Физиология динозавров
Физиология динозавров исторически была спорным вопросом, особенно их терморегуляция . В последнее время появилось много новых данных, касающихся физиологии динозавров в целом, включая не только метаболические системы и терморегуляцию, но также дыхательную и сердечно-сосудистую системы.
В первые годы палеонтологии динозавров широко считалось, что это были медлительные, громоздкие и расползающиеся хладнокровные ящерицы . [ сомнительно – обсудить ] Однако с открытием гораздо более полных скелетов на западе Соединенных Штатов, начиная с 1870-х годов, ученые смогли сделать более обоснованную интерпретацию биологии и физиологии динозавров. Эдвард Дринкер Коуп , противник Отниэля Чарльза Марша в « Костяных войнах» , считал по крайней мере некоторых динозавров активными и ловкими, как это видно на картине двух сражающихся лаелапов, созданной под его руководством Чарльзом Р. Найтом . [1]
Параллельно развитие дарвиновской эволюции и открытия археоптерикса и компсогната привели Томаса Генри Хаксли к предположению, что динозавры были тесно связаны с птицами. [2] Несмотря на эти соображения, образ динозавров как крупных рептилий уже укоренился. [1] и большинство аспектов их палеобиологии в первой половине двадцатого века интерпретировались как типично рептильные. [3] Начиная с 1960-х годов и с приходом эпохи Возрождения динозавров , взгляды на динозавров и их физиологию резко изменились, включая открытие пернатых динозавров в отложениях раннего мелового периода в Китае, что указывает на то, что птицы произошли от очень подвижных динозавров- манирапторов .
История [ править ]
Ранние интерпретации


Изучение динозавров началось в 1820-х годах в Англии. Пионеры в этой области, такие как Уильям Бакленд , Гидеон Мантелл и Ричард Оуэн , интерпретировали первые, очень фрагментарные останки как принадлежащие крупным четвероногим животным. [4] Их ранние работы можно увидеть сегодня в Хрустальном дворце динозавров , построенном в 1850-х годах, который представляет известных динозавров как слоновьих ящерицеподобных рептилий. [5] Несмотря на внешний вид рептилий, Оуэн предположил, что сердце и дыхательная система динозавров больше похожи на сердце и дыхательную систему млекопитающих, чем на рептилий. [4]
динозавров Изменение и ренессанс взглядов
В конце 1960-х годов аналогичные идеи появились вновь, начиная с Джона Острома работы о дейнонихах и эволюции птиц . [6] Его ученик Боб Баккер популяризировал меняющуюся мысль в серии статей, начиная с книги «Превосходство динозавров» в 1968 году. [7] В этих публикациях он энергично доказывал, что динозавры были теплокровными и активными животными, способными выдерживать продолжительные периоды высокой активности. В большинстве своих работ Баккер формулировал свои аргументы как новые доказательства, ведущие к возрождению идей, популярных в конце 19 века, часто ссылаясь на продолжающийся ренессанс динозавров . В защиту своей позиции он использовал множество анатомических и статистических аргументов. [8] [9] методология которого вызвала яростные споры среди учёных. [10]
Эти дебаты вызвали интерес к новым методам установления палеобиологии вымерших животных, таким как гистология костей , которые были успешно применены для определения темпов роста многих динозавров.
Сегодня принято считать, что многие или, возможно, все динозавры имели более высокий уровень метаболизма, чем современные рептилии, но также и что ситуация более сложна и разнообразна, чем первоначально предполагал Беккер. Например, в то время как более мелкие динозавры могли быть настоящими эндотермами , более крупные формы могли быть инерционными гомеотермами . [11] [12] или что многие динозавры могли иметь промежуточную скорость метаболизма. [13]
Питание и пищеварение [ править ]
Самые ранние динозавры почти наверняка были хищниками и имели несколько хищных черт со своими ближайшими родственниками, не являющимися динозаврами, такими как Lagosurus , в том числе: относительно большие, изогнутые, похожие на лезвия зубы в больших, широко раскрытых челюстях, которые закрывались, как ножницы; относительно небольшие брюшки , поскольку плотоядным животным не требуется большая пищеварительная система . Более поздние динозавры, считавшиеся хищниками, иногда вырастали намного крупнее, но сохраняли тот же набор особенностей. Вместо того, чтобы пережевывать пищу, эти хищники заглатывали ее целиком. [14]
Пищевые привычки орнитомимозавров и овирапторозавров остаются загадкой: хотя они произошли от хищных теропод , у них маленькие челюсти и отсутствуют лезвиеобразные зубы, как у типичных хищников, но нет никаких свидетельств их диеты или того, как они ее ели и переваривали. . [14]
Особенности других групп динозавров указывают на то, что они были травоядными. Эти функции включают в себя:
- Челюсти, которые лишь слегка открывались и закрывались, так что все зубы встретились одновременно.
- Большие брюшки, способные вместить большое количество растительности и хранить ее в течение длительного времени, необходимого для переваривания растительности.
- Кишки, которые, вероятно, содержали эндосимбиотические микроорганизмы, переваривающие целлюлозу , поскольку ни одно известное животное не может напрямую переваривать этот твердый материал. [14]
Зауроподы , которые были травоядными, не пережевывали пищу, поскольку их зубы и челюсти пригодны только для снятия листьев с растений. Орнитисхи , также травоядные, демонстрируют разнообразие подходов. Бронированные анкилозавры и стегозавры имели маленькие головы, слабые челюсти и зубы и, как полагают, питались почти так же, как зауроподы. были У пахицефалозавров маленькие головы, слабые челюсти и зубы, но отсутствие у них крупной пищеварительной системы предполагает другую диету, возможно, фрукты, семена или молодые побеги, которые были бы для них более питательными, чем листья. [14]
С другой стороны, орнитоподы, такие как гипсилофодон , игуанодон и различные гадрозавры, имели роговые клювы для отщипывания растительности, а также челюсти и зубы, которые были хорошо приспособлены для жевания. У рогатых цератопсов были схожие механизмы. [14]
Часто высказывалось предположение, что по крайней мере некоторые динозавры использовали проглатываемые камни, известные как гастролиты , для облегчения пищеварения, перемалывая пищу в мышечных желудках , и что эта особенность была у них общей с птицами. В 2007 году Оливер Уингс проанализировал упоминания гастролитов в научной литературе и обнаружил значительную путаницу, начиная с отсутствия согласованного и объективного определения «гастролита». Он обнаружил, что проглоченные твердые камни или песок могут способствовать пищеварению у птиц, которые в основном питаются зерном, но это может быть не так важно, и что птицы, которые едят насекомых летом, а зерно зимой, обычно избавляются от камней и песка летом. Гастролиты часто описывались как важные для динозавров -зауроподов , чья растительная диета требовала очень тщательного пищеварения, но Уингс пришел к выводу, что эта идея неверна: гастролиты встречаются лишь с небольшим процентом окаменелостей зауропод; там, где они были обнаружены, их количество слишком мало, а во многих случаях камни слишком мягкие, чтобы их можно было эффективно измельчать пищу; большинство этих гастролитов тщательно отполированы, но гастролиты, используемые современными животными для измельчения пищи, становятся шероховатыми от износа и разъедаются желудочными кислотами; следовательно, гастролиты зауроподов, вероятно, были проглочены случайно. С другой стороны, он пришел к выводу, что гастролиты, найденные в окаменелостях развитых динозавры- тероподы , такие как Sinornithomimus и Caudipteryx, напоминают птиц, и что использование гастролитов для измельчения пищи могло появиться на ранних стадиях развития группы динозавров, от которой произошли и эти динозавры, и птицы. [15] [16]
биология Репродуктивная
При откладке яиц у самок птиц в конечностях вырастает особый тип костей между твердой внешней костью и костным мозгом . [17] Эта мозговая кость, богатая кальцием , используется для изготовления яичной скорлупы, и птицы, которые ее производят, поглощают ее, когда закончили откладывать яйца. [18] Медуллярная кость была обнаружена в окаменелостях тероподов тираннозавра и аллозавра, а также орнитопод тенонтозавра . [18] [19]
Поскольку линия динозавров, включающая аллозавра и тираннозавра, на очень ранних этапах эволюции динозавров отошла от линии, которая привела к тенонтозаврам , наличие мозговой кости в обеих группах позволяет предположить, что динозавры в целом производили мозговую ткань. С другой стороны, крокодилы родственниками динозавров , которые являются вторыми ближайшими после птиц, не производят медуллярную кость. Эта ткань, возможно, впервые появилась у орнитодиров , триасовой группы архозавров, от которой, как полагают, произошли динозавры. [18]
Медуллярная кость была обнаружена у особей среднего размера, что позволяет предположить, что динозавры достигали половой зрелости до того, как стали взрослыми. Половая зрелость на уровне полувзрослой особи также наблюдается у рептилий и млекопитающих среднего и крупного размера, но птицы и мелкие млекопитающие достигают половой зрелости только после того, как они становятся взрослыми, что происходит в течение первого года жизни. Ранняя половая зрелость связана и с особенностями жизненного цикла животных: детеныши рождаются относительно хорошо развитыми, а не беспомощными; и уровень смертности среди взрослых высок. [18]
Дыхательная система [ править ]
Подушки безопасности [ править ]

Примерно с 1870 года ученые пришли к общему мнению, что посткраниальные скелеты многих динозавров содержали множество полостей, заполненных воздухом ( посткраниальная скелетная пневматизация , особенно в позвонках). Пневматизация черепа (например, околоносовых пазух ) встречается как у синапсидов , так и у архозавров. , но посткраниальная пневматизация встречается только у птиц, нептичьих ящеротазовых динозавров и птерозавров .
Долгое время эти полости считались просто устройствами для снижения веса, но Беккер предположил, что они соединены с воздушными мешками, птиц подобными тем, которые делают дыхательную систему наиболее эффективной из всех животных. [9]
Джон Рубен и др. (1997, 1999, 2003, 2004) оспаривали это и предположили, что у динозавров была «приливная» дыхательная система (внутри и наружу), приводимая в действие крокодилоподобным печеночно-поршневым механизмом – мышцы, прикрепленные в основном к лобку , тянут печень назад, что заставляет легкие расширяются для вдоха; когда эти мышцы расслабляются, легкие возвращаются к своему прежнему размеру и форме, и животное выдыхает. Они также представили это как повод усомниться в том, что птицы произошли от динозавров . [20] [21] [22] [23] [24]
Критики утверждают, что без птичьих воздушных мешков скромные улучшения в некоторых аспектах кровеносной и дыхательной систем современной рептилии позволили бы рептилии получать от 50% до 70% потока кислорода, как у млекопитающего аналогичного размера. [25] и что отсутствие воздушных мешков у птиц не предотвратит развитие эндотермии. [26] В научных журналах было опубликовано очень мало официальных опровержений утверждения Рубена и др. о том, что у динозавров не могло быть воздушных мешков птичьего типа; но один указывает на то, что окаменелость Sinosauropteryx , на которой они основывали большую часть своих аргументов, была сильно сплющена, и поэтому было невозможно сказать, имела ли печень правильную форму, чтобы действовать как часть печеночного поршневого механизма. [27] В некоторых недавних статьях просто отмечается без дальнейших комментариев, что Ruben et al. выступал против наличия воздушных мешков у динозавров. [28]
Исследователи представили доказательства и аргументы в пользу наличия воздушных мешков у зауроподов , « прозауроподов », целурозавров , цератозавров и теропод Aerosteon и Coelophys .
У продвинутых зауроподов («неозавроподов») на позвонках нижней части спины и тазобедренных суставов имеются признаки наличия воздушных мешков. У ранних зауроподов такие особенности наблюдаются только в шейных позвонках. последовательность развития, обнаруженную у эмбрионов птиц Если ориентироваться на , то воздушные мешки на самом деле возникли раньше, чем каналы в скелете, которые вмещают их в более поздних формах. [29] [30]

Признаки наличия воздушных мешков также были обнаружены у теропод. Исследования показывают, что окаменелости целурозавров , [31] цератозавры , [28] а у теропод Coelophys и Aerosteon имеются воздушные мешки. Целофиз из позднего триаса — один из самых ранних динозавров, чьи окаменелости свидетельствуют о наличии каналов для воздушных мешков. [30] У Аэростеона , аллозавра позднего мелового периода , были обнаружены самые птичьи воздушные мешки. [32]
Ранние зауроподоморфы , включая группу, традиционно называемую «просауроподами», также могли иметь воздушные мешки. Хотя возможные пневматические углубления были обнаружены у Платеозавра и Текодонтозавра , они очень маленькие. Одно исследование, проведенное в 2007 году, пришло к выводу, что у прозауроподов, вероятно, были брюшные и шейные воздушные мешки, основываясь на данных о их наличии у сестринских таксонов (теропод и зауроподов). Исследование пришло к выводу, что невозможно определить, имели ли прозауроподы проточные легкие, как у птиц, но воздушные мешки почти наверняка присутствовали. [33] Еще одним указанием на наличие воздушных мешков и их использование в вентиляции легких является реконструкция объема воздухообмена (объема воздуха, обмениваемого при каждом вдохе) у платеозавра , который выражается как отношение объема воздуха к массе тела у 29 мл/кг аналогичны значениям для гусей и других птиц и намного выше, чем типичные значения для млекопитающих. [34]
До сих пор не было обнаружено никаких свидетельств наличия воздушных мешков у птицетазовых динозавров. Но это не означает, что птицетазовые не могли иметь скорость метаболизма, сравнимую с таковой у млекопитающих, поскольку у млекопитающих также нет воздушных мешков. [35]
Было предложено три объяснения развития воздушных мешков у динозавров: [32]
- Увеличение дыхательной способности. Вероятно, это наиболее распространенная гипотеза, которая хорошо согласуется с идеей о том, что у многих динозавров был довольно высокий уровень метаболизма .
