Интерактом
В молекулярной биологии интерактом — это совокупность молекулярных взаимодействий в конкретной клетке . Этот термин конкретно относится к физическим взаимодействиям между молекулами (например, между белками, также известными как белок-белковые взаимодействия , ИПП; или между небольшими молекулами и белками). [1] ), но также может описывать наборы непрямых взаимодействий между генами ( генетические взаимодействия ).
Слово «интерактом» было первоначально придумано в 1999 году группой французских ученых во главе с Бернаром Жаком. [3] Математически интерактомы обычно отображаются в виде графиков . Хотя интерактомы можно назвать биологическими сетями , их не следует путать с другими сетями, такими как нейронные сети или пищевые сети .
Сети молекулярного взаимодействия
[ редактировать ]Молекулярные взаимодействия могут происходить между молекулами, принадлежащими к разным биохимическим семействам (белки, нуклеиновые кислоты, липиды, углеводы и др.), а также внутри одного семейства. Всякий раз, когда такие молекулы соединяются физическими взаимодействиями, они образуют сети молекулярных взаимодействий, которые обычно классифицируются по природе участвующих соединений. Чаще всего интерактом относится к сети белок-белкового взаимодействия (PPI) (PIN) или ее подмножествам. Например, интерактом белка Sirt-1 и интерактом второго порядка семейства Sirt. [4] [5] представляет собой сеть, включающую Sirt-1 и его непосредственно взаимодействующие белки, где в качестве интерактома второго порядка иллюстрируются взаимодействия до соседей второго порядка (соседей соседей). Другим широко изученным типом интерактома является интерактом белок-ДНК, также называемый генно-регуляторной сетью , сеть, образованная факторами транскрипции, регуляторными белками хроматина и их генами-мишенями. Даже метаболические сети можно рассматривать как сети молекулярного взаимодействия: метаболиты, то есть химические соединения в клетке, превращаются друг в друга с помощью ферментов , которым приходится физически связывать свои субстраты.
На самом деле все типы интерактомов взаимосвязаны. Например, белковые интерактомы содержат множество ферментов, которые, в свою очередь, образуют биохимические сети. Сходным образом, сети регуляции генов существенно перекрываются с сетями взаимодействия белков и сигнальными сетями.
Размер
[ редактировать ]
Было высказано предположение, что размер интерактома организма лучше, чем размер генома , коррелирует с биологической сложностью организма. [7] Хотя карты белок-белковых взаимодействий, содержащие несколько тысяч бинарных взаимодействий, теперь доступны для нескольких видов, ни одна из них в настоящее время не является полной, а размер интерактомов все еще остается предметом дискуссий.
Дрожжи
[ редактировать ]По оценкам , дрожжевой интерактом, то есть все белок-белковые взаимодействия между белками Saccharomyces cerevisiae , содержит от 10 000 до 30 000 взаимодействий. Разумная оценка может составлять порядка 20 000 взаимодействий. Более крупные оценки часто включают косвенные или прогнозируемые взаимодействия, часто на основе исследований аффинной очистки / масс-спектрометрии (AP/MS). [6]
Сети генетического взаимодействия
[ редактировать ]Гены взаимодействуют в том смысле, что влияют на функции друг друга. Например, мутация может быть безвредной, но в сочетании с другой мутацией такое сочетание может оказаться смертельным. Говорят, что такие гены «взаимодействуют генетически». Гены, связанные таким образом, образуют сети генетического взаимодействия . Некоторые из целей этих сетей: разработать функциональную карту клеточных процессов, идентифицировать мишени лекарств с помощью хемопротеомики и предсказать функцию неохарактеризованных генов.
В 2010 году наиболее «полный» генный интерактом, созданный на сегодняшний день, был составлен из примерно 5,4 миллионов сравнений двух генов для описания «профилей взаимодействия для ~ 75% всех генов почкующихся дрожжей », с ~ 170 000 взаимодействий генов. Гены были сгруппированы по схожим функциям, чтобы построить функциональную карту клеточных процессов. Используя этот метод, исследование смогло предсказать известные функции генов лучше, чем любой другой набор данных в масштабе генома, а также добавить функциональную информацию для генов, которые ранее не были описаны. С помощью этой модели генетические взаимодействия можно наблюдать в различных масштабах, что поможет в изучении таких концепций, как сохранение генов. Некоторые из наблюдений, сделанных в ходе этого исследования, заключаются в том, что отрицательных взаимодействий было в два раза больше, чем положительных , отрицательные взаимодействия были более информативными, чем положительные взаимодействия, а гены с большим количеством связей с большей вероятностью приводили к летальному исходу при нарушении. [8]
Интерактомика
[ редактировать ]Интерактомика – это дисциплина на стыке биоинформатики и биологии , которая занимается изучением как взаимодействий, так и последствий этих взаимодействий между белками и другими молекулами внутри клетки . [9] Таким образом, интерактомика направлена на сравнение таких сетей взаимодействий (т. е. интерактомов) между видами и внутри них, чтобы выяснить, как черты таких сетей либо сохраняются, либо изменяются.
Интерактомика — это пример системной биологии «сверху вниз» , которая рассматривает биосистему или организм сверху вниз. Собираются большие наборы общегеномных и протеомных данных и выводятся корреляции между различными молекулами. На основе полученных данных сформулированы новые гипотезы об обратных связях между этими молекулами. Эти гипотезы затем можно будет проверить с помощью новых экспериментов. [10]
Экспериментальные методы картирования интерактомов
[ редактировать ]Изучение интерактомов называется интерактомикой. Основной единицей белковой сети является белок-белковое взаимодействие (PPI). Несмотря на то, что существует множество методов изучения ИПП, лишь немногие из них использовались в больших масштабах для картирования целых интерактомов.
