшие выступы, расположенные линиями вдоль режущих кромок ( килей ) зубов. Близкий к нему термин — зубчик (также: зубчики ). Оба термина определялись по-разному, многие авторы использовали их как синонимы. Другие авторы применяли термин «зазубрины» к более мелким структурам зубов плотоядных, а термин «зубцы» - к более грубым выступам, наблюдаемым у многих травоядных зубов. Недавний терминологический обзор определяет зубцы как сложный тип зубцов, который образован как эмалью, так и подлежащим дентином , в то время как термин зубцы также охватывает структуры, образованные исключительно из эмали. Согласно этому определению, выступы, наблюдаемые у хищных динозавров, следует называть зубчиками. [114] Для описания сложной морфологии зубцов плотоядных зубов используется подробная терминология:
- целлы (единственное число: целла ; также: межзубное пространство ): Пространства между отдельными зубцами. [114]
- диафиз (множественное число: диафизы ): точка, где эмалевые шляпки двух соседних зубцов расходятся или располагаются ближе всего друг к другу. [115] [116] [114]
- ампула (множественное число: ампулы ): камера в форме колбы под диафизом. [114]
- operculum : Эмалевой слой зубца. [114]
- радикс : ядро зубца под жаберной крышкой. [114]
- межзубная борозда (множественное число: борозды, также: кровеносная канавка ): Борозда, которая проходит из промежутков между соседними зубцами на небольшое расстояние к поверхности зуба с обеих сторон перпендикулярно килю . [114]
- хвост (множественное число: caudae ): выпуклость, возникающая от основания зубца и идущая перпендикулярно килю на поверхность зуба между двумя межзубными бороздками. [114]
- череп
- Череп включает в себя кости и зубы головы. Он состоит из верхней части, черепа (множественное число: crania ), а также нижней части, нижней челюсти . Череп состоит из черепной коробки , крыши черепа , верхней челюсти и щечной области; и нёбо . Черепная коробка почти полностью эндохондрального происхождения, а остальная часть черепа состоит преимущественно из кожных костей. Нижние челюсти состоят как из дермальных, так и из эндохондральных костей. В черепе имеется ряд отверстий, которые являются важными ориентирами в анатомических описаниях. К ним относятся орбита (множественное число: орбита ) или глазница, в которой располагался глаз, а также внешняя ноздря . Кроме того, в черепе имелись дополнительные отверстия, или фенестры (единственное число: fenestra ), типичные для диапсидных рептилий : предглазничное отверстие между наружным ноздрем и глазницей; подвисочное окно позади глазницы и надвисочное окно в задней части крыши черепа. [1] : 138–140 [25] : 32–36 [17]
- крыша черепа
- Крыша черепа – это верхняя поверхность черепа. У динозавров она состоит из четырех парных костей: носовые , лобные и теменные — крупные элементы, а префронтальные — мелкие и располагаются между носовой, лобной и слезной . В задней части черепа находятся надвисочные окна . [25] : 38
- сфенетмоид
- Сфенетмоид — трубчатая кость в передней части черепной коробки . Расположенный под лобными костями, позади мезетмовидной кости и перед орбитосфеноидом , он содержал обонятельные луковицы , в которых располагалось обоняние. Как и в случае мезетмоида и орбитосфеноида, сфенетмоид у многих видов не окостенел и поэтому редко сохраняется. [73] [20] [25] : 39
- селезеночный
- Сплениальная кость — кожная кость нижней челюсти , расположенная позади и медиально (внутри) зубной кости . Сплениальная кость покрывает большую часть внутренней стороны зубной кости и часто является самой обширной костью нижней челюсти при взгляде медиально. [25] : 40 [5]
- чешуйчатый
- Чешуйчатая кость — парная кость в задних углах черепа. [25] : 38
- субнариальное отверстие
- Субнариальное отверстие представляет собой небольшое отверстие между предчелюстной и верхней челюстью у ящеротазовых и некоторых базальных птицетазовых животных. [25] : 23
- субнариальный разрыв
- Субнариальный зазор (также называемый «перегиб») — это зазор между предчелюстной и верхней челюстью в морде целофизоидных , дилофозавридных и спинозавридных теропод. В результате субнариального разрыва образуется диастема – разрыв в зубном ряду. Пространство между субнариальной щелью рыла и загнутым вверх кончиком нижней челюсти называется субростральной вырезкой . [117] [118] [119] [120] У дилофозавра щель содержала глубокую выемку позади зубного ряда предчелюстной кости, называемую субнариальной ямкой . [117]
- надзатылочный
- Надзатылочная кость — непарная кость затылка в задней части черепа. Он образует верхний край большого затылочного отверстия и дорсально соединяется с теменными костями. На нем часто виден срединный гребень для прикрепления мышц шеи. [20] [25] : 38–39
- надвисочное окно
- Надвисочное окно (также: верхнее височное окно) [121] ) — отверстие в задней части черепа. [1] : 140 Обычно он граничит с заглазничной , чешуйчатой и теменной областью . [25] : 38
- надугольный
- Надугольная кость — это дермальная кость нижней челюсти, которая видна при виде сбоку и расположена позади зубной кости и над угловой . [25] : 40



Т
[ редактировать ]- предплюсны
- Предплюсны представляют собой набор костей задней конечности между плюсневыми костями внизу и большеберцовой и малоберцовой костями вверху. [1] : 148 Их все вместе называют предплюсной . Предплюсна состоит из верхнего ряда, образованного таранной и пяточной костями , и нижнего ряда мелких костей, расположенных на вершине плюсневых костей. Верхний и нижний ряд могут двигаться друг против друга, образуя голеностопный сустав . [25] : 53
- зубы
- У динозавров зубы имеются на верхней и предчелюстной кости верхней челюсти, а также на зубной кости нижней челюсти. [25] : 40 Лишь самые базальные динозавры Eoraptor и Eodromaeus несут рудиментарные зубы на крыловидной ( кости неба небные зубы ). [122] Челюстная кость без зубов называется беззубой . [1] : 140 Зубы можно разделить на две части: коронку , то есть верхнюю, открытую часть, и корень , расположенный внутри десны. [48] Зубы в основном состоят из двух типов тканей: эмали и дентина. Эмаль образует твердый и блестящий внешний слой, а более мягкий и похожий на кость дентин заполняет внутреннюю часть. третий тип — цемент . Иногда поверх слоя эмали можно обнаружить [41]
