Архозавр
| Архозавры | ||
|---|---|---|
 | ||
| Птицы и крокодилы (в данном случае большая цапля и крокодил-грабитель ) — единственные живые группы архозавров. | ||
| Научная классификация | ||
| Домен: | Эукариоты | |
| Королевство: | животное | |
| Тип: | Хордовые | |
| Сорт: | Рептилии | |
| Клэйд : | Архозавроморфа | |
| Клэйд : | Архозавриформы | |
| Клэйд : | Евкрокопода | |
| Клэйд : | Архозаврия Коуп , 1869 г. | |
| Подгруппы | ||
| ||
| Синонимы | ||
| ||
Архозавры ( букв. « правящие рептилии » ) или архозавры ( / ˈ ɑːr k ə ˌ s ɔːr / [3] ) — клада диапсидных представителями зауропсидов четвероногих , являются птицы и крокодилы единственными дошедшими до нас . Хотя их широко классифицируют как рептилий , которые традиционно исключают птиц, кладистический смысл этого термина включает всех ныне живущих и вымерших родственников птиц и крокодилов, таких как нептичьи динозавры , птерозавры , фитозавры , этоозавры и раузухи, а также многие мезозойские морские рептилии . Современные палеонтологи определяют архозаврию как кроновую группу , включающую самого недавнего общего предка ныне живущих птиц и крокодилов, а также всех его потомков.
Основа Archosauria распадается на две клады: Pseudosuria , включающую крокодилов и их вымерших родственников; и Avemetatarsalia , куда входят птицы и их вымершие родственники (такие как нептичьи динозавры и птерозавры). [4] Более старые определения группы Archosauria основаны на общих морфологических характеристиках, таких как предглазничное отверстие в черепе, зазубренные зубы и вертикальное положение. Некоторые вымершие рептилии, такие как протерозухиды и эупаркериииды , также обладали этими особенностями, но возникли до разделения линий крокодилов и птиц. Старое морфологическое определение Archosauria в настоящее время примерно соответствует Archosauriformes , группе, названной так, чтобы объединить архозавров коронной группы и их близких родственников. [4]
Самые старые настоящие окаменелости архозавров известны из раннего триасового периода, хотя первые архозавроформы и архозавроморфы (рептилии, более близкие к архозаврам, чем к ящерицам или другим лепидозаврам ) появились в перми . Архозавры быстро диверсифицировались после пермско-триасового массового вымирания (~ 252 млн лет назад ), которое уничтожило большинство доминирующих тогда конкурентов терапсидов, таких как горгонопсии и аномодонты , а последующий засушливый триасовый климат позволил к засухе. архозаврам стать более устойчивыми (во многом благодаря их мочевой кислоте , основанной на мочевой системе ), которые в конечном итоге стали крупнейшими и наиболее экологически доминирующими наземными позвоночными от среднего триаса до мел-палеогенового вымирания (~ 66 млн лет назад). [5] Птицы и несколько крокодилообразных линий были единственными архозаврами, пережившими вымирание K-Pg , которые вновь диверсифицировались в последующую кайнозойскую эпоху. В частности, в наши дни птицы стали одной из наиболее богатых видами групп наземных позвоночных.
Отличительные характеристики
[ редактировать ]Архозавров традиционно можно отличить от других четвероногих на основе нескольких синапоморфий или общих характеристик, которые присутствовали у их последнего общего предка . Многие из этих характеристик появились до возникновения клады Archosauria, поскольку они присутствовали у архозаврообразных, таких как Proterosuchus и Euparkeria , которые находились за пределами коронной группы . [4]
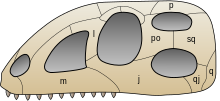
Наиболее очевидные особенности включают зубы, посаженные в глубокие лунки, анторбитальные и нижнечелюстные отверстия (отверстия перед глазами и в челюсти соответственно), [6] и выраженный четвертый вертел (выступающий гребень на бедренной кости ). [7] Будучи вставленными в лунки, зубы с меньшей вероятностью вырывались во время кормления. Эта особенность отвечает за название « текодонт » (что означает «зубы в лунке»). [8] которое ранние палеонтологи применили ко многим архозаврам триаса. [7] Кроме того, немышечная ткань щек и губ встречается в различных формах по всей кладе, при этом у всех современных архозавров отсутствуют немышечные губы, в отличие от большинства нептичьих ящеротазовых динозавров. [9] Некоторые архозавры, например птицы, вторично беззубы. Анторбитальные отверстия уменьшали вес черепа, который у ранних архозавров был относительно большим, как и у современных крокодилов . Нижнечелюстные окна в некоторых формах также могли уменьшить вес челюсти. Четвертый вертел обеспечивает большое место прикрепления мышц бедренной кости. Более сильные мышцы позволяли ранним архозаврам ходить прямо, а также могли быть связаны со способностью архозавров или их непосредственных предков пережить катастрофическое пермско-триасовое вымирание . [ нужна ссылка ]
В отличие от своих ныне живущих близких родственников лепидозавров, архозавры лишились сошниково-носового органа . [10]
Происхождение
[ редактировать ]Архозавры — подгруппа архозавроформ , которые сами являются подгруппой архозавроморфов . И древнейший архозавроморф ( Protorosaurus speneri ), и древнейший архозаврообразный ( Archosaurus rossicus ) жили в поздней перми. Древнейшие настоящие архозавры появились на оленекском этапе (247–251 млн лет назад) раннего триаса . несколько фрагментарных окаменелостей крупных хищных архозавров крокодиловой линии (неофициально называемых « рауизухами На этом этапе известно »). К ним относятся Scythoзарух и Цилмозух (оба обнаружены в России ), [11] а также Xilousuchus , гребневик из Китая . [4] Самые старые известные окаменелости архозавров птичьего типа относятся к анизийскому этапу (247–242 млн лет назад) Танзании и включают Асилизавра (раннего силезаврида ), Телеократера ( афанозавра ) и Ньясазавра (возможно, раннего динозавра). [ нужна ссылка ]
