Саванназавр
| Саванназавр | |
|---|---|
| |
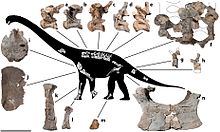
| Схема скелета и избранные окаменелости | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Динозаврия |
| Клэйд : | Саурисхия |
| Клэйд : | † Зауроподоморфа |
| Клэйд : | † Зауропода |
| Клэйд : | † Макронария |
| Род: | † Саванназавр Поропат и др . 2016 год |
| Разновидность: | † С. elliottorum
|
| Биномиальное имя | |
| † Savannasaurus elliottorum Поропат и др . 2016 год
| |
Savannasaurus — род титанозавровых динозавров- зауроподов из позднемеловой формации Винтон в Квинсленде , Австралия . Он содержит один вид, Savannasaurus elliottorum , названный в 2016 году Стивеном Поропатом и его коллегами. Голотип и единственный известный экземпляр, первоначально названный «Уэйд», является наиболее полным экземпляром австралийского зауропода и хранится в музее Австралийской эпохи динозавров . Динозавры, известные из современных пород, включают его близкого родственника Diamantinasaurus и теропода Australovenator ; связанные зубы позволяют предположить, что австраловенатор , возможно, питался образцом голотипа.
длиной 15 метров (49 футов) Саванназавр был титанозавром среднего размера. Он примечателен своими широкими бедрами, ширина которых в самых широких точках составляла более 1,1 метра (3 фута 7 дюймов). Это позволило бы распределить вес его тела более равномерно, а также создать крепкую плечевую кость (кости плеча) и, возможно, также высокую таранную кость (лодыжку). В сочетании с гибким позвоночником эти черты позволили бы саванназаврам лучше передвигаться по илистой почве пойм , на которых они жили. Другие линии титанозавров также демонстрируют некоторые из этих черт, которые могли быть приобретены независимо от аналогичного давления окружающей среды.
Различные черты позволяют предположить, что саванназавр был раннерасходящимся (или базальным ) представителем Titanosauria, но не членом более специализированной группы Lithostrotia . Среди них отсутствие сочленений гипосфен-гипантр в его позвонках, что было одной из черт, повышающих гибкость, которая могла появиться у многих линий титанозавров. Палеобиогеографический анализ предполагает, что предки саванназавра и диамантиназавра были либо из Азии, либо с одного из южных континентов Гондваны ; тем не менее, они, вероятно, мигрировали в Австралию через Антарктиду около 105–100 миллионов лет назад.
Открытие и присвоение имени
[ редактировать ]
В марте 2005 года первый экземпляр саванназавра был обнаружен Дэвидом Эллиотом, основателем Австралийской эпохи динозавров (AODF), вместе с его женой Джуди и их детьми. Образец, теперь внесенный в каталог как AODF 660, был найден на овцеводческой станции Бельмонт, в 60 километрах (37 миль) к северо-востоку от Винтона, Квинсленд ; этому месту был присвоен номер AODF 82, и его неофициально называют «сайтом Хо-Хум». [ 1 ] В ходе раскопок Музея Квинсленда и сотен волонтеров этот экземпляр был обнаружен в июле и сентябре того же года. Алевролитовая , конкреция содержащая образец, была расколота по заполненным гипсом трещинам с помощью отбойных молотков и долот, при этом куски были отмечены для облегчения повторной сборки. Подготовка образца заняла около десяти лет. [ 2 ] с использованием воздушных писцов и микроразъемов. клеи, такие как суперклей и аралдит Затем для соединения частей использовались . Метки позволили ориентировать образец, когда он был реконструирован, что послужило основой для трехмерной карты местности, построенной с помощью фотограмметрии . [ 3 ]
AODF 660 состоит примерно из 40 костей: одной шейной (шея), спинных с третьего по десятый ( туловище), как минимум четырех крестцовых (бедро) и как минимум пяти хвостовых (хвостовых) позвонков ; шейные и (фрагментарные) спинные ребра; части плечевого пояса, включая фрагментированную лопатку , левый клювовидный отросток и обе грудинные пластинки ; части передних конечностей, включая части обеих плечевых костей , фрагментированную локтевую кость , левую лучевую кость и кости кисти (левая первая-пятая пястные кости , правая четвертая пястная кость и две фаланги ); сросшиеся бедренные кости, а именно левая и правая лобковая и седалищная кости ; кости стопы (левая таранная кость и правая третья плюсневая кость ); и другие фрагменты. [ 1 ] Первоначально считалось, что одна из костей стопы принадлежала тероподу. [ 4 ] в то время как пястные кости первоначально также интерпретировались как происходящие из противоположных рук. [ 3 ] Этот экземпляр является наиболее полным экземпляром зауропода, известным из Австралии, и составляет примерно от 20% до 25% животного. [ 2 ]

Кости сохранились на площади менее 20 м2. 2 (220 кв. футов). Хотя спинные позвонки не были сочленены, они сохранились последовательно перед бедренными костями, что позволяет их идентифицировать. Ребра на левой стороне, по-видимому, были раздавлены еще до окаменения, прежде чем все они подверглись дальнейшей фрагментации. Пятый спинной позвонок и обе плечевые кости имеют следы растоптанных другими динозаврами перед окаменением, что также может объяснить разбросанные кости; однако кости были менее разбросаны, чем другие экземпляры динозавров из современных пород. Третьи раскопки, проведенные в сентябре 2006 года, не обнаружили никаких дополнительных останков, но в 2013 году на поверхности был обнаружен частичный центр хвостового позвонка. [ 3 ]
