Системная биология
| Сложные системы |
|---|
| Темы |

Системная биология — это вычислительный и математический анализ и моделирование сложных биологических систем . Это междисциплинарная область исследований, основанная на биологии , которая фокусируется на сложных взаимодействиях внутри биологических систем с использованием целостного подхода ( холизма вместо более традиционного редукционизма ) к биологическим исследованиям. [1]
В частности, начиная с 2000 года, эта концепция широко использовалась в биологии в самых разных контекстах. Проект «Геном человека» является примером прикладного системного мышления в биологии, которое привело к новым совместным способам работы над проблемами в области биологической генетики. [2] Одной из целей системной биологии является моделирование и обнаружение эмерджентных свойств , свойств клеток , тканей и организмов, функционирующих как система , теоретическое описание которых возможно только с использованием методов системной биологии. [1] [3] Обычно они включают метаболические сети или клеток . сигнальные сети [1] [4]
Обзор [ править ]
Этот раздел написан как личное размышление, личное эссе или аргументативное эссе , в котором излагаются личные чувства редактора Википедии или представлены оригинальные аргументы по определенной теме. ( декабрь 2022 г. ) |
Системную биологию можно рассматривать с нескольких различных аспектов.
В качестве области исследования, в частности, изучение взаимодействий между компонентами биологических систем и того, как эти взаимодействия приводят к функционированию и поведению этой системы (например, ферменты и метаболиты в метаболическом пути или сердцебиение). ). [5] [6] [7]
Как парадигму системную биологию обычно определяют в противоположность так называемой редукционистской парадигме ( биологической организации ), хотя она согласуется с научным методом . Различие между двумя парадигмами упоминается в этих цитатах: « Редукционистский подход успешно определил большинство компонентов и многие взаимодействия, но, к сожалению, не предлагает убедительных концепций или методов для понимания того, как возникают свойства системы... плюрализм Причины и следствия в биологических сетях лучше решать путем наблюдения с помощью количественных измерений за несколькими компонентами одновременно и путем строгой интеграции данных с математическими моделями». (Зауэр и др. ) [8] «Системная биология... занимается объединением, а не разъединением, интеграцией, а не редукцией. Она требует, чтобы мы разработали способы мышления об интеграции, которые были бы такими же строгими, как наши редукционистские программы, но отличались бы... Это означает изменение нашей философии. в полном смысле этого слова». ( Денис Ноубл ) [7]
В качестве серии операционных протоколов , используемых для проведения исследований, а именно цикла, состоящего из теории, аналитического или компьютерного моделирования для предложения конкретных проверяемых гипотез о биологической системе, экспериментальной проверки, а затем использования вновь полученного количественного описания клеток или клеточных процессов для уточнения. вычислительная модель или теория. [9] Поскольку целью является модель взаимодействий в системе, экспериментальные методы, которые больше всего подходят для системной биологии, являются общесистемными и стараются быть как можно более полными. Поэтому транскриптомика , метаболомика , протеомика и высокопроизводительные методы используются для сбора количественных данных для построения и проверки моделей. [10]
Как приложение теории динамических систем к молекулярной биологии . Действительно, сосредоточенность на динамике изучаемых систем является основным концептуальным отличием системной биологии от биоинформатики . [11]
Как социально-научное явление, определяемое стратегией интеграции сложных данных о взаимодействиях в биологических системах из различных экспериментальных источников с использованием междисциплинарных инструментов и персонала. [12]
История [ править ]