( Двухгибридная система дрожжей Y2H) подходит для изучения бинарных взаимодействий между двумя белками одновременно. Аффинная очистка и последующая масс-спектрометрия подходят для идентификации белкового комплекса. Оба метода можно использовать с высокой пропускной способностью (HTP). Дрожжевой двухгибридный скрининг допускает ложноположительные взаимодействия между белками, которые никогда не экспрессируются в одно и то же время и в одном и том же месте; Масс-спектрометрия с аффинным захватом не имеет этого недостатка и в настоящее время является золотым стандартом. Данные о двухгибридных дрожжах лучше указывают на неспецифическую тенденцию к липким взаимодействиям, тогда как масс-спектрометрия с аффинным захватом лучше указывает на функциональные белок-белковые взаимодействия in vivo. [11] [12]
Вычислительные методы исследования интерактомов
[ редактировать ]После создания интерактома существует множество способов проанализировать его свойства. Однако есть две важные цели такого анализа. Во-первых, ученые пытаются выяснить системные свойства интерактомов, например, топологию их взаимодействий. Во-вторых, исследования могут быть сосредоточены на отдельных белках и их роли в сети. Такие анализы в основном проводятся с использованием методов биоинформатики и включают, среди многих других, следующие:
Валидация
[ редактировать ]Во-первых, необходимо оценить охват и качество интерактома. Интерактомы никогда не бывают полными, учитывая ограничения экспериментальных методов. Например, было подсчитано, что типичные скрининги Y2H обнаруживают только 25% или около того всех взаимодействий в интерактоме. [13] Охват интерактома можно оценить, сравнив его с эталонами хорошо известных взаимодействий, которые были обнаружены и подтверждены независимыми анализами. [14] Другие методы отфильтровывают ложноположительные результаты, вычисляя сходство известных аннотаций участвующих белков, или определяют вероятность взаимодействия, используя субклеточную локализацию этих белков. [15]
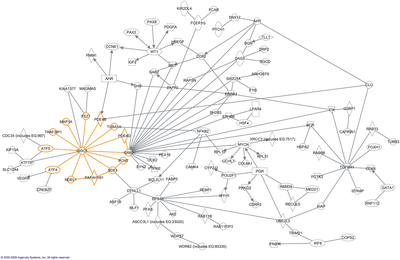
Прогнозирование ИЦП
[ редактировать ]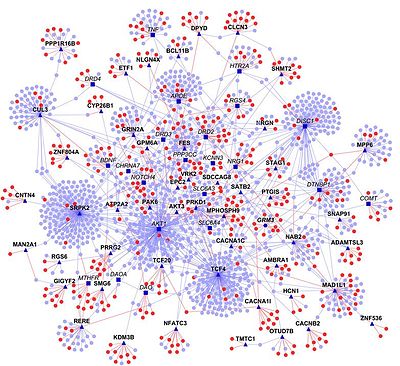
Используя экспериментальные данные в качестве отправной точки, перенос гомологии является одним из способов прогнозирования интерактомов. Здесь ИПП одного организма используются для прогнозирования взаимодействий между гомологичными белками в другом организме (« интерологи »). Однако этот подход имеет определенные ограничения, прежде всего потому, что исходные данные могут быть ненадежными (например, содержать ложноположительные и ложноотрицательные результаты). [17] Кроме того, белки и их взаимодействия изменяются в ходе эволюции и, следовательно, могут быть потеряны или приобретены. Тем не менее, были предсказаны многочисленные интерактомы, например, у Bacillus licheniformis . [18]
Некоторые алгоритмы используют экспериментальные данные о структурных комплексах, атомные детали интерфейсов связывания и создают подробные атомные модели белок-белковых комплексов. [19] [20] а также другие белок-молекулярные взаимодействия. [21] [22] Другие алгоритмы используют только информацию о последовательности, тем самым создавая объективные полные сети взаимодействия со многими ошибками. [23]
Некоторые методы используют машинное обучение, чтобы отличить, чем взаимодействующие пары белков отличаются от пар невзаимодействующих белков с точки зрения парных особенностей, таких как клеточная колокализация, совместная экспрессия генов, насколько близко расположены на ДНК гены, кодирующие два белка, и т. д. на. [16] [24] Случайный лес оказался наиболее эффективным методом машинного обучения для прогнозирования взаимодействия белков. [25] Такие методы были применены для обнаружения белковых взаимодействий в интерактоме человека, в частности в интерактоме мембранных белков. [24] и интерактом белков, связанных с шизофренией. [16]
Текстовый анализ ИЦП
[ редактировать ]Были предприняты некоторые усилия по систематическому извлечению сетей взаимодействия непосредственно из научной литературы. По сложности такие подходы варьируются от простой статистики совместного появления объектов, которые упоминаются вместе в одном и том же контексте (например, в предложении), до сложных методов обработки естественного языка и машинного обучения для обнаружения взаимосвязей. [26]
Прогнозирование функции белка
[ редактировать ]Сети взаимодействия белков использовались для предсказания функций белков с неизвестными функциями. [27] [28] Обычно это основано на предположении, что неохарактеризованные белки имеют те же функции, что и взаимодействующие с ними белки ( вина по ассоциации ). Например, было обнаружено, что YbeB, белок с неизвестной функцией, взаимодействует с рибосомальными белками, а позже было показано, что он участвует в бактериальной и эукариотической (но не архейной) трансляции . [29] Хотя такие предсказания могут быть основаны на единичных взаимодействиях, обычно обнаруживаются несколько взаимодействий. Таким образом, вся сеть взаимодействий может быть использована для предсказания функций белка, учитывая, что определенные функции обычно обогащаются среди интеракторов. [27] Термин «гипотом» использовался для обозначения интерактома, в котором по крайней мере один из генов или белков является гипотетическим белком . [30]
Возмущения и болезни
[ редактировать ]Топология возмущение интерактома позволяет предсказать, как сеть реагирует на ( например, удаление) узлов (белков) или ребер (взаимодействий). [31] Подобные возмущения могут быть вызваны мутациями генов и, следовательно, их белков, а сетевая реакция может проявиться как заболевание . [32] Сетевой анализ может выявить мишени для лекарств и биомаркеры заболеваний. [33]
Структура и топология сети
[ редактировать ]Сети взаимодействия можно анализировать с помощью инструментов теории графов . Свойства сети включают в себя распределение степеней , коэффициенты кластеризации , центральность по посредничеству и многие другие. Распределение свойств среди белков интерактома показало, что сети интерактома часто имеют безмасштабную топологию. [34] где функциональные модули внутри сети обозначают специализированные подсети. [35] Такие модули могут быть функциональными, как в сигнальном пути , или структурными, как в белковом комплексе. Фактически, идентификация белковых комплексов в интерактоме представляет собой сложную задачу, учитывая, что сеть сама по себе не выявляет напрямую наличие стабильного комплекса.
Изучены интерактомы
[ редактировать ]Вирусные интерактомы
[ редактировать ]Интерактомы вирусных белков состоят из взаимодействий между вирусными или фаговыми белками. Они были одними из первых проектов интерактомов, поскольку их геномы невелики, и все белки можно анализировать с ограниченными ресурсами. Вирусные интерактомы связаны с интерактомами своих хозяев, образуя сети взаимодействия вирус-хозяин. [36] Некоторые опубликованные интерактомы вирусов включают
Бактериофаг
- Escherichia coli Бактериофаг лямбда [37]
- Бактериофаг Escherichia coli Т7 [38]
- Бактериофаг Streptococcus pneumoniae Dp-1 [39]
- Бактериофаг Streptococcus pneumoniae Cp-1 [40]
Интерактомы лямбда и VZV важны не только для биологии этих вирусов, но и по техническим причинам: они были первыми интерактомами, которые были картированы с помощью нескольких векторов Y2H , что доказывает улучшенную стратегию более полного исследования интерактомов, чем показали предыдущие попытки.
Вирусы человека (млекопитающих)
- человека Вирус ветряной оспы (VZV) [41]
- Вирус Чандипура [42]
- Вирус Эпштейна-Барра (ВЭБ) [43]
- вирус гепатита С (ВГС), [44] Взаимодействие человека и ВГС [45]
- Вирус гепатита Е (ВГЕ) [46]
- Вирус простого герпеса 1 (ВПГ-1) [43]
- Герпесвирус, ассоциированный с саркомой Капоши (KSHV) [43]
- Мышиный цитомегаловирус (мЦМВ) [43]
Бактериальные интерактомы
[ редактировать ]Относительно небольшое количество бактерий было всесторонне изучено на предмет их белок-белковых взаимодействий. Однако ни один из этих интерактомов не является полным в том смысле, что они охватывают все взаимодействия. Фактически, было подсчитано, что ни один из них не охватывает более 20–30% всех взаимодействий, главным образом потому, что в большинстве этих исследований использовался только один метод, каждый из которых обнаруживает только подмножество взаимодействий. [13] Среди опубликованных бактериальных интерактомов (в том числе частичных)
| Разновидность | белки всего | взаимодействия | тип | ссылка |
| Хеликобактер пилори | 1,553 | ~3,004 | Y2H | [47] [48] |
| Кампилобактер жеюни | 1,623 | 11,687 | Y2H | [49] |
| Трепонема бледная | 1,040 | 3,649 | Y2H | [50] |
| кишечная палочка | 4,288 | (5,993) | АП/МС | [51] |
| кишечная палочка | 4,288 | 2,234 | Y2H | [52] |
| Мезоризобиум лоти | 6,752 | 3,121 | Y2H | [53] |
| Микобактерия туберкулеза | 3,959 | >8000 | B2H | [54] |
| Микоплазма гениталиум | 482 | АП/МС | [55] | |
| Синехоцистис сп. PCC6803 | 3,264 | 3,236 | Y2H | [56] |
| Золотистый стафилококк (MRSA) | 2,656 | 13,219 | АП/МС | [57] |
Интерактомы E. coli и Mycoplasma были проанализированы с использованием крупномасштабной аффинной очистки белковых комплексов и масс-спектрометрии (AP/MS), поэтому сделать вывод о прямых взаимодействиях нелегко. Остальные использовали обширный двухгибридный скрининг дрожжей (Y2H). Интерактом Mycobacterium Tuberculosis анализировали с помощью бактериального двухгибридного скрининга (B2H).