- большеберцовая кость
- Большеберцовая кость (множественное число: tibiae ) — большая из двух костей голени. Он расположен медиальнее малоберцовой кости . Большеберцовая кость имеет выступающий гребень, идущий от ее передней поверхности на верхнем конце, - кнемиальный гребень (также: большеберцовый гребень ). Кнемиальный гребень фиксировал сухожилия трехглавой мышцы бедра, которые отвечали за разгибание колена (вытягивание ноги), и увеличивал силу воздействия этих мышц. [4] : 178 [1] : 147
- тагомайзер
- Тагомайзер . — это от четырех до десяти шипов на хвосте, встречающихся у стегозавров Термин был придуман в 1982 году карикатуристом Гэри Ларсоном . [26]
- поперечный отросток
- Поперечные отростки представляют собой крыловидные выступы, отходящие в стороны от нервных дуг шейных, спинных, крестцовых и самых передних хвостовых позвонков. Их типично горизонтальная ориентация приводит к характерной Т-образной форме позвонков при виде спереди или сзади. поперечный отросток сочленяется с бугорком ребра На его вершине ; расширенная область сочленения ребра называется диапофизом ( множественное число: диапофизы ). [25] : 42 Некоторые авторы, однако, называют весь поперечный отросток диапофизом, используя оба термина как синонимы. [11] : 224–225 [75] : 8

В
[ редактировать ]- локтевая кость
- Локтевая кость (множественное число: ulnae ) — более крупная и расположенная сзади из двух костей предплечья. [1] : 145 На верхнем конце локтевой кости часто виден большой отросток, выходящий за пределы локтевого сустава, локтевой отросток , к которому прикреплялись трехглавые мышцы (мышцы-разгибатели предплечья). Иногда значительное удлинение локтевого отростка обеспечивало этим мышцам повышенную рычаговую нагрузку. [25] : 50 [4] : 173–174
V
[ редактировать ]- позвонки
- Позвонки (единственное число: позвонок ) — это костные компоненты позвоночного столба (т. е. позвоночника или позвоночника). У динозавров позвонки подразделяются на шейные (шейные позвонки), спинные (туловищные позвонки), крестцовые (тазовые позвонки) и хвостовые (хвостовые позвонки). Каждый позвонок имеет нижний компонент в форме катушки, центр , и дугообразный верхний компонент, нервную дугу . Между этими компонентами находится трубчатый туннель — позвоночный канал . Спинной мозг нервной системы проходит через позвоночный канал каждого позвонка позвоночника. Шейные, спинные и крестцовые позвонки соединяются с ребрами через поперечные отростки по бокам нервной дуги. Хвостовые позвонки соединяются с шевронами на нижней стороне их центров.
- рвота
- Сошник — непарная узкая кость передней части неба, разделяющая левую и правую верхнечелюстные кости и хоаны . [25] : 39–40
В
[ редактировать ]Х
[ редактировать ]И
[ редактировать ]С
[ редактировать ]- ряд зубов
- Zahnreihe . ( по-немецки «зубной ряд», множественное число Zahnreihen ) — это ряд позиций зубов, образующих замещающую единицу Зубной ряд обычно состоит из нескольких Цанрейхенов, содержащих по два и более зуба каждый. Замена зуба всегда начинается с самого заднего положения зуба Цанрайхе и распространяется вперед. Таким образом, можно избежать одновременной замены двух соседних позиций зубов, что привело бы к образованию промежутков в зубном ряду. Количество зубьев от одного Цанрайхе к другому и есть Z-расстояние . Когда расстояние по оси Z равно ровно 2, зубы заменяются поочередно; если расстояние по оси Z больше 2, зубы будут заменяться последовательно. [123]
- зифодонт
- Зифодонт относится к зубам, похожим на лезвия, с загнутыми и уплощенными коронками, которые обычно имеют зазубрины. Зубы зифодонтов, адаптированные к плотоядной диете, примитивно присутствуют у динозавров и встречаются у большинства нептичьих теропод. [41]
- зигапофизы
- Зигапофизы (единственное число: зигапофиз ), или суставные отростки , представляют собой проекции дуги позвонка, соединяющие соседние позвонки. Каждый позвонок имеет две пары зигапофизов, которые сочленяются с зигапофизами следующего и предыдущего позвонка: Спереди расположены презигапофизы ( также: краниальные зигапофизы ), выступающие вперед и имеющие суставную поверхность, обращенную вверх. В задней части позвонка расположены постзигапофизы (также: каудальные зигапофизы ), которые отходят назад суставной поверхностью вниз. [1] : 143
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа аб и объявление но из в ах есть также и аль являюсь Хольц, ТР; Бретт-Сурман, МК (2012). «Остеология динозавров». В Бретт-Сурмане, МК; Хольц, ТР; Фарлоу, Дж. О. (ред.). Полный динозавр (2-е изд.). Издательство Университета Индианы. стр. 135–149. ISBN 978-0-253-00849-7 .
- ^ Jump up to: а б с Карри, Пи Джей; Падиан, К., ред. (1997). «Глоссарий». Энциклопедия динозавров . стр. 813–827 . ISBN 978-0-12-226810-6 .
- ^ Jump up to: а б с д и ж Падиан, К. (1997). «Пекторальный пояс». В Карри, Пи Джей; Падиан, К. (ред.). Энциклопедия динозавров . стр. 530–536 . ISBN 978-0-12-226810-6 .
- ^ Jump up to: а б с д и ж г час Дилкс, Д.В.; Хатчинсон-младший; Холлидей, СМ; Витмер, LM (2012). «Реконструкция мускулатуры динозавров». В Бретт-Сурмане, МК; Хольц, ТР; Фарлоу, Дж. О. (ред.). Полный динозавр (2-е изд.). Издательство Университета Индианы. стр. 150–190. ISBN 978-0-253-00849-7 .
- ^ Jump up to: а б с д и ж г Ромер, А.С. (1976). «Человек». Остеология рептилий . Издательство Чикагского университета. стр. 198–217. ISBN 978-0-226-72491-1 .
- ^ Ведель, MJ (2009). «Доказательства существования птичьих воздушных мешков у ящеротазовых динозавров» . Журнал экспериментальной зоологии. Часть A: Экологическая генетика и физиология . 311 (8): 611–628. Бибкод : 2009JEZA..311..611W . дои : 10.1002/jez.513 . ПМИД 19204909 .