Хотя концепция системного взгляда на клеточные функции была хорошо понята, по крайней мере, с 1930-х годов, [13] технологические ограничения затрудняли проведение общесистемных измерений. Появление технологии микрочипов в 1990-х годах открыло совершенно новые возможности для изучения клеток на системном уровне. В 2000 году в Сиэтле был основан Институт системной биологии с целью привлечь людей «вычислительного» типа, которых, как считалось, не привлекала академическая среда университета. У института не было четкого определения того, чем на самом деле была эта область: примерное объединение людей из разных областей для использования компьютеров для целостного изучения биологии новыми способами. [14] Кафедра системной биологии Гарвардской медицинской школы была открыта в 2003 году. [15] В 2006 году было предсказано, что ажиотаж, вызванный «очень модной» новой концепцией, приведет к тому, что всем крупным университетам понадобятся факультеты системной биологии, таким образом, появится карьера, доступная для выпускников с хоть какими-то способностями в компьютерном программировании и биологии. [14] В 2006 году Национальный научный фонд поставил задачу построить математическую модель всей клетки. [ нужна ссылка ] первая цельноклеточная модель Mycoplasmagentium В 2012 году в лаборатории Карра Медицинской школы Маунт-Синай в Нью-Йорке была создана . Цельноклеточная модель способна предсказать жизнеспособность клеток M. Genitalium в ответ на генетические мутации. [16]
Более ранним предшественником системной биологии, как отдельной дисциплины, возможно, был системный теоретик Михайло Месарович в 1966 году на международном симпозиуме в Технологическом институте Кейса в Кливленде , штат Огайо, под названием «Теория систем и биология» . Месарович предсказал, что, возможно, в будущем появится такая вещь, как «системная биология». [17] [18] Другими ранними предшественниками, которые придерживались мнения, что биологию следует анализировать как систему, а не как простой набор частей, были «Анализ метаболического контроля» , разработанный Хенриком Качером и Джимом Бернсом. [19] позже тщательно переработанный, [20] и Рейнхарт Генрих и Том Рапопорт , [21] и теория биохимических систем, разработанная Майклом Сэвиджо. [22] [23] [24]
По словам Роберта Розена в 1960-х годах, к началу 20-го века целостная биология ушла в прошлое, поскольку стала популярной эмпирическая наука, в которой доминировала молекулярная химия. [18] Вторя ему сорок лет спустя, в 2006 году, Клинг пишет, что успех молекулярной биологии на протяжении 20-го века подавил целостные вычислительные методы. [14] К 2011 году Национальные институты здравоохранения выделили гранты на поддержку более десяти центров системной биологии в Соединенных Штатах. [25] но к 2012 году Хантер пишет, что системной биологии еще предстоит что-то сделать, чтобы полностью раскрыть свой потенциал. Тем не менее, сторонники надеялись, что в будущем это может оказаться более полезным. [26]
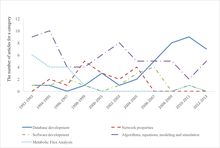
Важной вехой в развитии системной биологии стал международный проект Physiome . [ нужна ссылка ]
Связанные дисциплины [ править ]
Согласно интерпретации системной биологии как использования больших наборов данных с использованием междисциплинарных инструментов, типичным применением является метаболомика , которая представляет собой полный набор всех продуктов метаболизма, метаболитов , в системе на уровне организма, клетки или ткани. [28]
Элементы, которые могут представлять собой компьютерную базу данных, включают: феномику организма , вариации фенотипа по мере его изменения в течение его жизни; геномика , последовательность дезоксирибонуклеиновой кислоты (ДНК) организма, включая внутриклеточные специфичные вариации. (т.е. теломер изменение длины ); эпигеномика / эпигенетика , организменные и соответствующие клеточно-специфичные транскриптомные регулирующие факторы, не закодированные эмпирически в геномной последовательности. (т.е. метилирование ДНК , ацетилирование и деацетилирование гистонов и т.д.); транскриптомика в организмах, тканях или целых клетках экспрессии генов , измерения с помощью микрочипов ДНК или серийный анализ экспрессии генов ; интерферомика , корректирующие факторы транскрипта на организменном, тканевом или клеточном уровне (т.е. РНК-интерференция ), протеомика , измерения белков и пептидов на организменном, тканевом или клеточном уровне с помощью двумерного гель-электрофореза , масс-спектрометрии или методов многомерной идентификации белков. (современные ВЭЖХ системы в сочетании с масс-спектрометрией ). Поддисциплины включают в себя фосфопротеомика , гликопротеомика и другие методы обнаружения химически модифицированных белков; гликомические , организменные, тканевые или клеточные измерения углеводов ; липидомика на организменном, тканевом или клеточном уровне , измерение липидов . [ нужна ссылка ]