Доминирование архозавров в триасе
[ редактировать ]Синапсиды — клада, включающая млекопитающих и их вымерших предков . Последнюю группу часто называют млекопитающими рептилиями, но ее следует называть протомлекопитающими, стволовыми млекопитающими или базальными синапсидами, они не являются настоящими рептилиями поскольку по современной кладистической классификации . Они были доминирующими наземными позвоночными на протяжении всей перми , но большинство из них погибло во время пермско-триасового вымирания . Очень немногие крупные синапсиды пережили это событие, но одна форма, Lystrosaurus ( травоядный дицинодонт ), получила широкое распространение вскоре после вымирания. [12] После этого архозавры и другие архозавроформы быстро стали доминирующими наземными позвоночными в раннем триасе . Окаменелости, существовавшие до массового вымирания, были найдены только вокруг экватора, но после этого события окаменелости можно найти по всему миру. [13] Предлагаемые объяснения этого включают:
- Архозавры добились более быстрого прогресса в выпрямлении конечностей, чем синапсиды, и это дало им большую выносливость, поскольку они избегали ограничений Кэрриера . Возражением против этого объяснения является то, что архозавры стали доминировать, хотя у них все еще были раскидистые или полупрямые конечности, как у листрозавров и других синапсидов. [ нужна ссылка ]
- Архозавры обладают более эффективными [ нужны разъяснения ] дыхательные системы с однонаправленным потоком воздуха, в отличие от приливного дыхания синапидов. [14] Способность более эффективно дышать в условиях гипоксии могла быть полезна ранним архозаврам во время предполагаемого падения уровня кислорода в конце перми. [14]
- Ранний триас был преимущественно засушливым, поскольку большая часть суши Земли была сосредоточена на суперконтиненте Пангея . Архозавры, вероятно, лучше сохраняли воду, чем ранние синапсиды, потому что:
- Современные диапсиды (ящерицы, змеи, крокодилы, птицы) выделяют мочевую кислоту , которая может выделяться в виде пасты, что приводит к низкой потере воды в отличие от более разбавленной мочи. Разумно предположить, что архозавры (предки крокодилов, динозавров и птерозавров) также выделяли мочевую кислоту и поэтому хорошо сохраняли воду. Железистые (безжелезистые) шкурки диапсид также способствовали сохранению воды. [ нужна ссылка ]
- Современные млекопитающие выделяют мочевину , которая требует относительно высокой частоты мочеиспускания, чтобы не допустить ее выхода с мочой путем диффузии в почечные канальцы. Их шкура также содержит множество желез, которые также теряют воду. Если предположить, что ранние синапсиды обладали сходными особенностями, как, например, утверждают авторы « Палеоса» , они оказались в невыгодном положении в преимущественно засушливом мире. Тот же уважаемый сайт указывает, что «на протяжении большей части плио-плейстоценовой истории Австралии, где условия, вероятно, были схожими, крупнейшими наземными хищниками были не млекопитающие, а гигантские вараниды ( Мегалания ) и наземные крокодилы». [8]
Однако эта теория была подвергнута сомнению, поскольку она подразумевает, что синапсиды обязательно имели меньшие преимущества в удержании воды, что сокращение количества синапсид совпадает с изменениями климата или разнообразием архозавров (ни то, ни другое не проверялось), а также тот факт, что млекопитающие, обитающие в пустынях, также хорошо адаптированы в этом отделе. как архозавры, [15] а некоторые цинодонты, такие как Trucidocynodon, были хищниками крупных размеров. [16] Исследование предполагает, что конкуренция среди млекопитающих является основным объяснением небольшого размера мезозойских млекопитающих. [17]
Основные формы
[ редактировать ]

С 1970-х годов ученые классифицировали архозавров главным образом на основании их лодыжек. [18] У самых ранних архозавров были «примитивные мезотарзальные» лодыжки: таранная кость и пяточная кость были прикреплены к большеберцовой и малоберцовой , костям швами а сустав сгибался вокруг контакта между этими костями и стопой.
Псевдозухии начале появились в триаса . В лодыжках таранная кость была соединена с большеберцовой костью с помощью шва , и сустав вращался вокруг штифта на таранной кости, который входил в гнездо в пяточной кости. Ранние «круротарсаны» все еще ходили с раскидистыми конечностями, но у некоторых более поздних круротарсанов конечности развились полностью прямо. Современные крокодилы — это круротарсаны, которые могут использовать самые разные походки в зависимости от скорости. [19]
У Euparkeria и Ornithosuchidae были «перевернутые голеностопные» лодыжки с выступом на пяточной кости и гнездом на таранной кости.
Самые ранние окаменелости Avemetatarsalia («птичьи лодыжки») появляются в анизийском возрасте среднего триаса . У большинства орнитодиров были «продвинутые мезотарзальные» лодыжки. Эта форма лодыжки включала в себя очень большую таранную кость и очень маленькую пяточную кость и могла двигаться только в одной плоскости, как простой шарнир. Такое расположение, подходившее только для животных с прямыми конечностями, обеспечивало большую устойчивость при беге. Самые ранние авеметатарзалии, такие как Teleocrater и Asilisaurus, сохранили «примитивные мезотарзальные» лодыжки. Орнитодиры отличались от других архозавров и в других отношениях: они были легкого телосложения и обычно небольшого размера, их шеи были длинными и имели S-образный изгиб, их черепа были гораздо более легковесными, а многие орнитодиры были полностью двуногими . Четвертый вертел архозавров на бедре, возможно, облегчил орнитодиранам возможность стать двуногими, поскольку он обеспечивал больше рычагов для мышц бедра. В позднем триасе орнитодиры диверсифицировались и стали производить динозавры и птерозавры .
