Вирус гепатита С
| Гепацивирус С | |
|---|---|
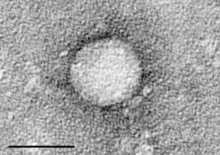 | |
| Электронная микрофотография гепацивируса С, выделенного из культуры клеток. Масштабная линейка = 50 нанометров | |
| Классификация вирусов | |
| (без рейтинга): | Вирус |
| Область : | Рибовирия |
| Королевство: | Орторнавиры |
| Тип: | Китриновирикота |
| Сорт: | Фласувирицеты |
| Заказ: | Желтовирусы |
| Семья: | Флавивирусиды |
| Род: | гепацивирус |
| Разновидность: | Гепацивирус С |
| Синонимы [1] [2] | |
Вирус гепатита С | |
Вирус гепатита С ( HCV ) [3] представляет собой небольшой ( 55–65 нм размером ) оболочечный РНК с положительным смыслом с одноцепочечной вирус из семейства Flaviviridae . Вирус гепатита С является причиной гепатита С и некоторых видов рака, таких как рак печени ( гепатоцеллюлярная карцинома , сокращенно ГЦК) и лимфомы у людей. [4] [5]
Таксономия
[ редактировать ]Вирус гепатита С принадлежит к роду Hepacivirus , члену семейства Flaviviridae . До 2011 года он считался единственным представителем этого рода. был обнаружен представитель этого рода Однако у собак : собачий гепацивирус . [6] Существует также по крайней мере один вирус этого рода, поражающий лошадей. [7] Несколько дополнительных вирусов этого рода были описаны у летучих мышей и грызунов. [8] [9]
Структура
[ редактировать ]
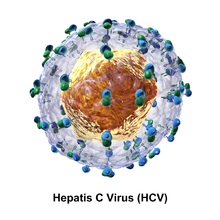
Частица вируса гепатита С состоит из липидной мембранной оболочки диаметром от 55 до 65 нм. [10] [11] вирусной оболочки Два гликопротеина , Е1 и Е2 , встроены в липидную оболочку. [12] Они принимают участие в прикреплении вируса и проникновении в клетку. [10] Внутри оболочки находится икосаэдрическое ядро диаметром от 33 до 40 нм. [11] Внутри ядра находится РНК-материал вируса. [10]
Гликопротеины Е1 и Е2
[ редактировать ]E1 и E2 соединяются ковалентно при внедрении в оболочку ВГС и стабилизируются дисульфидными связями . E2 имеет глобулярную форму и, судя по изображениям электронного микроскопа, кажется, выступает на 6 нм из оболочки оболочки. [11]
Эти гликопротеины играют важную роль во взаимодействии гепатита С с иммунной системой. Гипервариабельная область , гипервариабельная область 1 (HVR1), находится на гликопротеине E2. [10] HVR1 гибок и вполне доступен для окружающих молекул. [13] HVR1 помогает E2 защитить вирус от иммунной системы. Он предотвращает захват CD81 соответствующего рецептора вируса. [13] Кроме того, E2 может защитить E1 от иммунной системы. [13] Хотя аминокислотная последовательность HVR1 весьма вариабельна, эта область имеет схожие химические, физические и конформационные характеристики во многих гликопротеинах E2. [14]
Геном
[ редактировать ]
Вирус гепатита С имеет с положительной смысловой одноцепочечной РНК геном . Геном состоит из одной открытой рамки считывания длиной 9600 нуклеотидных оснований. [15] Эта единственная открытая рамка считывания транслируется для получения одного белкового продукта, который затем подвергается дальнейшей обработке для получения более мелких активных белков. Вот почему в общедоступных базах данных, таких как Европейский институт биоинформатики , вирусный протеом состоит только из 2 белков. [ нужна ссылка ]