Также изучаются молекулярные взаимодействия внутри клетки, это называется интерактомикой . [29] Дисциплиной в этой области исследований являются белок-белковые взаимодействия , хотя интерактомика включает взаимодействия других молекул. [ нужна ссылка ] Нейроэлектродинамика , где вычислительная функция компьютера или мозга как динамической системы изучается наряду с его (био)физическими механизмами; [30] и флюксомика — измерение скорости метаболических реакций в биологической системе (клетке, ткани или организме). [28]
При подходе к проблеме системной биологии существует два основных подхода. Это подходы «сверху вниз» и «снизу вверх». Подход «сверху вниз» учитывает как можно большую часть системы и во многом опирается на экспериментальные результаты. Метод RNA-Seq является примером экспериментального подхода «сверху вниз». И наоборот, подход «снизу вверх» используется для создания подробных моделей с включением экспериментальных данных. Примером подхода «снизу вверх» является использование схемных моделей для описания простой генной сети. [31]
Различные технологии, используемые для отслеживания динамических изменений в мРНК, белках и посттрансляционных модификациях. Механобиология , силы и физические свойства во всех масштабах, их взаимодействие с другими регуляторными механизмами; [32] биосемиотика , анализ системы знаковых отношений организма или других биосистем; Физиомика — систематическое изучение физиомы в биологии.
Системная биология рака является примером подхода системной биологии, который можно отличить по конкретному объекту исследования ( онкогенез и лечение рака ). Он работает с конкретными данными (образцы пациентов, данные высокой пропускной способности с особым вниманием к характеристикам генома рака в образцах опухолей пациентов) и инструментами (иммортализованные линии раковых клеток , мышиные модели онкогенеза, модели ксенотрансплантатов , методы высокопроизводительного секвенирования , миРНК- на основе нокдауна генов высокопроизводительные скрининги , компьютерное моделирование последствий соматических мутаций и нестабильности генома ). [33] Долгосрочной целью системной биологии рака является способность лучше диагностировать рак, классифицировать его и лучше прогнозировать результат предлагаемого лечения, что является основой персонализированной медицины рака и виртуального онкологического пациента в более отдаленной перспективе. Значительные усилия в вычислительной системной биологии рака были предприняты для создания реалистичных многомасштабных in silico моделей различных опухолей. [34]
Подход системной биологии часто предполагает разработку механистических моделей, таких как реконструкция динамических систем на основе количественных свойств их элементарных строительных блоков. [35] [36] [37] [38] Например, сотовую сеть можно смоделировать математически, используя методы химической кинетики. [39] и теория управления . Из-за большого количества параметров, переменных и ограничений в сотовых сетях часто используются численные и вычислительные методы (например, анализ баланса потоков ). [37] [39]
Биоинформатика и анализ данных [ править ]