Получив прозвище «Уэйд», [ 2 ] AODF 660 был официально описан и назван в 2016 году Стивеном Поропатом и его коллегами в статье, в которой также описывался череп, принадлежащий диамантиназурусу . Они идентифицировали AODF 660 как голотип нового рода и вида Savannasaurus elliottorum ; Родовое название Savannasaurus , от Taíno zavana (корень слова «саванна»), относится к среде, в которой он был найден, а видовое название elliottorum отдает дань уважения семье Эллиоттов и их вкладу в австралийскую палеонтологию. [ 1 ] Предварительное анатомическое описание AODF 660 в этой статье было дополнено в 2020 году другой статьей многих тех же авторов, в которой дано полное описание его остеологии . [ 3 ]
Описание
[ редактировать ]
Саванназавр среднего размера был титанозавром , длиной около 15 метров (49 футов), весом около 20 тонн (22 коротких тонны) и высотой в плечах около 3 метров (9,8 футов). [ 4 ]
Позвоночник
[ редактировать ]Как и у большинства других представителей Titanosauriformes , позвонки саванназавра были камелярными или пневматизированы небольшими отверстиями, образованными воздушными мешками, и у них не было сочленений гипосфен-гипантр , которые позволяли соседним позвонкам сцепляться. Шейные и спинные позвонки саванназавра были опистоцельными или имели центр (тело позвонка), выпуклое спереди и вогнутое сзади. У всех остальных представителей Eusauropoda шейные позвонки опистоцельные, а у всех остальных представителей Macronaria - опистоцельные спинные кости. [ 1 ] Необычно то, что передняя сочленяющаяся поверхность одного из каудальных центров была волнистой, вогнутой в верхней половине и выпуклой в нижней. Поропат и его коллеги предположили, что это отличительная особенность саванназавра . Остальные хвостовые части были амфицельными или имели центры, вогнутые на обоих концах, что также было редкостью среди титанозавров, за исключением Baotianmansaurus и Dongyangosaurus . [ 3 ]
Из-за своих пропорций считается, что единственный известный шейный позвонок саванназавра произошел из задней части шеи. В отличие от большинства титанозавров, но похожего на «предковое» ( плезиоморфное ) состояние зауроподов, позвонок имел тонкий киль, идущий продольно вдоль его нижней поверхности; у Мендозазавра , Оверозавра , Австропосейдона и Рапетозавра Такие кили есть также . По бокам позвонка имелась хорошо развитая продольная ямка , или выемка, простирающаяся почти на всю его длину, в отличие от многих титанозавров, у которых она была слабой или отсутствовала вообще. В отличие от более специализированных титанозавров, таких как Saltasauridae , парапофиз (одно из реберных сочленений) был ограничен передней половиной центра. Соседние фрагменты шейных ребер позволили предположить, что они были относительно длинными, простираясь как минимум на три центра. [ 1 ] [ 3 ]

Как и у многих титанозавров, сочленяющиеся поверхности спинных центров у саванназавра были короче по вертикали, чем в ширину. Спинные кости имеют слабо развитые гребни, окаймляющие боковые стороны их нижних граней, как у Diamantinasaurus и Opisthocoelicaudia (где они были лучше развиты), но у саванназавра отсутствует киль на нижней части центральной части, как у этих видов. Как и у большинства других представителей Somphospondyli , по бокам центральной части имелись каплевидные пневматические отверстия. Суставные отростки в передней части центра, известные как презигапофизы, были соединены костным листком ( пластинкой ), транспрезигапофизарной пластинкой; в передней спинной части этот лист имел V-образную форму, как у большинства титанозавров, но в задней части спинной части он был практически плоским, в отличие от большинства других титанозавров. Примечательно, что пластинки спинной части саванназавра были двусторонне асимметричными ; левая сторона позвонков была бы лучше укреплена дополнительными пластинками. Также в отличие от Opisthocoelicaudia дорсальная нервные отростки на вершине центров увеличивались в высоту по направлению к задним дорсальным отделам, не были разделены на две части и были выше соответствующих им суставных поверхностей. [ 1 ] [ 3 ]
Помимо волнистой сочленяющейся поверхности в одном из передних каудальных центров (хвостовая часть «А»), две передние каудальные кости (каудальные кости «А» и «В») саванназавра имеют еще одну отличительную особенность. Как и у нетитанозаврового сомфоспондилана Padillasaurus , передняя часть хвостового отдела имела неглубокие пневматические выемки по бокам, окружающие небольшие отверстия, тогда как у большинства представителей Somphospondyli есть только отверстия, а не выемки. [ 1 ] Эта характеристика ранее считалась отличительной характеристикой несомфоспондилановых брахиозавридов . [ 5 ] Два других хвостовых позвонка (хвостовые позвонки «C» и «D») известны из задней части хвоста. Центры каудальных костей B, C и D были примерно одинаковой длины, что позволяет предположить, что длина центра существенно не различалась. Однако обе сочленяющие поверхности были шире, чем высота в каудальной части C, а передняя была выше ширины в каудальной части D, что позволяет предположить, что она была дальше вперед, чем каудальная D. Как и у других титанозавроформ, отростки, окружающие нервный канал — ножки — в каудальных отделах C и D составляли две трети длины центра и были смещены вперед относительно средней линии. [ 3 ]