Обратите внимание, что с помощью вычислительных методов было предсказано множество дополнительных интерактомов (см. раздел выше).
Эукариотические интерактомы
[ редактировать ]Было предпринято несколько попыток картировать эукариотические интерактомы с помощью методов HTP. Хотя ни один биологический интерактом не был полностью охарактеризован, более 90% белков в Saccharomyces cerevisiae были проверены и охарактеризованы их взаимодействия, что делает его наиболее охарактеризованным интерактомом. [27] [58] [59] Виды, интерактомы которых изучены достаточно подробно, включают
- Шизосахаромицеты помбе [60] [61]
- Ценорабдитис элегантный
- Дрозофила меланогастер
- Мудрый человек
Недавно взаимодействие патоген-хозяин вируса гепатита С/человека (2008 г.) [62] Вирус Эпштейна-Барра/Человек (2008), Вирус гриппа/Человек (2009) были выделены с помощью HTP для идентификации основных молекулярных компонентов патогенов и иммунной системы хозяина. [63]
Предсказанные интерактомы
[ редактировать ]Как описано выше, можно предсказать ИПП и, следовательно, целые интерактомы. Хотя надежность этих предсказаний является спорной, они дают гипотезы, которые можно проверить экспериментально. Интерактомы были предсказаны для ряда видов, например
- Человек ( Homo sapiens ) [64]
- Рис ( Oryza sativa ) [65]
- Ксантомонада oryzae [66]
- Арабидопсис Талиана [67]
- Помидор ( помидор пасленум ) [68]
- Горчица полевая ( Brassica rapa ) [69]
- Кукуруза, кукуруза ( Zea mays ) [70]
- Тополь ( Populus trichocarpa ) [71]
- SARS-CoV-2 [72]

Свойства сети
[ редактировать ]Сети взаимодействия белков можно анализировать с помощью того же инструмента, что и другие сети. Фактически, они имеют много общих свойств с биологическими или социальными сетями . Некоторые из основных характеристик заключаются в следующем.

Распределение степеней
[ редактировать ]Распределение по степени описывает количество белков, имеющих определенное количество связей. Большинство сетей взаимодействия белков демонстрируют безмасштабное ( степенное ) распределение степеней, при котором распределение связности P(k) ~ k -с где k — степень. Эту зависимость также можно рассматривать как прямую линию на логарифмическом графике , поскольку приведенное выше уравнение равно log(P(k)) ~ —y•log(k). Одной из характеристик таких распределений является то, что существует много белков с небольшим количеством взаимодействий и мало белков с множеством взаимодействий, причем последние называются «концентраторами».
Хабы
[ редактировать ]Сильно связанные узлы (белки) называются хабами. Хан и др. [73] придумали термин « партийный хаб » для хабов, выражение которых коррелирует с их партнерами по взаимодействию. Концентраторы Party также соединяют белки внутри функциональных модулей, таких как белковые комплексы. Напротив, « концентраторы данных » не демонстрируют такой корреляции и, по-видимому, соединяют различные функциональные модули. Центры вечеринок встречаются преимущественно в наборах данных AP/MS, тогда как концентраторы данных встречаются преимущественно в сетевых картах двоичных интерактомов. [74] Обратите внимание, что достоверность различия между центром дат и центром вечеринки была оспорена. [75] [76] Хабы Party обычно состоят из белков с несколькими интерфейсами, тогда как концентраторы дат чаще представляют собой белки интерфейса с одним взаимодействием. [77] В соответствии с ролью дата-центров в соединении различных процессов у дрожжей количество бинарных взаимодействий данного белка коррелирует с количеством фенотипов, наблюдаемых для соответствующего мутантного гена в различных физиологических условиях. [74]
Модули
[ редактировать ]Узлы, участвующие в одном и том же биохимическом процессе, тесно взаимосвязаны. [33]
Эволюция
[ редактировать ]Эволюция сложности интерактома описана в исследовании, опубликованном в журнале Nature . [78] В этом исследовании впервые отмечается, что границы между прокариотами , одноклеточными эукариотами и многоклеточными эукариотами сопровождаются сокращением на порядки величины эффективного размера популяции с одновременным усилением эффектов случайного генетического дрейфа . Получающееся в результате снижение эффективности отбора, по-видимому, является достаточным для того, чтобы неадаптивным образом влиять на широкий спектр признаков на геномном уровне. Исследование Nature показывает, что изменение силы случайного генетического дрейфа также способно влиять на филогенетическое разнообразие на субклеточном и клеточном уровнях. Таким образом, размер популяции следует рассматривать как потенциальную детерминанту механистических путей, лежащих в основе долгосрочной фенотипической эволюции. В исследовании далее показано, что существует филогенетически широкая обратная связь между силой дрейфа и структурной целостностью белковых субъединиц. Таким образом, накопление умеренно вредных мутаций в популяциях небольшого размера вызывает вторичный отбор для белок-белковые взаимодействия , которые стабилизируют ключевые функции генов, смягчая структурную деградацию, вызванную неэффективным отбором. Таким образом, сложные белковые архитектуры и взаимодействия, необходимые для генезиса фенотипического разнообразия, могут изначально возникать посредством неадаптивных механизмов.
Критика, вызовы и ответы
[ редактировать ]Возможно, этот раздел содержит оригинальные исследования . ( Август 2015 г. ) |
Кимер и Чезарени [9] вызывают следующие опасения по поводу состояния (около 2007 г.) поля, особенно в отношении сравнительной интерактомики: Экспериментальные процедуры, связанные с полем, подвержены ошибкам, приводящим к «зашумленным результатам». Это приводит к тому, что 30% всех зарегистрированных взаимодействий являются артефактами. Фактически, две группы, использовавшие одни и те же методы на одном и том же организме, обнаружили менее 30% общих взаимодействий. Однако некоторые авторы утверждают, что такая невоспроизводимость является результатом чрезвычайной чувствительности различных методов к небольшим экспериментальным вариациям. Например, идентичные условия в анализах Y2H приводят к очень разным взаимодействиям при использовании разных векторов Y2H. [13]
Методы могут быть предвзятыми, т.е. метод определяет, какие взаимодействия будут обнаружены. Фактически, любой метод имеет предвзятость, особенно белковые методы. Поскольку каждый белок уникален, ни один метод не может выявить свойства каждого белка. Например, большинство аналитических методов, которые хорошо работают с растворимыми белками, плохо работают с мембранными белками. Это также справедливо для технологий Y2H и AP/MS.