Другие аспекты информатики, информатики и статистики также используются в системной биологии. К ним относятся новые формы вычислительных моделей, такие как использование исчисления процессов для моделирования биологических процессов (известные подходы включают стохастическое π-исчисление , BioAmbients, Beta Binders, BioPEPA и исчисление Брана) и ограничений моделирование на основе ; интеграция информации из литературы с использованием методов извлечения информации и интеллектуального анализа текста ; [40] разработка онлайн-баз данных и хранилищ для обмена данными и моделями, подходов к интеграции баз данных и совместимости программного обеспечения посредством слабой связи программного обеспечения, веб-сайтов и баз данных или коммерческих костюмов; сетевые подходы для анализа многомерных наборов геномных данных. Например, анализ сети взвешенной корреляции часто используется для идентификации кластеров (называемых модулями), моделирования взаимосвязей между кластерами, расчета нечетких показателей членства в кластере (модуле), идентификации внутримодульных узлов и для изучения сохранения кластеров в других наборах данных; методы анализа омических данных, основанные на путях, например, подходы к идентификации и оценке путей с различной активностью их генов, белков или метаболитов. [41] Большая часть анализа наборов геномных данных также включает выявление корреляций. Кроме того, поскольку большая часть информации поступает из разных областей, необходима разработка синтаксически и семантически обоснованных способов представления биологических моделей. [42]
Создание биологических моделей [ править ]
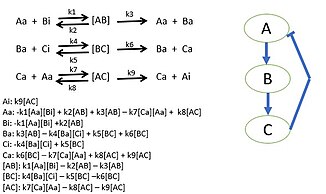
Исследователи начинают с выбора биологического пути и построения диаграмм всех белковых, генных и/или метаболических путей. После определения всех взаимодействий кинетика действующих масс или законы кинетической скорости ферментов для описания скорости реакций в системе используются . Используя закон сохранения массы, можно построить дифференциальные уравнения биологической системы. Можно провести эксперименты или подобрать параметры, чтобы определить значения параметров, которые будут использоваться в дифференциальных уравнениях . [44] Эти значения параметров будут различными кинетическими константами, необходимыми для полного описания модели. Эта модель определяет поведение видов в биологических системах и дает новое представление о конкретной деятельности системы. Иногда невозможно собрать все скорости реакций системы. Неизвестные скорости реакции определяются путем моделирования модели известных параметров и целевого поведения, которая обеспечивает возможные значения параметров. [45] [43]
Использование методов реконструкции и анализа на основе ограничений (COBRA) стало популярным среди системных биологов для моделирования и прогнозирования метаболических фенотипов с использованием моделей масштаба генома. Одним из методов является подход анализа баланса потоков (FBA), с помощью которого можно изучать биохимические сети и анализировать поток метаболитов через определенную метаболическую сеть путем оптимизации интересующей целевой функции (например, максимизации производства биомассы для прогнозирования роста). . [46]

См. также [ править ]
- Уравнение биохимических систем
- Биологические вычисления
- БиоСистемы (журнал)
- Вычислительная биология
- Экспосома
- Интерактом
- Список тем омики по биологии
- Список программного обеспечения для моделирования системной биологии
- Живые системы
- Анализ метаболического контроля
- Моделирование метаболической сети
- Моделирование биологических систем
- Молекулярная патологическая эпидемиология
- Сетевая биология
- Сетевая медицина
- Синтетическая биология
- Системная биомедицина
- Системная иммунология
- Системная медицина
- ТИАРА (база данных)
Ссылки [ править ]
- ^ Jump up to: Перейти обратно: а б с Тавассоли, Иман; Гольдфарб, Джозеф; Айенгар, Рави (04 октября 2018 г.). «Букварь по системной биологии: основные методы и подходы». Очерки по биохимии . 62 (4): 487–500. дои : 10.1042/EBC20180003 . ISSN 0071-1365 . ПМИД 30287586 . S2CID 52922135 .
- ^ Зеваил, Ахмед (2008). Физическая биология: от атомов к медицине . Издательство Имперского колледжа. п. 339.
- ^ Лонго, Джузеппе; Монтевиль, Маэль (2014). Перспективы организмов — Спрингер . Конспект лекций по морфогенезу. дои : 10.1007/978-3-642-35938-5 . ISBN 978-3-642-35937-8 . S2CID 27653540 .
- ^ Бу Зи, Callaway DJ (2011). «Белки ДВИГАЮТСЯ! Динамика белков и дальняя аллостерия в передаче сигналов в клетках». Структура белка и болезни . Достижения в области химии белков и структурной биологии. Том. 83. стр. 163–221. дои : 10.1016/B978-0-12-381262-9.00005-7 . ISBN 978-0-123-81262-9 . ПМИД 21570668 .
