Диамантиназавр
| Диамантиназавр | |
|---|---|
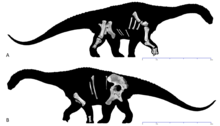
| |
| Скелет голотипа на видах справа (а) и слева (б) | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Динозаврия |
| Клэйд : | Саурисхия |
| Клэйд : | † Зауроподоморфа |
| Клэйд : | † Зауропода |
| Клэйд : | † Макронария |
| Клэйд : | † Сомфоспондилы |
| Клэйд : | † Диамантиназаврия |
| Род: | † Диамантиназавр Хокнулл и др. , 2009 г. |
| Разновидность: | † Д. matildae
|
| Биномиальное имя | |
| † Диамантиназавр матильда Хокнулл и др. , 2009 г.
| |
Диамантиназавр — род титанозавров- зауроподов из Австралии , живший в раннем позднем меловом периоде , около 94 миллионов лет назад. Типовым видом этого рода является D. matildae , впервые описанный и названный в 2009 году Скоттом Хокнуллом и его коллегами на основе находок окаменелостей в формации Винтон . Что означает «Диамантинская ящерица», название происходит от местоположения близлежащей реки Диамантина и греческого слова sauros , «ящерица». Видовой эпитет из австралийской песни Waltzing Matilda также места обитания голотипа , и паратипа . Известный скелет включает большую часть передних конечностей , плечевого пояса , таза , задних конечностей и ребер голотипа, а также одну плечевую кость, лучевую кость и некоторые позвонки паратипа.
История открытия
[ редактировать ]
Голотип диамантиназавра Винтона был впервые обнаружен в течение четырех сезонов раскопок недалеко от , Квинсленд , Австралия . Кости были найдены рядом с голотипом австраловенатора , а также крокодиломорфами и моллюсками . [ 1 ] Два найденных динозавра, известные по экземплярам, каталогизированным как AODF 603 и 604, были описаны в 2009 году Скоттом Хокнуллом и его коллегами. Экземпляр AODF 603 стал основой для рода Diamantinasaurus и вида D. matildae . Название вида является отсылкой к песне «Вальсирующая Матильда», написанной Банджо Патерсоном в Винтоне, а родовое название происходит от реки Диамантина , протекающей неподалеку от типовой местности, в сочетании с греческим словом sauros , что означает «ящерица». AODF 603 Голотип включает правую лопатку , обе плечевые кости , правую локтевую кость , обе неполные кисти , спинные ребра и гастралию , частичный таз и правую заднюю конечность без стопы. [ 3 ] Паратип того же экземпляра включает спинные и крестцовые позвонки, правую грудинную пластинку , которая, как теперь считается, представляет собой остаток клювовидного отростка , лучевую кость и одну фалангу кисти . Все эти кости происходят из AODL 85, прозванного «Место Матильды» на овечьей станции Элдерсли , расположенной примерно в 60 км (37 миль) к западу-северо-западу от Уинтона в центральном Квинсленде . Это местонахождение находится в верхней средней части формации Винтон , которая датируется сеноманом позднего мела . [ 1 ] [ 3 ]
Открытие диамантиназавра положило конец паузе в открытии новых динозавров в Австралии, как первого зауропода, названного за более чем 75 лет. Наряду с австраловенатором , голотип диамантиназавра был назван в честь австралийской песни «Вальсирующая Матильда», причем австраловенатора называли «Банджо», а диамантиназавра - «Матильдой». Винтонотитан , также с сайта, получил прозвище «Клэнси». [ 4 ] [ 5 ] Эта находка, по-видимому, стала крупнейшей находкой динозавров в Австралии, которая была задокументирована после открытия Муттабурразавра в 1981 году. [ 5 ]