Плечи и передние конечности
[ редактировать ]Две отличительные характеристики саванназавра обнаружены в грудинных пластинах. Во-первых, они имели D-образную форму с прямыми внешними краями, если смотреть снизу, а не почкообразную форму, как у других титанозавров. Во-вторых, у них отсутствовали длинные гребни, которые присутствовали на нижней поверхности у большинства представителей Neosauropoda . В целом нижние поверхности лишены борозд и ямок на верхних поверхностях, свидетельствующих о наличии хрящевых покрытий. Однако, как и у большинства других титанозавров, длина грудинных пластинок должна была составлять не менее 65% длины плечевых костей (у саванназавра это соотношение составляло примерно 71%). Коракоид саванназавра имел овальную форму при виде сбоку, в отличие от характерных для сальтазавридов четырехугольных коракоидов, а суставная ямка на нем (которая сочленялась с плечевой костью) была намного крупнее, чем у диамантиназавра . [ 1 ] В отличие от большинства титанозавроформ, за исключением Daxiatitan , Ligabuesaurus и Sauroposeidon , сочленение с лопаткой в задней части кости было выше, чем длина всей кости спереди назад. [ 3 ]
Как и у диамантиназавра , опистоцеликаудии и сальтазавридов, плечевая кость саванназавра была крепкой. Радиус был скручен вокруг своей оси, причем длинные оси двух концов были ориентированы в разных плоскостях, что иначе наблюдалось только у Huabeisaurus , Epachthosaurus и Rapetosaurus . Более типичными были максимальная ширина ее верхнего конца (33% длины кости) и наличие мощного гребня у задненаружного угла (межкостного гребня). Как и у всех неозауроподов, пальцы рук саванназавра были устроены как подкова. Как и у Диамантиназавра и Винтонотитана , самыми длинными были третьи пястные кости, за ними следовали вторая, первая, четвертая и пятая. Третья пястная кость составляла 49% длины лучевой кости, что было коротко для титанозавра, но составляло не менее 45%, как у всех макронарий. Отличительной особенностью является то, что нижняя часть четвертой пястной кости имела форму песочных часов, а не трапециевидную или шестиугольную форму. [ 1 ] При жизни пястные кости были бы связаны сверху и раздвинуты снизу. Наличие фаланг необычно; у всех титанозавров, кроме саванназавра и диамантиназавра, руки были образованы только пястными костями. Все пальцы, кроме первой, вероятно, имели фаланги, хотя их сочленяющие поверхности на нижней части пястных костей не простирались вперед, как можно было бы ожидать. [ 3 ]
Таз и задние конечности
[ редактировать ]Самой отличительной особенностью саванназавра была ширина его таза. [ 4 ] - его крестец имел ширину 1,07 метра (3 фута 6 дюймов), а сросшийся седалищно-лобковый комплекс - не менее 1,14 метра (3 фута 9 дюймов) в ширину. Его грудные пластины имели ширину около 85 сантиметров (2 фута 9 дюймов), чтобы вместить это. Хотя титанозавры в целом были широкотелыми, эти пропорции делали саванназавра чрезвычайно широкобедрым, уступая только Opisthocoelicaudia и сальтазавринам, таким как Neuquensaurus . [ 1 ] Другие пропорции таза саванназавра также были необычными. Высота сочленения седалищной кости на лобке составляла 56% от длины кости, что выше, чем у большинства титанозавров. Седалищная кость у саванназавра была относительно короткой и составляла 63% длины лобковой кости; она была как минимум на 70% длиннее у всех других титанозавров, кроме Rapetosaurus (54%) и Opisthocoelicaudia (64%). Характерной чертой, которая отличала саванназавра от любого другого зауропода, является то, что длина седалищной кости спереди назад составляла всего 42% ширины кости. На седалищной кости подвздошная ножка, один из отростков, окружающих вертлужную впадину (тазобедренный сустав), составляла всего 32% длины кости, что ниже, чем у большинства других зауроподов. Кроме того, ширина переднего конца седалищной кости составляла 49% длины кости; только у Диамантиназавра было такое же высокое соотношение (51%). [ 3 ]
Еще одной отличительной особенностью саванназавра является наличие тонкого гребня, который отделяет переднюю верхнюю часть лобка от задней пластины. Гребень параллелен другому гребню чуть ниже отверстия, известного как запирательное отверстие ; этот последний гребень соответствует множественным гребням у Diamantinasaurus . Также, как и у Диамантиназавра , на кончике лобка имелся расширенный «ботинок». На седалищной кости гребень сзади, отделявший ее от лобковой кости, простирался дальше в сторону, чем у Diamantinasaurus , но меньше, чем у Wintonotitan . Ниже, в области лодыжки, высота астрагала превышала его диаметр как спереди назад, так и из стороны в сторону, что необычно для титанозаврообразных. [ 1 ] Отличительными особенностями являются два необычно низких соотношения астрагала: поперечный диаметр составлял 87% высоты, а поперечный диаметр составлял 98% поперечного диаметра. В отличие от Диамантиназавра , на таранной кости не было выступа ниже того места, где она могла бы сочленяться с малоберцовой костью . [ 3 ]