- Улучшение баланса и маневренности за счет понижения центра тяжести и уменьшения инерции вращения . Однако это не объясняет расширение воздушных мешков у четвероногих зауроподов.
- В качестве охлаждающего механизма. Похоже, что воздушные мешки и перья у целурозавров развились примерно в одно и то же время . Если бы перья сохраняли тепло, их владельцам потребовались бы средства рассеивания избыточного тепла. Эта идея правдоподобна, но нуждается в дальнейшей эмпирической поддержке.
Расчеты объемов различных частей дыхательной системы зауроподов подтверждают Apatosaurus данные о птичьих воздушных мешках у зауроподов:
- Если предположить, что апатозавр , как и ближайшие выжившие родственники динозавров, крокодилы и птицы, не имел диафрагмы , объем мертвого пространства 30-тонного экземпляра составил бы около 184 литров. Это общий объём рта, трахеи и воздухоносных трубок. Если животное выдыхает меньше этого количества, застоявшийся воздух не выбрасывается и всасывается обратно в легкие при следующем вдохе.
- Оценки его дыхательного объема – количества воздуха, поступающего в легкие или выходящего из них за один вдох – зависят от типа дыхательной системы животного: 904 литра для птиц; 225 литров для млекопитающих; 19 литров если рептилоид.
Исходя из этого, у апатозавра не могла быть рептильной дыхательной системы, поскольку его дыхательный объем был меньше объема мертвого пространства, поэтому застоявшийся воздух не выбрасывался, а всасывался обратно в легкие. Аналогичным образом, система млекопитающих обеспечивает легкие только примерно 225–184 = 41 литров свежего, насыщенного кислородом воздуха при каждом вдохе. Следовательно, у апатозавра должна была быть либо неизвестная в современном мире система, либо система, подобная птичьим , с множеством воздушных мешков и проточным легким. Более того, птичьей системе потребовался бы объем легких около 600 литров, тогда как млекопитающему потребовался бы около 2950 литров, что превысило бы предполагаемые 1700 литров пространства, доступного в 30-тонном сундуке апатозавра . [36]
Дыхательная система динозавров с воздушными мешками, напоминающими птичьи, возможно, была способна поддерживать более высокий уровень активности, чем могут выдержать млекопитающие аналогичного размера и телосложения. Помимо обеспечения очень эффективной подачи кислорода, быстрый поток воздуха мог бы стать эффективным механизмом охлаждения, который необходим для активных, но слишком крупных животных, чтобы избавиться от всего избыточного тепла через кожу. [35]
Палеонтолог Питер Уорд утверждал, что эволюция системы воздушных мешков, которая впервые появилась у самых ранних динозавров, могла быть реакцией на очень низкий уровень кислорода в атмосфере (11%) в карнийский и норийский периоды триасового периода. [37]

Крючковатые отростки на ребрах [ править ]
есть шпоры, называемые « крючковидными отростками У птиц на задних краях ребер », которые дают грудным мышцам больше рычагов при прокачивании грудной клетки для улучшения снабжения кислородом. Размер крючковидных отростков связан с образом жизни птиц и потребностями в кислороде: самые короткие они у ходячих птиц и самые длинные у ныряющих, которым при всплытии на поверхность необходимо быстро пополнять запасы кислорода. Нептичьи динозавры -манирапторы также имели эти крючковидные отростки, и они были пропорционально такой же длины, как и у современных ныряющих птиц, что указывает на то, что манирапторам требовалась мощная подача кислорода. [38] [39]
Пластины, которые, возможно, функционировали так же, как крючковидные отростки, наблюдались в окаменелостях птицетазового динозавра Thescelosaurus и были интерпретированы как свидетельство высокого потребления кислорода и, следовательно, высокой скорости метаболизма. [40]
Носовые раковины [ править ]
Носовые раковины представляют собой извитые структуры тонкой кости в полости носа . У большинства млекопитающих и птиц они присутствуют и выстланы слизистыми оболочками , выполняющими две функции. Они улучшают обоняние за счет увеличения площади, доступной для поглощения переносимых по воздуху химикатов, согревают и увлажняют вдыхаемый воздух, а также извлекают тепло и влагу из выдыхаемого воздуха, чтобы предотвратить высыхание легких.
Джон Рубен и другие утверждали, что у динозавров не было обнаружено никаких свидетельств наличия носовых раковин. У всех динозавров, которые они исследовали, носовые ходы были слишком узкими и короткими, чтобы вместить носовые раковины, поэтому динозавры не могли поддерживать частоту дыхания, необходимую для метаболизма млекопитающих или птиц в состоянии покоя, потому что их легкие высыхали бы. вне. [20] [21] [41] [42] Однако против этого аргумента были высказаны возражения. Носовые раковины отсутствуют или очень малы у некоторых птиц (например, бескилевых , Procellariiformes и Falconiformes ) и млекопитающих (например, китов, муравьедов , летучих мышей , слонов и большинства приматов ), хотя эти животные полностью эндотермичны и в некоторых случаях очень активны. [43] [44] [45] [46] Другие исследования пришли к выводу, что носовые раковины хрупкие и редко встречаются в окаменелостях. В частности, ничего не было обнаружено у ископаемых птиц. [47]
В 2014 году Джейсон Бурк и другие в Anatomical Record сообщили об обнаружении носовых раковин у пахицефалозавров. [48]
Сердечно-сосудистая система [ править ]

В принципе, можно было бы ожидать, что у динозавров было двухчастное кровообращение, приводимое в движение четырехкамерным сердцем, поскольку многим из них требовалось высокое кровяное давление для доставки крови к головам, которые находились высоко над землей, но легкие позвоночных могут переносить только довольно низкое давление. артериальное давление. [35] В 2000 году скелет тесцелозавра , ныне выставленный в Музее естественных наук Северной Каролины , был описан как включающий остатки четырехкамерного сердца и аорты . Авторы интерпретировали структуру сердца как указание на повышенную скорость метаболизма тесцелозавра на , а не рептилий хладнокровность . [49] Их выводы были оспорены; другие исследователи опубликовали статью, в которой утверждают, что сердце на самом деле представляет собой конкрецию полностью минерального «цемента». Они отмечают: анатомия объекта неверна, например, предполагаемая «аорта» наиболее узкая там, где она встречается с «сердцем», и артерии в ней отсутствуют ответвляющиеся от нее ; «сердце» частично охватывает одно из ребер и имеет внутреннее строение местами из концентрических слоев; и еще один конкремент сохранился позади правой ножки. [50] Первоначальные авторы отстаивали свою позицию; они согласились, что в грудной клетке действительно был какой-то тип конкремента, но он сформировался вокруг и частично сохранил более мышечные части сердца и аорты. [51]
Независимо от идентичности объекта, он может иметь мало отношения к внутренней анатомии и скорости метаболизма динозавров. И современные крокодилы , и птицы, ближайшие ныне живущие родственники динозавров, имеют четырехкамерное сердце, хотя и видоизмененное у крокодилов, поэтому, вероятно, оно было и у динозавров. Однако такие сердца не обязательно связаны со скоростью метаболизма. [52]
и цикл жизненный Рост
Не было обнаружено ни одного яйца динозавра размером больше баскетбольного мяча, а эмбрионы крупных динозавров были обнаружены в относительно небольших яйцах, например Майазавра . [53] Как и млекопитающие, динозавры перестали расти, когда достигли типичных для своего вида размеров взрослой особи, в то время как взрослые рептилии продолжали медленно расти, если у них было достаточно еды. Динозавры всех размеров росли быстрее, чем современные рептилии аналогичного размера; но результаты сравнений с «теплокровными» современными животными такого же размера зависят от их размеров: [54] [55]
| Вес (кг) | Сравнительная скорость роста динозавров | Современные животные в этом размерном диапазоне |
|---|---|---|
| 0.22 | Медленнее сумчатых | Крыса |
| 1 – 20 | Похож на сумчатых, медленнее ранних птиц (тех, которые рождаются способными бегать). | От морской свинки до андского кондора |
| 100 – 1000 | Быстрее, чем сумчатые, похожи на скороспелых птиц, медленнее, чем плацентарные млекопитающие. | От рыжего кенгуру до белого медведя |
| 1500 – 3500 | Похож на большинство плацентарных млекопитающих. | От американского бизона до носорога |
| 25000 и выше | Очень быстрые, похожие на современных китов ; но примерно вдвое меньше, чем у увеличенной альтрициальной птицы (той, которая рождается беспомощной) - если бы можно было увеличить птицу до 25 000 килограммов (25 длинных тонн; 28 коротких тонн) | Киты |

Тираннозавр рекс продемонстрировал «подростковый скачок роста»: [56] [57]
- ½ тонны в 10 лет
- очень быстрый рост примерно до 2 тонн в подростковом возрасте (около ½ тонны в год).
- незначительный рост после второго десятилетия.
Исследование одного скелета гадрозавра Hypacrosaurus, проведенное в 2008 году , пришло к выводу, что этот динозавр рос еще быстрее, достигнув своего полного размера примерно в 15 лет; Основным доказательством было количество и расположение годичных колец в его костях. Авторы обнаружили, что это согласуется с теорией жизненного цикла, согласно которой виды-жертвы должны расти быстрее, чем их хищники, если они теряют много молоди из-за хищников, а местная окружающая среда обеспечивает достаточно ресурсов для быстрого роста. [58]
Похоже, что отдельные динозавры жили довольно недолго: например, самому старому (на момент смерти) тираннозавру, обнаруженному до сих пор, было 28 лет, а самому старому зауроподу — 38 лет. [56] Хищничество, вероятно, было причиной высокой смертности очень молодых динозавров, а сексуальная конкуренция - высокой смертности половозрелых динозавров. [59]
Метаболизм [ править ]
Научное мнение об образе жизни, метаболизме и терморегуляции динозавров со временем менялось с момента открытия динозавров в середине 19 века. Активность метаболических ферментов зависит от температуры, поэтому контроль температуры жизненно важен для любого организма, будь то эндотермического или экзотермного. Организмы можно разделить на пойкилотермных (пойкило – меняющийся), которые устойчивы к колебаниям внутренней температуры, и гомеотермных (гомео – тот же), которые должны поддерживать постоянную внутреннюю температуру. Животных можно разделить на эндотермных , которые регулируют свою температуру изнутри, и эктотермных , которые регулируют температуру с помощью внешних источников тепла.
« Теплокровность » — сложный и довольно неоднозначный термин, поскольку включает в себя некоторые или все из:
- Гомеотермия , т.е. поддержание достаточно постоянной температуры тела. Современные эндотермы поддерживают различные температуры: от 28 ° C (82 ° F) до 30 ° C (86 ° F) у однопроходных и ленивцев ; от 33 ° C (91 ° F) до 36 ° C (97 ° F) у сумчатых ; От 36 °C (97 °F) до 38 °C (100 °F) в большинстве плацентарных клеток ; и около 41 ° C (106 ° F) у птиц. [35]
- Тахиметаболизм , то есть поддержание высокой скорости обмена веществ , особенно в состоянии покоя. Для этого необходима достаточно высокая и стабильная температура тела, так как биохимические процессы протекают примерно вдвое быстрее, если температура животного снижается на 10°С; большинство ферментов имеют оптимальную рабочую температуру , и их эффективность быстро падает за пределы предпочтительного диапазона. [60]
- Эндотермия , то есть способность генерировать тепло внутри себя, например, путем «сжигания» жира, а не посредством таких действий, как купание или мышечная активность. Хотя эндотермия в принципе является наиболее надежным способом поддержания достаточно постоянной температуры, она дорогая; например, современным млекопитающим нужно в 10–13 раз больше еды, чем современным рептилиям. [35]
Крупные динозавры, возможно, также поддерживали свою температуру с помощью инерционной гомеотермии, также известной как «объемная гомеотермия» или «массовая гомеотермия». Другими словами, теплоемкость таких крупных животных была настолько велика, что для существенного изменения их температуры потребовалось бы два дня и более, и это сгладило бы колебания, вызванные суточными температурными циклами. Этот эффект сглаживания наблюдался у крупных черепах и крокодилов , но Платеозавр , весивший около 700 килограммов (1500 фунтов), возможно, был самым маленьким динозавром, на котором он был бы эффективен. Инерционная гомеотермия была бы невозможна ни для мелких видов, ни для молоди более крупных видов. [35] Растительность, бродящая в кишках крупных травоядных, также может выделять значительное количество тепла, но этот метод поддержания высокой и стабильной температуры был бы невозможен для хищников, мелких травоядных или молоди более крупных травоядных. [61]
Поскольку внутренние механизмы вымерших существ непознаваемы, большая часть дискуссий сосредоточена на гомеотермии и тахиметаболизме.