- ^ Карри, Пи Джей; Падиан, К., ред. (1997). «Текодонтия». Энциклопедия динозавров . стр. 728 . ISBN 978-0-12-226810-6 .
- ^ Фиорилло, Арканзас (1997). «Тафономия». В Карри, Пи Джей; Падиан, К. (ред.). Энциклопедия динозавров . стр. 713–716 . ISBN 978-0-12-226810-6 .
- ^ Jump up to: а б Падиан, К. (1997). «Двуногость». В Карри, Пи Джей; Падиан, К. (ред.). Энциклопедия динозавров . стр. 68–70 . ISBN 978-0-12-226810-6 .
- ^ Цуихидзи, Таканобу; Маковицкий, Питер Дж. (1 сентября 2007 г.). «Гомология неоцератопсических элементов шейного перекладины» . Журнал палеонтологии . 81 (5): 1132–1138. Бибкод : 2007JPal...81.1132T . дои : 10.1666/pleo05-164.1 . ISSN 0022-3360 . S2CID 131118800 .
- ^ Jump up to: а б с д и ж г Ромер, А.С. (1976). «Осевой скелет». Остеология рептилий . Издательство Чикагского университета. стр. 218–297. ISBN 978-0-226-72491-1 .
- ^ Гальтон, Питер М.; Кермак, Дайан (2010). «Анатомия Pantydraco caducus, очень базального динозавра-зауроподоморфа из ретийского периода (верхний триас) Южного Уэльса, Великобритания». Ревю палеобиологии . 29 : 341–404.
- ^ Хатчинсон-младший; Падиан, К. (1997). «Арктометатарзалия». В Карри, Пи Джей; Падиан, К. (ред.). Энциклопедия динозавров . стр. 24–26 . ISBN 978-0-12-226810-6 .
- ^ Хендерсон, Д. (2012). «Инженерия динозавра». У Майкла К. Бретта-Сурмана; Томас Р. Хольц; Джеймс Орвилл Фарлоу (ред.). Полный динозавр . Издательство Университета Индианы. стр. 637–665.
- ^ Jump up to: а б с д Леонарди, Джузеппе (1987). Глоссарий и руководство по палеоихнологии следов четвероногих . Бразилиа: Публикация Национального департамента добычи полезных ископаемых Бразилии. стр. 43–51.
- ^ Jump up to: а б Прието-Маркес, Альберт; Норелл, Марк А. (2011). «Переописание почти полного черепа платеозавра (Dinosauria: Sauropodomorpha) из позднего триаса Троссингена (Германия)» . Новитаты Американского музея (3727): 1–58. дои : 10.1206/3727.2 . S2CID 55272856 .
- ^ Jump up to: а б с д и ж Ромер, А.С. (1976). «Череп – Общая структура». Остеология рептилий . Издательство Чикагского университета. стр. 55–83. ISBN 978-0-226-72491-1 .
- ^ Ромер, А.С. (1976). «Череп в различных группах рептилий». Остеология рептилий . Издательство Чикагского университета. стр. 85–197. ISBN 978-0-226-72491-1 .
- ^ Роджерс, Кристина Карри; Форстер, Кэтрин А. (2004). «Череп Rapetosaurus krausei (sauropoda: Titanosauria) из позднего мела Мадагаскара». Журнал палеонтологии позвоночных . 24 (1): 121–144. Бибкод : 2004JVPal..24..121R . дои : 10.1671/A1109-10 . S2CID 83855113 .
- ^ Jump up to: а б с д и ж г час я дж к л Карри, Пи Джей (1997). «Анатомия черепной коробки». В Карри, Пи Джей; Падиан, К. (ред.). Энциклопедия динозавров . стр. 81–85 . ISBN 978-0-12-226810-6 .
- ^ Jump up to: а б с Падиан, К. (1997). «Скелетные структуры». В Карри, Пи Джей; Падиан, К. (ред.). Энциклопедия динозавров . стр. 668 . ISBN 978-0-12-226810-6 .
- ^ Бочеренс, Х. (1997). «Химический состав окаменелостей динозавров». В Карри, Пи Джей; Падиан, К. (ред.). Энциклопедия динозавров . стр. 111–117 . ISBN 978-0-12-226810-6 .
- ^ Удары, Уильям Т. (2001). «Кожный доспех полукантинских динозавров». В Кеннете Карпентере (ред.). Бронированные динозавры . Издательство Университета Индианы. стр. 363–385. ISBN 978-0-253-33964-5 .
- ^ Арбур, Виктория М.; Карри, Филип Дж. (2013). « Euoplocephalus tutus и разнообразие динозавров-анкилозаврид в позднем мелу Альберты, Канада, и Монтаны, США» . ПЛОС ОДИН . 8 (5): –62421. Бибкод : 2013PLoSO...862421A . дои : 10.1371/journal.pone.0062421 . ПМЦ 3648582 . ПМИД 23690940 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа аб и объявление но из в ах есть также и аль являюсь а к ап ак с как в В из хорошо топор является тот нет бб до нашей эры др. быть парень бг чб с минет БК Брусатте, Стивен Л. (2012). Палеобиология динозавров . Джон Уайли и сыновья.
- ^ Jump up to: а б с д и ж г час я дж к л м н Бретт-Сурман, депутат Кнессета; Хольц, ТР; Фарлоу, Дж.О., ред. (2012). «Глоссарий». Полный динозавр (2-е изд.). Издательство Университета Индианы. стр. 1075–1081. ISBN 978-0-253-00849-7 .
- ^ Гальтон, премьер-министр (2014). «Заметки о посткраниальной анатомии гетеродонтозавридного динозавра Heterodontosaurus tucki , базального птицетазового животного из нижней юры Южной Африки» (PDF) . Revue de Paléobiologie, Женева . 1. 33 : 97–141. ISSN 1661-5468 .
- ^ Лэнг, Эмили; Гуссар, Флоран (2007). «Переописание запястья и кисти ? Bothriospondylus madagascariensis : новые данные о морфологии запястья у Sauropoda». Геодиверситас . 29 (4): 549–560.