По состоянию на 2023 год с момента его первоначального описания было описано еще три экземпляра: каждый экземпляр получил прозвище «Алекс» для AODF 836, «Оливер» для AODF 663 и «Энн» для AODF 0906. [ 6 ] Второй экземпляр, AODF 836, был описан в 2016 году. Он включает части черепа, в том числе левую чешуйчатую кость , почти полную черепную коробку , правую надугольную часть и различные фрагменты. Кроме того, образец также включает атлас , ось , пять других шейных позвонков, три спинных позвонка, дополнительные спинные ребра, части бедра и еще одну правую лопатку. [ 7 ] В 2021 году указанный материал был подробно описан. [ 8 ] В 2022 году третий экземпляр, AODF 663, был назван молодым экземпляром. [ 9 ] В 2023 г. был подробно описан четвертый экземпляр, AODF 0906, состоящий из частичного посткраниального скелета и более полного черепа с неизвестными ранее многочисленными неповрежденными краниальными элементами. [ 6 ]
Описание
[ редактировать ]

Диамантиназавр был титанозавром среднего размера, длиной 16 м (52 фута) и весом до 25 т (28 коротких тонн). [ 10 ] [ 11 ] Как и другие зауроподы, диамантиназавр был крупным четвероногим травоядным . [ 12 ] Со времени первоначального описания единственные серьезные изменения включают неправильную идентификацию «грудинной пластинки», неправильное расположение ручных фаланг III-1 и IV-1 как III-1 и V-1 соответственно, а также идентификацию недостающей части малоберцовой кости. . [ 1 ]
Череп
[ редактировать ]Череп диамантиназавра известен не полностью, сохранились только задняя часть черепа и черепная коробка. Подобно сальтазавру и рапетозавру и в отличие от немегтозавра , надвисочное окно граничило с лобной костью . В отличие от обоих последних родов, Diamantinasaurus имеет низкую надзатылочную часть над черепным отверстием, которая впоследствии менее чем в 1,5 раза превышает высоту базальных бугров (которые имеют отверстие). Однако все эти черты характерны для сальтазавра . У производных титанозавров обнаружено множество других черт, в том числе наклон черепа вниз, зубчатые боковые отростки мозговой оболочки, ненарушенная ямка гипофиза и более центральное расположение отверстия для внутренней сонной артерии . [ 7 ]
Позвонки
[ редактировать ]Как это типично для Titanosauriformes , все шейные и спинные позвонки Diamantinasaurus опистоцельные . и камелярные (много мелких внутренних камер) Осевой позвонок этого рода короткий, что потенциально характерно для Saltasauridae . в отличие от и рапетозавра , Однако, презигапофизы диамантиназавра сальтазавра простираются впереди центра. Лишь в некоторых известных средних дорсальных отделах постспинальная пластинка (гребень на задней поверхности позвоночника) простирается ниже самого позвоночника. [ 7 ] Как и у других базальных зауроподов Europasaurus и Euhelopus , спинные позвонки имеют выемку на вершине задней центральной поверхности, что придает ей сердцевидный вид, в отличие от более продвинутых титанозавров или жираффатитов , которые обладают уплощенными центрами. различаются по форме центра, Хотя Opisthocoelicaudia и Diamantinasaurus они являются единственными титанозаврами, имеющими общий вентральный киль, расположенный внутри резко выраженной впадины под спинной частью. Дорсальные презигапофизы связаны с позвоночником спинопрезигапофизарной пластинкой , которая отсутствует у Opisthocoelicaudia и большинства дорсальных костей Rapetosaurus , а постзигодиапофизарная пластинка, обнаруженная у Diamantinasaurus, также отсутствует у большинства производных титанозавров. Нет никаких признаков сочленения гипосфен-гипантрум , диагностического признака производных титанозавров. Как и Opisthocoelicaudia , Alamosaurus и Lirainosaurus, за исключением других титанозавров, Diamantinasaurus имеет простой неделимый гребень между задним центром и диапофизом ( задняя центродиапофизарная пластинка). ). Плохо сохранившейся особенностью между презигапофизом и центром может быть задняя центропрезигапофизарная пластинка , обнаруженная у некоторых брахиозаврид, базальных титанозавров и Opisthocoelicaudia . Диамантиназавр обладал по меньшей мере пятью, возможно, шестью крестцовыми позвонками . [ 1 ]