Интерактомы далеко не полные, за исключением, пожалуй, S. cerevisiae. На самом деле это не критика, поскольку любая научная область изначально является «неполной», пока методологии не будут усовершенствованы. Интерактомика в 2015 году — это то место, где секвенирование генома происходило в конце 1990-х годов, учитывая, что доступно лишь несколько наборов данных интерактома (см. таблицу выше).
Хотя геномы стабильны, интерактомы могут различаться в зависимости от тканей, типов клеток и стадий развития. Опять же, это не критика, а скорее описание проблем в этой области.
Трудно сопоставить эволюционно родственные белки у отдаленно родственных видов. Хотя гомологичные последовательности ДНК можно найти относительно легко, гораздо сложнее предсказать гомологичные взаимодействия («интерологи»), поскольку гомологи двух взаимодействующих белков не нуждаются во взаимодействии. Например, даже внутри протеома два белка могут взаимодействовать, а их паралоги — нет.
Каждый белок-белковый интерактом может представлять собой лишь частичную выборку потенциальных взаимодействий, даже если предположительно окончательная версия опубликована в научном журнале. Дополнительные факторы могут играть роль во взаимодействиях белков, которые еще не включены в интерактомы. Сила связывания различных белковых интеракторов, факторы микроокружения, чувствительность к различным процедурам и физиологическое состояние клетки — все это влияет на белок-белковые взаимодействия, но обычно не учитывается в исследованиях интерактома. [79]
См. также
[ редактировать ]- Биоинформатика , Омика , Протеомика , Геномика
- БиоПлекс
- Коннектом
- Глоссарий теории графов
- Человеческий интерактом
- Список тем омики по биологии
- Математическая биология
- Метаболическая сеть
- Моделирование метаболической сети
- Метаболический путь
- Сетевая медицина
Ссылки
[ редактировать ]- ^ Ван Л., Эфтехари П., Шахнер Д., Игнатова И.Д., Пальме В., Шильхер Н., Ладурнер А., Хейсс Э.Х., Штангл Х., Дирш В.М., Атанасов А.Г. Новый интерактивный подход идентифицирует ABCA1 как прямую мишень эводиамина, который увеличивает отток холестерина из макрофагов . Sci Rep. 23 июля 2018 г.; 8 (1): 11061. дои: 10.1038/s41598-018-29281-1 .
- ^ Хна В., Портеус Д. (2009). Рейф А (ред.). «Путь DISC1 модулирует экспрессию генов нейроразвития, синаптогенности и сенсорного восприятия» . ПЛОС ОДИН . 4 (3): e4906. Бибкод : 2009PLoSO...4.4906H . дои : 10.1371/journal.pone.0004906 . ПМЦ 2654149 . ПМИД 19300510 .

- ^ Санчес С; Лашез С; Джаноди Ф; и др. (январь 1999 г.). «Изучение молекулярных взаимодействий и генетических сетей у Drosophila melanogaster с использованием базы данных Интернета FlyNets» . Нуклеиновые кислоты Рез . 27 (1): 89–94. дои : 10.1093/нар/27.1.89 . ПМК 148104 . ПМИД 9847149 .
- ^ Шарма, Анкуш; Гаутам В.К.; Константини С; Паладино А; Колонна G (февраль 2012 г.). «Интерактомные и фармакологические данные о человеческом Sirt-1» . Передний. Фармакол . 3 : 40. дои : 10.3389/fphar.2012.00040 . ПМК 3311038 . ПМИД 22470339 .
- ^ Шарма, Анкуш; Константини С; Колонна G (март 2013 г.). «Сеть белок-белкового взаимодействия семейства сиртуинов человека». Biochimica et Biophysical Acta (BBA) - Белки и протеомика . 1834 (10): 1998–2009. arXiv : 1302.6423 . Бибкод : 2013arXiv1302.6423S . дои : 10.1016/j.bbapap.2013.06.012 . ПМИД 23811471 . S2CID 15003130 .
- ^ Jump up to: а б Уэц П. и Григорьев А. (2005) Интерактом дрожжей. В Джорде, Л.Б., Литтле, ПФР, Данне, М.Дж. и Субраманиаме, С. (редакторы), Энциклопедия генетики, геномики, протеомики и биоинформатики. John Wiley & Sons Ltd: Чичестер, том 5, стр. 2033–2051 гг.
- ^ Штумпф депутат; Торн Т; де Сильва Э; и др. (май 2008 г.). «Оценка размеров интерактома человека» . Учеб. Натл. акад. наук. США . 105 (19): 6959–64. Бибкод : 2008PNAS..105.6959S . дои : 10.1073/pnas.0708078105 . ПМЦ 2383957 . ПМИД 18474861 .
- ^ Костанцо М; Барышникова А; Беллей Дж.; и др. (22 января 2010 г.). «Генетический ландшафт клетки» . Наука . 327 (5964): 425–431. Бибкод : 2010Sci...327..425C . дои : 10.1126/science.1180823 . ПМК 5600254 . ПМИД 20093466 .
- ^ Jump up to: а б Кимер, Л; Дж. Чезарени (2007). «Сравнительная интерактомика: сравниваем яблоки и груши?». Тенденции в биотехнологии . 25 (10): 448–454. дои : 10.1016/j.tibtech.2007.08.002 . ПМИД 17825444 .
- ^ Брюггеман, Ф.Дж.; Х.В. Вестерхофф (2006). «Природа системной биологии». Тенденции в микробиологии . 15 (1): 45–50. дои : 10.1016/j.tim.2006.11.003 . ПМИД 17113776 .
- ^ Бреттнер, Леандра М.; Джоанна Мазель (2012). «Липкость белка, а не количество функциональных белок-белковых взаимодействий, предсказывает шум экспрессии и пластичность у дрожжей» . Системная биология BMC . 6 : 128. дои : 10.1186/1752-0509-6-128 . ПМК 3527306 . ПМИД 23017156 .
- ^ Мукерджи, К; Славсон; Кристманн; Гриффит (июнь 2014 г.). «Нейрон-специфические белковые взаимодействия дрозофилы CASK-ß выявлены с помощью масс-спектрометрии» . Передний. Мол. Нейроски . 7:58 . doi : 10.3389/fnmol.2014.00058 . ПМК 4075472 . ПМИД 25071438 .
- ^ Jump up to: а б с Чен, ЮК; Раджагопала, СВ; Стеллбергер, Т.; Утц, П. (2010). «Исчерпывающий бенчмаркинг дрожжевой двухгибридной системы» . Природные методы . 7 (9): 667–668, автор 668 668. doi : 10.1038/nmeth0910-667 . ПМЦ 10332476 . ПМИД 20805792 . S2CID 35834541 .
- ^ Раджагопала, СВ; Хьюз, КТ; Уец, П. (2009). «Сравнительный анализ дрожжевых двухгибридных систем с использованием взаимодействия белков подвижности бактерий» . Протеомика . 9 (23): 5296–5302. дои : 10.1002/pmic.200900282 . ПМЦ 2818629 . ПМИД 19834901 .