- ^ Лица IV, В. Скотт; Карри, Филип Дж. (2011). «Хвост тираннозавра: переоценка размера и двигательной роли M. caudofemoralis у нептичьих теропод» . Анатомические записи: достижения в интегративной анатомии и эволюционной биологии . 294 (1): 119–131. дои : 10.1002/ar.21290 . ПМИД 21157923 . S2CID 26158462 .
- ^ Jump up to: а б с Мэдсен, Дж. Х.; Уэллс, СП (2000). Цератозавр (Динозавр, Теропода): пересмотренная остеология . Геологическая служба Юты . стр. 1–80. ISBN 978-1-55791-380-7 .
- ^ Эверс С.В., Раухут О.В.М., Милнер А.С., Макфитерс Б., Аллен Р. 2015. Переоценка морфологии и систематического положения тероподного динозавра Sigilmassasaurus из «среднего» мелового периода Марокко. PeerJ 3:e1323 https://doi.org/10.7717/peerj.1323
- ^ Фронимос, Дж.А.; Уилсон, Дж.А.; Баумиллер, ТК (2016). «Полярность вогнуто-выпуклых межпозвонковых суставов шей и хвостов динозавров-зауроподов». Палеобиология . 42 (4): 624–642. Бибкод : 2016Pbio...42..624F . дои : 10.1017/pab.2016.16 . S2CID 89348860 .
- ^ Кардонг, Кеннет В. «Скелетная система: осевой скелет». Позвоночные животные - сравнительная анатомия, функции, эволюция (6-е изд.). Мак Грау Хилл. стр. 294–324. ISBN 978-9-33-922211-6 .
- ^ Карпентер, К.; Тидвелл, В. (2005). «Переоценка зауроподов раннего мела Astrodon johnsoni Leidy 1865 (Titanosauriformes)». В Карпентере, К.; Тидвелл, К. (ред.). Громовые ящерицы: динозавры-зауроподоморфы . Издательство Университета Индианы. стр. 78–114. ISBN 978-0-253-34542-4 .
- ^ Jump up to: а б Хольц, ТР; Бретт-Сурман, МК (2012). «Палеоневрология динозавров». В Бретт-Сурмане, МК; Хольц, ТР; Фарлоу, Дж. О. (ред.). Полный динозавр (2-е изд.). Издательство Университета Индианы. стр. 191–208. ISBN 978-0-253-00849-7 .
- ^ Кундрат, Мартин (2007). «Птичьи атрибуты виртуальной модели мозга теропода-овираптора Conchoraptor gracilis ». Naturwissenschaften . 94 (6): 499–504. Бибкод : 2007NW.....94..499K . дои : 10.1007/s00114-007-0219-1 . ПМИД 17277940 . S2CID 25723660 .
- ^ Удары, Уильям Т. (2014). «Уведомление о нодозавре (Dinosauria, Ankylosauria) сохранилось из середины мела в Кембридже, Англия, с комментариями к шейному полукольцу» (PDF) . Труды Ассоциации геологов . 125 (1): 56–62. Бибкод : 2014ПрГА..125...56Б . дои : 10.1016/j.pgeola.2013.07.001 .
- ^ Кристиан, А.; Дземский, Г. (2007). «Реконструкция положения шейного скелета Brachiosaurus brancai Janensch, 1914 путем анализа межпозвонкового напряжения вдоль шеи и сравнения с результатами разных подходов» . Ископаемый рекорд . 10 (1): 38–49. дои : 10.1002/mmng.200600017 .
- ^ Кляйн, Н.; Кристиан, А.; Сандер, премьер-министр (2012). «Гистология показывает, что удлиненные шейные ребра у динозавров-зауроподов представляют собой окостеневшие сухожилия» . Письма по биологии . 8 (6): 1032–1035. дои : 10.1098/rsbl.2012.0778 . ПМЦ 3497149 . ПМИД 23034173 .
- ^ Вейшампель, Дэвид Б. (1981). «Носовая полость ламбеозавриновых гадрозаврид (Reptilia: Ornithischia): сравнительная анатомия и гомологии». Журнал палеонтологии : 1046–1057.
- ^ Jump up to: а б с д Сандер, премьер-министр (1997). «Зубы и челюсти». В Карри, Пи Джей; Падиан, К. (ред.). Энциклопедия динозавров . стр. 717–725 . ISBN 978-0-12-226810-6 .
- ^ Jump up to: а б с Чопп, Эмануэль; Матеус, Октавио (2013). «Ключицы, межключицы, гастралии и грудинные ребра у динозавров-зауроподов: новые сообщения о Diplodocidae и их морфологические, функциональные и эволюционные последствия» . Журнал анатомии . 222 (3): 321–340. дои : 10.1111/joa.12012 . ПМЦ 3582252 . ПМИД 23190365 .
- ^ Ву, Сяо-Чун; Рассел, Энтони П. (1997). «Функциональная морфология». У Кевина Падиана; Филип Дж. Карри (ред.). Энциклопедия динозавров . Сан-Диего: Академическая пресса. стр. 258–268 . ISBN 978-0-12-226810-6 .
- ^ Jump up to: а б с д и Холлидей, Кейси М.; Уитмер, Лоуренс М. (2008). «Краниальный кинезис у динозавров: внутричерепные суставы, мышцы-протракторы и их значение для эволюции и функции черепа у диапсид». Журнал палеонтологии позвоночных . 28 (4): 1073–1088. Бибкод : 2008JVPal..28.1073H . дои : 10.1671/0272-4634-28.4.1073 . S2CID 15142387 .
- ^ Jump up to: а б Фальшивый, Синтия Маршалл; Падиан, Кевин (2007). «Опистотоническая поза скелетов позвоночных: посмертное сокращение или предсмертная агония?» . Палеобиология . 33 (2): 201–226. Бибкод : 2007Pbio...33..201F . дои : 10.1666/06015.1 . ISSN 0094-8373 . S2CID 86181173 .
- ^ Jump up to: а б с д Райсдорф, АГ; Вуттке, М. (2012). «Переоценка гипотезы опистотонической позы Муди у ископаемых позвоночных. Часть I: Рептилии - Тафономия двуногих динозавров Compsognathus longipes и Juravenator starki с архипелага Зольнхофен (юрский период, Германия)». Палеобиоразнообразие и палеосреда . 92 (1): 119–168. Бибкод : 2012ПдПе...92..119Р . дои : 10.1007/s12549-011-0068-y . S2CID 129785393 .