Классификация
[ редактировать ]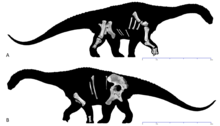
Для описания в 2016 году Поропат и его коллеги провели филогенетический анализ , чтобы проверить сходство саванназавра и нового образца черепа диамантиназавра . Они добавили эти экземпляры в набор данных, созданный Филипом Мэннионом (одним из соавторов описания саванназавра ) и коллегами для переописания лусотитана в 2013 году . [ 6 ] Анализ поместил Savannasaurus и Diamantinasaurus вместе в кладу (единую группу), близкую к корню Titanosauria, вне производной (специализированной) группы Lithostrotia . [ 1 ] Аналогичные результаты были получены Рафаэлем Ройо-Торресом и его коллегами при описании Сориатитана в 2017 году ; [ 7 ] Маннион и его коллеги в описании Вуиврии в 2017 году ; [ 8 ] Александр Аверьянов и Владимир Ефимов в описании « Волгатитана» в 2018 году ; [ 9 ] Бернардо Гонсалес Рига и его коллеги в их переописании Мендозазавра в 2018 году ; [ 10 ] и Педро Мочо и его коллеги в их описаниях неопределенного литострота в 2019 году. [ 11 ] и Океанотитан . [ 12 ]
В 2020 году Поропат и его коллеги отметили ряд особенностей, подтверждающих положение саванназавра вне литостроции: горизонтальные транспрезигапофизарные пластинки, отсутствие сужения позвонков крестца, амфицельные каудальные центры, пневматические экскавации на передних каудальных отделах, Грудные пластинки D-образной формы, наличие фаланг и отсутствие продольного гребня на сторона лобка. В то же время отсутствие гипосфен-гипантральных сочленений в дорсальных и хвостовых позвонках свидетельствует против более базального (неспециализированного или направленного к корням) положения, учитывая их наличие у многих нетитанозавровых титанозавроформ и редкость среди титанозавров. [ 6 ] Некоторые титанозавры также имели такие сочленения; у базального Andesaurus и литострота Epachthosaurus они были в спинных отделах, тогда как у литостротиана Epachthosaurus , Opisthocoelicaudia , Volgatitan и Malawisaurus они были в хвостовом отделе. Однако Поропат и его коллеги признали возможность того, что они могли быть приобретены независимо несколько раз среди титанозавров. [ 3 ]

Тем не менее, другие филогенетические анализы, основанные на наборе данных Манниона и его коллег за 2013 год, дали другие результаты. Ройо-Торрес и его коллеги повторили свой анализ 2017 года после удаления титанозавра из формации Клеверли , чубутисавра и анголатитана ; вместо этого они обнаружили, что саванназавр и диамантиназавр были последовательно базальными к кладе, состоящей из Baotianmansaurus и Dongyanosaurus . [ 7 ] Саванназавр имел общие амфицельные каудальные центры с обоими родами, различные особенности позвоночных пластинок с Baotianmansaurus , а также некилевидные дорсальные центры и отсутствующий продольный гребень на стороне лобковой кости с Dongyanosaurus . Однако, в отличие от саванназавра , у Baotianmansaurus были сочленения гипосфен-гипантрум, и оба рода имели раздвоенные дорсальные нервные отростки (т. е. разделенные на две части). [ 3 ]
Аналогичным образом, для статьи 2019 года, называющей новую кладу , баотианманзавра , донгианозавра Colossosauria, Гонсалес Рига и его коллеги расширили свой филогенетический анализ 2018 года и обнаружили неразрешенную политомию саванназавра и клады более производных титанозавров. Диамантиназавр был поставлен в более производное положение как сестринская группа Lithostrotia. [ 13 ] Также в 2019 году Джулиан Сильва мл. и расширил анализ 2018 года, проведенный Гонсалесом Ригой и его коллегами на предмет их переописания Уберабатитана . Они нашли саванназавра за пределами Титанозаврии в политомии с Euhelopus , Erketu , Qiaowanlong , кладой Tangvayosaurus и Phuwiangosaurus , а также другими производными сомфоспондилами; Они также обнаружили, что Diamantinasaurus и Baotianmansaurus образуют кладу внутри Saltasauridae. [ 14 ] Поропат и его коллеги сочли свои результаты необычными в 2020 году. [ 3 ] В своем переописании Jiangshanosaurus и Dongyanosaurus в 2019 году Маннион и его коллеги обнаружили как Savannasaurus , так и Diamantinasaurus внутри Saltasauridae, используя одинаковые взвешивающие характеристики в своем наборе данных, но использование подразумеваемого взвешивания устранило этот результат. [ 15 ]
Филогенетическое древо анализа Поропата и его коллег, проведенного в 2016 году, воспроизведено ниже слева с акцентом на соответствующие клады. [ 1 ] Альтернативное дерево, созданное Ройо-Торресом и его коллегами в 2017 году из анализа, в котором были удалены титанозаврообразные Cloverly, Chubutisaurus и Angolatitan , также показано справа. [ 7 ]
Топология А: Поропат и др. (2016) [ 1 ]
| Титанозаврообразные |
| ||||||||||||||||||||||||||||||
Топология B: Ройо-Торрес и др. (2017) [ 7 ]
| Титанозаврообразные |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Палеобиогеография