- ^ Янай Офран , Гай Ячдав , Эял Мозес , Та-цен Сунг , Раджеш Наир и Буркхард Рост (июль 2006 г.). «Создавать и оценивать белковые сети с помощью молекулярных характеристик отдельных белков» . Биоинформатика . 22 (14): е402–е407. doi : 10.1093/биоинформатика/btl258 . ПМИД 16873500 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Jump up to: а б с Ганапатираджу М.К., Тахир М., Ханден А., Саркар С.Н., Свит Р.А., Нимгаонкар В.Л., Лошер К.Э., Бауэр Э.М., Чапарала С. (апрель 2016 г.). «Интерактом шизофрении с 504 новыми белок-белковыми взаимодействиями» . НПЖ Шизофрения . 2 : 16012. дои : 10.1038/npjschz.2016.12 . ПМЦ 4898894 . ПМИД 27336055 .
- ^ Мика С., Рост Б. (2006). «Белко-белковые взаимодействия более консервативны внутри видов, чем между видами» . PLOS Вычислительная биология . 2 (7): е79. Бибкод : 2006PLSCB...2...79M . дои : 10.1371/journal.pcbi.0020079 . ПМЦ 1513270 . ПМИД 16854211 .
- ^ Хан, Ю.-К.; и др. (2016). «Прогнозирование и характеристика сети белок-белковых взаимодействий у Bacillus licheniformis WX-02» . наук. Представитель . 6 : 19486. Бибкод : 2016NatSR...619486H . дои : 10.1038/srep19486 . ПМК 4726086 . ПМИД 26782814 .
- ^ Киттихотират В., Геркен М., Бамгарнер Р.Э., Самудрала Р. (2009). «Protinfo PPC: веб-сервер для прогнозирования белковых комплексов на атомном уровне» . Исследования нуклеиновых кислот . 37 (проблема с веб-сервером): W519–W525. дои : 10.1093/нар/gkp306 . ПМК 2703994 . ПМИД 19420059 .
- ^ Тьяги, М; Хасимото, К; Шумейкер, бакалавр искусств; Вухти, С; Панченко А.Р. (март 2012 г.). «Крупномасштабное картирование белкового интерактома человека с использованием структурных комплексов» . Представитель ЭМБО . 13 (3): 266–71. дои : 10.1038/embor.2011.261 . ПМК 3296913 . ПМИД 22261719 .
- ^ Макдермотт Дж., Геркен М., Фрейзер З., Чанг А.Н., Самудрала Р. (2005). «BIOVERSE: усовершенствования структуры структурных, функциональных и контекстуальных аннотаций белков и протеомов» . Исследования нуклеиновых кислот . 33 (проблема с веб-сервером): W324–W325. дои : 10.1093/nar/gki401 . ПМК 1160162 . ПМИД 15980482 .
- ^ Шумейкер, бакалавр искусств; Чжан, Д; Тьяги, М; Тангуду, РР; Фонг, Дж. Х.; Марчлер-Бауэр, А; Брайант, Ш.; Мадей, Т; Панченко А.Р. (январь 2012 г.). «IBIS (Сервер предполагаемого биомолекулярного взаимодействия) сообщает, прогнозирует и интегрирует множество типов консервативных взаимодействий белков» . Нуклеиновые кислоты Рез . 40 (Проблема с базой данных): D834–40. дои : 10.1093/nar/gkr997 . ПМЦ 3245142 . ПМИД 22102591 . Хопф Т.А., Шарфе К.П., Родригес Дж.П., Грин А.Г., Кольбахер О., Сандер С., Бонвин А.М., Маркс Д.С. (2014). «Коэволюция последовательностей дает трехмерные контакты и структуры белковых комплексов» . электронная жизнь . 3 : e03430. arXiv : 1405.0929 . Бибкод : 2014arXiv1405.0929H . doi : 10.7554/eLife.03430 . ПМЦ 4360534 . ПМИД 25255213 .
- ^ Котляр М, Пастрелло С, Пиветта Ф, Ло Сардо А, Кумбаа С, Ли Х, Наранян Т, Ню Ю, Дин З, Вафаи Ф, Брокс-Картер Ф, Петшнигг Дж, Миллс ГБ, Юрисикова А, Стагляр И, Маэстро Р , Юришица I (2015). «In silico предсказание физических взаимодействий белков и характеристика сирот интерактома». Природные методы . 12 (1): 79–84. дои : 10.1038/nmeth.3178 . ПМИД 25402006 . S2CID 5287489 . Хэмп Т., Рост Б. (2015). «Эволюционные профили улучшают прогнозирование межбелковых взаимодействий на основе последовательности» . Биоинформатика . 31 (12): 1945–1950. doi : 10.1093/биоинформатика/btv077 . ПМИД 25657331 . Питре С., Хушьяр М., Шенрок А., Саманфар Б., Джессулат М., Грин Дж.Р., Дене Ф., Гольшани А. (2012). «Короткие совпадающие полипептидные области могут предсказывать глобальные карты взаимодействия белков» . Научные отчеты . 2 : 239. Бибкод : 2012NatSR...2E.239P . дои : 10.1038/srep00239 . ПМК 3269044 . ПМИД 22355752 . Питре С., Хушьяр М., Шенрок А., Саманфар Б., Джессулат М., Грин Дж.Р., Дене Ф., Гольшани А. (2012). «Короткие совпадающие полипептидные области могут предсказывать глобальные карты взаимодействия белков» . Научные отчеты . 2 : 239. Бибкод : 2012NatSR...2E.239P . дои : 10.1038/srep00239 . ПМК 3269044 . ПМИД 22355752 .
- ^ Jump up to: а б Ци Ю, Дхиман Х.К., Бхола Н., Будяк И., Кар С., Ман Д., Дутта А., Тирупула К., Карр Б.И., Грандис Дж., Бар-Джозеф З., Кляйн-Ситхараман Дж. (декабрь 2009 г.). «Систематическое предсказание взаимодействий мембранных рецепторов человека» . Протеомика . 9 (23): 5243–55. дои : 10.1002/pmic.200900259 . ПМК 3076061 . ПМИД 19798668 .
- ^ Ци Ю, Бар-Джозеф З., Кляйн-Ситхараман Дж. (май 2006 г.). «Оценка различных биологических данных и методы вычислительной классификации для использования в прогнозировании взаимодействия белков» . Белки . 63 (3): 490–500. дои : 10.1002/прот.20865 . ПМК 3250929 . ПМИД 16450363 .
- ^ Хоффманн, Р; Краллингер, М; Андрес, Э; Тамамес, Дж; Блашке, К; Валенсия, А (2005). «Интеллектуальный анализ текста для метаболических путей, сигнальных каскадов и белковых сетей». Научная сигнализация . 2005 (283): пе21. дои : 10.1126/stke.2832005pe21 . ПМИД 15886388 . S2CID 15301069 .
- ^ Jump up to: а б с Швиковски, Б.; Утц, П.; Филдс, С. (2000). «Сеть белок-белковых взаимодействий у дрожжей». Природная биотехнология . 18 (12): 1257–1261. дои : 10.1038/82360 . ПМИД 11101803 . S2CID 3009359 .
- ^ Макдермотт Дж., Бамгарнер Р.Э., Самудрала Р. (2005). «Функциональная аннотация предсказанных сетей взаимодействия белков» . Биоинформатика . 21 (15): 3217–3226. doi : 10.1093/биоинформатика/bti514 . ПМИД 15919725 .
- ^ Раджагопала, СВ; Сикорский, П.; Кофилд, Дж. Х.; Товчигречко А.; Утц, П. (2012). «Изучение белковых комплексов по двугибридной системе дрожжей» . Методы . 58 (4): 392–399. дои : 10.1016/j.ymeth.2012.07.015 . ПМЦ 3517932 . ПМИД 22841565 .