- ^ ЛеБлан, Аарон Р.Х.; Рейс, Роберт Р.; Эванс, Дэвид С.; Байёль, Алида М. (2016). «Онтогенез раскрывает функцию и эволюцию зубной батареи гадрозаврид динозавров» . Эволюционная биология BMC . 16 (1): 152. Бибкод : 2016BMCEE..16..152L . дои : 10.1186/s12862-016-0721-1 . ISSN 1471-2148 . ПМЦ 4964017 . ПМИД 27465802 .
- ^ Jump up to: а б с Мартин, Энтони Дж. (2006). Введение в изучение динозавров (2-е изд.). Издательство Блэквелл. ISBN 978-1-4051-3413-2 .
- ^ Эриксон, Грегори М. (1997). «Схемы замены зубов». У Кевина Падиана; Филип Дж. Карри (ред.). Энциклопедия динозавров . Сан-Диего: Академическая пресса. стр. 739–740 . ISBN 978-0-12-226810-6 .
- ^ Кольбер, Эдвин Харрис (1964). «Взаимоотношения ящеротазовых динозавров». Новитаты Американского музея (2181): 1–24.
- ^ Гальтон, Питер М. (1 декабря 1977 г.). «О Staurikosaums Pricei , раннем ящеротазовом динозавре из триаса Бразилии, с примечаниями о Herrerasauridae и Poposauridae». Палеонтологическая газета . 51 (3): 234–245. Бибкод : 1977PalZ...51..234G . дои : 10.1007/BF02986571 . ISSN 1867-6812 . S2CID 129438498 .
- ^ Jump up to: а б с Додсон, П.; Форстер, Калифорния; Сэмпсон, С.Д. (2004), «Ceratopsidae», в Вейшампеле, Д.Б.; Додсон, П.; Осмольска, Х. (ред.), Динозаврия (2-е изд.), Беркли: University of California Press, стр. 494–513 , ISBN 978-0520254084
- ^ Jump up to: а б Маковицкий, П. (2012). «Маргиноцефалия». В Бретт-Сурмане, МК; Хольц, ТР; Фарлоу, Дж. О. (ред.). Полный динозавр (2-е изд.). Издательство Университета Индианы. стр. 526–549. ISBN 978-0-253-00849-7 .
- ^ Jump up to: а б с д Хорнер, Джон Р.; Гудвин, Марк Б. (2008). «Онтогенез черепных эпиоссификаций у трицератопса ». Журнал палеонтологии позвоночных . 28 (1): 134–144. doi : 10.1671/0272-4634(2008)28[134:OOCEIT]2.0.CO;2 . S2CID 86291412 .
- ^ Jump up to: а б с Уитмер, Лоуренс М. (2001). «Положение ноздрей у динозавров и других позвоночных и его значение для функции носа». Наука . 293 (5531): 850–853. дои : 10.1126/science.1062681 . ПМИД 11486085 . S2CID 7328047 .
- ^ Уитмер, Лоуренс М. (1997). «Эволюция анторбитальной полости архозавров: исследование реконструкции мягких тканей в летописи окаменелостей с анализом функции пневматическости». Журнал палеонтологии позвоночных . 17 (С1): 1–76. Бибкод : 1997JVPal..17S...1W . дои : 10.1080/02724634.1997.10011027 .
- ^ Классенс, Леон ПАМ (2004). «Гастралия динозавра; происхождение, морфология и функции» (PDF) . Журнал палеонтологии позвоночных . 24 (1): 89–106. Бибкод : 2004JVPal..24...89C . дои : 10.1671/A1116-8 . S2CID 53318713 .
- ^ Иероним, Тобин; Уитмер, Лоуренс (2004). «Морщинистость черепа и рога динозавров: носорог и жираф как модельные системы для реконструкции кожи ископаемых таксонов». Журнал палеонтологии позвоночных . Том. 24. С. 70–70А.
- ^ Апестегия, Себастьян (2005). «Эволюция комплекса гипосфен-гипантрум у зауропод». В Вирджинии Тидвелл; Кеннет Карпентер (ред.). Громовые ящерицы: динозавры-зауроподоморфы . Издательство Университета Индианы. ISBN 978-0-253-34542-4 .
- ^ Лангер, Макс К.; Бентон, Майкл Дж. (2006). «Ранние динозавры: филогенетическое исследование». Журнал систематической палеонтологии . 4 (4): 309–358. Бибкод : 2006JSPal...4..309L . дои : 10.1017/S1477201906001970 . S2CID 55723635 .
- ^ Jump up to: а б Готье, Жак (1986). «Монофилия ящеров и происхождение птиц». Мемуары Калифорнийской академии наук . 8 : 1–55.
- ^ Jump up to: а б Новас, Фернандо Э. (1996). «Монофилия динозавров». Журнал палеонтологии позвоночных . 16 (4): 723–741. Бибкод : 1996JVPal..16..723N . дои : 10.1080/02724634.1996.10011361 .
- ^ Jump up to: а б с д и Хатчинсон, Джон Р. (2001). «Эволюция остеологии таза и мягких тканей на пути к современным птицам (Neornithes)» . Зоологический журнал Линнеевского общества . 131 (2): 123–168. дои : 10.1111/j.1096-3642.2001.tb01313.x .
- ^ Несбитт, SJ (29 апреля 2011 г.). «Ранняя эволюция архозавров: взаимоотношения и происхождение основных клад» . Бюллетень Американского музея естественной истории . 352 : 1–292. дои : 10.1206/352.1 . hdl : 2246/6112 . ISSN 0003-0090 . S2CID 83493714 .
- ^ Бадни, Лиза А.; Колдуэлл, Майкл В.; Альбинос, Адриана (2006). «Гистология зубной лунки меловой змеи Dinilysia с обзором тканей прикрепления зубов амниот». Журнал палеонтологии позвоночных . 26 (1): 138–145. doi : 10.1671/0272-4634(2006)26[138:TSHITC]2.0.CO;2 . S2CID 86130973 .
- ^ Рид, REH (1997). «Гистология костей и зубов». У Кевина Падиана; Филип Дж. Карри (ред.). Энциклопедия динозавров . Сан-Диего: Академическая пресса. стр. 329–339 . ISBN 978-0-12-226810-6 .