Передняя конечность
[ редактировать ]
Почти вся правая передняя конечность известна у Diamantinasaurus , хотя в дополнение к правой известна левая плечевая кость, а также известна левая первая пястная кость, тогда как правая не сохранилась. Диагностика диамантиназавра ( заключается в том, что гленоидное плечевое) сочленение лопатки повернуто наружу, в отличие от всех других сомфоспондилов. Подобно аламозавру и таксонам основания титанозаврии, известен по крайней мере один вентральный отросток, хотя он плохо сохранился. Лопатка сомфоспондилов Diamantinasaurus . прочная, имеет более круглое поперечное сечение, чем у других Коракоид, ошибочно идентифицированный как грудина в первоначальном описании, простой и лишен каких-либо особенностей, контрастируя с Huabeisaurus , Lirainosaurus и Opisthocoelicaudia . Проксимальная титанозавров поверхность плечевой кости длиной 1,068 м (3,50 фута) заметно изогнута, как у производных Opisthocoelicaudia и Saltasaurus . Латеральный угол также имеет квадратную форму, что помещает его внутри сомфоспондила. Как и большинство сомфоспондилов, но в отличие от Euhelopus и Rapetosaurus , Diamantinasaurus имеет смещенный к середине дельтопекторальный гребень . Гребни для прикрепления мышц развиты слабее, чем у Opisthocoelicaudia и Magyarosaurus . В отличие от производных титанозавров, мыщелки, сочленяющиеся с предплечьем, не выражены. Диамантиназавр имеет локтевую кость по сравнению с производными титанозаврами по уровню прочности, а также очень выраженный локтевой отросток . Точно так же радиус диамантиназавра более прочный, чем у всех титанозавров, за исключением Opisthocoelicaudia . Длина локтевой кости составляет 70 см (28 дюймов), а радиус — 67,5 см (26,6 дюйма). [ 1 ]

Ввиду полноты материала передних конечностей отсутствие костей запястья предположил Поропат с соавт. среди сохранившегося материала. (2014), чтобы быть связанным с их подлинным отсутствием в жизни, как у Opisthocoelicaudia и Alamosaurus . Рука Diamantinasaurus сравнительно демонстрирует некоторые плезиоморфные особенности, в том числе: средняя пястная кость является самой длинной (41,2 см (16,2 дюйма) Mc III по сравнению со следующей по длине 37,5 см (14,8 дюйма) Mc II); наличие когтя большого пальца; и наличие нескольких фаланг, имеющих фаланговую формулу 2–1–1–1–1. Однако кисть диамантиназавра полностью цилиндрическая и вертикальная, как и у других титанозавров. Наличие большого количества фаланг у диамантиназавра было использовано Поропатом и др. (2014) предположили, что все титанозавры на самом деле имели окостеневшие фаланги, в отличие от более ранних исследований. Следуя этой логике, они предположили, что у Opisthocoelicaudia и Epachthosaurus , у которых сохранилась одна фаланга четвертого пальца, отсутствие других было связано с их утратой до окаменения предыдущих пальцев, а не с отсутствием. Полное отсутствие сохранившихся фаланг у Alamosaurus , Rapetosaurus , Neuquensaurus и Saltasaurus потенциально возникли из-за разчленения, а не из-за отсутствия окостенения. [ 1 ]
Задняя конечность
[ редактировать ]