- ^ Снуп, Джеки Л; Вестерхофф, Ганс В. (2005). «От изоляции к интеграции: подход системной биологии к созданию кремниевой ячейки». В Альбергине, Лилия; Вестерхофф, Ганс В. (ред.). Системная биология: определения и перспективы . Темы современной генетики. Том. 13. Берлин: Шпрингер-Верлаг. стр. 13–30. дои : 10.1007/b106456 . ISBN 978-3-540-22968-1 .
- ^ «Системная биология: наука XXI века» . Институт системной биологии . Проверено 15 июня 2011 г.
- ^ Jump up to: Перейти обратно: а б Благородный, Денис (2006). Музыка жизни: Биология за пределами генома . Оксфорд: Издательство Оксфордского университета. п. 176. ИСБН 978-0-19-929573-9 .
- ^ Зауэр, Уве; Хайнеманн, Матиас; Замбони, Никола (27 апреля 2007 г.). «Генетика: приближаясь к полной картине» . Наука . 316 (5824): 550–551. дои : 10.1126/science.1142502 . ПМИД 17463274 . S2CID 42448991 .
- ^ Холоденко Борис Н; Сауро, Герберт М (2005). «Механистический и модульный подходы к моделированию и выводу клеточных регуляторных сетей». В Альбергине, Лилия; Вестерхофф, Ганс В. (ред.). Системная биология: определения и перспективы . Темы современной генетики. Том. 13. Берлин: Шпрингер-Верлаг. стр. 357–451. дои : 10.1007/b136809 . ISBN 978-3-540-22968-1 .
- ^ Кьяра Ромуальди; Джероламо Ланфранки (2009). «Статистические инструменты для анализа экспрессии генов и системной биологии и соответствующие веб-ресурсы». Стивен Кравец (ред.). Биоинформатика для системной биологии (2-е изд.). Хумана Пресс. стр. 181–205. дои : 10.1007/978-1-59745-440-7_11 . ISBN 978-1-59745-440-7 .
- ^ Войт, Эберхард (2012). Первый курс системной биологии . Гирляндная наука. ISBN 9780815344674 .
- ^ Байталук, М. (2009). «Системная биология регуляции генов». Биомедицинская информатика . Методы молекулярной биологии. Том. 569. стр. 55–87. дои : 10.1007/978-1-59745-524-4_4 . ISBN 978-1-934115-63-3 . ПМИД 19623486 .
- ^ Райт, Сьюэлл (1934). «Физиологические и эволюционные теории доминирования» . Американский натуралист . стр. 24–53.
- ^ Jump up to: Перейти обратно: а б с Клинг, Джим (3 марта 2006 г.). «Работа с системами» . Наука . Проверено 15 июня 2011 г.
- ^ «HMS открывает новый отдел по изучению системной биологии» . Гарвардская газета. 23 сентября 2003 г.
- ^ Карр, Джонатан Р.; Сангви, Джайодита К.; Маклин, Дерек Н.; Гучоу, Мириам В.; Джейкобс, Джаред М.; Боливаль, Бенджамин; Асад-Гарсия, Насира; Гласс, Джон И.; Коверт, Маркус В. (июль 2012 г.). «Целоклеточная вычислительная модель предсказывает фенотип на основе генотипа» . Клетка . 150 (2): 389–401. дои : 10.1016/j.cell.2012.05.044 . ПМЦ 3413483 . ПМИД 22817898 .
- ^ Месарович, Михаил Д. (1968). Теория систем и биология . Берлин: Springer-Verlag.
- ^ Jump up to: Перейти обратно: а б Розен, Роберт (5 июля 1968 г.). «Средство к новому холизму». Наука . 161 (3836): 34–35. Бибкод : 1968Sci...161...34M . дои : 10.1126/science.161.3836.34 . JSTOR 1724368 .