Классификация
[ редактировать ]Современная классификация
[ редактировать ]Архозаврию обычно определяют как кронную группу , что означает, что она включает только потомков последних общих предков ее ныне живущих представителей. В случае архозавров это птицы и крокодилы. Архозаврия входит в более крупную кладу Archosauriformes , в которую входят некоторые близкие родственники архозавров, такие как протерохампсиды и эупаркерииды . Этих родственников часто называют архозаврами, несмотря на то, что они находятся за пределами коронной группы Archosauria и занимают более базальное положение внутри Archosauriformes. [20] Исторически многие архозавроформы были описаны как архозавры, в том числе протерозухиды и эритрозухиды , на основании наличия анторбитального отверстия. Хотя многие исследователи предпочитают рассматривать Archosauria как кладу без ранга , некоторые продолжают присваивать ей традиционный биологический ранг. Традиционно Archosauria рассматривалась как надотряд, хотя некоторые исследователи 21 века отнесли ее к разным рангам, включая отдел. [21] и Класс. [22]
История классификации
[ редактировать ]Архозаврия как термин была впервые введена американским палеонтологом Эдвардом Дринкером Коупом в 1869 году и включала широкий спектр таксонов, включая динозавров , крокодилов , текодонтов , зауроптеригов (которые могут быть связаны с черепахами), ринхоцефалов (группа, в которую, по мнению Коупа, входили ринхозавры) . , которые в наше время считаются более базальными архозавроморфами , и туатары , которые являются лепидозаврами ), и аномодонты , которые сейчас считаются синапсидами. [23] Лишь в 1986 году Archosauria была определена как коронная клада, ограничив ее использование более производными таксонами. [24]
большеберцовая кость Фибула Астрагал каблук


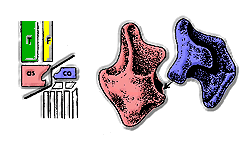
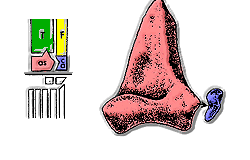
Термин Коупа представлял собой греко-латинский гибрид, предназначенный для обозначения черепных дуг, но позже его также понимали как «ведущие рептилии» или «правящие рептилии» из-за ассоциации с греческим ἀρχός «лидер, правитель». [25]
Термин «текодонт», ныне считающийся устаревшим, впервые был использован английским палеонтологом Ричардом Оуэном в 1859 году для описания триасовых архозавров и стал широко использоваться в 20 веке. Текодонты считались «базальной группой», от которой произошли более продвинутые архозавры. Они не обладали чертами, присущими более поздним линиям птиц и крокодилов, и поэтому считались более примитивными и предками этих двух групп. После кладистической революции 1980-х и 90-х годов, когда кладистика стала наиболее широко используемым методом классификации организмов, текодонты больше не считались действительной группировкой. Поскольку они считаются «базальной группой», текодонты являются парафилетическими , то есть они образуют группу, которая не включает всех потомков своего последнего общего предка: в этом случае более производные крокодилы и птицы исключаются из «Thecodontia», поскольку они раньше было понятно. Описание базальных орнитодиров Lagerpeton и Lagosurus в 1970-х годах предоставило доказательства, связывающие текодонтов с динозаврами, и способствовало отказу от использования термина «Thecodontia», который многие кладисты считают искусственной группировкой. [26]
После идентификации «крокодиловых нормальных» и «крокодиловых перевернутых» лодыжек Санкаром Чаттерджи в 1978 году было обнаружено базальное расщепление у Archosauria. Чаттерджи считал, что эти две группы — это Pseudosuria с «нормальной» лодыжкой и Ornithosuridae с «перевернутой» лодыжкой. В то время считалось, что орнитозухиды были предками динозавров. В 1979 году ARI Cruickshank определил базальный раскол и предположил, что круротарсановая лодыжка развивалась независимо в этих двух группах, но противоположным образом. Круикшенк также считал, что развитие этих типов лодыжек прогрессировало в каждой группе, что позволило продвинутым членам иметь полупрямую (в случае крокодилов) или прямую (в случае динозавров) походку. [26]
Филогения
[ редактировать ]Во многих филогенетических анализах было показано, что архозавры представляют собой монофилетическую группу, образуя таким образом настоящую кладу. Одно из первых исследований филогении архозавров было проведено французским палеонтологом Жаком Готье в 1986 году. Готье разделил архозавров на псевдозухий , линию крокодилов, и орнитозухий , линию динозавров и птерозавров. Псевдозухия определялась как все архозавры, более тесно связанные с крокодилами, а орнитозухия определялась как все архозавры, более тесно связанные с птицами. Протерохампсиды, эритрозухиды и протерозухиды последовательно выпали за пределы архозаврии в полученном дереве. Ниже представлена кладограмма Готье (1986): [27]
| Зауропсида |
| ||||||||||||||||||
В 1988 году палеонтологи Майкл Бентон и Дж. М. Кларк создали новое дерево в ходе филогенетического исследования базальных архозавров. Как и в случае с деревом Готье, Бентон и Кларк выявили базальное расщепление внутри архозаврии. Они назвали эти две группы Crocodylotarsi и Ornithosuria. Crocodylotarsi был определен как таксон, основанный на апоморфии, основанный на наличии «нормального для крокодила» голеностопного сустава (который считается определяющей апоморфией клады). Pseudosuria Готье, напротив, был таксоном, основанным на стебле . В отличие от дерева Готье, Бентон и Кларк помещают Euparkeria вне орнитозухий и вообще за пределы кроновой группы Archosauria. [28]