На 5'- и 3'-концах РНК находятся нетранслируемые области (UTR), которые не транслируются в белки, но важны для трансляции и репликации вирусной РНК. 5'-UTR имеет с рибосомой. сайт связывания [16] или внутренний сайт входа в рибосому (IRES), который инициирует трансляцию очень длинного белка, содержащего около 3000 аминокислот. Основной домен IRES HCV содержит четырехстороннее спиральное соединение Холлидея , которое интегрировано в предполагаемый псевдоузел . [17] Конформация этого корового домена ограничивает ориентацию открытой рамки считывания для позиционирования на 40S рибосомальной субъединице . Большой пре-белок позже расщепляется клеточными и вирусными протеазами на 10 более мелких белков, которые обеспечивают репликацию вируса внутри клетки-хозяина или собираются в зрелые вирусные частицы. [18] Структурные белки, вырабатываемые вирусом гепатита С, включают Core-белок, E1 и E2; неструктурные белки включают NS2 , NS3 , NS4A , NS4B , NS5A и NS5B . [ нужна ссылка ]
Молекулярная биология
[ редактировать ]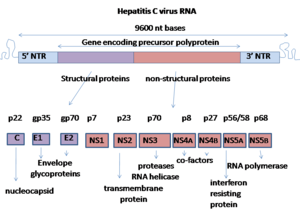
Этот раздел нуждается в дополнительных цитатах для проверки . ( Май 2016 г. ) |
Белки этого вируса расположены по геному в следующем порядке: N-конец-ядро-оболочка (E1)–E2–p7-неструктурный белок 2 (NS2)–NS3–NS4A–NS4B–NS5A–NS5B–C-терминал. Генерация зрелых неструктурных белков (от NS2 до NS5B) зависит от активности вирусных протеиназ. [19] Соединение NS2/NS3 расщепляется металлозависимой автокаталитической протеиназой, кодируемой NS2 и N-концом NS3. Остальные расщепления ниже этого сайта катализируются сериновой протеазой, также содержащейся в N-концевой области NS3.
- Коровый белок содержит 191 аминокислоту и по гидрофобности может быть разделен на три домена: домен 1 (остатки 1–117) содержит в основном основные остатки с двумя короткими гидрофобными участками ; домен 2 (остатки 118–174) менее основной и более гидрофобный, его С-конец находится на конце р21; домен 3 (остатки 175–191) обладает высокой гидрофобностью и действует как сигнальная последовательность для белка оболочки E1.
- Оба белка оболочки (E1 и E2) сильно гликозилированы и важны для проникновения в клетку. Е1 служит слияющей субъединицей, а Е2 действует как белок, связывающий рецептор. E1 имеет 4–5 N-связанных гликанов, а E2 — 11 сайтов N-гликозилирования.
- Белок NS1 (p7) необходим для репликации вирусного генома, но играет решающую роль в морфогенезе вируса. Этот белок представляет собой трансмембранный белок из 63 аминокислот, который локализуется в эндоплазматическом ретикулуме . Расщепление р7 опосредуется сигнальными пептидазами эндоплазматического ретикулума . Два трансмембранных домена р7 соединены цитоплазматической петлей и ориентированы в сторону просвета эндоплазматической сети.
- Белок NS2 представляет собой трансмембранный белок массой 21–23 килодальтон (кДа) с протеазной активностью.
- NS3 представляет собой белок массой 67 кДа, N-конец которого обладает активностью сериновой протеазы, а С-конец обладает активностью НТФазы / хеликазы . Он расположен внутри эндоплазматической сети и образует гетеродимерный комплекс с NS4A — мембранным белком из 54 аминокислот, который действует как кофактор протеиназы.
- NS4A — мембранный белок из 54 аминокислот, который действует как кофактор протеиназы.
- NS4B представляет собой небольшой (27 кДа) гидрофобный интегральный мембранный белок с четырьмя трансмембранными доменами. Он расположен внутри эндоплазматической сети и играет важную роль в привлечении других вирусных белков. Он вызывает морфологические изменения в эндоплазматической сети, образуя структуру, называемую мембранной сетью.