- ^ Качер, Х; Бернс, Дж. А. (1973). «Контроль потока». Симпозиумы Общества экспериментальной биологии . 27 : 65–104. PMID 4148886 .
- ^ Качер, Х; Бернс, Дж.А.; Фелл, Д.А. (1995). «Контроль потока». Труды Биохимического общества . 23 (2): 341–366. дои : 10.1042/bst0230341 . ПМИД 7672373 .
- ^ Генрих, Р; Рапопорт, Т.А. (1974). «Линейная стационарная теория ферментативных цепей: общие свойства, контроль и эффекторная сила» . Европейский журнал биохимии . 42 (1): 89–95. дои : 10.1111/j.1432-1033.1974.tb03318.x . ПМИД 4830198 .
- ^ Саважо, Майкл А. (декабрь 1969 г.). «Журнал теоретической биологии» . 25 (3): 365–369. дои : 10.1016/S0022-5193(69)80026-3 . ПМИД 5387046 .
{{cite journal}}: Для цитирования журнала требуется|journal=( помощь ) - ^ Саважо, Майкл А. (декабрь 1969 г.). «Журнал теоретической биологии» . 25 (3): 370–379. дои : 10.1016/S0022-5193(69)80027-5 . ПМИД 5387047 .
{{cite journal}}: Для цитирования журнала требуется|journal=( помощь ) - ^ Саважо, Майкл А. (февраль 1970 г.). «Журнал теоретической биологии» . 26 (2): 215–226. дои : 10.1016/S0022-5193(70)80013-3 . ПМИД 5434343 .
{{cite journal}}: Для цитирования журнала требуется|journal=( помощь ) - ^ «Системная биология — Национальный институт общих медицинских наук» . Архивировано из оригинала 19 октября 2013 года . Проверено 12 декабря 2012 г.
- ^ Хантер, Филип (май 2012 г.). «Возвращение на Землю: даже если системная биология еще не оправдала своих обещаний, она уже созрела и вот-вот принесет свои первые результаты» . Отчеты ЭМБО . 13 (5): 408–411. дои : 10.1038/embor.2012.49 . ПМЦ 3343359 . ПМИД 22491028 .
- ^ Цзоу, Явен; Лаубихлер, Манфред Д. (25 июля 2018 г.). «От систем к биологии: вычислительный анализ исследовательских статей по системной биологии с 1992 по 2013 год» . ПЛОС ОДИН . 13 (7): e0200929. Бибкод : 2018PLoSO..1300929Z . дои : 10.1371/journal.pone.0200929 . ISSN 1932-6203 . ПМК 6059489 . ПМИД 30044828 .
- ^ Jump up to: Перейти обратно: а б Касканте, Марта; Марин, Сильвия (30 сентября 2008 г.). «Метаболомика и флюксомика подходы». Очерки по биохимии . 45 : 67–82. дои : 10.1042/bse0450067 . ISSN 0071-1365 . ПМИД 18793124 .
- ^ Кьюсик, Майкл Э.; Клитгорд, Нильс; Видаль, Марк; Хилл, Дэвид Э. (15 октября 2005 г.). «Интерактом: ворота в системную биологию» . Молекулярная генетика человека . 14 (дополнение_2): Р171–Р181. дои : 10.1093/hmg/ddi335 . ISSN 0964-6906 . ПМИД 16162640 .
- ^ Аур, Дориан (2012). «От нейроэлектродинамики к думающим машинам». Когнитивные вычисления . 4 (1): 4–12. дои : 10.1007/s12559-011-9106-3 . ISSN 1866-9956 . S2CID 12355069 .
- ^ Лоор, Хурам Шахзад и Хуан Дж. (31 июля 2012 г.). «Применение системных подходов сверху вниз и снизу вверх в физиологии и метаболизме жвачных животных» . Современная геномика . 13 (5): 379–394. дои : 10.2174/138920212801619269 . ПМК 3401895 . ПМИД 23372424 .