Клады Crurotarsi и Ornithodira были впервые использованы вместе в 1990 году палеонтологами Полом Серено и А.Б. Аркуччи в их филогенетическом исследовании архозавров. Они были первыми, кто создал кладу Crurotarsi, а Ornithodira была названа Готье в 1986 году. Crurotarsi и Ornithodira заменили Pseudosuria и Ornithosuria соответственно, поскольку монофилия обеих этих клад была поставлена под сомнение. [26] [29] Серено и Аркуччи включили в свой анализ особенности архозавров, отличные от типов лодыжек, в результате чего получилось другое дерево, чем в предыдущих анализах. Ниже приведена кладограмма, основанная на Серено (1991), которая аналогична той, что была создана Серено и Аркуччи: [26]
| Архозавриформы |
| ||||||||||||||||||||||||
Ornithodira и Crurotarsi представляют собой клады, основанные на узлах , что означает, что они определяются как включающие последнего общего предка двух или более таксонов и всех его потомков. Орнитодира включает последнего общего предка птерозавров и динозавров (в том числе птиц), тогда как Круротарси включает последнего общего предка современных крокодилов и трех групп триасовых архозавров: орнитозухид , этозавров и фитозавров . Эти клады не эквивалентны архозаврам «птичьей линии» и «крокодиловой линии», которые представляют собой ветвистые клады, определяемые как все таксоны, более тесно связанные с одной живой группой (птицами или крокодилами), чем с другой.
Бентон предложил название Avemetatarsalia в 1999 году, чтобы включить всех архозавров птичьего типа (по его определению, все архозавры более тесно связаны с динозаврами, чем с крокодилами). Его анализ небольшого триасового архозавра Scleromochlus поместил его в число архозавров птичьего типа, но за пределами Орнитодиры, а это означает, что Орнитодира больше не была эквивалентом архозавров птичьего типа. Ниже представлена модифицированная кладограмма Бентона (2004), показывающая эту филогению: [24]
| Архозаврия |
| ||||||||||||||||||||||||||||||
В монографии Стерлинга Несбитта о ранних архозаврах 2011 года филогенетический анализ обнаружил убедительную поддержку того, что фитозавры не относятся к архозавриям. Многие последующие исследования подтвердили эту филогению. Поскольку Crurotarsi определяется включением фитозавров, размещение фитозавров за пределами Archosauria означает, что Crurotarsi должен включать всю Archosauria. Несбитт восстановил Pseudosuria в качестве названия клады архозавров крокодиловой линии, используя его в качестве таксона на основе стебля. Ниже представлена модифицированная кладограмма Несбитта (2011): [4]
| Зауропсида |
| ||||||
Вымирание и выживание
[ редактировать ]Крокодиломорфы, птерозавры и динозавры пережили триасовое и юрское вымирание около 200 миллионов лет назад, но другие архозавры вымерли на границе триаса и юрского периода или раньше.
Нептичьи динозавры и птерозавры погибли в результате мел-палеогенового вымирания , которое произошло примерно 66 миллионов лет назад, но птицы кроновой группы (единственная оставшаяся группа динозавров) и многие крокодилиформы выжили. Оба являются потомками архозавров и, следовательно, сами являются архозаврами согласно филогенетической таксономии .
Крокодилы (к которым относятся все современные крокодилы , аллигаторы и гавиалы ) и птицы процветают сегодня в голоцене . Принято считать, что среди всех наземных позвоночных птиц больше всего видов. [ нужна ссылка ]
Образ жизни архозавров
[ редактировать ]Тазобедренные суставы и локомоция
[ редактировать ]
Как и ранние четвероногие , ранние архозавры имели раскидистую походку, потому что их тазобедренные суставы были обращены в сторону, а выступы на вершинах их бедренных костей находились на одной линии с бедренной костью.В раннем и среднем триасе у некоторых групп архозавров развились тазобедренные суставы, которые позволяли (или требовали) более прямую походку. Это давало им большую выносливость, поскольку позволяло избежать ограничений Кэрриера , то есть они могли одновременно бегать и легко дышать. Существовало два основных типа суставов, позволяющих выпрямлять ноги:
- Тазобедренные суставы были обращены в сторону, но выступы на бедренных костях располагались под прямым углом к остальной части бедренной кости, поэтому они были направлены вниз. Динозавры произошли от архозавров с таким расположением бедер.
- Тазобедренные суставы были направлены вниз, а выступы на бедренных костях находились на одной линии с бедренной костью. Это «столбообразное» расположение, по-видимому, развилось независимо в различных линиях архозавров, например, оно было распространено у «Rauisuria» (некрокодиломорфных паракрокодиломорфов ), а также появилось у некоторых этозавров .
Было отмечено, что вертикальное положение требует больше энергии, поэтому может указывать на более высокий обмен веществ и более высокую температуру тела. [30]
Диета
[ редактировать ]Большинство из них были крупными хищниками, но представители различных линий заняли другие ниши. Этозавры были травоядными, и у некоторых из них был развит обширный панцирь. Некоторые крокодилиформные были травоядными, например, Simosurus , Phyllodontosurus . Крупный крокодилообразный Stomatosuchus , возможно, был фильтратором . Зауроподоморфы и птицетазовые динозавры были травоядными животными с разнообразными биомеханическими приспособлениями к питанию .