[ редактировать ]южный суперконтинент Гондвана . период распался Когда в меловой , фауна динозавров стала более разнообразной [ 16 ] Традиционные гипотезы предполагают, что фауны гондванских континентов (включая Австралию) были тесно связаны. [ 17 ] но было обнаружено, что австралийские динозавры среднего мела более тесно связаны со своими лавразийскими (северными) собратьями, несмотря на то, что Лавразия и Гондвана уже были разделены океаном Тетис в течение значительного периода времени. [ 18 ] [ 19 ] В 2016 году Поропат и его коллеги проверили эту гипотезу, проведя палеобиогеографический анализ с использованием результатов филогенетического анализа. Используя современную географию континентов, они установили, что предки австралийских зауроподов Savannasaurus , Diamantinasaurus и Wintonotitan , вероятно, были азиатами. Учитывая географию мелового периода, можно предположить, что ареалы предков также охватывали гондванские континенты Южной Америки, Африки и Индо-Мадагаскара. [ 1 ]

Объединив свой палеобиогеографический анализ с филогениями, калиброванными по времени, основанными на оценках возраста окаменелостей, Поропат и его коллеги пришли к выводу, что сомфоспондилы и титанозавры достигли широкого распространения по всему миру к барремской эпохе, от 131 до 125 миллионов лет назад, если не раньше. Однако в конечном итоге они разделились на несколько линий с ограниченным географическим ареалом; они объяснили этот эндемизм региональными событиями вымирания . Они подсчитали, что линии саванназавра + диамантиназавра и винтонотитана по отдельности достигли Австралии не позднее поздней альбской эпохи, от 105 до 100 миллионов лет назад, но точное время зависело от моделей, которые они использовали. К этому интервалу относятся древнейшие австралийские титанозавроформы, австрозавр и «зауропод Хьюэндена». [ 1 ] [ 20 ]
Хотя они признали возможность систематической ошибки выборки , Поропат и его коллеги считали вероятным позднеальбское расселение из-за наиболее вероятного маршрута расселения, который должен был начаться из Южной Америки и пересечь Антарктиду в юго-восточную Австралию. В эпоху апта -раннего альба этот регион Австралии располагался на 70° южной широты с прохладным умеренным климатом. [ 21 ] В этих широтах и средах зауроподы не были обнаружены, что позволяет предположить, что они были адаптированы к теплому климату и избегали этих регионов. Таким образом, они, вероятно, не смогли бы расселиться по Антарктиде до позднего альба, когда глобальное потепление привело к более плавному градиенту температуры. [ 22 ] [ 23 ] Поропат и его коллеги предположили, что климатические предпочтения также могут объяснить сходство австралийских динозавров с Лавразией. [ 1 ]
В 2020 году Тай Кубо применил сетевой анализ к филогенетическому «супердереву» австралийских динозавров. Он обнаружил прочную связь между фауной австралийских динозавров и другими динозаврами Гондваны и определил гондванских динозавров мелового периода как отдельное сообщество. Таким образом, он предположил, что большинство предыдущих результатов, определяющих сходство между австралийскими и лавразийскими динозаврами, были вызваны недостаточной выборкой гондванских динозавров и филогенетической нестабильностью австралийских динозавров. Например, он отметил, что мегараптор Australovenator считался ответвлением азиатской линии, пока не было обнаружено больше гондванских мегарапторов. Хотя он предположил, что то же самое можно сказать и об этих зауроподах, он также признал возможность того, что у них была другая биогеографическая история, чем у более мелких динозавров. [ 24 ]
Палеобиология
[ редактировать ]
Как и все титанозавры, саванназавр, вероятно, был зауроподом «широкой колеи», а это означает, что он мог стоять и ходить, расставив ноги далеко от средней линии. [ 25 ] В 2020 году Поропат и его коллеги указали на ширину его грудинных пластин и крестца как на свидетельство широкой позиции. Крепкая плечевая кость саванназавра также считалась вероятной адаптацией к широкой стойке, которая была конвергентно приобретена у сальтазавринов. Они также предположили, что необычная форма таранной кости также была адаптацией к нагрузке, но отсутствие других костей задних конечностей делало это неясным. Тем не менее, Поропат и его коллеги также отметили черты, которые могли бы увеличить гибкость позвоночного столба, а именно отсутствие сочленений гипосфен-гипантр и наличие заметных экскаваций (центропрезигапофизарных ямок) под транспрезигапофизарными пластинками. Оба они были общими с диамантиназаврами , но в остальном были редки среди зауроподов. [ 1 ] [ 3 ]
Поропат и его коллеги отметили, что и саванназавр , и диамантиназавр жили в пойме с богатой глиной почвой и большим количеством осадков (см. § Палеоэкология ), что означало бы, что земля иногда становилась илистой и опасной для крупных зауроподов. Они считали возможным, что давление окружающей среды отобрало особей , способных передвигаться по илистой земле, и что саванназавр мог проводить больше времени возле воды, чем другие зауроподы. Гибкие позвонки (чтобы спасаться от трясины ), бочкообразные тела, похожие на тела гиппопотамов , широкая стойка и крепкие передние конечности (все для распределения веса тела) — все это было бы полезно для такой ниши . Они предположили, что сальтазаврины могли подвергаться аналогичному избирательному давлению, и поэтому «типичный» план тела титанозавра мог быть получен конвергентно несколько раз. Хотя в той же среде могли быть выбраны и более длинные шеи, чтобы обеспечить более легкий доступ к воде, Поропат и его коллеги отметили, что Саванназавр и диамантиназавр, вероятно, имели относительно короткую шею, хотя перекрытие шейных ребер могло улучшить стабильность у саванназавра . [ 3 ]