- ^ Спилл, Фабиан; Бакал, Крис; Мак, Майкл (2018). «Механическая и системная биология рака» . Журнал вычислительной и структурной биотехнологии . 16 : 237–245. arXiv : 1807.08990 . Бибкод : 2018arXiv180708990S . дои : 10.1016/j.csbj.2018.07.002 . ПМК 6077126 . ПМИД 30105089 .
- ^ Барийо, Эммануэль; Кальцоне, Лоуренс; Хюп, Филипп; Верт, Жан-Филипп; Зиновьев, Андрей (2012). Вычислительная системная биология рака . Чепмен и Холл / CRCМатематическая и вычислительная биология. п. 461. ИСБН 978-1439831441 .
- ^ Бирн, Хелен М. (2010). «Рассечение рака с помощью математики: от клетки к животной модели». Обзоры природы Рак . 10 (3): 221–230. дои : 10.1038/nrc2808 . ПМИД 20179714 . S2CID 24616792 .
- ^ Гарднер, Тимоти .С; ди Бернардо, Диего; Лоренц, Дэвид; Коллинз, Джеймс Дж. (4 июля 2003 г.). «Вывод о генетических сетях и идентификация сложного механизма действия посредством профилирования экспрессии». Наука . 301 (5629): 102–105. Бибкод : 2003Sci...301..102G . дои : 10.1126/science.1081900 . ПМИД 12843395 . S2CID 8356492 .
- ^ ди Бернардо, Диего; Томпсон, Майкл Дж.; Гарднер, Тимоти С.; Чобот, Сара Э.; Иствуд, Эрин Л.; Войтович, Эндрю П.; Эллиотт, Шон Дж.; Шаус, Скотт Э.; Коллинз, Джеймс Дж. (март 2005 г.). «Хемогеномное профилирование в масштабе всего генома с использованием реконструированных генных сетей». Природная биотехнология . 23 (3): 377–383. дои : 10.1038/nbt1075 . ПМИД 15765094 . S2CID 16270018 .
- ^ Jump up to: Перейти обратно: а б Тавассоли, Иман (2015). Динамика решения клеточной судьбы, опосредованная взаимодействием аутофагии и апоптоза в раковых клетках . Спрингеровские тезисы. Международное издательство Спрингер. дои : 10.1007/978-3-319-14962-2 . ISBN 978-3-319-14961-5 . S2CID 89307028 .
- ^ Коркут, А; Ван, В; Демир, Э; Аксой, Б.А.; Цзин, X; Молинелли, Э.Дж.; Бабур, О; Бемис, Д.Л.; Онур Шумер, С; Солит, Д.Б.; Пратилас, Калифорния; Сандер, К. (18 августа 2015 г.). «Биология возмущений определяет комбинации лекарств выше и ниже в клетках меланомы, устойчивых к ингибитору RAF» . электронная жизнь . 4 . doi : 10.7554/eLife.04640 . ПМЦ 4539601 . ПМИД 26284497 .
- ^ Jump up to: Перейти обратно: а б Гупта, Анкур; Роулингс, Джеймс Б. (апрель 2014 г.). «Сравнение методов оценки параметров в стохастических химико-кинетических моделях: примеры из системной биологии» . Журнал Айше . 60 (4): 1253–1268. дои : 10.1002/aic.14409 . ISSN 0001-1541 . ПМЦ 4946376 . ПМИД 27429455 .
- ^ Ананаду, София ; Келл, Дуглас; Цудзи, Дзюнъити (декабрь 2006 г.). «Интеллектуальный анализ текста и его потенциальные применения в системной биологии». Тенденции в биотехнологии . 24 (12): 571–579. дои : 10.1016/j.tibtech.2006.10.002 . ПМИД 17045684 .
- ^ Глааб, Энрико; Шнайдер, Рейнхард (2012). «PathVar: анализ вариабельности экспрессии генов и белков в клеточных путях с использованием данных микрочипов» . Биоинформатика . 28 (3): 446–447. doi : 10.1093/биоинформатика/btr656 . ПМЦ 3268235 . ПМИД 22123829 .