Земля, вода и воздух
[ редактировать ]Архозавров в основном изображают как наземных животных, но:
- Многие фитозавры и крокодилообразные доминировали в реках и болотах и даже вторглись в моря (например, телеозавры , Metriorhynchidae и Dyrosauridae ) . Metriorhynchidae были скорее похожи на дельфинов, с веслообразными передними конечностями, хвостовым плавником и гладкой небронированной кожей.
- Две клады орнитодиров, птерозавры и птицы, доминировали в воздухе после того, как приспособились к летательному образу жизни.
- некоторые динозавры, такие как спинозавр, Утверждалось, что гесперорниты и пингвины . вели полуводный образ жизни. К такому образу жизни приспособились также
Метаболизм
[ редактировать ]Метаболизм архозавров до сих пор остается спорной темой. Они, безусловно, произошли от хладнокровных предков, а выжившие архозавры, не относящиеся к динозаврам, крокодилы, являются хладнокровными. Но у крокодилов есть некоторые особенности, которые обычно связаны с теплокровным обменом веществ, поскольку они улучшают снабжение животных кислородом:
- 4-камерное сердце. И птицы, и млекопитающие имеют 4-камерное сердце, в котором полностью разделены потоки насыщенной кислородом и лишенной кислорода крови . У рептилий, не являющихся крокодилами, трехкамерное сердце , которое менее эффективно, поскольку позволяет двум потокам смешиваться и, таким образом, отправлять некоторое количество лишенной кислорода крови в тело, а не в легкие. Сердце современных крокодилов четырехкамерное, но меньше по размеру по сравнению с размером тела и работает при более низком давлении, чем у современных птиц и млекопитающих . У них также есть шунтирование легких , что делает их функционально трехкамерными под водой, сохраняя кислород .
- вторичное небо , позволяющее животному есть и дышать одновременно.
- механизм печеночно-поршневой для перекачивания легких . Это отличается от механизмов перекачки легких млекопитающих и птиц, но похоже на то, что некоторые исследователи обнаружили у некоторых динозавров. [31] [32]
Исторически существовала неопределенность относительно того, почему естественный отбор благоприятствовал развитию этих особенностей, которые очень важны для активных теплокровных существ, но малопригодны для хладнокровных водных хищников, устраивающих засады , которые проводят подавляющую часть своего времени, плавая в воде. воды или лежать на берегах рек.


Палеонтологические свидетельства [ нужны разъяснения ] показывает, что предки современных крокодилов были активными и эндотермными (теплокровными). Некоторые эксперты [ ВОЗ? ] полагают, что их предки-архозавры также были теплокровными. Вероятно, это связано с тем, что в ходе эволюции пероподобные нити покрыли все тело и стали способны обеспечивать теплоизоляцию. [33] Физиологические, анатомические особенности и особенности развития сердца крокодилов подтверждают палеонтологические данные и показывают, что эта линия вернулась к эктотермии, когда она вторглась в нишу водных хищников, устраивающих засады. У эмбрионов крокодилов на ранней стадии развивается полностью четырехкамерное сердце. Модификации растущего сердца образуют легочный шунт, который включает левую дугу аорты , которая начинается из правого желудочка , отверстие Паницца между левой и правой дугами аорты и зубчатый клапан у основания легочной артерии . Шунт используется во время ныряния, чтобы сердце функционировало как трехкамерное, обеспечивая крокодилам нервно-контролируемое шунтирование, используемое эктотермными животными. Исследователи пришли к выводу, что предки современных крокодилов имели полностью четырехкамерное сердце и, следовательно, были теплокровными, прежде чем они вернулись к холоднокровному или экзотермическому метаболизму. Авторы также приводят другие доказательства эндотермии у стволовых архозавров. [34] [35] Разумно предположить, что позже у крокодилов развился легочный шунт, поскольку они стали хладнокровными, водными и менее активными.
Если бы предки крокодилов и другие архозавры триаса были теплокровными, это помогло бы решить некоторые эволюционные загадки:
- Самые ранние крокодиломорфы, например Terrestrisuchus , были стройными, длинноногими наземными хищниками, телосложение которых предполагает достаточно активный образ жизни, требующий довольно быстрого метаболизма. И некоторые другие архозавры-круротарсаны, по-видимому, имели прямые конечности, в то время как конечности райзухий очень плохо приспособлены к любой другой позе. Прямые конечности выгодны для активных животных, поскольку позволяют избежать ограничений Кэрриера , но невыгодны для более медлительных животных, поскольку увеличивают затраты энергии на вставание и лежание.
- Если бы ранние архозавры были полностью хладнокровными, а (что наиболее вероятно) динозавры были, по крайней мере, достаточно теплокровными , динозаврам пришлось бы развить теплокровный метаболизм менее чем в два раза быстрее, чем синапсидам , чтобы сделать то же самое.