Оценка скорости метаболизма осложняется различием между скоростью в состоянии покоя и во время активности. У всех современных рептилий, а также у большинства млекопитающих и птиц максимальные показатели при полной активности в 10–20 раз превышают минимальные показатели в состоянии покоя. Однако у некоторых млекопитающих эти скорости различаются в 70 раз. Теоретически у наземных позвоночных может быть уровень метаболизма рептилий в состоянии покоя и уровень метаболизма птиц, когда они работают на пределе своих возможностей. Однако животное с такой низкой скоростью отдыха не сможет быстро расти. Огромные травоядные зауроподы, возможно, так постоянно перемещались в поисках пищи, что их энергетические затраты были бы примерно одинаковыми, независимо от того, была ли скорость их метаболизма в состоянии покоя высокой или низкой. [62]
Теории [ править ]
Основные возможности заключаются в следующем: [35]
- Динозавры были хладнокровными, как и современные рептилии, за исключением того, что большой размер многих из них стабилизировал температуру их тела.
- Они были теплокровными и больше напоминали современных млекопитающих или птиц, чем современных рептилий.
- Они не были ни хладнокровными, ни теплокровными в современных терминах, но имели метаболизм, который отличался и в некотором смысле был промежуточным между метаболизмом современных хладнокровных и теплокровных животных.
- В их число вошли животные с двумя или тремя из этих типов обмена веществ.
Динозавры существовали около 150 миллионов лет, поэтому весьма вероятно, что у разных групп развился разный метаболизм и режимы терморегуляции, а у некоторых из них развилась физиология, отличная от первых динозавров.
Если бы все или некоторые динозавры имели промежуточный метаболизм, они могли бы иметь следующие особенности: [35]
- Низкая скорость метаболизма в состоянии покоя — что уменьшит количество необходимой им пищи и позволит им использовать больше этой пищи для роста, чем животные с высокой скоростью метаболизма в состоянии покоя.
- Инерционная гомеотермия
- Способность контролировать потерю тепла путем расширения и сужения кровеносных сосудов под кожей, как это делают многие современные рептилии.
- Двухчастное кровообращение осуществляется четырехкамерным сердцем.
- Высокая аэробная способность , позволяющая поддерживать постоянную активность.
Роберт Рид предположил, что таких животных можно рассматривать как «неудавшихся эндотермистов». Он предполагал, что и динозавры, и триасовые предки млекопитающих проходят через стадию с этими особенностями. Млекопитающие были вынуждены уменьшаться в размерах, поскольку архозавры стали доминировать в экологических нишах для средних и крупных животных. Их уменьшающийся размер сделал их более уязвимыми к потерям тепла, поскольку это увеличило соотношение площади поверхности к массе и, таким образом, вынудило их увеличить внутреннее тепловыделение и, таким образом, стать полными эндотермами. С другой стороны, динозавры превратились в животных от средних до очень крупных и, таким образом, смогли сохранить «промежуточный» тип метаболизма. [35]
Костная структура [ править ]
Арман де Риклес обнаружил гаверсовы каналы в костях динозавров и утверждал, что у динозавров имеются доказательства эндотермии. Эти каналы часто встречаются у «теплокровных» животных и связаны с быстрым ростом и активным образом жизни, поскольку помогают перерабатывать кость, способствуя быстрому росту и восстанавливая повреждения, вызванные стрессом или травмами. [63] Плотная вторичная гаверсова кость, образующаяся при ремоделировании, встречается у многих живых эндотермных животных, а также динозавров, птерозавров и терапсидов . Вторичные гаверсовы каналы коррелируют с размером и возрастом, механическим стрессом и кругооборотом питательных веществ. Наличие вторичных гаверсовых каналов предполагает сопоставимый рост костей и продолжительность жизни млекопитающих и динозавров. [64] Баккер утверждал, что присутствие фиброламеллярной кости (образующейся быстро и имеющей волокнистый, плетеный вид) в окаменелостях динозавров является свидетельством эндотермии. [9]
Однако в результате других, преимущественно более поздних исследований, структура костей не считается надежным индикатором обмена веществ у динозавров, млекопитающих или рептилий:
- Кости динозавров часто содержат линии задержки роста (LAG), образованные чередующимися периодами медленного и быстрого роста; Фактически, многие исследования подсчитывают годичные кольца, чтобы оценить возраст динозавров. [55] [56] Образование годичных колец обычно обусловлено сезонными изменениями температуры, и это сезонное влияние иногда рассматривается как признак медленного метаболизма и эктотермии. Но годичные кольца встречаются у белых медведей и у млекопитающих, впадающих в спячку. [65] [66] Связь между LAG и зависимостью от сезонного роста остается нерешенной. [67]
- Фиброламеллярная кость довольно часто встречается у молодых крокодилов и иногда встречается у взрослых. [68] [69]
- Гаверсова кость была обнаружена у черепах, крокодилов и черепах. [70] но часто отсутствует у мелких птиц, летучих мышей, землероек и грызунов. [69]
Тем не менее де Риклес упорно продолжал исследования строения костей динозавров и архозавров . В середине 2008 года он стал соавтором статьи, в которой исследовались образцы костей широкого спектра архозавров, включая ранних динозавров, и пришел к следующему выводу: [71]
- Даже самые ранние архозавроформы , возможно, были способны к очень быстрому росту, что позволяет предположить, что у них был довольно высокий уровень метаболизма. Хотя делать выводы о самых ранних архозаврообразных на основе более поздних форм сложно, поскольку весьма вероятны видоспецифичные вариации в структуре костей и скорости роста, существуют исследовательские стратегии, которые могут минимизировать риск того, что такие факторы вызовут ошибки в анализе.
- архозавры разделились на три основные группы В триасе : орнитодиры , от которых произошли динозавры, продолжали быстро расти; предки крокодилов переняли более типичные для «рептилий» медленные темпы роста; и большинство других архозавров триаса имели средние темпы роста.
Остеогистологический анализ плотности и плотности сосудов, формы и площади остеоцитов пришел к выводу, что нептичьи динозавры и большинство архозаврообразных (кроме Proterosuchus , крокодилов и фитозавров ) сохраняли тепло и имели скорость метаболизма в состоянии покоя, аналогичную таковой у современных млекопитающих и птиц. [72]
, артериальное давление и кровоток Скорость веществ обмена
Эндотермные животные в значительной степени полагаются на аэробный метаболизм и имеют высокие показатели потребления кислорода во время активности и покоя. Кислород, необходимый тканям, переносится кровью, и, следовательно, скорость кровотока и кровяное давление в сердце у теплокровных эндотермов значительно выше, чем у холоднокровных эктотермов. [73] Минимальное кровяное давление динозавров можно измерить, оценив расстояние по вертикали между сердцем и макушкой головы, поскольку этот столб крови должен иметь давление внизу, равное гидростатическому давлению, полученному из плотности крови и гравитация. К этому давлению добавляется давление, необходимое для перемещения крови по системе кровообращения. В 1976 году было отмечено, что из-за своего роста у многих динозавров минимальное кровяное давление было в пределах эндотермического диапазона и что у них должно было быть четырехкамерное сердце, чтобы отделить контур высокого давления, идущий к телу, от контура низкого давления к легкие. [74] Было неясно, было ли у этих динозавров высокое кровяное давление просто для поддержания кровотока или для поддержания высоких скоростей кровотока, необходимых для эндотермии, или для того и другого.
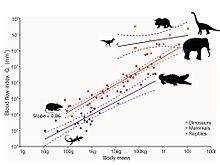
Однако недавний анализ крошечных отверстий в костях ископаемых ног динозавров позволяет оценить скорость кровотока и, следовательно, скорость метаболизма. [75] Отверстия называются питательными отверстиями, а питательная артерия — это главный кровеносный сосуд, проходящий внутрь кости, где он разветвляется на крошечные сосуды системы гаверсовых каналов . Эта система отвечает за замену старой кости новой, тем самым устраняя микроразрывы, возникающие естественным образом во время передвижения. Без этого восстановления микроразрывы будут накапливаться, что приведет к стрессовым переломам и, в конечном итоге, к катастрофическому разрушению кости. Размер питательного отверстия определяет показатель кровотока через него в соответствии с уравнением Хагена-Пуазейля . Разумеется, размер также связан с размером тела животного, поэтому этот эффект устраняется анализом аллометрии . Индекс кровотока питательного отверстия бедренных костей у современных млекопитающих увеличивается прямо пропорционально максимальной скорости метаболизма животных, измеряемой во время максимального устойчивого передвижения. Индекс кровотока у млекопитающих примерно в 10 раз больше, чем у экзотермных рептилий. У десяти видов ископаемых динозавров из пяти таксономических групп показатели даже выше, чем у млекопитающих, с учетом размеров тела, что указывает на то, что они были высокоактивными аэробными животными. Таким образом, высокая скорость кровотока, высокое кровяное давление, четырехкамерное сердце и устойчивый аэробный метаболизм — все это соответствует эндотермия .
роста Темпы
Динозавры сравнительно быстро выросли из маленьких яиц до веса в несколько тонн. Естественная интерпретация этого состоит в том, что динозавры очень быстро преобразовывали пищу в массу тела, что требует довольно быстрого метаболизма как для активного добывания пищи, так и для быстрого ее усвоения. [76] Развивающаяся кость у молодых особей отчетливо пористая, что связано с васкуляризацией и скоростью отложения кости, что позволяет предположить, что темпы роста близки к тем, которые наблюдаются у современных птиц.
Но предварительное исследование взаимосвязи между размером взрослых особей, скоростью роста и температурой тела пришло к выводу, что у более крупных динозавров температура тела была выше, чем у более мелких; По оценкам, у апатозавра , самого крупного динозавра в выборке, температура тела превышала 41 °C (106 °F), тогда как у более мелких динозавров температура тела, по оценкам, составляла около 25 °C (77 °F). [77] – для сравнения, нормальная температура тела человека составляет около 37 °C (99 °F). [78] [79] На основании этих оценок исследование пришло к выводу, что крупные динозавры были инерционными гомеотермами (их температура стабилизировалась за счет их огромного размера) и что динозавры были экзотермными (в просторечии «хладнокровными», потому что они не выделяли столько тепла, сколько млекопитающие). когда не двигаешься и не перевариваешь пищу). [77] Эти результаты согласуются с взаимосвязью между размерами динозавров и скоростью роста (описанной выше). [80] [81] Исследования зауроподоморфа Massospondylus и раннего теропода Syntarsus ( Megapnosaurus ) показывают темпы роста 3 кг/год и 17 кг/год соответственно, что намного медленнее, чем у майазауры и наблюдаемых у современных птиц. [82]
кислорода в изотопов Соотношение костях
Соотношение изотопов 16 О и 18 Содержание кислорода в кости зависит от температуры, при которой образовалась кость: чем выше температура, тем больше 16 О. Баррик и Шауэрс (1999) проанализировали соотношения изотопов у двух теропод , обитавших в регионах с умеренным климатом и сезонными колебаниями температуры: тираннозавра (США) и гиганотозавра (Аргентина): [83]
- спинные позвонки обоих динозавров не показали никаких признаков сезонных изменений, что указывает на то, что оба сохраняли постоянную внутреннюю температуру, несмотря на сезонные колебания температуры воздуха.
- ребра и кости ног обоих динозавров демонстрировали большую изменчивость температуры и более низкую среднюю температуру по мере увеличения расстояния от позвонков.