- NS5A представляет собой гидрофильный фосфопротеин, который играет важную роль в репликации вируса, модуляции клеточных сигнальных путей и интерфероновом ответе. Известно, что он связывается с белками VAP человека, заякоренными в эндоплазматическом ретикулуме . [20]
- Белок NS5B (65 кДа) представляет собой вирусную РНК-зависимую РНК-полимеразу . NS5B выполняет ключевую функцию репликации вирусной РНК ВГС с использованием цепи вирусной положительной смысловой РНК в качестве матрицы и катализирует полимеризацию рибонуклеозидтрифосфатов (rNTP) во время репликации РНК . [21] [22] [23] Несколько кристаллических структур полимеразы NS5B в нескольких кристаллических формах были определены на основе одной и той же консенсусной последовательности BK (HCV-BK, генотип 1). [24] Структуру можно представить в виде правой руки с пальцами, ладонью и большим пальцем. Окруженный активный сайт , уникальный для NS5B, содержится в структуре белка «ладошки». Недавние исследования структуры белка NS5B генотипа 1b штамма J4 (HC-J4) указывают на наличие активного центра, где возможен контроль связывания нуклеотидов и инициация синтеза РНК de novo. De-novo добавляет необходимые праймеры для инициации репликации РНК. [25] Текущие исследования пытаются связать структуры с этим активным сайтом, чтобы изменить его функциональность и предотвратить дальнейшую репликацию вирусной РНК. [26]
Также описан 11-й белок. [27] [28] Этот белок кодируется сдвигом рамки +1 в капсидном гене. Похоже, что он антигенен , но его функция неизвестна.
Репликация
[ редактировать ]
Репликация ВГС включает несколько этапов. Вирус размножается главным образом в , где гепатоцитах печени , по оценкам, ежедневно каждая инфицированная клетка производит около пятидесяти вирионов (вирусных частиц), при этом общее количество генерируемых вирионов составляет один триллион. Вирус также может реплицироваться в мононуклеарных клетках периферической крови , что потенциально является причиной высокого уровня иммунологических нарушений, обнаруживаемых у хронически инфицированных пациентов с ВГС. В печени частицы ВГС попадают в печеночные синусоиды с током крови. Эти синусоиды соседствуют с клетками гепатоцитов. [10] ВГС способен проходить через эндотелий синусоидов и достигать базолатеральной поверхности клеток гепатоцитов. [10]
ВГС имеет большое разнообразие генотипов вируса и быстро мутирует из-за высокой частоты ошибок со стороны РНК-зависимой РНК-полимеразы . Скорость мутаций приводит к появлению такого количества вариантов вируса, что его считают квазивидом, а не обычным видом вируса. [29] Проникновение в клетки-хозяева происходит посредством сложных взаимодействий между вирионами, особенно через их гликопротеины, и молекулами клеточной поверхности CD81 , рецептором ЛПНП , SR-BI , DC-SIGN , Клаудином-1 и Окклюдином . [30] [31]
Оболочка ВГС аналогична липопротеинам очень низкой плотности (ЛПОНП) и липопротеинам низкой плотности (ЛПНП). [10] Из-за этого сходства считается, что вирус способен связываться с аполипопротеинами . Он может окружить себя липопротеинами, частично закрывающими Е1 и Е2. Недавние исследования показывают, что эти аполипопротеины взаимодействуют с рецептором-мусорщиком B1 (SR-B1). SR-B1 способен удалять липиды из липопротеинов вокруг вируса, чтобы обеспечить лучший контакт с HVR1. Клаудин 1, который представляет собой белок с плотными соединениями , и CD81 связываются, образуя комплекс, подготавливая их к последующим процессам заражения ВГС. Когда иммунная система активируется, макрофаги увеличивают количество TNF-α вокруг инфицированных гепатоцитов. Это запускает миграцию окклюдина, который представляет собой еще один комплекс плотных соединений, к базолатеральной мембране. Частица ВГС готова войти в клетку. [10]