Дыхательная система
[ редактировать ]Недавнее исследование легких Alligator Mississippiensis ( Американский аллигатор ) показало, что поток воздуха через них однонаправленный, двигаясь в одном и том же направлении при вдохе и выдохе. [36] Это также наблюдается у птиц и многих нептичьих динозавров, у которых есть воздушные мешки , способствующие дыханию. И птицы, и аллигаторы достигают однонаправленного потока воздуха благодаря наличию парабронхов , которые отвечают за газообмен . Исследование показало, что у аллигаторов воздух входит через вторую бронхиальную ветвь , движется через парабронхи и выходит через первую бронхиальную ветвь. Однонаправленный поток воздуха как у птиц, так и у аллигаторов позволяет предположить, что этот тип дыхания присутствовал у основания архозаврий и сохранялся как у динозавров, так и у нединозавровых архозавров, таких как этоозавры, «рауизухи» (некрокодиломорфные паракрокодиломорфы), крокодиломорфы и птерозавры. [36] Использование однонаправленного воздушного потока в легких архозавров, возможно, дало группе преимущество перед синапсидами, у которых были легкие, в которых воздух приливно входил и выходил через сеть бронхов, заканчивающихся альвеолами , которые представляли собой тупики. Более высокая эффективность газообмена, наблюдаемая в легких архозавров, возможно, была полезна во времена низкого содержания кислорода в атмосфере, которые, как полагают, существовали в мезозое. [37]
Воспроизведение
[ редактировать ]Большинство (если не все) архозавров яйцекладущие . Птицы и крокодилы откладывают яйца с твердой скорлупой, а также вымершие динозавры и крокодиломорфы. Яйца с твердой скорлупой присутствуют как у динозавров, так и у крокодилов, что использовалось как объяснение отсутствия живорождения или яйцеживорождения у архозавров. [38] Однако оба птерозавра [39] и баурузухиды [40] имеют яйца с мягкой скорлупой, а это означает, что твердая скорлупа не является плезиоморфным состоянием. Анатомия таза крикозавров и других метриоринхид. [41] и окаменелые эмбрионы, принадлежащие неархозавру архозавроморфу Dinocephalosaurus , [42] вместе предполагают, что отсутствие живорождения среди архозавров может быть следствием ограничений, специфичных для линии. [ нужны разъяснения ]
Архозавры по своей природе являются сверхранними , о чем свидетельствуют различные динозавры, птерозавры и крокодиломорфы. [43] Однако родительская забота несколько раз развивалась независимо друг от друга у крокодилов, динозавров и этозавров . [44] У большинства таких видов животные закапывают яйца и полагаются на определение пола в зависимости от температуры . Заметным исключением являются неорниты , которые высиживают яйца и полагаются на генетическое определение пола – черта, которая могла бы дать им преимущество в выживании перед другими динозаврами. [45]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Ричард Дж. Батлер; Стивен Л. Брусатте; Майк Райх; Стерлинг Дж. Несбитт; Райнер Р. Шох; Ян Дж. Хорнунг (2011). «Парусная рептилия Ctenosauriscus из последнего раннего триаса Германии, а также время и биогеография раннего излучения архозавров» . ПЛОС ОДИН . 6 (10): e25693. Бибкод : 2011PLoSO...625693B . дои : 10.1371/journal.pone.0025693 . ПМК 3194824 . ПМИД 22022431 .
- ^ Фот, К.; Эскурра, доктор медицины; Сукиас, РБ; Брусатте, СЛ; Батлер, Р.Дж. (2016). «Недооцененная диверсификация стволовых архозавров в среднем триасе предшествовала доминированию динозавров» . Эволюционная биология BMC . 16 (1). Кристиан Фот, Мартин Д. Эскурра, Роланд Б. Сукиас, Стивен Л. Брусатте и Ричард Дж. Батлер.: 188. Бибкод : 2016BMCEE..16..188F . дои : 10.1186/s12862-016-0761-6 . ПМК 5024528 . ПМИД 27628503 .
- ^ «Определение и значение архозавра» . Словарь.com . Проверено 01 марта 2022 г.
- ↑ Перейти обратно: Перейти обратно: а б с д и Несбитт, SJ (2011). «Ранняя эволюция архозавров: взаимоотношения и происхождение основных клад» . Бюллетень Американского музея естественной истории . 352 : 1–292. дои : 10.1206/352.1 . hdl : 2246/6112 . S2CID 83493714 .
- ^ Эскурра, Миннесота; Шайер, ТМ; Батлер, Р.Дж. (2014). «Происхождение и ранняя эволюция саурий: переоценка пермских ископаемых ящериц и время расхождения крокодилов и ящериц» . ПЛОС ОДИН . 9 (2): e89165. Бибкод : 2014PLoSO...989165E . дои : 10.1371/journal.pone.0089165 . ПМЦ 3937355 . ПМИД 24586565 .
- ^ Гарет Дайк; Гэри Кайзер, ред. (2011). Живые динозавры: эволюционная история современных птиц . Джон Уайли и сыновья. стр. 10 . ISBN 978-0-470-65666-2 .
- ↑ Перейти обратно: Перейти обратно: а б Ханна, ДР (2004). Биология рептилий . Издательство Дискавери. стр. 78 и далее. ISBN 978-81-7141-907-4 .
- ↑ Перейти обратно: Перейти обратно: а б Уайт, Т.; Казлев М.А. «Архозавроморфы: Обзор» . Палеос.com. Архивировано из оригинала 20 декабря 2010 года . Проверено 6 сентября 2012 г.
- ^ Пол, Грегори С. (2016). Принстонский полевой справочник по динозаврам (2-е изд.). Принстон, Нью-Джерси: Издательство Принстонского университета. п. 26. ISBN 978-0-691-16766-4 .
- ^ Понселе, Г., и Шимелд, С.М. (2020). Эволюционное происхождение обонятельной системы позвоночных. Открытая биол. 10:200330. дои: 10.1098/rsob.200330
- ^ Гауэр, диджей; Сенников, АГ (2003). «Ранние архозавры России». В Бентоне, MJ; Шишкин, М.А.; Анвин, Д.М. (ред.). Эпоха динозавров в России и Монголии . Кембридж: Издательство Кембриджского университета. стр. 140–159.