Баррик и Шоуерс пришли к выводу, что оба динозавра были эндотермными животными, но имели более низкий уровень метаболизма, чем современные млекопитающие, и что инерционная гомеотермия была важной частью их терморегуляции во взрослом возрасте. Их аналогичный анализ некоторых птицетазовых позднемелового периода в 1996 году пришел к выводу, что эти животные демонстрируют аналогичную картину. [84]
Однако эта точка зрения была оспорена. Данные указывают на гомеотермию, но сами по себе не могут доказать эндотермию. Во-вторых, производство кости, возможно, не было непрерывным в областях вблизи конечностей конечностей – в скелетах аллозавров линии задержанного роста («LAG»; скорее как кольца роста ) редки или отсутствуют в крупных костях конечностей, но распространены в пальцах и пальцы ног. Хотя нет абсолютных доказательств того, что ЛАГ связаны с температурой, они могут отмечать моменты, когда конечности были настолько прохладными, что кости переставали расти. Если бы это было так, данные о соотношении изотопов кислорода были бы неполными, особенно в те времена, когда конечности были наиболее прохладными. Соотношения изотопов кислорода могут быть ненадежным методом оценки температуры, если невозможно доказать, что рост костей был одинаково непрерывным во всех частях животного. [35]
- жертва Соотношение хищник
- хладнокровным хищникам нужно гораздо меньше еды, чем теплокровным, поэтому данная масса добычи может прокормить гораздо больше хладнокровных хищников, чем теплокровных.
- соотношение общей массы хищников и жертв в сообществах динозавров было гораздо больше похоже на соотношение современных и новейших теплокровных сообществ, чем на соотношение современных или ископаемых хладнокровных сообществ.
- следовательно, хищные динозавры были теплокровными. А поскольку самые ранние динозавры (например, Staurikosaurus , Herrerasaurus ) были хищниками, все динозавры, должно быть, были теплокровными.
Этот аргумент подвергся критике по нескольким причинам и больше не воспринимается всерьез (следующий список критических замечаний далеко не исчерпывающий): [86] [87]
- Оценки веса динозавров сильно различаются, и даже небольшое отклонение может существенно повлиять на рассчитанное соотношение хищника и жертвы.
- Его выборка , возможно, не была репрезентативной. Баккер получил свои цифры путем подсчета музейных образцов, но они имеют уклон в сторону редких или особенно хорошо сохранившихся экземпляров и не отражают то, что существует в пластах окаменелостей. Даже пласты окаменелостей могут неточно отражать реальные популяции: например, более мелкие и молодые животные имеют менее крепкие кости и, следовательно, вероятность их сохранения меньше.
- Для крупных экзотермных хищников опубликованных соотношений хищник-жертва нет, поскольку такие хищники очень редки и в основном встречаются только на довольно небольших островах. Столь же редки крупные экзотермические травоядные. Поэтому Баккеру пришлось сравнивать соотношение хищников и жертв среди млекопитающих с соотношениями в сообществах рыб и беспозвоночных, где продолжительность жизни намного короче, а другие различия также искажают сравнение.
- Концепция предполагает, что популяции хищников ограничены только наличием добычи. Однако другие факторы, такие как нехватка мест гнездования, каннибализм или нападение одного хищника на другого, могут удерживать популяцию хищников ниже предела, установленного биомассой добычи, и это может ошибочно уменьшить соотношение хищник-жертва.
- Экологические факторы могут, например, ошибочно уменьшить соотношение хищник-жертва: хищник может охотиться только на некоторые из присутствующих видов «добычи»; болезни, паразиты и голод могут убить некоторых животных-жертв прежде, чем хищники получат возможность охотиться на них.
- Очень сложно точно сказать, что на что охотится. Например, на молодняк травоядных животных могут охотиться ящерицы и змеи, а на взрослых особей охотятся млекопитающие. И наоборот, молодь многих хищников питается в основном беспозвоночными и по мере роста переходит на позвоночных.
Осанка и походка [ править ]
Этот раздел нуждается в дополнительных цитатах для проверки . ( сентябрь 2008 г. ) |

Конечности динозавров были прямыми и держались под телом, а не раскинулись в стороны, как у ящериц и тритонов. Об этом свидетельствуют углы суставных поверхностей и места прикрепления мышц и сухожилий на костях. Попытки изобразить динозавров с раскинутыми конечностями приводят к созданию существ с вывихнутыми бедрами, коленями, плечами и локтями. [88]
Ограничение Кэрриера гласит, что позвоночным животным, дышащим воздухом, с двумя легкими, которые сгибают свое тело в стороны во время передвижения, трудно двигаться и дышать одновременно. Это серьезно ограничивает выносливость и заставляет их проводить больше времени в отдыхе, чем в движении. [89]
Раскидистые конечности требуют бокового сгибания во время передвижения (за исключением черепах и черепах, которые очень медлительны и чья броня делает их тела довольно жесткими). Однако, несмотря на ограничение Кэрриера, раскидистые конечности эффективны для существ, которые проводят большую часть своего времени, отдыхая на животе и двигаясь только в течение нескольких секунд за раз, поскольку такое расположение сводит к минимуму затраты энергии на вставание и лежание.
Прямые конечности увеличивают затраты на вставание и лежание, но позволяют избежать ограничений Кэрриера. Это указывает на то, что динозавры были активными животными, поскольку естественный отбор благоприятствовал бы сохранению разросшихся конечностей, если бы динозавры были вялыми и большую часть времени бодрствования проводили в отдыхе. Активный образ жизни требует метаболизма, который быстро восстанавливает запасы энергии и расщепляет продукты жизнедеятельности, вызывающие утомление, т. е. требует достаточно быстрого обмена веществ и значительной степени гомеотермии.
Кроме того, прямая осанка требует точного баланса, что является результатом быстро функционирующей нервно-мышечной системы. Это предполагает эндотермический метаболизм, поскольку экзотермическое животное не сможет ходить или бегать и, таким образом, уклоняться от хищников, когда его внутренняя температура будет понижена. Другие доказательства эндотермии включают длину конечностей (многие динозавры обладали сравнительно длинными конечностями) и прямохождение, которые сегодня встречаются только у эндотермных животных. [90] Многие двуногие динозавры обладали изящными костями ног и короткими бедрами по сравнению с длиной икры. Обычно это адаптация к частому продолжительному бегу, характерная для эндотермных животных, которые, в отличие от эктотермных животных, способны производить достаточно энергии, чтобы предотвратить начало анаэробного метаболизма в мышцах. [91]
Баккер и Остром указали, что у всех динозавров задние конечности были прямостоячими, а передние конечности у всех четвероногих динозавров были прямостоячими; и что среди живых животных только эндотермические («теплокровные») млекопитающие и птицы имеют прямые конечности (Остром признал, что случайная «высокая походка» крокодилов была частичным исключением). Баккер утверждал, что это явное свидетельство эндотермии у динозавров, в то время как Остром считал это убедительным, но не убедительным. [9] [92]
Исследование 2009 года подтвердило гипотезу о том, что эндотермия была широко распространена, по крайней мере, у более крупных нептичьих динозавров и что она, вероятно, была наследственной для всех динозаврообразных, основываясь на биомеханике бега. [93] хотя также было высказано предположение, что эндотермия появилась гораздо раньше в эволюции архозавроморфов , возможно, даже до возникновения Archosauriformes . [72]

Перья и нити [ править ]
Сейчас нет сомнений в том, что многие виды тероподных динозавров имели перья, в том числе Шувууя , Синозавроптерикс и Дилонг (ранний тираннозавр ). [94] [27] [95] Их интерпретировали как изоляцию и, следовательно, свидетельство теплокровности.
Но прямые, недвусмысленные отпечатки перьев были обнаружены только у целурозавров (к которым относятся, среди прочего, птицы и тираннозавры), поэтому в настоящее время перья не дают нам никакой информации о метаболизме других основных групп динозавров, например, целофизид , цератозавров , карнозавров , или зауроподы . Нитчатый покров также присутствовал, по крайней мере, у некоторых птицетазовых, таких как Tianyulong , Kulindadromeus и Psittacosaurus , что не только указывает на эндотермию в этой группе, но также и на то, что перья уже присутствовали у первых орнитодиранов (последнего общего предка динозавров и птерозавров). Их отсутствие в некоторых группах, таких как Ankylosauria, может быть результатом подавления генов перьев. [96] Хотя нити впервые появились у Coelurosauria, согласно реконструкциям максимального правдоподобия, и что покровные структуры Psittacosaurus , Tianyulong и Kulindadromeus независимо развились из нитей, но это было основано на предположении, что примитивные предки птерозавров были чешуйчатыми. [97]
Окаменевшая кожа карнотавра ( абелизаврида и, следовательно, не целурозавра) представляет собой неоперенную кожу, похожую на рептилию, с рядами выпуклостей. [98] но вывод о том, что Карнотавр обязательно был без перьев, подвергся критике, поскольку отпечатки не покрывают все тело и обнаруживаются только в боковой области, но не на спине. [99] Взрослый карнотавр весил около 2 тонн. [100] а млекопитающие такого размера и крупнее имеют либо очень короткую, редкую шерсть, либо голую кожу, поэтому, возможно, кожа карнотавра ничего не говорит нам о том, были ли перья у более мелких теропод, не являющихся целурозаврами. Известно, что тираннозавроид Ютираннус имел перья и весил 1,1 тонны. [100]
Отпечатки кожи пелорозавра и других зауроподов (динозавров со слоновьим телом и длинной шеей) показывают большие шестиугольные чешуи, а у некоторых зауроподов, таких как сальтазавр , на коже были костные пластины. [101] Кожа цератопсов состояла из крупных полигональных чешуек, иногда с разбросанными круглыми пластинками. [102] «Мумифицированные» останки и отпечатки кожи гадрозавридов обнаруживают галечные чешуйки. Маловероятно, что анкилозавриды, такие как Euoplocephalus , имели изоляцию, поскольку большая часть их поверхности была покрыта костными выступами и пластинами. [103] Точно так же нет никаких доказательств изоляции у стегозавров . Таким образом, изоляция и повышенная скорость метаболизма, лежащие в основе их развития, могли быть ограничены тероподами или даже лишь их подгруппой. [ нужна ссылка ] Отсутствие перьев или другого вида изоляции не указывает на эктотермию или низкий метаболизм, как это наблюдается в относительной безволосости мегафауны млекопитающих, свиней, человеческих детей и лысых летучих мышей , что совместимо с эндотермией. [100]
Полярные динозавры [ править ]
Окаменелости динозавров были найдены в регионах, которые в то время находились близко к полюсам, особенно в юго-восточной Австралии , Антарктиде и на склоне Аляски Северном . Нет никаких свидетельств серьезных изменений угла наклона земной оси , поэтому полярным динозаврам и остальным представителям этих экосистем пришлось бы справляться с такими же экстремальными изменениями продолжительности дня в течение года, которые происходят сегодня на аналогичных широтах (вплоть до полный день без темноты летом и полный день без солнечного света зимой). [104]
Исследования окаменелой растительности показывают, что на Северном склоне Аляски максимальная температура составляла 13 ° C (55 ° F) и минимальная температура от 2 ° C (36 ° F) до 8 ° C (46 ° F) за последние 35 миллионов человек. годы мелового периода (немного прохладнее, чем в Портленде , штат Орегон, но немного теплее, чем в Калгари , Альберта). Несмотря на это, на Северном склоне Аляски нет окаменелостей крупных хладнокровных животных, таких как ящерицы и крокодилы, которые были распространены в то же время в Альберте, Монтане и Вайоминге . Это говорит о том, что по крайней мере некоторые нептичьи динозавры были теплокровными. [104] Было высказано предположение, что североамериканские полярные динозавры могли мигрировать в более теплые регионы с приближением зимы, что позволило бы им обитать на Аляске летом, даже если бы они были хладнокровными. [105] Но поездка туда и обратно туда и в Монтану, вероятно, потребовала бы больше энергии, чем хладнокровное наземное позвоночное животное производит за год; другими словами, динозавры Аляски должны были быть теплокровными, независимо от того, мигрировали они или оставались на зимовку. [106] В статье 2008 года Фила Р. Белла и Эрика Снивли о миграции динозавров предположено, что большинство полярных динозавров, включая теропод , зауроподов , анкилозавров и гипсилофодонтов , вероятно, перезимовали , хотя гадрозавриды, такие как эдмонтозавр, вероятно, были способны совершать ежегодные путешествия туда и обратно на расстояние 2600 км (1600 миль). . [107] [108]

Сложнее определить климат юго-восточной Австралии, когда пласты окаменелостей динозавров были заложены 115–105 миллионов лет назад , ближе к концу раннего мела: эти отложения содержат свидетельства вечной мерзлоты , клиньев льда и бугристого грунта, образованного движение подземного льда, что предполагает, что среднегодовая температура колебалась от -6 ° C (21 ° F) до 5 ° C (41 ° F); исследования изотопов кислорода в этих отложениях дают среднюю годовую температуру от 1,5 ° C (34,7 ° F) до 2,5 ° C (36,5 ° F). Однако разнообразие ископаемой растительности и большие размеры некоторых ископаемых деревьев превосходят то, что встречается сегодня в таких холодных условиях, и никто не объяснил, как такая растительность могла выжить при низких температурах, предполагаемых физическими индикаторами - для сравнения Фэрбенкс На Аляске в настоящее время средняя годовая температура составляет 2,9 ° C (37,2 ° F). [104] Ежегодная миграция из и в юго-восточную Австралию была бы очень трудной для довольно маленьких динозавров, таких как Leaellynasaura , травоядное животное длиной от 60 до 90 сантиметров (3,0 фута), потому что морские пути на север блокировали проход в более теплые места. широты. [104] Образцы костей Leaellynasaura и Timimus , орнитомимида длиной около 3,5 метров (11 футов) и высотой в бедрах 1,5 метра (4,9 футов), позволяют предположить, что у этих двух динозавров были разные способы выживания в холодные и темные зимы: образец Тимимуса имел линии задержанный рост (сокращенно LAG; похож на годичные кольца ), и он, возможно, впал в спячку ; но у образца Leaellynasaura не было обнаружено никаких признаков LAG, поэтому он мог оставаться активным всю зиму. [109] Исследование 2011 года, посвященное костям гипсилофодонтов и теропод, также пришло к выводу, что эти динозавры не впадали в спячку зимой, а оставались активными. [110]
Доказательства поведенческой терморегуляции
Некоторые динозавры, например Спинозавр и Уранозавр , имели на спине «паруса», поддерживаемые шипами, растущими из позвонков . (Это, кстати, справедливо и для синапсидного диметродона .) Такие динозавры могли использовать эти паруса для:
- согревайтесь, греясь «парусами» под прямым углом к солнечным лучам.