Палеосреда
[ редактировать ]
Овечья станция Бельмонт является частью содержащих окаменелости горных пород в верхней части формации Винтон , которая была датирована примерно или сразу после границы между сеноманской и туронской эпохами мелового периода - около 93,9 миллиона лет назад - на основе урановых месторождений. свинцовое датирование обломочных цирконов . [ 26 ] Это геологически самая молодая порода мезозойской эры в бассейне Эроманга . шесть основных фаций В формации Уинтон обнаружено , или отдельных типов пород: два разных типа комбинаций песчаника и алевролита (связанные с извилистыми реками, трещинами и дамбами ), аргиллиты (связанные с старицами и прудами), песчаные алевролиты и алевритовые аргиллиты (связанные с приливными отмелями ), богатые растительностью аргиллиты и уголь (связанные с болотистая местность), внутриформационные конгломераты (связанные с затоплением). Эти камни позволяют предположить, что саванназавр жил в пресноводной пойме, покрытой широкими извилистыми речными руслами с низкоэнергетическим потоком воды, которая время от времени подвергалась наводнениям. [ 27 ] [ 28 ]
В сеноманском и туронском периоде район Винтона находился на широте около 50° ю.ш. Окаменелости листьев показывают, что климат был теплым, со средней годовой температурой 16 ° C (61 ° F), и влажным, со средним годовым количеством осадков более 1300 миллиметров (51 дюйм). Хотя погода была сезонной, не было экстремального цикла между сезоном дождей и сезоном засухи , как сегодня. Вегетационный период длился восемь-девять месяцев. Однако погодные условия также оказались циклическими на многолетней основе, подобно современным тихоокеанским десятилетним колебаниям ; это также могло повлиять на спорадические наводнения. Хвойные и покрытосеменные растения были наиболее распространенными растениями формации Винтон; хвойные деревья включали кипарисовик Austrosequoia , араукарии Araucaria и Emwadea и подокарпий Protophyllocladoxylon , а покрытосеменные включали Lovellea и различные безымянные формы. Другие растения включали папоротники Phyllopteroides и Tempskya ; печеночник Marchantites ; хвощ Equisetites ; тот bennettitaleans Otozamites и Ptilophyllum ; и гинкгоальский Гинкго . [ 3 ] [ 28 ]

Саванназавр жил рядом с разнообразной фауной позвоночных в верхней части формации Винтон. [ 26 ] Среди современных динозавров были зауроподы Diamantinasaurus matildae и Wintonotitan watsi , теропод-мегараптор Australovenator wintonensis , а также неопределенные анкилозавры и гипсилофодонты . Зуб, принадлежащий австраловенатору, был найден рядом с голотипом саванназавра , что позволяет предположить, что его питание было причиной вычленения экземпляра. Среди других архозавров был птерозавр Ферродрако Лентони. [ 29 ] и крокодил Isisfordia duncani , а также другие неописанные птерозавры и крокодилы. Дополнительные позвоночные, известные из формации Винтон, - это -хелиды черепахи ; индетерминантная вараноидная ящерица; [ 30 ] рыба двоякодышащая Metaceratodus ellioti и M. wollastoni ; и лучепёрые рыбы, включая Cladocyclus geddesi . [ 31 ] Среди беспозвоночных преобладали двустворчатые моллюски Alathyria jaqueti , Hyridella goondiwindiensis , Megalovirgus wintonensis и Prohyria macmichaeli , но также были брюхоногие моллюски Melanoides , стрекозы , скорпионы и панцирные клещи, известные по следам окаменелостей, обнаруженных в окремненной древесине . [ 3 ] [ 28 ] [ 32 ]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р Поропат, Сан-Франциско; Мэннион, Вашингтон; Апчерч, П.; Хокнулл, ЮАР; Кир, BP; Кундрат, М.; Тишлер, ТР; Слоан, Т.; Синапий, GHK; Эллиотт, Дж.А.; Эллиотт, Д.А. (2016). «Новые австралийские зауроподы проливают свет на палеобиогеографию динозавров мелового периода» . Научные отчеты . 6 : 34467. Бибкод : 2016NatSR...634467P . дои : 10.1038/srep34467 . ПМК 5072287 . ПМИД 27763598 .
- ^ Перейти обратно: а б с Сен-Флер, Николас (20 октября 2016 г.). «Знакомьтесь, новый титанозавр. Вы можете называть его Уэйдом» . Нью-Йорк Таймс . Проверено 21 октября 2016 г.
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с Поропат, Сан-Франциско; Мэннион, Вашингтон; Апчерч, П.; Тишлер, ТР; Слоан, Т.; Синапий, GHK; Эллиотт, Дж.А.; Эллиотт, Д.А. (2020). «Остеология широкобедрого титанозаврового динозавра-зауропода Savannasaurus elliottorum из верхнемеловой формации Винтон в Квинсленде, Австралия» (PDF) . Журнал палеонтологии позвоночных . 40 (3): e1786836. дои : 10.1080/02724634.2020.1786836 . S2CID 224850234 .
- ^ Перейти обратно: а б с Геггель, Л. (20 октября 2016 г.). «Широкобедрый динозавр размером с автобус, однажды проезжавший по Австралии» . Живая наука . Покупка . Проверено 28 ноября 2020 г.
- ^ Д'Эмик, доктор медицины (2013). «Ранняя эволюция титанозаврообразных динозавров-зауроподов» . Зоологический журнал Линнеевского общества . 166 (3): 624–671. дои : 10.1111/j.1096-3642.2012.00853.x . hdl : 2027.42/94293 .