- ^ Деслер С., Замбах С., Сураваджхала П., Расмуссен Л.Дж. (2014). «Представляем гипотезу: способ интеграции предсказанных белков в интерактомы». Международный журнал исследований и приложений в области биоинформатики . 10 (6): 647–52. дои : 10.1504/IJBRA.2014.065247 . ПМИД 25335568 .
- ^ Бараб А.-Л.; Олтвай, З. (2004). «Сетевая биология: понимание функциональной организации клетки». Обзоры природы Генетика . 5 (2): 101–113. дои : 10.1038/nrg1272 . ПМИД 14735121 . S2CID 10950726 .
- ^ Гох, К. -И.; Цой, И. -Г. (2012). «Изучение болезней человека: сеть болезней человека» . Брифинги по функциональной геномике . 11 (6): 533–542. дои : 10.1093/bfgp/els032 . ПМИД 23063808 .
- ^ Jump up to: а б Барабаси, Алабама; Гулбахче, Н.; Лоскальцо, Дж (2011). «Сетевая медицина: сетевой подход к болезням человека» . Обзоры природы Генетика . 12 (1): 56–68. дои : 10.1038/nrg2918 . ПМК 3140052 . ПМИД 21164525 .
- ^ Альберт-Ласло Барабаши и Золтан Н. Олтваи (февраль 2004 г.). «Сетевая биология: понимание функциональной организации клетки». Обзоры природы. Генетика . 5 (2): 101–113. дои : 10.1038/nrg1272 . ПМИД 14735121 . S2CID 10950726 .
- ^ Гао, Л.; Сан, ПГ; Сонг, Дж. (2009). «Алгоритмы кластеризации для обнаружения функциональных модулей в сетях взаимодействия белков». Журнал биоинформатики и вычислительной биологии . 7 (1): 217–242. дои : 10.1142/S0219720009004023 . ПМИД 19226668 .
- ^ Навратил В.; и др. (2009). «VirHostNet: база знаний для управления и анализа общепротеомных сетей взаимодействия вируса и хоста» . Нуклеиновые кислоты Рез . 37 (Проблема с базой данных): D661–8. дои : 10.1093/нар/gkn794 . ПМЦ 2686459 . ПМИД 18984613 .
- ^ Раджагопала СВ.; и др. (2011). «Карта белкового взаимодействия бактериофага лямбда» . БМК Микробиол . 11 : 213. дои : 10.1186/1471-2180-11-213 . ПМЦ 3224144 . ПМИД 21943085 .
- ^ Бартель П.Л., Рокляйн Дж.А., СенГупта Д., Филдс С. (1996). «Карта белковых связей бактериофага T7 Escherichia coli». Нат. Жене . 12 (1): 72–7. дои : 10.1038/ng0196-72 . ПМИД 8528255 . S2CID 37155819 .
- ^ Сабри М.; и др. (2011). «Аннотация генома и внутривирусный интерактом Streptococcus pneumoniae вирулентного фага Dp-1 » . Дж. Бактериол . 193 (2): 551–62. дои : 10.1128/JB.01117-10 . ПМК 3019816 . ПМИД 21097633 .
- ^ Хойзер Р.; и др. (2011). «Протеом и интерактом Streptococcus pneumoniae фага Cp-1 » . Дж. Бактериол . 193 (12): 3135–8. дои : 10.1128/JB.01481-10 . ПМК 3133188 . ПМИД 21515781 .
- ^ Стеллбергер, Т.; и др. (2010). «Улучшение двугибридной системы дрожжей с помощью пермутированных слитых белков: интерактом вируса Varicella Zoster» . Протеомная наука . 8 :8. дои : 10.1186/1477-5956-8-8 . ПМЦ 2832230 . ПМИД 20205919 .
- ^ Кумар, К.; Рана, Дж.; Шриджит, Р.; Габрани, Р.; Шарма, СК; Гупта, А.; Чаудхари, ВК; Гупта, С. (2012). «Интравирусные белковые взаимодействия вируса Чандипура». Архив вирусологии . 157 (10): 1949–1957. дои : 10.1007/s00705-012-1389-5 . ПМИД 22763614 . S2CID 17714252 .
- ^ Jump up to: а б с д Фоссум, Э; и др. (2009). Сан, Рен (ред.). «Эволюционно консервативные сети взаимодействия герпесвирусных белков» . ПЛОС Патог . 5 (9): e1000570. дои : 10.1371/journal.ppat.1000570 . ПМЦ 2731838 . ПМИД 19730696 .
- ^ Хаген, Н.; Байер, К; Роеш, К; Шиндлер, М. (2014). «Сеть внутривирусного взаимодействия белков вируса гепатита С» . Молекулярная и клеточная протеомика . 13 (7): 1676–89. дои : 10.1074/mcp.M113.036301 . ПМК 4083108 . ПМИД 24797426 .
- ^ Хан, Ю; Ню, Дж; Ван, Д; Ли, Ю (2016). «Анализ сети взаимодействия белков вируса гепатита С на основе гепатоцеллюлярной карциномы» . ПЛОС ОДИН . 11 (4): e0153882. Бибкод : 2016PLoSO..1153882H . дои : 10.1371/journal.pone.0153882 . ПМЦ 4846009 . ПМИД 27115606 .
- ^ Остерман А., Стеллбергер Т., Гебхардт А., Курц М., Фридель К.С., Утц П., Ничко Х., Байкер А., Визосо-Пинто М.Г. (2015). «Интравирусный интерактом вируса гепатита Е» . Научный представитель . 5 : 13872. Бибкод : 2015NatSR...513872O . дои : 10.1038/srep13872 . ПМЦ 4604457 . ПМИД 26463011 .
- ^ Рейн, Джей Си; Селиг, Л.; Де Рёз, Х.; Батталья, VR; Реверди, CL; Саймон, СП; Ленцен, Г.; Петель, Ф.; Войчик, JRM; Шехтер, В.; Чемама, Ю.; Лабинь, AS; Легрен, П. (2001). «Карта белок-белкового взаимодействия Helicobacter pylori». Природа . 409 (6817): 211–215. Бибкод : 2001Natur.409..211R . дои : 10.1038/35051615 . ПМИД 11196647 . S2CID 4400094 .
- ^ Хойзер, Р; Сеол, А; Раджагопала, СВ; Моска, Р; Сислер, Г; Вермке, Н; Сикорский, П; Шварц, Ф; Шик, М; Вухти, С; Элой, П; Утц, П. (2014). «Сеть белок-белкового взаимодействия второго поколения Helicobacter pylori» . Молекулярная и клеточная протеомика . 13 (5): 1318–29. дои : 10.1074/mcp.O113.033571 . ПМК 4014287 . ПМИД 24627523 .
- ^ Пэрриш, младший; и др. (2007). «Карта взаимодействия белков по всему протеому для Campylobacter jejuni» . Геном Биол . 8 (7): 130 рандов. дои : 10.1186/gb-2007-8-7-r130 . ПМЦ 2323224 . ПМИД 17615063 .
- ^ Jump up to: а б Раджагопала, СВ; Титц, БР; Голл, Дж.; Хойзер, Р.; Маккевитт, Монтана; Палцкилл, Т.; Утц, П. (2008). Холл, Нил (ред.). «Бинарный белковый интерактом бледной трепонемы – спирохеты сифилиса» . ПЛОС ОДИН . 3 (5): e2292. Бибкод : 2008PLoSO...3.2292T . дои : 10.1371/journal.pone.0002292 . ПМК 2386257 . ПМИД 18509523 .