- ^ Бардини, Р.; Политано, Г.; Бенсо, А.; Ди Карло, С. (01 января 2017 г.). «Многоуровневые и гибридные подходы к моделированию в системной биологии» . Журнал вычислительной и структурной биотехнологии . 15 : 396–402. дои : 10.1016/j.csbj.2017.07.005 . ISSN 2001-0370 . ПМЦ 5565741 . ПМИД 28855977 .
- ^ Jump up to: Перейти обратно: а б Транструм, Марк К.; Цю, Пэн (17 мая 2016 г.). «Соединение механистических и феноменологических моделей сложных биологических систем» . PLOS Вычислительная биология . 12 (5): e1004915. arXiv : 1509.06278 . Бибкод : 2016PLSCB..12E4915T . дои : 10.1371/journal.pcbi.1004915 . ISSN 1553-7358 . ПМЦ 4871498 . ПМИД 27187545 .
- ^ Челлабойна, В.; Бхат, СП; Хаддад, ВМ; Бернштейн, DS (август 2009 г.). «Моделирование и анализ кинетики действия масс». Журнал IEEE Control Systems . 29 (4): 60–78. дои : 10.1109/MCS.2009.932926 . ISSN 1941-000X . S2CID 12122032 .
- ^ Браун, Кевин С.; Сетна, Джеймс П. (12 августа 2003 г.). «Статистически-механические подходы к моделям со многими плохо известными параметрами». Физический обзор E . 68 (2): 021904. Бибкод : 2003PhRvE..68b1904B . дои : 10.1103/physreve.68.021904 . ISSN 1063-651X . ПМИД 14525003 .
- ^ Орт, Джеффри Д.; Тиле, Инес; Палссон, Бернхард О (март 2010 г.). «Что такое анализ баланса потоков?» . Природная биотехнология . 28 (3): 245–248. дои : 10.1038/nbt.1614 . ISSN 1087-0156 . ПМК 3108565 . ПМИД 20212490 .
Дальнейшее чтение [ править ]
- Клип, Эдда; Либермейстер, Вольфрам; Вирлинг, Кристоф; Ковальд, Аксель (2016). Системная биология. Учебник, 2-е издание . Уайли. ISBN 978-3-527-33636-4 .
- Асфар С. Азми, изд. (2012). Системная биология в исследованиях рака и открытии лекарств . Спрингер. ISBN 978-94-007-4819-4 .
- Китано, Хироаки (15 октября 2001 г.). Основы системной биологии . МТИ Пресс. ISBN 978-0-262-11266-6 .
- Вернер, Эрик (29 марта 2007 г.). «Все системы работают» . Природа . 446 (7135): 493–494. Бибкод : 2007Natur.446..493W . дои : 10.1038/446493а . приводится сравнительный обзор трех книг:
- Алон, Ури (7 июля 2006 г.). Введение в системную биологию: принципы проектирования биологических цепей . Чепмен и Холл. ISBN 978-1-58488-642-6 .
- Канеко, Кунихико (15 сентября 2006 г.). Жизнь: введение в биологию сложных систем . Спрингер-Верлаг. Бибкод : 2006lics.book.....K . ISBN 978-3-540-32666-3 .
- Палссон, Бернхард О. (16 января 2006 г.). Системная биология: свойства реконструированных сетей . Издательство Кембриджского университета. ISBN 978-0-521-85903-5 .
- Вернер Дубицкий; Олаф Волькенхауэр; Хироки Ёкота; Кван-Хён Чо, ред. (13 августа 2013 г.). Энциклопедия системной биологии . Издательство Спрингер. ISBN 978-1-4419-9864-4 .
Внешние ссылки [ править ]
 СМИ, связанные с системной биологией, на Викискладе?
СМИ, связанные с системной биологией, на Викискладе? - Биологические системы в биофизике-вики