Эти взаимодействия приводят к эндоцитозу вирусной частицы. Этому процессу способствуют белки клатрина. Попав внутрь ранней эндосомы, эндосома и вирусная оболочка сливаются, и РНК попадает в цитоплазму. [10]
ВГС берет на себя часть внутриклеточного механизма для репликации. [32] Геном ВГС транслируется с образованием одного белка, содержащего около 3011 аминокислот. Затем полипротеин подвергается протеолитическому процессингу вирусными и клеточными протеазами с образованием трех структурных (связанных с вирионами) и семи неструктурных (NS) белков. Альтернативно, сдвиг рамки считывания может произойти в области ядра для создания белка альтернативной рамки считывания (ARFP). [33] ВГС кодирует две протеазы: цистеиновую аутопротеазу NS2 и сериновую протеазу NS3-4A. Затем белки NS рекрутируют вирусный геном в комплекс репликации РНК, который связан с перестроенными цитоплазматическими мембранами. Репликация РНК происходит с помощью вирусной РНК-зависимой РНК-полимеразы NS5B, которая продуцирует промежуточную РНК с отрицательной цепью. РНК с отрицательной цепью затем служит матрицей для производства новых вирусных геномов с положительной цепью. Возникающие геномы затем можно транслировать, реплицировать или упаковывать в новые вирусные частицы. [ нужна ссылка ]
Вирус реплицируется на внутриклеточных липидных мембранах. [34] Эндоплазматическая сеть, в частности, деформируется в мембранные структуры уникальной формы, называемые «мембранозными сетями». Эти структуры могут быть индуцированы экспрессией только вирусного белка NS4B. [35] Коровый белок связывается с липидными каплями и использует микротрубочки и динеины, чтобы изменить их расположение на перинуклеарное распределение. [36] Высвобождение из гепатоцита может происходить по секреторному пути ЛПОНП. [37] Другая гипотеза утверждает, что вирусная частица может секретироваться из эндоплазматического ретикулума через эндосомальный сортировочный комплекс, необходимый для пути транспорта (ESCRT). [10] Этот путь обычно используется для выхода пузырьков из клетки. Единственным ограничением этой гипотезы является то, что этот путь обычно используется для отпочкования клеток , и неизвестно, как ВГС сможет использовать путь ESCRT для использования с эндоплазматическим ретикулумом. [10]
Генотипы
[ редактировать ]На основании генетических различий между изолятами ВГС виды вируса гепатита С подразделяются на шесть генотипов (1–6) с несколькими подтипами внутри каждого генотипа (обозначенными строчными буквами). [38] [39] Подтипы далее разбиваются на квазивиды в зависимости от их генетического разнообразия. Генотипы различаются на 30–35% нуклеотидных сайтов по всему геному. [40] Разница в геномном составе подтипов генотипа обычно составляет 20–25%. Подтипы 1a и 1b встречаются во всем мире и вызывают 60% всех случаев. [ нужна ссылка ]
Клиническое значение
[ редактировать ]Части этой статьи (относящиеся к противовирусным препаратам прямого действия) необходимо обновить . ( январь 2024 г. ) |
Генотип клинически важен для определения потенциального ответа на терапию интерфероном и необходимой продолжительности такой терапии. Генотипы 1 и 4 менее чувствительны к лечению интерфероном, чем другие генотипы (2, 3, 5 и 6). [41] Продолжительность стандартной терапии интерфероном для генотипов 1 и 4 составляет 48 недель, тогда как для генотипов 2 и 3 лечение завершается за 24 недели. Устойчивый вирусологический ответ наблюдается в 70% случаев генотипа 1, ~90% генотипов 2 и 3, ~65% генотипа 4 и ~80% генотипа 6. [42] Кроме того, люди африканского происхождения гораздо реже реагируют на лечение при инфицировании генотипами 1 или 4. [43] Предполагается, что значительная часть отсутствия ответа на лечение вызвана однонуклеотидным полиморфизмом (SNP) на хромосоме 19 генома человека, который является предиктором успеха лечения. [44] Генотипы ВГС 1 и 4 были эндемично распространены в перекрывающихся районах Западной и Центральной Африки, на протяжении веков заражая человеческие популяции, несущие рассматриваемый генетический полиморфизм. Это побудило ученых предположить, что длительное сохранение генотипов 1 и 4 ВГС у людей африканского происхождения является эволюционной адаптацией ВГС на протяжении многих столетий к иммуногенетическим реакциям этих популяций. [45]