- терять тепло, используя «паруса» в качестве радиаторов, стоя в тени или лицом прямо к солнцу или от него.
Но это было очень небольшое меньшинство известных видов динозавров.Одна из распространенных интерпретаций пластин на спинах стегозавров - это теплообменники для терморегуляции, поскольку пластины заполнены кровеносными сосудами, которые теоретически могут поглощать и рассеивать тепло. [111]
Это могло бы сработать для стегозавра с большими пластинами, такого как Стегозавр , но другие стегозавры, такие как Wuerhosaurus , Tuojiangosaurus и Kentrosaurus, обладали гораздо меньшими пластинами с площадью поверхности, имеющей сомнительную ценность для терморегуляции. Однако идея использования пластин стегозавра в качестве теплообменников в последнее время подвергается сомнению. [112]
Другие доказательства [ править ]
Эндотермия требует частого дыхания, что может привести к потере воды. У современных птиц и млекопитающих потеря воды ограничивается за счет вытягивания влаги из выдыхаемого воздуха покрытыми слизистой оболочкой дыхательных раковин, покрытыми тканями костными листками в полости носа. У некоторых динозавров есть обонятельные носовые раковины, используемые для обоняния, но ни у одного из них еще не были идентифицированы дыхательные носовые раковины. [113]
Поскольку эндотермия обеспечивает более совершенный нервно-мышечный контроль, а вещество мозга требует большого количества энергии для поддержания, некоторые предполагают, что увеличение размера мозга указывает на повышенную активность и, следовательно, на эндотермию. Коэффициент энцефализации (EQ) динозавров, показатель размера мозга, рассчитанный с использованием эндокастов мозга, варьируется в диапазоне от птичьего до рептилийного. Если использовать только EQ, целурозавры, по-видимому, были столь же активны, как и современные млекопитающие, в то время как тероподы и орнитоподы находятся где-то между млекопитающими и рептилиями, а другие динозавры напоминают рептилий. [113]
Исследование, опубликованное Роджером Сеймуром в 2013 году, еще больше подтвердило идею о том, что динозавры были эндотермическими животными. Изучив морских крокодилов, Сеймур обнаружил, что даже если их большие размеры могут обеспечивать стабильную и высокую температуру тела, во время активности экзотермический метаболизм крокодилов обеспечивает меньшие аэробные способности и генерирует только 14% общей мышечной силы эндотермного млекопитающего аналогичного размера до полной усталости. . Сеймур рассуждал, что динозавры должны были быть эндотермными, поскольку им требовались лучшие аэробные способности и более высокая выработка энергии, чтобы конкурировать и доминировать над млекопитающими как активными наземными животными на протяжении всей мезозойской эры. [114]
архозавров Ранний метаболизм
Этот раздел нуждается в дополнительных цитатах для проверки . ( сентябрь 2008 г. ) |
Похоже, что самые ранние динозавры обладали особенностями, которые составляют основу аргументов в пользу теплокровных динозавров, особенно прямостоячими конечностями. Возникает вопрос: «Как динозавры стали теплокровными?» Наиболее очевидные возможные ответы:
- «Их непосредственные предки ( архозавры ) были хладнокровными, а у динозавров очень рано в своей эволюции начала развиваться теплокровность». Это означает, что динозавры развили значительную степень теплокровности за очень короткое время, возможно, менее чем за 20 миллионов лет. Но у предков млекопитающих эволюция теплокровности, по-видимому, заняла гораздо больше времени, начиная с зарождения вторичного нёба примерно в начале средней перми. [115] и продолжалось, возможно, до появления волос около 164 миллионов лет назад в середине юрского периода. [116] ).
- «Непосредственные предки динозавров ( архозавры ) были, по крайней мере, достаточно теплокровными, и динозавры развивались дальше в этом направлении». Этот ответ поднимает две проблемы: ( А ) Ранняя эволюция архозавров до сих пор очень плохо изучена – большое количество особей и видов обнаружено с начала триаса , но только 2 вида известны из очень поздней перми ( Archosaurus rossicus и Protorosaurus) . спанери ); ( Б ) Крокодилы эволюционировали незадолго до динозавров и тесно с ними связаны, но хладнокровны (см. ниже).
Крокодилы представляют собой некоторую загадку, если рассматривать динозавров как активных животных с довольно постоянной температурой тела. Крокодилы развились незадолго до динозавров и, уступая птицам, являются ближайшими живыми родственниками динозавров, однако современные крокодилы хладнокровны. Это вызывает некоторые вопросы:
- Если динозавры были в значительной степени «теплокровными», то когда и как быстро в их линии появилась теплокровность?
- Современные крокодилы хладнокровны, но имеют несколько особенностей, связанных с теплокровностью. Как они приобрели эти качества?
Современные крокодилы хладнокровны, но могут передвигаться с выпрямленными конечностями и имеют несколько особенностей, обычно связанных с теплокровностью, поскольку они улучшают снабжение животных кислородом: [117]
- 4-камерное сердце. У млекопитающих и птиц сердце четырехкамерное. У рептилий, не являющихся крокодилами, трехкамерное сердце, которое менее эффективно, поскольку позволяет смешиваться насыщенной кислородом и лишенной кислорода крови и, следовательно, посылать часть лишенной кислорода крови в организм, а не в легкие. Сердце современных крокодилов четырехкамерное, но меньше по размеру по сравнению с размером тела и работает при более низком давлении, чем у современных млекопитающих и птиц. У них также есть обходной канал, который делает их функционально трехкамерными, когда они находятся под водой, что позволяет экономить кислород. [118]
- диафрагма , которая помогает дыханию.
- вторичное небо , позволяющее животному есть и дышать одновременно.
- механизм печеночно-поршневой для перекачивания легких. Это отличается от механизмов перекачивания легких млекопитающих и птиц, но похоже на то, что некоторые исследователи обнаружили у некоторых динозавров. [20] [22]
Так почему же естественный отбор отдал предпочтение этим особенностям, которые важны для активных теплокровных существ, но малопригодны для хладнокровных водных хищников, устраивающих засады, которые проводят большую часть своего времени, плавая в воде или лежа на берегах рек?

В конце 1980-х годов было высказано предположение, что крокодилы изначально были активными теплокровными хищниками и что их предки-архозавры были теплокровными. [89] [117] Совсем недавно исследования развития показали, что у эмбрионов крокодилов сначала развивается полностью четырехкамерное сердце, а затем развиваются модификации, которые заставляют их сердца функционировать как трехкамерные под водой. Используя принцип, согласно которому онтогенез повторяет филогению , исследователи пришли к выводу, что первоначальные крокодилы имели полностью 4-камерное сердце и, следовательно, были теплокровными, и что позже у крокодилов развился обходной путь, когда они снова стали хладнокровными водными хищниками, устраивающими засады. [119] [120]
Более поздние исследования костных структур архозавров и их влияния на скорость роста также показывают, что ранние архозавры имели довольно высокий уровень метаболизма и что триасовые предки крокодилов вернулись к более типичному «рептильному» уровню метаболизма. [71]
Если эта точка зрения верна, то развитие теплокровности у архозавров (достигающее своего пика у динозавров) и у млекопитающих заняло бы примерно одинаковое количество времени. Это также согласуется с ископаемыми свидетельствами:
- Самые ранние крокодиломорфы, например Terrestrisuchus , были стройными, длинноногими наземными хищниками.
- Прямые конечности появились довольно рано в эволюции архозавров , а у райзухий очень плохо приспособлены к какой-либо другой позе. [121]
См. также [ править ]
- Классификация динозавров
- Ренессанс динозавров
- Эволюция динозавров
- Эволюционная физиология
- Список динозавров
- Происхождение птиц
- Аргентинский черно-белый тегу # Теплокровность
Ссылки [ править ]
- ^ Jump up to: Перейти обратно: а б Лукас, Спенсер Г. (1999). Динозавры: Учебник (3-е изд.). McGraw-Hill Companies, Inc., стр. 3–9 . ISBN 978-0-07-303642-7 .
- ^ Фастовский Д.Э., Вейшампель Д.Б. (2005). «Теропода I: Природа красная в зубах и когтях». В Фастовский Д.Э., Вейшампель Д.Б. (ред.). Эволюция и вымирание динозавров (2-е изд.). Издательство Кембриджского университета. стр. 265–299. ISBN 978-0-521-81172-9 .
- ^ Кольбер, Э.Х.; Коулз, РБ; Богерт, CM (июнь 1947 г.). «Температурная устойчивость американского аллигатора и ее влияние на привычки, эволюцию и вымирание динозавров». Ежеквартальный обзор биологии . 22 (2): 145–146. дои : 10.1086/395712 .
- ^ Jump up to: Перейти обратно: а б Лукас, Спенсер Г. (2000). Динозавры: Учебник (3-е изд.). McGraw-Hill Companies, Inc., стр. 1–3 . ISBN 978-0-07-303642-7 .
- ^ Торренс, Хью (1997). «Политика и палеонтология». В Фарлоу, Джеймс О.; Бретт-Сурман, Майкл К. (ред.). Полный динозавр . Блумингтон: Издательство Университета Индианы. стр. 175–190 . ISBN 978-0-253-33349-0 .
- ^ Бентон, Майкл Дж. (2000). «Краткая история палеонтологии динозавров». У Пола, Грегори С. (ред.). Научно-американская книга о динозаврах . Нью-Йорк: Пресса Святого Мартина. стр. 10–44. ISBN 978-0-312-26226-6 .
- ^ Баккер, RT (1968). «Превосходство динозавров». Открытие . 3 (2): 11–22.
- ^ Баккер, RT (1987). «Возвращение танцующих динозавров» . В Черкасе, SJ; Олсон, ЕС (ред.). Динозавры прошлого и настоящего, том. Я. Вашингтонский университет Press. ISBN 978-0-295-96541-3 .
- ^ Jump up to: Перейти обратно: а б с д Баккер, Роберт Т. (1972). «Анатомические и экологические доказательства эндотермии у динозавров». Природа . 238 (5359): 81–85. Бибкод : 1972Natur.238...81B . дои : 10.1038/238081a0 . S2CID 4176132 .
- ^ Томас, РДК; Олсон, ЕС (1980). Холодный взгляд на теплокровных динозавров . Вествью Пресс. ISBN 978-0-89158-464-3 .
- ^ Бентон, MJ (2005). Палеонтология позвоночных (3-е изд.). Уайли-Блэквелл. стр. 221–223. ISBN 978-0-632-05637-8 .
- ^ Паладино, ФВ; О'Коннор, член парламента; Спотила, младший (1990). «Метаболизм кожистых черепах, гигантотермия и терморегуляция динозавров». Природа . 344 (6269): 858–860. Бибкод : 1990Natur.344..858P . дои : 10.1038/344858a0 . S2CID 4321764 .
- ^ Баррик, RE; Душ, штат Вашингтон; Фишер, АГ (1996). «Сравнение терморегуляции четырех птицетазовых динозавров и ящерицы-вараниды из меловой формации Два Медицина: данные по изотопам кислорода». ПАЛЕОС . 11 (4): 295–305. Бибкод : 1996Палай..11..295B . дои : 10.2307/3515240 . JSTOR 3515240 .