- ^ Перейти обратно: а б Мэннион, Вашингтон; Апчерч, П.; Барнс, Р.Н.; Матеус, О. (2013). «Остеология позднеюрского португальского динозавра-зауропода Lusotitan atalaiensis (Macronaria) и история эволюции базальных титанозавроформ» . Зоологический журнал Линнеевского общества . 168 : 98–206. дои : 10.1111/zoj.12029 .
- ^ Перейти обратно: а б с д Ройо-Торрес, Р.; Фуэнтес, К.; Мейхиде-Фуэнтес, Ф.; Мейхиде-Фуэнтес, М. (2017). «Новый динозавр Brachiosauridae Sauropod из нижнего мела Европы (провинция Сория, Испания)». Меловые исследования . 80 : 38–55. дои : 10.1016/j.cretres.2017.08.012 .
- ^ Мэннион, Вашингтон; Аллен, Р.; Мойн, О. (2017). «Самый ранний известный титанозавроподобный динозавр-зауропод и эволюция брахиозаврид» . ПерДж . 5 : е3217. дои : 10.7717/peerj.3217 . ПМК 5417094 . ПМИД 28480136 .
- ^ Аверьянов А.; Ефимов, В. (2018). «Старейший титанозавр-зауропод Северного полушария» . Биологические коммуникации . 63 (6): 145–162. дои : 10.21638/spbu03.2018.301 . hdl : 11701/15099 .
- ^ Гонсалес Рига, BJ; Мэннион, Вашингтон; Поропат, Сан-Франциско; Ортис Дэвид, Л.; Кориа, Япония (2018). «Остеология позднемелового аргентинского динозавра-зауропода Mendozasaurus neguyelap : значение для базальных взаимоотношений с титанозаврами». Зоологический журнал Линнеевского общества . 184 (1): 136–181. doi : 10.1093/zoolinnean/zlx103 . hdl : 10044/1/53967 .
- ^ Мочо, П.; Перес-Гарсия, А.; Мартин Хименес, М.; Ортега, Ф. (2019). «Новые останки из испанского сеномана проливают свет на гондванское происхождение европейских титанозавров раннего мела». Меловые исследования . 95 : 164–190. дои : 10.1016/j.cretres.2018.09.016 .
- ^ МОЧО, П.; Ройо-Торрес, Р.; Ортега, Ф. (2019). «Новый макронарий зауропод из поздней юры Португалии». Журнал палеонтологии позвоночных . 39 (1): e1578782. дои : 10.1080/02724634.2019.1578782 . S2CID 182239988 .
- ^ Гонсалес Рига, BJ; Ламанна, MC; Отеро, А.; Ортис Дэвид, LD; Келлнер, AWA; Ибирику, LM (2019). «Обзор аппендикулярной анатомии скелета южноамериканских зауроподов-титанозавров с определением недавно признанной клады» . Анаис да Академия Бразилиа де Сиенсиас . 91 (Приложение 2): e20180374. дои : 10.1590/0001-3765201920180374 . hdl : 11336/106658 . ПМИД 31340217 .
- ^ Сильва, JCG младший; Мариньо, ТС; Мартинелли, АГ; Лангер, MC (2019). «Остеология и систематика Uberabatitan Ribeiroi (Dinosauria; Sauropoda): позднемелового титанозавра из Минас-Жерайс, Бразилия». Зоотакса . 4577 (3): 401–438. дои : 10.11646/zootaxa.4577.3.1 . ПМИД 31715707 .
- ^ Мэннион, Вашингтон; Апчерч, П.; Джин, X.; Чжэн, В. (2019). «Новая информация о меловых динозаврах-зауроподах провинции Чжэцзян, Китай: влияние на лавразийскую филогению и биогеографию титанозаврообразных» . Королевское общество открытой науки . 6 (8): 191057. Бибкод : 2019RSOS....691057M . дои : 10.1098/rsos.191057 . ПМК 6731702 . ПМИД 31598266 .
- ^ Фанти, Ф. (2012). «Меловые континентальные мосты, изолированность и викариантность в южном полушарии: какой путь выбрали динозавры?». В Таланте, Дж. А. (ред.). Земля и Жизнь . Международный год планеты Земля. Дордрехт: Спрингер. стр. 883–911. дои : 10.1007/978-90-481-3428-1_31 . ISBN 978-90-481-3427-4 .
- ^ Мольнар, Р.Э. (1992). «Палеозоогеографические взаимоотношения австралийских мезозойских четвероногих». В Чаттерджи, С.; Хоттон, Н. (ред.). Новые концепции в глобальной тектонике . Лаббок: Издательство Техасского технологического университета. стр. 259–266.
- ^ Барретт, премьер-министр; Бенсон, РБЖ; Рич, TH; Викерс-Рич, П. (2011). «Первый динозавр-спинозавр из Австралии и космополитизм меловой фауны динозавров» . Письма по биологии . 7 (6): 933–936. дои : 10.1098/rsbl.2011.0466 . ПМК 3210678 . ПМИД 21693488 .
- ^ Бенсон, РБЖ; Рич, TH; Викерс-Рич, П.; Холл, М. (2012). «Фауна теропод из Южной Австралии указывает на высокое полярное разнообразие и обусловленную климатом провинциальность динозавров» . ПЛОС ОДИН . 7 (5): e37122. Бибкод : 2012PLoSO...737122B . дои : 10.1371/journal.pone.0037122 . ПМЦ 3353904 . ПМИД 22615916 .