Заражение одним генотипом не дает иммунитета против других, возможно одновременное заражение двумя штаммами. В большинстве таких случаев один из штаммов за короткое время превосходит другой. Это открытие может быть полезно при лечении, при замене штаммов, не реагирующих на лекарства, другими, которые легче поддаются лечению. [46]
Рекомбинация
[ редактировать ]Когда два вируса заражают одну и ту же клетку, генетическая рекомбинация . может произойти [47] Хотя и нечасто, рекомбинация ВГС наблюдалась между разными генотипами, между подтипами одного и того же генотипа и даже между штаммами одного и того же подтипа. [47]
Эпидемиология
[ редактировать ]Вирус гепатита С преимущественно передается через кровь , с очень низким риском передачи половым или вертикальным путем . [48] Из-за этого пути распространения основными группами риска являются потребители внутривенных наркотиков (ПИН), реципиенты продуктов крови и иногда пациенты, находящиеся на гемодиализе . Распространенной средой передачи ВГС также является внутрибольничная ( нозокомиальная правила гигиены и стерилизации . ) передача, когда в клинике не соблюдаются [49] В качестве потенциального исторического способа распространения ВГС был предложен ряд культурных или ритуальных практик, включая обрезание, калечащие операции на половых органах, ритуальное скарификацию, традиционное нанесение татуировок и иглоукалывание. [48] Утверждалось также, что, учитывая чрезвычайно длительные периоды персистенции ВГС у людей, даже очень низкие и необнаружимые уровни механической передачи через кусающих насекомых могут быть достаточными для поддержания эндемической инфекции в тропиках, где люди получают большое количество укусов насекомых. [50]
Эволюция
[ редактировать ]Идентифицировать происхождение этого вируса было сложно, но генотипы 1 и 4, по-видимому, имеют общее происхождение. [51] Байесовский анализ показывает , что основные генотипы разошлись примерно 300–400 лет назад от общего предка вируса. [52] Минорные генотипы разошлись около 200 лет назад от своих основных генотипов. Все существующие генотипы, по-видимому, произошли от генотипа 1 подтипа 1b. [ нужна ссылка ]
Изучение штаммов генотипа 6 предполагает более раннюю дату эволюции: примерно от 1100 до 1350 лет до настоящего времени . [53] Предполагаемая частота мутаций составила 1,8 × 10. −4 . Экспериментальное исследование оценило частоту мутаций в 2,5–2,9 × 10. −3 базовых замен на сайт в год. [54] Этот генотип может быть предком других генотипов. [53]
Исследование изолятов из Европы, США и Японии показало, что дата происхождения генотипа 1b приходится примерно на 1925 год. [55] Предполагаемые даты возникновения типов 2а и 3а — 1917 и 1943 годы соответственно. Время дивергенции типов 1а и 1б оценивалось в 200–300 лет. [ нужна ссылка ]
Исследование генотипов 1a и 1b показало, что даты происхождения — 1914–1930 годы для типа 1a и 1911–1944 годы для типа 1b. [56] В период с 1940 по 1960 годы эффективная численность популяции обоих типов 1a и 1b претерпела массовое увеличение. Распространение подтипа 1b ВГС предшествовало распространению подтипа 1a как минимум на 16 лет. Оба типа, по-видимому, распространились из развитого мира в развивающийся.