- ^ Jump up to: Перейти обратно: а б с д и Норман, Д.Б. (апрель 2001 г.). «Кормление динозавров» . Энциклопедия наук о жизни . Джон Уайли и сыновья. дои : 10.1038/npg.els.0003321 . ISBN 978-0470016176 . Проверено 10 сентября 2009 г.
- ^ Крылья, О. (2007). «Обзор функции гастролитов с последствиями для ископаемых позвоночных и пересмотренная классификация» (PDF) . Acta Palaeontologica Polonica . 52 (1): 1–16 . Проверено 10 сентября 2008 г.
- ^ Уингз, О. и Сандер. ПМ (март 2007 г.). «У динозавров-зауроподов нет желудочной мельницы: новые данные анализа массы и функции гастролита у страусов» . Труды Королевского общества Б. 274 (1610): 635–640. дои : 10.1098/rspb.2006.3763 . ПМК 2197205 . ПМИД 17254987 .
- ^ http://jeb.biologies.org/content/184/1/63.full.pdf [ пустой URL PDF ]
- ^ Jump up to: Перейти обратно: а б с д Ли, Эндрю Х.; Вернинг, Сара (2008). «Половая зрелость растущих динозавров не соответствует моделям роста рептилий» . Труды Национальной академии наук . 105 (2): 582–587. Бибкод : 2008PNAS..105..582L . дои : 10.1073/pnas.0708903105 . ПМК 2206579 . ПМИД 18195356 .
- ^ Швейцер, Миннесота; Виттмейер, Дж.Л.; Хорнер, младший (2005). «Полоспецифичная репродуктивная ткань бескилевых и тираннозавра рекса » (PDF) . Наука . 308 (5727): 1456–1460. Бибкод : 2005Sci...308.1456S . дои : 10.1126/science.1112158 . ПМИД 15933198 . S2CID 30264554 .
- ^ Jump up to: Перейти обратно: а б с Рубен, Дж.А.; Джонс, Т.Д.; Гейст, Северная Каролина; Хиллениус, WJ (ноябрь 1997 г.). «Структура легких и вентиляция легких тероподовых динозавров и ранних птиц». Наука . 278 (5341): 1267–1270. Бибкод : 1997Sci...278.1267R . дои : 10.1126/science.278.5341.1267 .
- ^ Jump up to: Перейти обратно: а б Рубен, Дж.А.; Джонс, Т.Д.; Гейст, Северная Каролина; Лейтч, А.; Хиллениус, WJ (1997). «Вентиляция легких и газообмен у тероподовых динозавров». Наука . 278 (5341): 1267–1270. Бибкод : 1997Sci...278.1267R . дои : 10.1126/science.278.5341.1267 .
- ^ Jump up to: Перейти обратно: а б Рубен, Дж.А.; Даль Сассо, К.; Гейст, Северная Каролина; Хиллениус, WJ; Джонс, Т.Д.; Синьор, М. (январь 1999 г.). «Функция легких и метаболическая физиология динозавров-теропод» (PDF) . Наука . 283 (5401): 514–516. Бибкод : 1999Sci...283..514R . дои : 10.1126/science.283.5401.514 . ПМИД 9915693 .
- ^ Рубен, Дж.А.; Джонс, Т.Д.; Гейст, NR (2003). «Дыхание и репродуктивная палеофизиология динозавров и ранних птиц» (PDF) . Физиол. Биохим. Зоол . 76 (2): 141–164. дои : 10.1086/375425 . hdl : 10211.1/1472 . ПМИД 12794669 . S2CID 222819 .
- ^ Хиллениус, В.Дж. и Рубен, Дж.А. (ноябрь – декабрь 2004 г.). «Эволюция эндотермии у наземных позвоночных: кто? Когда? Почему?». Физиологическая и биохимическая зоология . 77 (6): 1019–1042. дои : 10.1086/425185 . ПМИД 15674773 . S2CID 29300018 .
- ^ Хикс, Дж. У. и Фармер, К. Г. (ноябрь 1997 г.). «Легочная вентиляция и газообмен у теропод-динозавров» . Наука . 278 (5341): 1267–1270. Бибкод : 1997Sci...278.1267R . дои : 10.1126/science.278.5341.1267 .
- ^ Хикс, Дж. В. и Фармер, К. Г. (сентябрь 1999 г.). «Потенциал газообмена в легких рептилий: последствия для связи динозавров и птиц». Физиология дыхания . 117 (2–3): 73–83. дои : 10.1016/S0034-5687(99)00060-2 . ПМИД 10563436 .
- ^ Jump up to: Перейти обратно: а б Карри, П.Дж. и Чен, П.Дж. (декабрь 2001 г.). «Анатомия Sinosauropteryx prima из Ляонина, северо-восточный Китай» (PDF) . Канадский журнал наук о Земле . 38 (12): 1705–1727. Бибкод : 2001CaJES..38.1705C . doi : 10.1139/cjes-38-12-1705 .
- ^ Jump up to: Перейти обратно: а б О'Коннор П. и Классенс Л. (июль 2005 г.). «Базовая конструкция птичьих легких и проточная вентиляция у нептичьих динозавров-теропод». Природа . 436 (7048): 253–256. Бибкод : 2005Natur.436..253O . дои : 10.1038/nature03716 . ПМИД 16015329 . S2CID 4390587 .
- ^ Ведель, MJ (2003). «Пневматичность позвонков, воздушные мешки и физиология динозавров-зауроподов» . Палеобиология . 29 (2): 243–255. doi : 10.1666/0094-8373(2003)029<0243:VPASAT>2.0.CO;2 . S2CID 46619244 . Полный текст в настоящее время доступен на сайте «Findarticles.com: Пневматичность позвонков, воздушные мешки и физиология динозавров-зауроподов» . Палеобиология . 2003 г. и «Пневматичность позвонков, воздушные мешки и физиология динозавров-зауроподов» (PDF) . Архивировано из оригинала (PDF) 17 декабря 2008 г. Подробный анатомический анализ можно найти по адресу. Ведель, MJ (2003). «Эволюция пневматичности позвонков у динозавров-зауроподов» (PDF) . Журнал палеонтологии позвоночных . 23 (2): 344–357. doi : 10.1671/0272-4634(2003)023[0344:TEOVPI]2.0.CO;2 . S2CID 55884062 .
- ^ Jump up to: Перейти обратно: а б Ведель, MJ (июнь 2006 г.). «Происхождение посткраниальной пневматическости скелета у динозавров» (PDF) . Интегративная зоология . 1 (2): 80–85. дои : 10.1111/j.1749-4877.2006.00019.x . ПМИД 21395998 .
- ^ Нэйш, Д.; Мартилл, DM; Фрей, Э. (июнь 2004 г.). «Экология, систематика и биогеографические взаимоотношения динозавров, включая нового теропода, из формации Сантана (? Альб, ранний мел) Бразилии». Историческая биология . 16 (2–4): 57–70. CiteSeerX 10.1.1.394.9219 . дои : 10.1080/08912960410001674200 . S2CID 18592288 . Это также одна из нескольких тем, затронутых в сообщении в блоге Нэйша. «Базальные тираны-динозавры и моя любимица Миришия » . – обратите внимание, что Миришия была целурозавром , который, по мнению Нэйша, был тесно связан с Компсогнатом .
- ^ Jump up to: Перейти обратно: а б Серено П.С., Мартинес Р.Н., Уилсон Дж.А., Варриккио Д.Д., Алькобер О.А. и др. (сентябрь 2008 г.). Кемп Т. (ред.). «Доказательства существования птичьих внутригрудных воздушных мешков у нового хищного динозавра из Аргентины» . ПЛОС ОДИН . 3 (9): е3303. Бибкод : 2008PLoSO...3.3303S . дои : 10.1371/journal.pone.0003303 . ПМЦ 2553519 . ПМИД 18825273 .
- ^ Ведель, Мэтью (2007). «Что пневматическость говорит нам о «прозауроподах» и наоборот» (PDF) . Специальные статьи по палеонтологии . 77 : 207–222. Архивировано из оригинала (PDF) 5 июля 2008 года . Проверено 31 октября 2007 г.
- ^ Мэллисон, Х. (2010). «Цифровой Платеозавр II: оценка диапазона движений конечностей и позвоночного столба, а также предыдущих реконструкций с использованием цифрового скелетного крепления» . Acta Palaeontologica Polonica . 55 (3): 433–458. дои : 10.4202/app.2009.0075 .
- ^ Jump up to: Перейти обратно: а б с д и ж г час я дж Рид, REH (1997). «Физиология динозавров: аргументы в пользу «промежуточных» динозавров». В Фарлоу, Джо; Бретт-Сурман, депутат Кнессета (ред.). Полный динозавр . Блумингтон: Издательство Университета Индианы. стр. 449–473. ISBN 978-0-253-33349-0 . Проверено 12 сентября 2008 г.
- ^ Паладино, ФВ; Спотила, младший; Додсон, П. (1997). «Проект гигантов: моделирование физиологии крупных динозавров». В Фарлоу, Джо; Бретт-Сурман, депутат Кнессета (ред.). Полный динозавр . Блумингтон, Индиана: Издательство Университета Индианы. стр. 491–504. ISBN 978-0-253-21313-6 .
- ^ Уорд, Питер (2006) Из воздуха: динозавры, птицы и древняя атмосфера Земли, страницы 159–198, National Academies Press. ISBN 9780309141239 .
- ^ Кодд, младший; Мэннинг, Польша; Норелл, Массачусетс; Перри, Сан-Франциско (январь 2008 г.). «Птичье дыхание у динозавров-манирапторов» . Труды Королевского общества Б. 275 (1631): 157–161. дои : 10.1098/rspb.2007.1233 . ПМК 2596187 . ПМИД 17986432 . Сводка новостей на «Почему у динозавров было птичье дыхание» . 7 ноября 2007 г.
- ^ Щекотать, ПГ; Эннос, Арканзас; Леннокс, Ле; Перри, Сан-Франциско; Кодд, младший (ноябрь 2007 г.). «Функциональное значение крючковидных отростков птиц» . Журнал экспериментальной биологии . 210 (Часть 22): 3955–3961. дои : 10.1242/jeb.008953 . ПМИД 17981863 .
- ^ Фишер, ЧП; Рассел, Д.А.; Стоскопф, МК; Баррик, RE; Хаммер, М.; Кузьмиц А.А. (апрель 2000 г.). «Сердечно-сосудистые доказательства средней или более высокой скорости метаболизма у птицетазовых динозавров». Наука . 288 (5465): 503–505. Бибкод : 2000Sci...288..503F . дои : 10.1126/science.288.5465.503 . ПМИД 10775107 . Но обратите внимание, что основная тема этой статьи заключается в том, что окаменелость предоставила убедительные доказательства существования 4-камерного сердца, что не получило широкого признания.
- ^ Рубен, Дж.А.; Хиллениус, WJ; Гейст, Северная Каролина; Лейтч, А.; Джонс, Т.Д.; Карри, Пи Джей; Хорнер, младший; Эспе, Г. (август 1996 г.). «Метаболический статус некоторых динозавров позднего мела» (PDF) . Наука . 273 (5279): 1204–1207. Бибкод : 1996Sci...273.1204R . дои : 10.1126/science.273.5279.1204 . S2CID 84693210 .
- ^ Рубен, Дж. А. и Джонс, Т. Д. (2000). «Селективные факторы, связанные с происхождением меха и перьев» . Американский зоолог . 40 (4): 585–596. дои : 10.1093/icb/40.4.585 .
- ^ Банг, Б.Г. (1966). «Обонятельный аппарат Procellariiformes». Акта Анатомика . 65 (1): 391–415. дои : 10.1159/000142884 . ПМИД 5965973 .
- ^ Банг, Б.Г. (1971). «Функциональная анатомия обонятельной системы 23 отрядов птиц». Акта Анатомика . 79. 79 : 1–76. дои : 10.1159/isbn.978-3-318-01866-0 . ISBN 978-3-8055-1193-3 . ПМИД 5133493 .
- ^ Скотт, Дж. Х. (1954). «Теплорегулирующая функция слизистой оболочки носа». Журнал ларинологии и отологии . 68 (5): 308–317. дои : 10.1017/S0022215100049707 . ПМИД 13163588 . S2CID 32082759 .
- ^ Куломб, HN; Сэм Х. Риджуэй; Ш; Эванс, МЫ (1965). «Дыхательный водообмен у двух видов морских свиней». Наука . 149 (3679): 86–88. Бибкод : 1965Sci...149...86C . дои : 10.1126/science.149.3679.86 . ПМИД 17737801 . S2CID 38947951 .
- ^ Уитмер, LM (август 2001 г.). «Положение ноздрей у динозавров и других позвоночных и его значение для функции носа». Наука . 293 (5531): 850–853. CiteSeerX 10.1.1.629.1744 . дои : 10.1126/science.1062681 . ПМИД 11486085 . S2CID 7328047 .