- ^ Уилсон, Дж. А. (1999). «Номенклатура позвоночных пластинок зауроподов и других ящеротазовых динозавров» . Журнал палеонтологии позвоночных . 19 (4): 639–653. Бибкод : 1999JVPal..19..639W . дои : 10.1080/02724634.1999.10011178 .
- ^ Франкфурт, Николас Г.; Кьяппе, Луис М. (1999). «Возможный овирапторозавр из позднего мела северо-западной Аргентины» . Журнал палеонтологии позвоночных . 19 (1): 101–105. Бибкод : 1999JVPal..19..101F . дои : 10.1080/02724634.1999.10011126 . ISSN 0272-4634 .
- ^ Шюнке, М.; Шульте, Э.; Шумахер, У. (2006). Росс, LM; Ламперти, ЭД (ред.). Атлас анатомии Тиме: общая анатомия и скелетно-мышечная система . Тиме. ISBN 978-3-13-142071-8 .
- ^ Вагнер, Гюнтер П.; Чиу, Чи-Хуа (2001). «Конечность четвероногих: гипотеза о ее происхождении». Журнал экспериментальной зоологии. Часть A: Экологическая генетика и физиология . 291 (3): 226–240. Бибкод : 2001JEZ...291..226W . дои : 10.1002/jez.1100 . ПМИД 11598912 .
- ^ Бейсо, Эйприл М. (2013). Идентификация и интерпретация костей животных: Руководство . Издательство Техасского университета A&M. ISBN 978-1-62349-082-9 .
- ^ Падиан, Кевин (1997). «Саурисхия». У Кевина Падиана; Филип Дж. Карри (ред.). Энциклопедия динозавров . Сан-Диего: Академическая пресса. стр. 647–653 . ISBN 978-0-12-226810-6 .
- ^ Jump up to: а б с Али, Фархин; Зеленицкий, Дарла К.; Терриен, Франсуа; Вейшампель, Дэвид Б. (2008). «Гомология «решетчатого комплекса» тираннозаврид и его значение для реконструкции обонятельного аппарата нептичьих теропод». Журнал палеонтологии позвоночных . 28 (1): 123–133. doi : 10.1671/0272-4634(2008)28[123:HOTECO]2.0.CO;2 . S2CID 85751740 .
- ^ Браун, Калеб Маршалл; Рассел, Энтони П. (2012). «Гомология и архитектура хвостовой корзины Pachycephalosauria (Dinosauria: Ornithischia): первое появление миорхабдоев у четвероногих» . ПЛОС ОДИН . 7 (1): –30212. Бибкод : 2012PLoSO...730212B . дои : 10.1371/journal.pone.0030212 . ПМК 3260247 . ПМИД 22272307 .
- ^ Jump up to: а б с Новас, Фернандо Э. (2009). Эпоха динозавров в Южной Америке . Блумингтон: Издательство Университета Индианы. ISBN 978-0-253-35289-7 .
- ^ Jump up to: а б с Бейли, Джек Боуман (1 ноября 1997 г.). «Удлинение нервного отдела позвоночника у динозавров: парусники или буйволы?» . Журнал палеонтологии . 71 (6): 1124–1146. Бибкод : 1997JPal...71.1124B . дои : 10.1017/S0022336000036076 . ISSN 0022-3360 . JSTOR 1306608 . S2CID 130861276 .
- ^ Апчерч, П.; Барретт, премьер-министр; Додсон, П. (2004). «Зауропода». В Вейшампеле, Д.Б.; Додсон, П.; Осмольская, Х. (ред.). Динозаврия, второе издание . Университет Калифорнии Пресс, Беркли. стр. 259–322 . ISBN 978-0-520-24209-8 .
- ^ Jump up to: а б Хольц-младший, ТР; Мольнар, Р.Э.; Карри, Пи Джей (2004). «Базальная столбняк». У Дэвида Б. Вейшампеля; Питер Додсон; Х. Осмольска (ред.). Динозаврия (2-е изд.). Беркли: Издательство Калифорнийского университета. стр. 71 –110. ISBN 978-0-520-24209-8 .
- ^ Хольц, Томас Р. (2004). «Тираннозавроидея». У Дэвида Б. Вейшампеля; Питер Додсон; Х. Осмольска (ред.). Динозаврия (2-е изд.). Беркли: Издательство Калифорнийского университета. стр. 111–136 . ISBN 978-0-520-24209-8 .
- ^ Осмольска, Гальшка; Карри, Филип Дж.; Барсболд, Ринчен (2004). «Овирапторозаврия». У Дэвида Б. Вейшампеля; Питер Додсон; Х. Осмольска (ред.). Динозаврия (2-е изд.). Беркли: Издательство Калифорнийского университета. стр. 165–183 . ISBN 978-0-520-24209-8 .
- ^ Jump up to: а б с д Фастовский, Дэвид Э. и Вейшампель, Дэвид Б. (2009). Динозавры: краткая естественная история . Кембридж, Великобритания: Издательство Кембриджского университета. п. 358. ИСБН 978-0-511-47789-8 .
- ^ Форд, Трейси Л. (2000). «Обзор остеодерм анкилозавра из Нью-Мексико и предварительный обзор панциря анкилозавра». Бюллетень Музея естественной истории и науки Нью-Мексико . 17 : 157–176.
- ^ Арбур, Виктория М.; Бернс, Майкл Э.; Белл, Фил Р.; Карри, Филип Дж. (2014). «Эпидермальные и дермальные покровные структуры анкилозавровых динозавров». Журнал морфологии . 275 (1): 39–50. дои : 10.1002/Jmor.20194 . ПМИД 24105904 . S2CID 35121589 .
- ^ Д'Эмик, Майкл Д.; Уилсон, Джеффри А.; Чаттерджи, Санкар (2009). «Записи остеодермы титанозавра (Dinosauria: Sauropoda): обзор и первый окончательный образец из Индии». Журнал палеонтологии позвоночных . 29 (1): 165–177. Бибкод : 2009JVPal..29..165D . дои : 10.1671/039.029.0131 . S2CID 85991253 .
- ^ Падиан, Кевин (1997). «Орнитишия». У Кевина Падиана; Филип Дж. Карри (ред.). Энциклопедия динозавров . Сан-Диего: Академическая пресса. стр. 494–498 . ISBN 978-0-12-226810-6 .