Штаммы генотипа 2 из Африки можно разделить на четыре клады, которые коррелируют со страной их происхождения: (1) Камерун и Центральноафриканская Республика (2) Бенин, Гана и Буркина-Фасо (3) Гамбия, Гвинея, Гвинея-Бисау и Сенегал ( 4) Мадагаскар. [57] Имеются также убедительные доказательства распространения генотипа 2 ВГС из Западной Африки в страны Карибского бассейна в результате трансатлантической работорговли . [58]
Считается, что генотип 3 возник в Юго-Восточной Азии. [59]
Эти данные из разных стран позволяют предположить, что этот вирус, возможно, развился в Юго-Восточной Азии и был распространен в Западной Африке торговцами из Западной Европы. [60] Позже он был завезен в Японию, когда в этой стране была снята добровольная изоляция . После завоза в страну на его распространение повлияло множество местных факторов, включая переливание крови, программы вакцинации, внутривенное употребление наркотиков и режимы лечения. Учитывая снижение темпов распространения после того, как в 1990-х годах был введен скрининг ВГС в продуктах крови, может показаться, что ранее переливание крови было важным методом распространения. Требуется дополнительная работа для определения дат эволюции различных генотипов и сроков их распространения по земному шару. [ нужна ссылка ]
Вакцинация
[ редактировать ]В отличие от гепатита А и В, в настоящее время не существует вакцины для предотвращения заражения гепатитом С. [61]
Текущие исследования
[ редактировать ] этой статьи Фактическая точность может быть нарушена из-за устаревшей информации . Причина такова: в результате медицинских исследований ВГС теперь излечим: https://web.archive.org/web/20180919174730/http://www.who.int/hepatitis/news-events/direct-acting- противовирусное лечение-гепатит-c/en/ . ( ноябрь 2019 г. ) |
Изучение ВГС затруднено из-за узкого круга хозяев ВГС. [62] Использование репликонов оказалось успешным, но они были обнаружены лишь недавно. [63] ВГС, как и большинство РНК-вирусов, существует как квазивид вируса , что очень затрудняет выделение отдельного штамма или типа рецептора для изучения. [64] [65]
Текущие исследования сосредоточены на низкомолекулярных ингибиторах вирусной протеазы , РНК-полимеразы и других неструктурных генов. Два препарата — боцепревир от Merck [66] и телапревир от Vertex Pharmaceuticals — оба ингибитора протеазы NS3 были одобрены к использованию 13 мая 2011 г. и 23 мая 2011 г. соответственно.
о возможной связи между низким уровнем витамина D и плохим ответом на лечение. Сообщалось [67] [68] [69] [70] Исследования in vitro показали, что витамин D может снижать репликацию вируса. [71] Хотя эта работа выглядит многообещающе [72] [73] ожидаются результаты клинических испытаний. [74] [75] Однако было высказано предположение, что добавление витамина D важно в дополнение к стандартному лечению, чтобы улучшить ответ на лечение. [76]
Было показано, что нарингенин , флавоноид, содержащийся в грейпфрутах и других фруктах и травах, блокирует сборку внутриклеточных инфекционных вирусных частиц, не влияя на внутриклеточные уровни вирусной РНК или белка. [76]
Другие агенты, находящиеся на стадии исследования, включают нуклеозидные и нуклеотидные аналоги- ингибиторы и ненуклеозидные ингибиторы РНК-зависимой РНК-полимеразы, ингибиторы NSP5A и соединения, нацеленные на хозяина, такие как ингибиторы циклофилина и силибинин . [77]