левая подвздошная кость , левая и правая лобковые кости , левая и правая седалищные кости сохранились У диамантиназавра и вся правая нога без ступни , хотя некоторые кости сильно фрагментированы и плохо сохранились. Внешняя часть подвздошной кости хорошо сохранилась, но ее размер и хрупкость означают, что внутреннюю сторону невозможно увидеть по анатомическим особенностям. Верхний край подвздошной кости сломан, обнажая многочисленные маленькие внутренние камеры, которые имеются у титанозавров Alamosaurus , Epachthosaurus , Lirainosaurus , Saltasaurus и Sonidosaurus . Как и у других производных титанозавров, передний отросток подвздошной кости расширяется в сторону и вращается, так что вертикальное тело подвздошной кости превращается в горизонтальную полку. Диамантиназавр также демонстрирует производные от зауропода черты округлой подвздошной кости, уменьшенную суставную поверхность седалищной кости и выступ над седалищным сочленением (общий только с Opisthocoelicaudia среди Titanosauriformes). Лобковая кость, как и у продвинутых зауроподов, представляет собой уплощенную кость, лишенную переднего крючка диплодокоидов, но с потенциально аутапоморфными бороздками, окружающими лобковую кость. запирательное отверстие . Сочленение с седалищной костью занимает 46% длины лонной кости, как и у большинства макронарий, но в отличие от Alamosaurus и Opisthocoelicaudia , у которых оно редуцировано. Вся седалищная кость составляет всего 68% длины лобковой кости, как и у других титанозавров, а также расширяется медиально, поэтому все дно таза закрыто. В отличие от некоторых титанозавров, седалищная кость диамантиназавра не имеет сужения по ширине или выступающего внутрь фланца. У Diamantinasaurus также отсутствует заметный мышечный рубец для внутреннего сгибателя большеберцовой кости 3 на стороне дистальной части седалищной кости, что является диагностическим признаком этого таксона среди Neosauropoda . [ 1 ]

Бедренная кость длиной 1,345 м (4,41 фута) примерно в два раза шире своей длины, как и у других производных зауроподов, хотя она слегка раздавлена. Раздавливание не помешало сохранению гребня linea intermuscularis cranialis , также присутствующего у Saltasaurus , Neuquensaurus , Bonatitan , Rocasaurus и Alamosaurus . Как это типично для зауроподов, головка бедренной кости немного выше большого вертела , имеется умеренный вертельный выступ. Присутствует умеренная латеральная выпуклость, над которой бедренная кость смещена медиально, как и у большинства макронарий, за исключением Opisthocoelicaudia , Saltasaurus и Rapetosaurus . Мыщелки сочленения с большеберцовой и малоберцовой костью простираются высоко на заднюю поверхность бедренной кости, но в отличие от Neuquensaurus и Opisthocoelicaudia не заходят на переднюю поверхность. Мыщелок малоберцовой кости разделен депрессией, которая имеет небольшой гребень, также встречающийся у Magyarosaurus и других титанозавров, хотя его выступание уникально для Diamantinasaurus . Мыщелок малоберцовой кости больше большеберцовой кости и простирается дальше вниз, придавая бедренной кости скошенный вид, что потенциально является диагностическим признаком Saltasauridae, но также встречается у Рапетозавр и Донгбейтитан нетитанозавр [ 1 ]
Большеберцовая кость составляет 59% длины бедренной кости и, как обычно для неозауроподов, на проксимальной поверхности шире, чем длинна. Диамантиназавр имеет множество ямок и гребней на голени, которые не наблюдаются у других зауроподов, что делает их набором диагностических признаков. Как и у многих титанозавров, дистальный конец большеберцовой кости расширен более чем в два раза по ширине средней части тела, хотя тонкий выступ вдоль средней части тела может быть диагностическим признаком диамантиназавра . Первоначально восстановленная недостающая часть стержня малоберцовой кости имеет длину 76,9 см (30,3 дюйма) и имеет среднюю прочность, хотя и близка к изящной. Кость плохо