- ^ Jump up to: а б Талборн, Ричард А. (1990). Следы динозавров . Лондон, Нью-Йорк: Чепмен и Холл. ISBN 978-0-412-32890-9 .
- ^ Карри, Филип Дж.; Сарджант, Уильям А.С. (1979). «Следы динозавров нижнего мелового периода из каньона Пис-Ривер, Британская Колумбия, Канада» (PDF) . Палеогеография, Палеоклиматология, Палеоэкология . Следы окаменелостей. 28 : 103–115. Бибкод : 1979PPP....28..103C . дои : 10.1016/0031-0182(79)90114-7 . ISSN 0031-0182 .
- ^ Хайман, Либби Генриетта (1942). Сравнительная анатомия позвоночных (2-е изд.). Чикаго, Иллинойс, США: Издательство Чикагского университета. п. 177 .
- ^ Кумбс, Уолтер П. (1972). «Костяное веко Euoplocephalus (Reptilia, Ornithischia)» . Журнал палеонтологии . 46 (5): 637–650. ISSN 0022-3360 . JSTOR 1303019 .
- ^ Додсон, Питер (1993). «Сравнительная краниология Ceratopsia». Американский научный журнал . 293 (А): 200–234. Бибкод : 1993AmJS..293..200D . дои : 10.2475/ajs.293.A.200 .
- ^ Нэйш, Д. (2012). «Птицы». У Майкла К. Бретта-Сурмана; Томас Р. Хольц; Джеймс Орвилл Фарлоу (ред.). Полный динозавр . Издательство Университета Индианы. стр. 379–423.
- ^ Jump up to: а б Полина Карабахал, Ариана; Карбаллидо, Хосе Л.; Карри, Филип Дж. (2014). «Мозговая черепная коробка, нейроанатомия и положение шеи Amargasaurus cazaui (Sauropoda, Dicraeosauridae) и их значение для понимания положения головы у зауроподов». Журнал палеонтологии позвоночных . 34 (4): 870–882. Бибкод : 2014JVPal..34..870P . дои : 10.1080/02724634.2014.838174 . hdl : 11336/19365 . S2CID 85748606 .
- ^ Эдингер, Тилли (1955). «Размер теменного отверстия и органа у рептилий: уточнение» . Вестник Музея сравнительной зоологии . 114 : 1–34. Архивировано из оригинала 1 декабря 2017 года.
- ^ Леман, Томас М. (1996). «Рогатый динозавр из формации Эль-Пикачо в Западном Техасе и обзор цератопсовых динозавров с юго-запада Америки» . Журнал палеонтологии . 70 (3): 494–508. Бибкод : 1996JPal...70..494L . дои : 10.1017/S0022336000038427 . S2CID 132470348 .
- ^ Харрис, Джеральд Д. (2006). «Значение Suuwassea emilieae (Dinosauria: Sauropoda) для внутрисвязей и эволюции жгутиконосцев». Журнал систематической палеонтологии . 4 (2): 185–198. Бибкод : 2006JSPal...4..185H . дои : 10.1017/S1477201906001805 . S2CID 9646734 .
- ^ Апалдетти, Сесилия; Мартинес, Рикардо Н.; Пол, Диего; Саутер, Тибо (2014). «Переописание черепа Coloradisaurus brevis (Dinosauria, Sauropodomorpha) из позднетриасовой формации Лос-Колорадос в бассейне Союза Исчигуаласто-Вилла, северо-запад Аргентины». Журнал палеонтологии позвоночных . 34 (5): 1113–1132. Бибкод : 2014JVPal..34.1113A . дои : 10.1080/02724634.2014.859147 . hdl : 11336/36518 . S2CID 86158311 .
- ^ Лангер, Макс К. (2004). «Базальная Саурисхия». У Дэвида Б. Вейшампеля; Питер Додсон; Х. Осмольска (ред.). Динозаврия (2-е изд.). Беркли: Издательство Калифорнийского университета. стр. 25–46 . ISBN 978-0-520-24209-8 .
- ^ Падиан, Кевин (1997). «Филогения динозавров». У Кевина Падиана; Филип Дж. Карри (ред.). Энциклопедия динозавров . Сан-Диего: Академическая пресса. стр. 646–651 . ISBN 978-0-12-226810-6 .
- ^ Раскин-Гутман, Д. (1997). «Таз, сравнительная анатомия». У Кевина Падиана; Филип Дж. Карри (ред.). Энциклопедия динозавров . Сан-Диего: Академическая пресса. стр. 536–540 . ISBN 978-0-12-226810-6 .
- ^ Шахнер, Эмма Р.; Лайсон, Тайлер Р.; Додсон, Питер (2009). «Эволюция дыхательной системы у нептичьих теропод: данные морфологии ребер и позвонков» . Анатомические записи: достижения в интегративной анатомии и эволюционной биологии . 292 (9): 1501–1513. дои : 10.1002/ar.20989 . ПМИД 19711481 .
- ^ Jump up to: а б Хон, DWE; Хольц, ТР (2017). «Век спинозавров - обзор и пересмотр спинозавров с комментариями по их экологии» . Acta Geologica Sinica — английское издание . 91 (3): 1120–1132. Бибкод : 2017AcGlS..91.1120H . дои : 10.1111/1755-6724.13328 . ISSN 1000-9515 . S2CID 90952478 .
- ^ Ортега, Ф.; Эскасо, Ф.; Санс, JL (2010). «Причудливая горбатая Carcharodontosauria (Theropoda) из нижнего мела Испании» (PDF) . Природа . 467 (7312): 203–206. Бибкод : 2010Natur.467..203O . дои : 10.1038/nature09181 . ПМИД 20829793 . S2CID 4395795 . Архивировано из оригинала (PDF) 8 октября 2018 г. Проверено 27 ноября 2018 г.
- ^ Аллен, Р.; Ксайсанавонг, Т.; Ричир, П.; Хентавонг, Б. (2012). «Первый окончательный азиатский спинозаврид (Dinosauria: Theropoda) из раннего мела Лаоса». Naturwissenschaften . 99 (5): 369–377. Бибкод : 2012NW.....99..369A . дои : 10.1007/s00114-012-0911-7 . ПМИД 22528021 . S2CID 2647367 .