- ^ Кулик, Зоя Т.; Лунгмус, Жаклин К.; Ангельчик, Кеннет Д.; Сидор, Кристиан А. (05.11.2021). «Быстрая жизнь в триасе: новые данные об истории жизни листрозавра (Therapsida: Dicynodontia) из северо-восточной Пангеи» . ПЛОС ОДИН . 16 (11): e0259369. Бибкод : 2021PLoSO..1659369K . дои : 10.1371/journal.pone.0259369 . ISSN 1932-6203 . ПМК 8570511 . ПМИД 34739492 .
- ^ «Двоюродный брат динозавра размером с игуану обнаружен в Антарктиде — ScienceDaily» .
- ↑ Перейти обратно: Перейти обратно: а б Броклхерст, Роберт Дж.; Шахнер, Эмма Р.; Кодд, Джонатан Р.; Селлерс, Уильям И. (02 марта 2020 г.). «Дыхательная эволюция архозавров» . Философские труды Королевского общества B: Биологические науки . 375 (1793): 20190140. doi : 10.1098/rstb.2019.0140 . ПМК 7017431 . ПМИД 31928195 .
- ↑ Даррен Нэйш , Эпизод 38: Не слишком потрепанные подкарты. Архивировано 27 января 2016 г. в Wayback Machine.
- ^ Оливейра, ТВ; Соареш, МБ; Шульц, CL (2010). «Trucidocynodon riograndensis gen. nov. et sp. nov. (Eucynodontia), новый цинодонт из верхнего триаса Бразилии (формация Санта-Мария)». Зоотакса . 2382 : 1–71. дои : 10.11646/zootaxa.2382.1.1 .
- ^ Броклхерст, Нил; Панчироли, Эльза; Беневенто, Джемма Луиза; Бенсон, Роджер Би Джей (2021). «Вымирание млекопитающих как движущая сила морфологической радиации кайнозойских млекопитающих» . Современная биология . 31 (13): 2955–2963.е4. Бибкод : 2021CBio...31E2955B . дои : 10.1016/j.cub.2021.04.044 . ПМИД 34004143 . S2CID 234782605 .
- ^ Archosauromorpha: Archosauria - Palaeos. Архивировано 5 апреля 2005 г. в Wayback Machine.
- ^ Хатчинсон, Джон Р.; Фелклер, Дин; Хьюстон, Кэти; Чанг, Ю-Мэй; Брюгген, Джон; Кледзик, Дэвид; Влит, Кент А. (17 декабря 2019 г.). «Дивергентная эволюция наземных двигательных способностей у современных крокодилов» . Научные отчеты . 9 (1): 19302. Бибкод : 2019NatSR...919302H . дои : 10.1038/s41598-019-55768-6 . ISSN 2045-2322 . ПМЦ 6917812 . ПМИД 31848420 .
- ^ Гауэр, диджей; Уилкинсон, М. (1996). «Есть ли консенсус относительно филогении базальных архозавров?» (PDF) . Труды Королевского общества Б. 263 (1375): 1399–1406. дои : 10.1098/rspb.1996.0205 . S2CID 86610229 .
- ^ Бентон, MJ (2005). Палеонтология позвоночных , 3-е изд. Блэквелл Сайенс Лтд.
- ^ Гёлих, UB; Кьяппе, LM; Кларк, Дж. М.; Сьюс, Х.-Д. (2005). «Систематическое положение предполагаемого динозавра позднеюрского периода Macelognathus (Crocodylomorpha: Sphenosuchia)». Канадский журнал наук о Земле . 42 (3): 307–321. Бибкод : 2005CaJES..42..307G . дои : 10.1139/e05-005 .
- ^ Коуп, Эдвард Дринкер (1869). «Краткий обзор вымерших батрахий, рептилий и авесов Северной Америки» . Труды Американского философского общества . 14 (1): 1–252. дои : 10.2307/1005355 . hdl : 2027/nyp.33433090912423 . JSTOR 1005355 .
- ↑ Перейти обратно: Перейти обратно: а б Бентон, MJ (2004). «Происхождение и родство динозавров». В Вейшампеле, Д.Б.; Додсон, Пр; Осмольска, Х. (ред.). Динозаврия (2-е изд.). Беркли: Издательство Калифорнийского университета. стр. 7–19 . ISBN 978-0-520-24209-8 .
- ^ Брошюры по биологии: Коллекция кофоидов , том. 2900 (1878), с. 731
- ↑ Перейти обратно: Перейти обратно: а б с д Серено, ПК (1991). «Базальные архозавры: филогенетические связи и функциональные последствия». Мемуары (Общество палеонтологии позвоночных) . 2 :1–53. дои : 10.2307/3889336 . JSTOR 3889336 .
- ^ Готье, Дж. А. (1986). «Монофилия ящеров и происхождение птиц». В Падиане, К. (ред.). Происхождение птиц и эволюция полета. Мемуары Калифорнийской академии наук . Том. 8. Сан-Франциско: Калифорнийская академия наук. стр. 1–55.
- ^ Бентон, MJ; Кларк, Дж. М. (1985). «Филогения архозавров и взаимоотношения крокодилов». В Бентоне, MJ (ред.). Филогения и классификация четвероногих . Том. 1. Оксфорд: Кларендон Пресс. стр. 295–338. ISBN 978-0-19-857712-6 .
- ^ Серено, ПК; Аркуччи, AB (1990). «Монофилия голеностопных архозавров и происхождение голеностопных суставов птиц и крокодилов». Новогодний альбом по геологии и палеонтологии, Трактаты . 180 :21-52. дои : 10.1127/njgpa/180/1990/21 . S2CID 256805773 .
- ^ Десмонд, Адриан Дж., Горячие динозавры: революция в палеонтологии. 1976, Dial Press, стр. 87.