сохранилась, но все еще имеет диагностическое расширение рубца малоберцовой мышцы и диагностический медиальный гребень с окружающими бороздками. Как и у многих титанозавроформ, астрагал диамантиназавра менее чем в 1,5 раза шире длины, а проксимальная поверхность разделена на восходящий отросток и ямку для большеберцовой кости. На внешней стороне таранной кости также имеется неглубокая ямка для малоберцовой кости, придающая кости подтреугольную форму. В переднем основании восходящего отростка нет впадин и отверстий, что типично для Эузавроподы . Отросток на задней стороне тела астрагала уникален среди всех зауроподов, что делает его аутапоморфией диамантиназавра . [ 1 ]
Классификация
[ редактировать ]Когда он был первоначально описан, Diamantinasaurus был отнесен к Lithostrotia incertae sedis . В обеих филогениях, в которые он был помещен, Diamantinasaurus находился либо недалеко от Saltasauridae , либо сестринским таксоном Opisthocoelicaudia внутри семейства. [ 3 ] В исследовании 2014 года было обнаружено, что этот род занимал как литостротии в обеих крупных филогениях, занимая относительно производное положение в Titanosauria. Их первая филогения была изменена по сравнению с филогенией Карбадилло и Сандера (2014), причем матрица косвенно основана на филогении Уилсона 2002 года. На этой кладограмме было обнаружено, что Diamantinasaurus является сестринским таксоном Tapuiasaurus , их родством за пределами Saltasauridae . В этой филогении поддержка Бремера для каждой группы составляла не более 1. Пять особенностей скелета поддерживали размещение диамантиназавра в литостроции. [ 1 ]


В том же исследовании отношения с использованием Mannion et al. (2013). Они разрешились с Диамантиназавром как сальтазавридом, сестрой Opisthocoelicaudia , и Dongyangosaurus как следующим ближайшим. Было обнаружено, что два признака подтверждают размещение Диамантиназавра в Литостротии, а третий не удалось оценить. [ 1 ]
Другой филогенетический анализ, проведенный в 2016 году, частично воспроизведенный ниже, показал, что он является нелитостротианским титанозавром и сестринским таксоном современного саванназавра . [ 7 ] [ 13 ]

Горскак и О'Коннор (2019) в своем описании Мньямавамтуки выявили Диамантиназавра как сальтазаврида с помощью экономного филогенетического анализа, в то время как байесовский филогенетический анализ с переменной скоростью показал, что он находится сразу за пределами Saltasauridae. [ 14 ]
Исследование 2021 года выявило аналогичную топологию, обнаружив тесную связь с саванназавром, а также с сармиентозавром из раннего позднего мела Патагонии, чей череп имел сходство с упомянутым черепным материалом диамантиназавра . Клада, содержащая эти таксоны, получила название Diamantinasauria . [ 8 ] Бистон и др. (2024) рассматривали австралотитана как младший синоним диамантиназавра или неопределенного вида диамантиназавра-титанозавра, поскольку голотип австралотитана не обладает различимыми аутапоморфными признаками, позволяющими классифицировать его как действительный род, и имеет много общего с известными экземплярами диамантиназавра . [ 15 ]
Палеобиология
[ редактировать ]
Рост
[ редактировать ]В 2011 году был описан самый маленький положительно идентифицированный эмбрион титанозавра. Хотя он был обнаружен в Монголии , эмбрион имеет большинство общих черт с диамантиназавром и рапетозавром . Эмбрион из относительно сферического яйца размером 87,07–91,1 мм (3,428–3,587 дюйма) был идентифицирован как сохранившийся до литостротиана. Эмбрион был слегка крепким, промежуточным между крепостью рапетозавра и диамантиназавра . Яйцо является частью целого места гнездования литостротиевых титанозавров. Датировка региона также позволяет предположить, что это яйцо предшествует яйцам из Аука-Мауэво в Аргентине и что яйца были отложены в раннем меловом периоде . [ 16 ]