- ^ Эдди, доктор медицинских наук; Кларк, Дж. А. (2011). «Новая информация о краниальной анатомии Acrocanthosaurus atokensis и ее значении для филогении Allosauroidea (Dinosauria: Theropoda)» . ПЛОС ОДИН . 6 (3): e17932. Бибкод : 2011PLoSO...617932E . дои : 10.1371/journal.pone.0017932 . ПМК 3061882 . ПМИД 21445312 .
- ^ Бентон, Майкл Дж. (2012). Доисторическая жизнь . Эдинбург, Шотландия: Дорлинг Киндерсли. п. 338. ИСБН 978-0-7566-9910-9 .
- ^ Прието-Маркес, А.; Кьяппе, LM; Джоши, SH (2012). Додсон, Питер (ред.). «Ламбеозавровый динозавр Magnapaulia laticaudus из позднего мела Нижней Калифорнии, северо-запад Мексики» . ПЛОС ОДИН . 7 (6): e38207. Бибкод : 2012PLoSO...738207P . дои : 10.1371/journal.pone.0038207 . ПМЦ 3373519 . ПМИД 22719869 .
- ^ Пол, GS (2010). Принстонский полевой справочник по динозаврам . Издательство Принстонского университета . п. 208–209 .
- ^ Шмитц, Ларс (2009). «Количественные оценки особенностей зрительного восприятия ископаемых птиц». Журнал морфологии . 270 (6): 759–773. CiteSeerX 10.1.1.537.3346 . дои : 10.1002/jmor.10720 . ПМИД 19123246 . S2CID 309861 .
- ^ Ямасита, Момо; Кониси, Такуя; Сато, Тамаки (18 февраля 2015 г.). «Склеротические кольца у мозазавров (Squamata: Mosasauridae): структуры и таксономическое разнообразие» . ПЛОС ОДИН . 10 (2): –0117079. Бибкод : 2015PLoSO..1017079Y . дои : 10.1371/journal.pone.0117079 . ISSN 1932-6203 . ПМЦ 4334958 . ПМИД 25692667 .
- ^ Jump up to: а б Викариус, МК; Марьянская, ТМ; Вейшампель, Д.Б. (2004). «Анкилозаврия». У Дэвида Б. Вейшампеля; Питер Додсон; Х. Осмольска (ред.). Динозаврия (2-е изд.). Беркли: Издательство Калифорнийского университета. стр. 363–392 . ISBN 978-0-520-24209-8 .
- ^ Продажи, Маркос АФ; Шульц, Сезар Л. (06 ноября 2017 г.). «Таксономия спинозавров и эволюция краниодентальных особенностей: данные из Бразилии» . ПЛОС ОДИН . 12 (11): –0187070. Бибкод : 2017PLoSO..1287070S . дои : 10.1371/journal.pone.0187070 . ISSN 1932-6203 . ПМЦ 5673194 . ПМИД 29107966 .
- ^ Гальтон, премьер-министр (2012). «Стегозавры». В Бретт-Сурмане, МК; Хольц, ТР; Фарлоу, Дж. О. (ред.). Полный динозавр (2-е изд.). Издательство Университета Индианы. стр. 482–504. ISBN 978-0-253-00849-7 .
- ^ Гальтон, Питер М.; Апчерч, Пол (2004). «Прозауропода». У Дэвида Б. Вейшампеля; Питер Додсон; Х. Осмольска (ред.). Динозаврия (2-е изд.). Беркли: Издательство Калифорнийского университета. стр. 232–258 . ISBN 978-0-520-24209-8 .
- ^ Jump up to: а б с д и ж г час Хендрикс, Кристоф; Матеус, Октавио; Араужо, Рикардо (2015). «Предлагаемая терминология зубов теропод (Dinosauria, Saurischia)» . Журнал палеонтологии позвоночных . 35 (5): –982797. Бибкод : 2015JVPal..35E2797H . дои : 10.1080/02724634.2015.982797 . S2CID 85774247 .
- ^ Д'Амор, Доменик К.; Блюменшин, Роберт Дж. (2012). «Использование полосатых следов зубов на костях для прогнозирования размера тела динозавров-теропод: модель, основанная на наблюдениях за питанием Varanus komodoensis , комодского варана». Палеобиология . 38 (1): 79–100. Бибкод : 2012Pbio...38...79D . дои : 10.1017/S0094837300000415 .
- ^ Аблер, Уильям Л. (1992). «Зазубренные зубы динозавров-тираннозаврид и кусающие структуры других животных». Палеобиология . 18 (2): 161–183. Бибкод : 1992Pbio...18..161A . дои : 10.1017/S0094837300013956 . S2CID 88238481 .
- ^ Jump up to: а б Уэллс, СП (1984). « Dilophosaurus wetherilli (Dinosauria, Theropoda), остеология и сравнения». Палеонтографика Абтейлунг А. 185 : 85–180.
- ^ Пол, GS (1988). Хищные динозавры мира . Нью-Йорк: Саймон и Шустер. стр. 271–274 . ISBN 978-0-671-61946-6 .
- ^ Чариг, Эй Джей; Милнер, AC (1997). « Baryonyx walkeri , рыбоядный динозавр из Уэлдена в графстве Суррей» . Бюллетень Лондонского музея естественной истории . 53 : 11–70.
- ^ Уэллс, СП (1970). « Дилофозавр (Reptilia: Saurischia), новое название динозавра» . Журнал палеонтологии . 44 (5): 989. JSTOR 1302738 .
- ^ Бентон, Майкл (2014). Палеонтология позвоночных (4-е изд.). Джон Уайли и сыновья. ISBN 978-1-118-40764-6 .
- ^ Мартинес, Рикардо Н.; Серено, Пол С.; Алькобер, Оскар А.; Коломби, Карина Э.; Ренне, Пол Р.; Монтаньес, Изабель П.; Карри, Брайан С. (14 января 2011 г.). «Базальный динозавр с начала эры динозавров в юго-западной Пангее». Наука . 331 (6014): 206–210. Бибкод : 2011Sci...331..206M . дои : 10.1126/science.1198467 . hdl : 11336/69202 . ПМИД 21233386 . S2CID 33506648 .
- ^ Вейшампель, Дэвид Б. (1991). «Теоретический морфологический подход к замене зубов у низших позвоночных». В Шмидт-Киттлер, Н.; Фогель, Клаус (ред.). Конструктивная морфология и эволюция . Спрингер. стр. 295–310.