- ^ Рубен, Дж.; и др. (1996). «Метаболический статус некоторых динозавров позднего мела» (PDF) . Наука . 273 (5279): 120–147. Бибкод : 1996Sci...273.1204R . дои : 10.1126/science.273.5279.1204 . S2CID 84693210 .
- ^ Рубен, Дж.; и др. (1997). «Структура легких и вентиляция легких тероподовых динозавров и ранних птиц». Наука . 278 (5341): 1267–1270. Бибкод : 1997Sci...278.1267R . дои : 10.1126/science.278.5341.1267 .
- ^ Персоны, Уолтер С.; Карри, Филип Дж. (2015). «Щетинка перед пухом: новый взгляд на функциональное происхождение перьев» . Эволюция . 69 (4): 857–862. дои : 10.1111/evo.12634 . ISSN 1558-5646 . ПМИД 25756292 . S2CID 24319963 .
- ^ Сеймур, RS; Беннетт-Стампер, CL; Джонстон, Южная Дакота; Кэрриер, Д.Р. и Григг, Г.К. (2004). «Доказательства эндотермических предков крокодилов в основе эволюции архозавров». Физиол. Биохим. Зоол . 77 (6): 1051–1067. дои : 10.1086/422766 . HDL : 2440/1933 . ПМИД 15674775 . S2CID 10111065 .
- ^ Саммерс, AP (2005). «Эволюция: Добросердечные крокодилы» . Природа . 434 (7035): 833–834. Бибкод : 2005Natur.434..833S . дои : 10.1038/434833а . ПМИД 15829945 . S2CID 4399224 .
- ↑ Перейти обратно: Перейти обратно: а б Фермер, CG; Сандерс, К. (2010). «Однонаправленный поток воздуха в легких аллигаторов». Наука . 327 (5963): 338–340. Бибкод : 2010Sci...327..338F . дои : 10.1126/science.1180219 . ПМИД 20075253 . S2CID 206522844 .
- ^ Лиза Гроссман (14 января 2010 г.). «Аллигаторы дышат, как птицы» . Новости науки . Архивировано из оригинала 29 сентября 2012 года . Проверено 14 января 2010 г.
- ^ Робин М. Эндрюс; Том Мэтис (2000). «Естественная история развития рептилий: ограничения эволюции живорождения» . Бионаука . 50 (3): 227–238. doi : 10.1641/0006-3568(2000)050[0227:NHORDC]2.3.CO;2 .
- ^ Джи, Кью; Джи, Ю.А.; Ченг, Ю.Н.; и др. (декабрь 2004 г.). « » (декабрь 2004 г.). «Палеонтология: яйцо птерозавра с кожистой скорлупой» (PDF) . Природа . 432 (7017): 572. дои : 10.1038/432572a . ПМИД 15577900 . S2CID 4416203 .
- ^ Оливейра, CEM; Сантуччи, РМ; Андраде, МБ; Фульфаро, виджей; Базило, JAF; Бентон, MJ (2011). «Яйца и яичная скорлупа крокодиломорфов из формации Адамантина (группа Бауру), верхний мел Бразилии» . Палеонтология . 54 (2): 309–321. Бибкод : 2011Palgy..54..309O . дои : 10.1111/j.1475-4983.2010.01028.x .
- ^ Эррера, Янина; Фернандес, Марта С.; Ламас, Сусана Г.; Кампос, Лисандро; Талеви, Марианелла; Гаспарини, Зулма (01 февраля 2017 г.). «Морфология сакральной области и репродуктивные стратегии Metriorhynchidae: контриндуктивный подход» . Труды Королевского общества Эдинбурга по наукам о Земле и окружающей среде . 106 (4): 247–255. дои : 10.1017/S1755691016000165 . hdl : 11336/66599 . ISSN 1755-6910 .
- ^ Лю, Цзюнь; Орган, Крис Л.; Бентон, Майкл Дж.; Брэндли, Мэтью С.; Эйчисон, Джонатан К. (14 февраля 2017 г.). «Живорождение у архозавроморфной рептилии» . Природные коммуникации . 8 : 14445. Бибкод : 2017NatCo...814445L . дои : 10.1038/ncomms14445 . ISSN 2041-1723 . ПМК 5316873 . ПМИД 28195584 .
- ^ Марк П. Виттон (2013), Птерозавры: естественная история, эволюция, анатомия, Princeton University Press, ISBN 978-0-691-15061-1
- ^ Аванзини, М.; Далла; Миетто, П; Пьюбелли, Д; Прето, Н; Риго, М; Роги, Дж. (2007). «Место гнездования позвоночных на северо-востоке Италии обнаруживает неожиданно сложное поведение поздних карнийских рептилий» (PDF) . ПАЛЕОС . 22 (5): 465–475. Бибкод : 2007Палай..22..465А . дои : 10.2110/palo.2005.p05-137r . S2CID 131332250 .
- ^ Танака, Кохей (2015). «Пористость яичной скорлупы дает представление об эволюции гнездования динозавров» . ПЛОС ОДИН . 10 (11): e0142829. Бибкод : 2015PLoSO..1042829T . дои : 10.1371/journal.pone.0142829 . ПМЦ 4659668 . ПМИД 26605799 .
Источники
[ редактировать ]- Бентон, MJ (2004). Палеонтология позвоночных (3-е изд.). Блэквелл Наука.
- Кэрролл, РЛ (1988). Палеонтология и эволюция позвоночных» . Нью-Йорк: WH Freeman. ISBN 978-0-7167-1822-2 .
Внешние ссылки
[ редактировать ]- UCMP
- Палеос рассматривает запутанную историю филогении архозавров (генеалогическое древо) и имеет превосходное изображение различных типов лодыжек архозавров.
- Архив филогении Микко Архозаврия