Палеоэкология
[ редактировать ]
Диамантиназавр был найден примерно в 60 километрах (37 миль) к северо-западу от Винтона, недалеко от станции Элдерсли. [ 3 ] Он был извлечен из богатой окаменелостями части формации Винтон , возраст которой можно датировать примерно 93 миллионами лет назад. [ 2 ] Диамантиназавр был найден в слое глины между слоями песчаника , что интерпретируется как отложение старицы . На этом месте также был найден австраловенатор , который был напрямую связан с диамантиназавром , двустворчатыми моллюсками , рыбами , черепахами , крокодилами и различными растениями . В формации Уинтон была фауна, включающая двустворчатых моллюсков, брюхоногих моллюсков , насекомых , двоякодышащих рыб Metaceratodus , черепах, крокодилов Isisfordia , птерозавров и несколько типов динозавров, таких как вышеупомянутый австраловенатор , зауроподы Wintonotitan , Savannasaurus и Austrosaurus , а также безымянные анкилозавры . и гипсилофодонты . Кости диамантиназавра можно отличить от других зауроподов из-за общей прочности, а также множества специфических особенностей. Растения, известные из формации, включают папоротники , гинкго , голосеменные и покрытосеменные растения . [ 3 ]
Ссылки
[ редактировать ]- ^ Jump up to: а б с д и ж г час я дж к л Поропат, Сан-Франциско; Апчерч, П.; Мэннион, Вашингтон; Хокнулл, ЮАР; Кир, BP; Слоан, Т.; Синапий, GHK; Эллиот, окружной прокурор (2014). «Ревизия динозавра-зауропода Diamantinasaurus matildae Hocknull et al. 2009 из середины мела Австралии: последствия для гондванского расселения титанозаврообразных» . Исследования Гондваны . 27 (3): 995–1033. дои : 10.1016/j.gr.2014.03.014 . hdl : 10044/1/27497 .
- ^ Jump up to: а б Такер, RT; Робертс, EM; Ху, Ю.; Кемп, AIS; Солсбери, Юго-Запад (2013 г.). «Ограничения по возрасту обломочных цирконов формации Винтон, Квинсленд: контекстуализация фауны динозавров позднего мела в Австралии». Исследования Гондваны . 24 (2): 767–779. Бибкод : 2013GondR..24..767T . дои : 10.1016/j.gr.2012.12.009 .
- ^ Jump up to: а б с д и Хокнулл, Скотт А.; Уайт, Мэтт А.; Тишлер, Трэвис Р.; Кук, Алекс Г.; Каллеха, Наоми Д.; Слоан, Триш; Эллиотт, Дэвид А. (2009). Серено, Пол (ред.). «Новые среднемеловые (последние альбы) динозавры из Винтона, Квинсленд, Австралия» . ПЛОС ОДИН . 4 (7): е6190. Бибкод : 2009PLoSO...4.6190H . дои : 10.1371/journal.pone.0006190 . ПМК 2703565 . ПМИД 19584929 .
- ^ Мюссер, А. (3 июня 2010 г.). «Виды животных: Diamantinasaurus matildae » . Австралийский музей.
- ^ Jump up to: а б «Новые динозавры найдены в Австралии» . Новости Би-би-си . 3 июля 2009 г.
- ^ Jump up to: а б Поропат, Сан-Франциско; Мэннион, Вашингтон; Ригби, СЛ; Дункан, Р.Дж.; Пентленд, штат Айдахо; Бевитт, Джей-Джей; Слоан, Т.; Эллиотт, Д.А. (2023). «Почти полный череп динозавра-зауропода Diamantinasaurus matildae из верхнемеловой формации Винтон в Австралии и значение для ранней эволюции титанозавров» . Королевское общество открытой науки . 10 (4). 221618. дои : 10.1098/rsos.221618 . ПМЦ 10090887 . ПМИД 37063988 .
- ^ Jump up to: а б с д Поропат, Сан-Франциско; Мэннион, Вашингтон; Апчерч, П.; Хокнулл, ЮАР; Кир, BP; Кундрат, М.; Тишлер, ТР; Слоан, Т.; Синапий, GHK; Эллиотт, Дж.А.; Эллиотт, Д.А. (2016). «Новые австралийские зауроподы проливают свет на палеобиогеографию динозавров мелового периода» . Научные отчеты . 6 : 34467. Бибкод : 2016NatSR...634467P . дои : 10.1038/srep34467 . ПМК 5072287 . ПМИД 27763598 .