- ^ Ху, П; и др. (2009). Левченко, Андре (ред.). «Глобальный функциональный атлас Escherichia coli, включающий ранее не охарактеризованные белки» . ПЛОС Биол . 7 (4): е96. дои : 10.1371/journal.pbio.1000096 . ПМЦ 2672614 . ПМИД 19402753 .
- ^ Раджагопала, СВ; Сикорский, П; Кумар, А; Моска, Р; Власблом, Дж; Арнольд, Р; Франка-Ко, Дж; Пакала, СБ; Фанс, С; Сеол, А; Хойзер, Р; Сислер, Г; Вухти, С; Эмили, А; Бабу, М; Элой, П; Пипер, Р; Утц, П. (2014). «Ландшафт бинарного белок-белкового взаимодействия Escherichia coli» . Природная биотехнология . 32 (3): 285–90. дои : 10.1038/nbt.2831 . ПМК 4123855 . ПМИД 24561554 .
- ^ Симода, Ю.; Шинпо, С.; Кохара, М.; Накамура, Ю.; Табата, С.; Сато, С. (2008). «Крупномасштабный анализ белок-белковых взаимодействий у азотфиксирующей бактерии Mesorhizobium loti» . Исследование ДНК . 15 (1): 13–23. дои : 10.1093/dnares/dsm028 . ПМК 2650630 . ПМИД 18192278 .
- ^ Ван, Ю.; Кюи, Т.; Чжан, К.; Ян, М.; Хуанг, Ю.; Ли, В.; Чжан, Л.; Гао, К.; Привет.; Ли, Ю.; Хуанг, Ф.; Цзэн, Дж.; Хуанг, К.; Ян, К.; Тиан, Ю.; Чжао, К.; Чен, Х.; Чжан, Х.; Он, ЗГ (2010). «Глобальная сеть белок-белковых взаимодействий в патогене человека Mycobacterium TuberculosisH37Rv». Журнал исследований протеома . 9 (12): 6665–6677. дои : 10.1021/pr100808n . ПМИД 20973567 .
- ^ Кунер, С.; Ван Ноорт, В.; Беттс, MJ; Лео-Масиас, А.; Батисс, К.; Роде, М.; Ямада, Т.; Майер, Т.; Бадер, С.; Бельтран-Альварес, П.; Кастаньо-Дьес, Д.; Чен, В.-Х.; Девос, Д.; Гуэль, М.; Норамбуэна, Т.; Раке, И.; Рыбин В.; Шмидт, А.; Юс, Э.; Эберсольд, Р.; Херрманн, Р.; Бетчер, Б.; Франгакис, А.С.; Рассел, РБ; Серрано, Л.; Борк, П.; Гэвин, А.-К. (2009). «Организация протеома в бактерии с уменьшенным геномом». Наука . 326 (5957): 1235–1240. Бибкод : 2009Sci...326.1235K . дои : 10.1126/science.1176343 . ПМИД 19965468 . S2CID 19334426 .
- ^ Сато, С.; Симода, Ю.; Мураки, А.; Кохара, М.; Накамура, Ю.; Табата, С. (2007). «Крупномасштабный анализ взаимодействия белков в Synechocystis sp. PCC6803» . Исследование ДНК . 14 (5): 207–216. дои : 10.1093/dnares/dsm021 . ПМЦ 2779905 . ПМИД 18000013 .
- ^ Черкасов А; Хсинг, М; Зораги, Р; Фостер, LJ; Видите ли, Р.Х.; Стойнов Н.; Цзян, Дж; Каур, С; Лиан, Т; Джексон, Л; Гонг, Х; Суэйзи, Р; Амандорон, Э; Хормоздиари, Ф; Дао, П; Сахинальп, К; Сантос-Фильо, О; Аксерио-Силиес, П; Байлер, К; Макмастер, WR; Брунэм, Колорадо; Финли, BB; Райнер, Нью-Йорк (2011). «Картирование сети взаимодействия белков у метициллин-резистентного золотистого стафилококка ». Журнал исследований протеома . 10 (3): 1139–50. дои : 10.1021/pr100918u . ПМИД 21166474 .
- ^ Утц, П.; Джот, Л.; Кэгни, Г.; Мэнсфилд, штат Техас; Джадсон, РС; Найт, младший; Локшон, Д.; Нараян, В. (2000). «Комплексный анализ белок-белковых взаимодействий у Saccharomyces cerevisiae». Природа . 403 (6770): 623–627. Бибкод : 2000Natur.403..623U . дои : 10.1038/35001009 . ПМИД 10688190 . S2CID 4352495 .
- ^ Кроган, Нью-Джерси; и др. (2006). «Глобальный ландшафт белковых комплексов дрожжей Saccharomyeses Cerivisiae ». Природа . 440 (7084): 637–643. Бибкод : 2006Natur.440..637K . дои : 10.1038/nature04670 . ПМИД 16554755 . S2CID 72422 .
- ^ Панкальди В., Сарач О.С., Раллис С., Маклин Дж.Р., Пржеворовский М., Гулд К., Бейер А., Бэлер Дж. (2012). «Прогнозирование сети взаимодействия белков делящихся дрожжей» . G3: Гены, геномы, генетика . 2 (4): 453–67. дои : 10.1534/g3.111.001560 . ПМЦ 3337474 . ПМИД 22540037 .
- ^ Во, ТВ; и др. (2016). «Интерактом делящихся дрожжей по всему протеому раскрывает принципы эволюции сети от дрожжей к человеку» . Клетка . 164 (1–2): 310–323. дои : 10.1016/j.cell.2015.11.037 . ПМЦ 4715267 . ПМИД 26771498 .
- ^ де Шасси Б; Навратил V; Таффоро Л; и др. (04.11.2008). «Белковая сеть инфекции вируса гепатита С» . Молекулярная системная биология . 4 (4): 230. doi : 10.1038/msb.2008.66 . ПМК 2600670 . ПМИД 18985028 .
- ^ Навратил V; де Шасси Б; и др. (05.11.2010). «Сравнение белок-белковых взаимодействий между вирусами и сетью систем интерферона I типа человека на системном уровне». Журнал исследований протеома . 9 (7): 3527–36. дои : 10.1021/pr100326j . ПМИД 20459142 .
- ^ Браун КР, Юрисица I (2005). «Онлайн-база данных прогнозируемых человеческих взаимодействий» . Биоинформатика . 21 (9): 2076–82. doi : 10.1093/биоинформатика/bti273 . ПМИД 15657099 .
- ^ Гу Х, Чжу П, Цзяо Ю, Мэн Ю, Чен М (2011). «PRIN: предсказанная сеть интерактомов риса» . БМК Биоинформатика . 12 :161. дои : 10.1186/1471-2105-12-161 . ПМК 3118165 . ПМИД 21575196 .
- ^ Го Дж, Ли Х, Чанг Дж. В., Лэй Ю, Ли С, Чен Л. Л. (2013). «Прогнозирование и характеристика сети белок-белкового взаимодействия у Xanthomonas oryzae pv. oryzae PXO99 A» . Рез. Микробиол . 164 (10): 1035–44. дои : 10.1016/j.resmic.2013.09.001 . ПМИД 24113387 .