- ^ «Исследование дыхания динозавров показывает, что носы улучшают обоняние и охлаждают мозг» .
- ^ Фишер, Пол Э.; Рассел, Дейл А.; Стоскопф, Майкл К.; Баррик, Риз Э.; Хаммер, Майкл; Кузьмиц, Эндрю А. (апрель 2000 г.). «Сердечно-сосудистые доказательства средней или более высокой скорости метаболизма у птицетазовых динозавров». Наука . 288 (5465): 503–505. Бибкод : 2000Sci...288..503F . дои : 10.1126/science.288.5465.503 . ПМИД 10775107 .
- ^ Роу, Тимоти; Макбрайд, Эрл Ф.; Серено, Пол К. (февраль 2001 г.). «Технический комментарий: динозавр с каменным сердцем» . Наука . 291 (5505): 783а. дои : 10.1126/science.291.5505.783a . ПМИД 11157158 .
- ^ Рассел, Дейл А .; Фишер, Пол Э.; Баррик, Риз Э.; Стоскопф, Майкл К. (февраль 2001 г.). «Ответ: динозавр с каменным сердцем» . Наука . 291 (5505): 783а. дои : 10.1126/science.291.5505.783a . ПМИД 11157158 .
- ^ Чинсами, Анусуя; и Хиллениус, Виллем Дж. (2004). «Физиология нептичьих динозавров». Динозаврия , 2-я. 643–659.
- ^ Карпентер, К.; Хирш, К.Ф.; Хорнер, младший (1994). "Введение". В Карпентере, К.; Хирш, К.Ф.; Хорнер, младший (ред.). Яйца динозавров и детеныши . Издательство Кембриджского университета. ISBN 978-0-521-56723-7 .
- ^ Эриксон, генеральный директор; Карри Роджерс, К.; Йерби, SA (июль 2001 г.). «Модели роста динозавров и быстрые темпы роста птиц». Природа . 412 (6845): 429–433. Бибкод : 2001Natur.412..429E . дои : 10.1038/35086558 . ПМИД 11473315 . S2CID 4319534 . Обратите внимание, что Кристина Роджерс также публиковала статьи под своей девичьей фамилией Кристина Карри.
- ^ Jump up to: Перейти обратно: а б Карри, штат Калифорния (1999). «Онтогенетическая гистология апатозавра (Dinosauria: Sauropoda): новые взгляды на темпы роста и продолжительность жизни» . Журнал палеонтологии позвоночных . 19 (4): 654–665. Бибкод : 1999JVPal..19..654C . дои : 10.1080/02724634.1999.10011179 . Архивировано из оригинала 15 июля 2010 года.
- ^ Jump up to: Перейти обратно: а б с Эриксон, генеральный директор; Маковицкий, П.Дж.; Карри, Пи Джей; Норелл, Массачусетс; Йерби, ЮАР; Брочу, Калифорния (август 2004 г.). «Гигантизм и сравнительные параметры жизненного цикла динозавров-тираннозаврид» (PDF) . Природа . 430 (7001): 772–775. Бибкод : 2004Natur.430..772E . дои : 10.1038/nature02699 . ПМИД 15306807 . S2CID 4404887 .
- ^ Хорнер, младший; Падиан, К. (сентябрь 2004 г.). «Возраст и динамика роста тираннозавра рекса» . Труды Королевского общества Б. 271 (1551): 1875–1880. дои : 10.1098/rspb.2004.2829 . ПМК 1691809 . ПМИД 15347508 .
- ^ Купер, Л.Н.; Ли, АХ; Конус, МЛ; Хорнер, младший (август 2008 г.). «Относительные темпы роста хищников и динозавров-жертв отражают последствия хищничества» . Труды Королевского общества Б. 275 (1651): 2609–2615. дои : 10.1098/rspb.2008.0912 . ПМК 2605812 . ПМИД 18682367 .
- ^ Эриксон, генеральный директор; Карри, Пи Джей; Иноуе, Б.Д.; Винн, А.А. (июль 2006 г.). «Таблицы жизни тираннозавров: пример биологии популяции нептичьих динозавров». Наука . 313 (5784): 213–217. Бибкод : 2006Sci...313..213E . дои : 10.1126/science.1125721 . ПМИД 16840697 . S2CID 34191607 .
- ^ Макгоуэн, К. (1991). Динозавры, Спитфайры и Морские Драконы . Издательство Гарвардского университета. стр. 133–135 . ISBN 978-0-674-20769-1 .
- ^ Маки, Род-Айленд (2002). «Мутуалистическое ферментативное пищеварение в желудочно-кишечном тракте: разнообразие и эволюция» . Интегративная и сравнительная биология . 42 (2): 319–326. дои : 10.1093/icb/42.2.319 . ПМИД 21708724 .
- ^ Пол, GS (1998). «Правило Коупа» . В Карпентере, К.; Фрейзер, Н.; Чуре, Д.; Киркланд, Дж.И. (ред.). Материалы симпозиума Моррисона: специальный выпуск журнала «Современная геология» . Тейлор и Фрэнсис. п. 211. ИСБН 978-90-5699-183-8 . Проверено 12 сентября 2008 г.
- ^ Риклс, AJ де. (1974). Эволюция эндотермии: гистологические данные . Эволюционная теория 1: 51–80.
- ^ Фастовский и Вейшампель 2009, стр.258.
- ^ Чинсами, А.; Рич, Т.; Викерс-Рич, П. (1998). «Гистология костей полярных динозавров» . Журнал палеонтологии позвоночных . 18 (2): 385–390. Бибкод : 1998JVPal..18..385C . дои : 10.1080/02724634.1998.10011066 . Архивировано из оригинала 15 июля 2010 года.
- ^ Клевезаль, Джорджия ; Мина, М.В.; Орешкин А.В. (1996). Регистрирующие структуры млекопитающих. Определение возраста и реконструкция истории жизни . ЦРК Пресс. ISBN 978-90-5410-621-0 .
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Фастовский и Вейшампель 2009, стр.260.
- ^ Энлоу, Д.Х. (1963). Принципы ремоделирования кости. Описание процессов постнатального роста и ремоделирования длинных костей и нижней челюсти . Спрингфилд, Иллинойс: CC Thomas.
- ^ Jump up to: Перейти обратно: а б Рид, REH (1984). «Первичная кость и физиология динозавров». Геологический журнал . 121 (6): 589–598. Бибкод : 1984GeoM..121..589R . дои : 10.1017/S0016756800030739 . S2CID 128981835 .
- ^ Рид, REH (1997). «Как росли динозавры». В Фарлоу, Джо; Бретт-Сурман, депутат Кнессета (ред.). Полный динозавр . Блумингтон: Издательство Университета Индианы. стр. 403–413 . ISBN 978-0-253-33349-0 .
- ^ Jump up to: Перейти обратно: а б де Риклес, А.; Падиан, К .; Нолл, Ф.; Хорнер, младший (апрель – июнь 2008 г.). «О происхождении высоких темпов роста у архозавров и их древних родственников: дополнительные гистологические исследования триасовых архозавроформ и проблема «филогенетического сигнала» в гистологии костей». Анналы палеонтологии . 94 (2): 57–76. Бибкод : 2008AnPal..94...57D . дои : 10.1016/j.annpal.2008.03.002 . Резюме также онлайн по адресу «Происхождение высоких темпов роста архозавров» . 31 мая 2008 года . Проверено 3 июня 2008 г.
- ^ Jump up to: Перейти обратно: а б Лежандр, Лукас Дж.; Генар, Гийом; Бота-Бринк, Дженнифер; Кубо, Хорхе (1 ноября 2016 г.). «Палеогистологические доказательства наследственной высокой скорости метаболизма у архозавров» . Систематическая биология . 65 (6): 989–996. doi : 10.1093/sysbio/syw033 . ISSN 1063-5157 . ПМИД 27073251 .
- ^ Сеймур, Роджер С.; Беннетт-Стампер, Кристина Л.; Джонстон, Соня Д.; Кэрриер, Дэвид Р.; Григг, Гордон К. (2004). «Доказательства эндотермических предков крокодилов, лежащих в основе эволюции архозавров» (PDF) . Физиологическая и биохимическая зоология . 77 (6): 1051–1067. дои : 10.1086/422766 . HDL : 2440/1933 . ПМИД 15674775 . S2CID 10111065 .
- ^ Сеймур, Роджер С. (1976). «Динозавры, эндотермия и кровяное давление». Природа . 262 (5565): 207–208. Бибкод : 1976Natur.262..207S . дои : 10.1038/262207a0 . S2CID 4157084 .
- ^ Сеймур, Роджер С.; Смит, Сара Л; Уайт, Крейг Р.; Хендерсон, Дональд М.; Шварц-Уингс, Даниэла (2012). «Приток крови к длинным костям свидетельствует об активности метаболизма у млекопитающих, рептилий и динозавров» . Труды Королевского общества Б. 279 (1728): 451–456. дои : 10.1098/rspb.2011.0968 . ПМЦ 3234558 . ПМИД 21733896 .
- ^ Пол, GS (1988). Хищные динозавры мира . Нью-Йорк: Саймон и Шустер. п. 155 . ISBN 978-0-671-61946-6 .
- ^ Jump up to: Перейти обратно: а б Гиллули, Дж. Ф.; Аллен, AP; Чарнов Е.Л. (август 2006 г.). «Окаменелости динозавров предсказывают температуру тела» . ПЛОС Биология . 4 (8): е248. doi : 10.1371/journal.pbio.0040248 . ПМЦ 1489189 . ПМИД 16817695 . Менее техническое резюме можно найти на странице Гросс, Л. (август 2006 г.). «Математика и окаменелости решают спор о метаболизме динозавров» . ПЛОС Биология . 4 (8): е255. doi : 10.1371/journal.pbio.0040255 . ПМК 1489188 . ПМИД 20076616 .
- ^ Маковяк, Пенсильвания; СС Вассерман; М. М. Левин (23 сентября 1992 г.). «Критическая оценка 98,6 ° F (37,0 ° C), верхнего предела нормальной температуры тела и других наследий Карла Рейнхольда Августа Вундерлиха». ДЖАМА . 268 (12): 1578–1580. дои : 10.1001/jama.1992.03490120092034 . ПМИД 1302471 .
- ^ Сунд-Левандер, Марта; Форсберг, Кристина; Варен, Лис Карин (2002). «Нормальная оральная, ректальная, барабанная и подмышечная температура тела у взрослых мужчин и женщин: систематический обзор литературы». Скандинавский журнал наук о заботе . 16 (2): 122–128. дои : 10.1046/j.1471-6712.2002.00069.x . ПМИД 12000664 .
- ^ Эриксон, Грегори М.; Роджерс, Кристина Карри; Йерби, Скотт А. (2001). «Модели роста динозавров и быстрые темпы роста птиц». Природа . 412 (6845): 429–433. Бибкод : 2001Natur.412..429E . дои : 10.1038/35086558 . ПМИД 11473315 . S2CID 4319534 .
- ^ Карри, Кристина А. (1999). «Онтогенетическая гистология апатозавра (Dinosauria: Sauropoda): новые сведения о темпах роста и продолжительности жизни». Журнал палеонтологии позвоночных . 19 (4): 654–665. Бибкод : 1999JVPal..19..654C . дои : 10.1080/02724634.1999.10011179 .
- ^ Фастовский, Дэвид Э.; Вейшампель, Дэвид Б. (2009). Динозавры: краткая естественная история (PDF) . п. 258.
- ^ Баррик, Р.Э. и Шауэрс, У.Дж. (1999). «Термофизиология и биология гиганотозавра : сравнение с тираннозавром » . Электронная палеонтология . 2 (2) . Проверено 10 сентября 2008 г.
- ^ Баррик, RE; Душ, штат Вашингтон; Фишер, АГ (август 1996 г.). «Сравнение терморегуляции четырех птицетазовых динозавров и ящерицы-вараниды из меловой формации Два Медицина: данные по изотопам кислорода». ПАЛЕОС . 11 (4): 295–305. Бибкод : 1996Палай..11..295B . дои : 10.2307/3515240 . JSTOR 3515240 .
- ^ Баккер, RT (сентябрь 1974 г.). «Биоэнергетика динозавров - ответ Беннетту, Далзеллу и Федуччиа». Эволюция . 28 (3): 497–503. дои : 10.2307/2407178 . JSTOR 2407178 . ПМИД 28564852 .
- ^ Фастовский, Д.Э.; Вейшампель, Д.Б.; Сиббик, Дж. (2005). Эволюция и вымирание динозавров . Издательство Кембриджского университета. ISBN 978-0-521-81172-9 .