- ^ Jump up to: а б Поропат, Стивен Ф; Кундрат, Мартин; Мэннион, Филип Д.; Апчерч, Пол; Тишлер, Трэвис Р.; Эллиотт, Дэвид А. (20 января 2021 г.). «Второй экземпляр позднемелового австралийского динозавра-зауропода Diamantinasaurus matildae предоставляет новую анатомическую информацию о черепе и шее ранних титанозавров» . Зоологический журнал Линнеевского общества . 192 (2): 610–674. doi : 10.1093/zoolinnean/zlaa173 . ISSN 0024-4082 .
- ^ Ригби С.Л., Поропат С.Ф., Маннион П.Д., Пентленд А.Х., Слоан Т., Рамболд С.Дж., Вебстер CB, Эллиотт Д.А. (2022). «Молодая особь Diamantinasaurus matildae (Dinosauria: Titanosauria) из верхнемеловой формации Винтон в Квинсленде, Австралия, имеющая значение для онтогенеза зауроподов» . Журнал палеонтологии позвоночных . 41 (6): e2047991. дои : 10.1080/02724634.2021.2047991 . S2CID 248187418 .
- ^ Хольц, ТР; Рей, Л.В. (2007). Динозавры: самая полная и современная энциклопедия для любителей динозавров всех возрастов . Случайный дом. ISBN 9780375824197 . Список родов Holtz 2012 Информация о весе
- ^ Ада Дж. Клинкхамер; Генрих Мэллисон; Стивен Ф. Поропат; Джордж Х.К. Синапиус; Стивен Роу (2018). «Трехмерное скелетно-мышечное моделирование задней конечности зауроподоморфа: влияние изменения позы на мышечный рычаг» . Анатомическая запись . 301 (12): 2145–2163. дои : 10.1002/ar.23950 . ПМИД 30299598 . S2CID 52940848 .
- ^ Апчерч, П.; Барретт, премьер-министр; Додсон, П. (2004). «Зауропода». В Вейшампеле, Дэвид Б.; Додсон, Питер; Осмольская, Гальшка (ред.). Динозаврия (Второе изд.) . Издательство Калифорнийского университета. стр. 259–322 . ISBN 978-0-520-24209-8 .
- ^ Сен-Флер, Николас (20 октября 2016 г.). «Знакомьтесь, новый титанозавр. Вы можете называть его Уэйдом» . Нью-Йорк Таймс . Проверено 21 октября 2016 г.
- ^ Горчак, Э.; О'Коннор, П. (2019). «Новый африканский титанозавровый динозавр-зауропод из формации Галула среднего мела (пачка Мтука), рифтовый бассейн Руква, юго-западная Танзания» . ПЛОС ОДИН . 14 (2): e0211412. Бибкод : 2019PLoSO..1411412G . дои : 10.1371/journal.pone.0211412 . ПМК 6374010 . ПМИД 30759122 .
- ^ Бистон, СЛ; Поропат, Сан-Франциско; Мэннион, Вашингтон; Пентленд, штат Айдахо; Энчелмайер, MJ; Слоан, Т.; Эллиотт, Д.А. (2024). «Переоценка разнообразия динозавров-зауроподов в верхнемеловой формации Винтон в Квинсленде, Австралия, посредством 3D-оцифровки и описания новых экземпляров» . ПерДж . 12 . е17180. дои : 10.7717/peerj.17180 . ПМК 11011616 . ПМИД 38618562 .
- ^ Грелле-Тиннер, Г.; Сим, СМ; Ким, Д.Х.; Тримби, П.; Хига, А.; Ан, СЛ; Ох, ХС; Ким, Ти Джей; Карджилов, Н. (2011). «Описание первого литостротиального эмбриона титанозавра in ovo с нейтронной характеристикой и последствиями для миграции и расселения литостротианского апта». Исследования Гондваны . 20 (2–3): 621–629. Бибкод : 2011GondR..20..621G . дои : 10.1016/j.gr.2011.02.007 .