- ^ Гейслер-Ли Дж., О'Тул Н., Аммар Р., Проварт Н.Дж., Миллар А.Х., Гейслер М. (2007). «Предсказанный интерактом арабидопсиса» . Физиол растений . 145 (2): 317–29. дои : 10.1104/стр.107.103465 . ПМК 2048726 . ПМИД 17675552 .
- ^ Хуан, Шэнсюн; Тан, Сяофэн; Лю, Юншэн (01.01.2016 Юэ, Цзюнь, Вэй ; ) . .6 . : 25047. Бибкод : 2016NatSR...625047Y . doi : srep25047 . ISSN 2045-2322 . PMC 4848565. 10.1038 PMID 27121261 /
- ^ Ян, Цзяньхуа; Осман, Ким; Икбал, Мудассар; Штекель, Дов Дж.; Ло, Цзевэй; Армстронг, Сьюзен Дж.; Франклин, Ф. Крис Х. (1 января 2012 г.). «Вывод об интерактоме Brassica rapa с использованием данных межбелкового взаимодействия Arabidopsis thaliana» . Границы в науке о растениях . 3 : 297. doi : 10.3389/fpls.2012.00297 . ISSN 1664-462X . ПМЦ 3537189 . ПМИД 23293649 .
- ^ Цзиндун; Чэнь, Луонань; 01 февраля 2016 г. Чжу, Гуанхуэй; Сюй, Синь-Цзянь; Лу, Ле , База данных белок-белковых взаимодействий кукурузы» . Физиология растений . 170 (2): 618–626. : 10.1104 /pp.15.01821 . ISSN 1532-2548 . PMC 4734591. . PMID 26620522 doi
- ^ Роджерс-Мельник, Эли; Калп, Марк; ДиФазио, Стивен П. (01 января 2013 г.). «Прогнозирование сетей взаимодействия полногеномных белков на основе данных о первичных последовательностях в модельных и немодельных организмах с использованием ENTS» . БМК Геномика . 14 :608. дои : 10.1186/1471-2164-14-608 . ISSN 1471-2164 . ПМЦ 3848842 . ПМИД 24015873 .
- ^ Jump up to: а б Гуцци П.Х., Меркателли Д., Сераоло С., Джорджи Ф.М. (2020). «Основной регуляторный анализ взаимодействия SARS-CoV-2 и человека» . Журнал клинической медицины . 9 (4): 982–988. дои : 10.3390/jcm9040982 . ПМК 7230814 . ПМИД 32244779 .
- ^ Хан, доктор юридических наук; Бертен, Н; Хао, Т; Гольдберг, Д.С.; Берриз, Г.Ф.; Чжан, Л.В.; Дюпюи, Д; Уолхаут, Эй Джей; Кьюсик, Мэн; Рот, ФП; Видаль, М. (2004). «Доказательства динамически организованной модульности в сети межбелковых взаимодействий дрожжей». Природа . 430 (6995): 88–93. Бибкод : 2004Natur.430...88H . дои : 10.1038/nature02555 . ПМИД 15190252 . S2CID 4426721 .
- ^ Jump up to: а б Ю, Х; Браун, П; Йылдирим, Массачусетс; Лемменс, Я; Венкатесан, К; Сахали, Дж; Хиродзанэ-Кисикава, Т; Гебреаб, Ф; Ли, Н; Симонис, Н; Хао, Т; Руаль, Дж. Ф.; Дрико, А; Васкес, А; Мюррей, РР; Саймон, К; Тардиво, Л; Тэм, С; Сврзикапа, Н; Фан, С; Де Смет, А.С.; Мотыль, А; Хадсон, Мэн; Парк, Дж; Синь, Х; Кьюсик, Мэн; Мур, Т; Бун, К; Снайдер, М; Рот, ФП (2008). «Высококачественная карта взаимодействия бинарных белков сети интерактома дрожжей» . Наука . 322 (5898): 104–10. Бибкод : 2008Sci...322..104Y . дои : 10.1126/science.1158684 . ПМЦ 2746753 . ПМИД 18719252 .
- ^ Батада, штат Нью-Йорк; Регуля, Т; Брейткройц, А; Баучер, Л; Брейткройц, Б.Дж.; Херст, Л.Д.; Тайерс, М. (2006). «Слоистые, а не высококучевые: новый взгляд на сеть взаимодействия дрожжевых белков» . ПЛОС Биология . 4 (10): е317. дои : 10.1371/journal.pbio.0040317 . ПМК 1569888 . ПМИД 16984220 .
- ^ Бертен, Н; Симонис, Н; Дюпюи, Д; Кьюсик, Мэн; Хан, доктор юридических наук; Фрейзер, HB; Рот, ФП; Видаль, М. (2007). «Подтверждение организованной модульности в интерактоме дрожжей» . ПЛОС Биология . 5 (6): е153. doi : 10.1371/journal.pbio.0050153 . ПМК 1892830 . ПМИД 17564493 .
- ^ Ким, премьер-министр; Лу, Эл Джей; Ся, Ю; Герштейн, МБ (2006). «Связь трехмерных структур с белковыми сетями дает эволюционное понимание». Наука . 314 (5807): 1938–41. Бибкод : 2006Sci...314.1938K . дои : 10.1126/science.1136174 . ПМИД 17185604 . S2CID 2489619 .
- ^ Фернандес, А; М. Линч (2011). «Неадаптивные причины сложности интерактома» . Природа . 474 (7352): 502–505. дои : 10.1038/nature09992 . ПМК 3121905 . ПМИД 21593762 .
- ^ Уэлч, Дж. Рики (январь 2009 г.). «Нечеткий интерактом». Тенденции биохимических наук . 34 (1): 1–2. дои : 10.1016/j.tibs.2008.10.007 . ПМИД 19028099 .
Дальнейшее чтение
[ редактировать ]- Парк Дж., Лаппе М., Тейхманн С.А. (март 2001 г.). «Картирование взаимодействий семейства белков: репертуар внутримолекулярных и межмолекулярных взаимодействий семейства белков в PDB и дрожжах». Дж Мол Биол . 307 (3): 929–38. дои : 10.1006/jmbi.2001.4526 . ПМИД 11273711 .
Внешние ссылки
[ редактировать ]Веб-серверы Interactome
[ редактировать ]- Protinfo PPC предсказывает атомную трехмерную структуру белково-белковых комплексов. Киттихотират В., Геркен М., Бамгарнер Р., Самудрала Р. (2009). «Protinfo PPC: веб-сервер для прогнозирования белковых комплексов на атомном уровне» . Исследования нуклеиновых кислот . 37 (проблема с веб-сервером): W519–W525. дои : 10.1093/нар/gkp306 . ПМК 2703994 . ПМИД 19420059 .
- IBIS (сервер) сообщает, прогнозирует и интегрирует несколько типов консервативных взаимодействий белков.
Инструменты визуализации Interactome
[ редактировать ]- GPS-Prot Веб-визуализация данных о взаимодействии белков
- PINV - Визуализатор сети взаимодействия белков
Интерактомные базы данных
[ редактировать ]- БиоГРИД База данных
- Mentha — браузер Interactome Кальдероне; и др. (2013). «mentha: ресурс для просмотра интегрированных сетей взаимодействия белков» . Природные методы . 10 (8): 690–691. дои : 10.1038/nmeth.2561 . ПМИД 23900247 . S2CID 9733108 .
- IntAct: база данных молекулярных взаимодействий
- Interactome.org — специализированный интерактивный веб-сайт.