- ^ Фарлоу, Дж. О. (1980). «Соотношение биомассы хищника и жертвы, пищевые сети сообщества и физиология динозавров» (PDF) . В Томасе, РДК; Олсон, ЕС (ред.). Холодный взгляд на теплокровных динозавров . Боулдер, Колорадо: Американская ассоциация содействия развитию науки. стр. 55–83. Архивировано из оригинала (PDF) 17 декабря 2008 года.
- ^ Это было признано не позднее 1909 года: «Доктор Холланд и расползающиеся зауроподы» . Архивировано из оригинала 12 июня 2011 года. Аргументы и многие изображения также представлены в Десмонд, А. (1976). Горячие динозавры . ДаблДэй. ISBN 978-0-385-27063-2 .
- ^ Jump up to: Перейти обратно: а б Кэрриер, ДР (1987). «Эволюция двигательной выносливости четвероногих: обход механических ограничений». Палеобиология . 13 (3): 326–341. Бибкод : 1987Pbio...13..326C . дои : 10.1017/S0094837300008903 . JSTOR 2400734 . S2CID 83722453 .
- ^ Фастовский и Вейшампель 2009, стр.251.
- ^ Фастовский и Вейшампель 2009, стр.252.
- ^ Остром, Дж. Х. (1980). «Свидетельства эндотермии у динозавров» (PDF) . В Томасе, РДК; Олсон, ЕС (ред.). Холодный взгляд на теплокровных динозавров . Боулдер, Колорадо: Американская ассоциация содействия развитию науки. стр. 82–105. Архивировано из оригинала (PDF) 27 мая 2008 года.
- ^ Понцер, Х. .; Аллен, В. .; Хатчинсон, Дж. .; Фарке, А.А. (2009). Фарке, Эндрю Аллен (ред.). «Биомеханика бега указывает на эндотермию у прямоногих динозавров» . ПЛОС ОДИН . 4 (11): е7783. Бибкод : 2009PLoSO...4.7783P . дои : 10.1371/journal.pone.0007783 . ПМЦ 2772121 . ПМИД 19911059 .
- ^ Швейцер, Миннесота; Дж. А. Ватт; Р. Авчи; Л. Кнапп; Л. Кьяппе; М. Норелл; М. Маршалл. (июль 1999 г.). «Специфическая иммунологическая реактивность бета-кератина в перьевых структурах мелового альваресзавриды Shuvuuia Deserti » . Журнал экспериментальной зоологии . 285 (2): 146–157. doi : 10.1002/(SICI)1097-010X(19990815)285:2<146::AID-JEZ7>3.0.CO;2-A . ПМИД 10440726 . Архивировано из оригинала 10 декабря 2012 года.
- ^ Сюй С.; Норелл, Массачусетс; Куанг Х.; Ван Х.; Чжао Ц. и Цзя К. (2004). «Базальные тираннозавроиды из Китая и свидетельства существования протоперьев у тираннозавроидов» (PDF) . Природа . 431 (7009): 680–684. Бибкод : 2004Natur.431..680X . дои : 10.1038/nature02855 . ПМИД 15470426 . S2CID 4381777 .
- ^ Бентон, Майкл Дж.; Дуайи, Даниэль; Цзян, Баоюй; Макнамара, Мария (1 сентября 2019 г.). «Раннее происхождение перьев» . Тенденции в экологии и эволюции . 34 (9): 856–869. дои : 10.1016/j.tree.2019.04.018 . hdl : 10468/8068 . ISSN 0169-5347 . ПМИД 31164250 . S2CID 174811556 .
- ^ Барретт, Пол М.; Эванс, Дэвид С.; Кампионе, Николас Э. (30 июня 2015 г.). «Эволюция эпидермальных структур динозавров» . Письма по биологии . 11 (6): 20150229. doi : 10.1098/rsbl.2015.0229 . ПМЦ 4528472 . ПМИД 26041865 .
- ^ Бонапарт, Дж. Ф.; Новас, Э.Э.; Кориа, РА (1990). « Carnotaurus sastrei Bonaparte, рогатый, легкого телосложения карнозавр из среднего мела Патагонии» . Вклад Музея естественной истории округа Лос-Анджелес в науку . 416 : 1–42. Архивировано из оригинала 10 мая 2008 года.
- ^ Доктор философии, Андреа Кау (1 марта 2016 г.). «Теропода: постмодернистские мезозойские мифы и легенды о динозаврах: Чешуя Карнотавра» . Теропода . Проверено 25 апреля 2020 г.
- ^ Jump up to: Перейти обратно: а б с Пол, Грегори С. (2016). Принстонский полевой справочник по динозаврам - 2-е издание . Издательство Принстонского университета. ISBN 978-0-691-16766-4 .
- ^ Черкас, С.А. (1994). Локли, МГ; дос Сантос, В.Ф.; Мейер, Калифорния; Хант, AP (ред.). «История и интерпретация отпечатков кожи зауроподов». Гайя . 10 . Лиссабон, Португалия.
- ^ Додсон П. и Барлоу В.Д. (1996). Рогатые динозавры: естественная история . Издательство Принстонского университета. ISBN 978-0-691-05900-6 . См. также изображение на «Кожа цератопса» . Архивировано из оригинала 3 февраля 2014 года.
- ^ Гослайн, А. (16 ноября 2004 г.). «Раскрыта «пуленепробиваемая» броня динозавров» . Новый новостной сервис Scientist.com.
- ^ Jump up to: Перейти обратно: а б с д Рич, TH; Викерс-Рич, П.; Ганглофф, Р.А. (февраль 2002 г.). «Полярные динозавры» . Наука . 295 (5557): 979–980. дои : 10.1126/science.1068920 . ПМИД 11834803 . S2CID 28065814 . См. также Викерс-Рич, П. и Рич, TH (2004). «Динозавры Антарктики» (PDF) . Научный американец. Архивировано из оригинала (PDF) 17 мая 2008 г. и Фиорилло, Арканзас (2004). «Динозавры Арктической Аляски» (PDF) . Научный американец . 291 (6): 84–91. Бибкод : 2004SciAm.291f..84F . doi : 10.1038/scientificamerican1204-84 . ПМИД 15597984 . Архивировано из оригинала (PDF) 17 мая 2008 года.
- ^ Хоттон, Н. (1980). «Альтернатива эндотермии динозавров: счастливые странники». В Томасе, РДК; Олсон, ЕС (ред.). Холодный взгляд на теплокровных динозавров . Боулдер, Колорадо: Американская ассоциация содействия развитию науки. стр. 311–350. ISBN 978-0-89158-464-3 .
- ^ Пол, GS (1988). Хищные динозавры мира . Нью-Йорк: Саймон и Шустер. стр. 160–161 . ISBN 978-0-671-61946-6 .
- ^ Белл, Фил Р.; Снивли, Э. (2008). «Полярные динозавры на параде: обзор миграции динозавров» . Алчеринга . 32 (3): 271–284. дои : 10.1080/03115510802096101 .
- ^ Ллойд, Робин (4 декабря 2008 г.). «Полярные динозавры пережили холодные темные зимы» . LiveScience.com . Имагинова . Проверено 11 декабря 2008 г.
- ^ Чинсами, А.; Рич, TH; Викерс-Рич, П. (1998). «Гистология костей полярных динозавров». Журнал палеонтологии позвоночных . 18 (2): 385–390. Бибкод : 1998JVPal..18..385C . дои : 10.1080/02724634.1998.10011066 . JSTOR 4523908 . См. также Лесли, М. (декабрь 2007 г.). «Измерение температуры у динозавра» . Смитсоновский журнал. [ постоянная мертвая ссылка ]
- ^ Вудворд, Холли Н.; Рич, Томас Х.; Чинсами, Анусуя; Викерс-Рич, Патрисия (2011). «Динамика роста полярных динозавров Австралии» . ПЛОС ОДИН . 6 (8): e23339. Бибкод : 2011PLoSO...623339W . дои : 10.1371/journal.pone.0023339 . ПМК 3149654 . ПМИД 21826250 .
- ^ де Буфрениль, В.; Фарлоу, Джо; де Риклс, А. (1986). «Рост и функция пластин стегозавра : данные гистологии костей». Палеобиология . 12 (4): 459–473. Бибкод : 1986Pbio...12..459D . дои : 10.1017/S0094837300003171 . JSTOR 2400518 . S2CID 89139852 .
- ^ «Пластины и шипы стегозавра только для красоты» . 18 мая 2005 г. Архивировано из оригинала 27 сентября 2011 г.
- ^ Jump up to: Перейти обратно: а б Фастовский и Вейшампель 2009, стр.255.
- ^ Сеймур Роджер С. (5 июля 2013 г.). «Максимальная аэробная и анаэробная выработка энергии у крупных крокодилов по сравнению с млекопитающими: последствия гигантотермии динозавров» . ПЛОС ОДИН . 8 (7): e69361. Бибкод : 2013PLoSO...869361S . дои : 10.1371/journal.pone.0069361 . ПМК 3702618 . ПМИД 23861968 .
- ^ Кермак, Д.М. и Кермак, К.А. (1984). Эволюция характеров млекопитающих . Лондон: Издательство Крум Хелм Капитан Сабо. п. 149. ИСБН 978-0-7099-1534-8 .
- ^ Самые ранние явные свидетельства наличия волос или меха встречаются в окаменелостях Castorocauda , датированных 164 миллионами лет назад в середине юрского периода . Джи, К.; Луо, ZX; Юань, CX; Табрум, Арканзас (февраль 2006 г.). «Плавательная форма млекопитающих средней юры и экоморфологическая диверсификация ранних млекопитающих» . Наука . 311 (5764): 1123–7. Бибкод : 2006Sci...311.1123J . дои : 10.1126/science.1123026 . ПМИД 16497926 . S2CID 46067702 . См. также новость на сайте «Найден юрский бобр; переписывает историю млекопитающих» . Архивировано из оригинала 3 марта 2006 года. Самое позднее с 1950-х годов утверждалось, что могут быть свидетельства наличия волос у раннего триаса, цинодонтов таких как Thrinaxodon : Бринк, А.С. (1955). «Исследование скелета Диадемодона ». Палеонтология Африканская . 3 :3–39. и Кемп, Т.С. (1982). Млекообразные рептилии и происхождение млекопитающих . Лондон: Академическая пресса. п. 363 . ISBN 978-0-12-404120-2 . но отверстия (небольшие туннели) в мордовых костях цинодонтов являются в лучшем случае неоднозначным свидетельством, поскольку подобные отверстия встречаются у некоторых ныне живущих рептилий: Эстес, Р. (1961). «Краниальная анатомия рептилии-цинодонта Thrinaxodon liorhinus ». Бюллетень Музея сравнительной зоологии (1253 г.): 165–180. и Рубен, Дж. А. и Джонс, Т. Д. (2000). «Селективные факторы, связанные с происхождением меха и перьев» . Американский зоолог . 40 (4): 585–596. дои : 10.1093/icb/40.4.585 .
- ^ Jump up to: Перейти обратно: а б Макгоуэн, К. (1991). Динозавры, Спитфайры и Морские Драконы . Издательство Гарвардского университета. стр. 151–152 . ISBN 978-0-674-20769-1 .
- ^ «Тайна крокодильего сердца» . Австралийская радиовещательная корпорация .
- ^ Саммерс, AP (2005). «Эволюция: Добросердечные крокодилы» . Природа . 434 (7035): 833–834. Бибкод : 2005Natur.434..833S . дои : 10.1038/434833а . ПМИД 15829945 . S2CID 4399224 .
- ^ Сеймур, RS; Беннетт-Стампер, CL; Джонстон, Южная Дакота; Перевозчик, ДР; Григг, GC (2004). «Доказательства эндотермических предков крокодилов, лежащих в основе эволюции архозавров» (PDF) . Физиологическая и биохимическая зоология . 77 (6): 1051–1067. дои : 10.1086/422766 . HDL : 2440/1933 . ПМИД 15674775 . S2CID 10111065 . См. также объяснение этого вопроса с полезными иллюстрациями по адресу: Майерс, П.З. (19 апреля 2005 г.). «Горячие крокодилы?» . Архивировано из оригинала 16 февраля 2008 года.
- ^ Хатчинсон, младший (март – апрель 2006 г.). «Эволюция передвижения архозавров» (PDF) . Comptes Рендус Палевол . 5 (3–4): 519–530. Бибкод : 2006CRPal...5..519H . дои : 10.1016/j.crpv.2005.09.002 .
Внешние ссылки [ править ]
- Термофизиология и биология гиганотозавра: сравнение с тираннозавром Р.Э. Барриком и У.Дж. Шауэрсом (1999)
- Сообщается, что найдено сердце динозавра
- Эволюция крокодила не согревает сердца
физиологии Типы | ||
|---|---|---|
| Животные |  | |
| Растения | ||
| Клетки | ||
| Связанные темы | ||