- ^ Поропат, Сан-Франциско; Наир, Япония; Сайм, CE; Мэннион, Вашингтон; Апчерч, П.; Хокнулл, ЮАР; Кук, АГ; Тишлер, ТР; Холланд, Т. (2017). «Переоценка Austrosaurus mckillopi Longman, 1933 из аргиллита Аллару в Квинсленде, первого в Австралии названного динозавра-зауропода мелового периода». Алчеринга . 41 (4): 543–580. дои : 10.1080/03115518.2017.1334826 . hdl : 10044/1/48659 . S2CID 134237391 .
- ^ Деттманн, Мэн; Мольнар, Р.Э.; Дуглас, Дж. Г. (1992). «Австралийские наземные фауны и флоры мелового периода: биостратиграфические и биогеографические значения». Меловые исследования . 13 (3): 207–262. дои : 10.1016/0195-6671(92)90001-7 .
- ^ Хасэгава, Х.; Тада, Р.; Цзян, X.; Суганума, Ю.; Имсамут, С.; Чарусири, П.; Ичинноров Н.; Кханд, Ю. (2012). «Резкое сокращение циркуляции Хэдли во время супероранжереи середины мелового периода» . Климат прошлого . 8 (4): 1323–1337. Бибкод : 2012CliPa...8.1323H . дои : 10.5194/cp-8-1323-2012 .
- ^ Уфнар, DF; Гонсалес, Луизиана; Людвигсон, Джорджия; Бреннер, РЛ; Витцке, Би Джей (2004). «Доказательства увеличения переноса скрытого тепла во время парникового потепления мелового (альбского) периода». Геология . 32 (12): 1049–1052. Бибкод : 2004Geo....32.1049U . дои : 10.1130/G20828.1 .
- ^ Кубо, Т. (2020). «Биогеографический сетевой анализ австралийских динозавров мелового периода». Исследования Гондваны . 82 : 39–47. Бибкод : 2020GondR..82...39K . дои : 10.1016/j.gr.2019.12.012 .
- ^ Уилсон, Дж.А.; Каррано, Монтана (1999). «Титанозавры и происхождение троп «широкой колеи»: биомеханический и систематический взгляд на передвижение зауроподов». Палеобиология . 25 (2): 252–267. дои : 10.1017/S0094837300026543 . JSTOR 2666093 .
- ^ Перейти обратно: а б Такер, RT; Робертс, EM; Ху, Ю.; Кемп, AIS; Солсбери, Юго-Запад (2013 г.). «Ограничения по возрасту обломочных цирконов формации Винтон, Квинсленд: контекстуализация фауны динозавров позднего мела в Австралии». Исследования Гондваны . 24 (2): 767–779. Бибкод : 2013GondR..24..767T . дои : 10.1016/j.gr.2012.12.009 .
- ^ Такер, RT; Робертс, EM; Дарлингтон, В.; Солсбери, Юго-Запад (2017). «Исследование стратиграфии и палеосреды ряда недавно обнаруженных местонахождений окаменелостей позвоночных среднего мела в формации Винтон, Квинсленд, Австралия» (PDF) . Осадочная геология . 358 : 210–229. Бибкод : 2017SedG..358..210T . дои : 10.1016/j.sedgeo.2017.05.004 .
- ^ Перейти обратно: а б с Флетчер, ТЛ; Мосс, ПТ; Солсбери, Юго-Запад (2018 г.). «Палеосреда верхнемеловой (сеноман-туронской) части формации Винтон, Квинсленд, Австралия» . ПерДж . 6 : е5513. дои : 10.7717/peerj.5513 . ПМК 6130253 . ПМИД 30210941 .
- ^ Пентленд, штат Айдахо; Поропат, Сан-Франциско; Тишлер, ТР; Слоан, Т.; Эллиотт, РА; Эллиотт, штат Ха; Эллиотт, Дж.А.; Эллиотт, Д.А. (2019). « Ferrodraco lentoni gen. et sp. nov., новый птерозавр-орнитохейрид из формации Винтон (сеноман – нижний турон) в Квинсленде, Австралия» . Научные отчеты . 9 (1): 13454. Бибкод : 2019НатСР...913454П . дои : 10.1038/s41598-019-49789-4 . ПМК 6776501 . ПМИД 31582757 .
- ^ Кир, BP (2016). «Меловые морские амниоты Австралии: перспективы десятилетия новых исследований» . Мемуары музея Виктории . 74 : 17–28. дои : 10.24199/j.mmv.2016.74.03 .
- ^ Беррелл, RW; Альварадо-Ортега, Дж.; Ябумото, Ю.; Солсбери, Юго-Запад (2014 г.). «Первая запись ихтиодектиформной рыбы Cladocyclus из восточной Гондваны: новый вид из нижнего мела Квинсленда, Австралия» . Acta Palaeontologica Polonica . 59 (4): 903–920. дои : 10.4202/app.2012.0019 .
- ^ Хокнулл, ЮАР; Уайт, Массачусетс; Тишлер, ТР; Кук, АГ; Каллеха, Северная Дакота; Слоан, Т.; Эллиотт, Д.А. (2009). «Новые среднемеловые (последние альбы) динозавры из Винтона, Квинсленд, Австралия» . ПЛОС ОДИН . 4 (7): е6190. Бибкод : 2009PLoSO...4.6190H . дои : 10.1371/journal.pone.0006190 . ПМК 2703565 . ПМИД 19584929 .