Софосбувир для лечения хронического гепатита С был одобрен FDA 6 декабря 2013 года. Сообщается, что это первый препарат, продемонстрировавший безопасность и эффективность для лечения определенных типов инфекции ВГС без необходимости одновременного применения интерферона. . [78] 22 ноября FDA одобрило симепревира применение в сочетании с пегинтерфероном-альфа и рибавирином . [79] Симепревир одобрен в Японии для лечения хронического гепатита С генотипа 1. [80]
В настоящее время также проводятся экспериментальные исследования немедикаментозных методов лечения. Оксиматрин , например, представляет собой экстракт корня, обнаруженный на азиатском континенте, который, как сообщается, обладает противовирусной активностью против ВГС в клеточных культурах и исследованиях на животных. Небольшие и многообещающие испытания на людях показали положительные результаты и отсутствие серьезных побочных эффектов, но они были слишком малы, чтобы обобщать выводы. [76]
5 октября 2020 года было объявлено, что Харви Дж. Альтер , Майкл Хоутон и Чарльз М. Райс были удостоены Нобелевской премии 2020 года по физиологии и медицине за открытие ВГС. [81]
См. также
[ редактировать ]- Заболевания, передающиеся через кровь
- Вирус рака
- Открытие и разработка ингибиторов NS5A
- ВГС ИРЭС
- Вирус гепатита С «стебель-петля» VII
- Элемент 3'X вируса гепатита С
- Цис-действующий репликационный элемент вируса гепатита С (HCV) (CRE)
- Инфекция, передающаяся половым путем
Ссылки
[ редактировать ]- ^ « Гепацивирус С » . Национальный центр биотехнологической информации (NCBI) . Проверено 3 января 2022 г.
- ^ Смит, Дональд Б.; и др. (23 июня 2016 г.). «Создайте 13 новых видов рода Hepacivirus и переименуйте 1 вид (семейство Flaviviridae )» (PDF) . Международный комитет по таксономии вирусов (ICTV) . Проверено 13 марта 2019 г.
- ^ «Род: Гепацивирус» . Международный комитет по таксономии вирусов . Июль 2018. Архивировано из оригинала 19 февраля 2020 года.
- ^ Ферри, Клодовео (2015). «Синдром ВГС: совокупность органо- и неорганоспецифических аутоиммунных заболеваний, В-клеточной неходжкинской лимфомы и рака» . Всемирный журнал гепатологии . 7 (3): 327–43. дои : 10.4254/wjh.v7.i3.327 . ISSN 1948-5182 . ПМЦ 4381161 . ПМИД 25848462 .
- ^ Русин I, Лимон С.М. (2014). «Механизмы рака печени, вызванного ВГС: что мы узнали из исследований in vitro и на животных?» . Рак Летт . 345 (2): 210–5. дои : 10.1016/j.canlet.2013.06.028 . ПМК 3844040 . ПМИД 23871966 .
- ^ Капур А. и др. (2011). «Характеристика собачьего гомолога вируса гепатита С» . Proc Natl Acad Sci США . 108 (28): 11608–13. Бибкод : 2011PNAS..10811608K . дои : 10.1073/pnas.1101794108 . ПМК 3136326 . ПМИД 21610165 .
- ^ Бурбело П.Д., Дубови Э.Дж., Симмондс П. и др. (июнь 2012 г.). «Обнаружение генетически разнообразных гепацивирусов у нового хозяина с помощью серологии» . Дж. Вирол . 86 (11): 6171–8. дои : 10.1128/JVI.00250-12 . ПМК 3372197 . ПМИД 22491452 .
- ^ Цюань П.Л., Ферт С., Конте Дж.М. и др. (май 2013 г.). «Летучие мыши являются основным естественным резервуаром гепацивирусов и пегивирусов» . Учеб. Натл. акад. наук. США . 110 (20): 8194–9. Бибкод : 2013PNAS..110.8194Q . дои : 10.1073/pnas.1303037110 . ПМЦ 3657805 . ПМИД 23610427 .
- ^ Капур А., Симмондс П., Шил Т.К. и др. (2013). «Идентификация гомологов вируса гепатита С и пегивирусов у грызунов» . мБио 4 (2):e00216–13. дои : 10.1128/mBio.00216-13 . ПМЦ 3622934 . ПМИД 23572554 .
- ^ Jump up to: а б с д и ж г час я дж к Дюбюиссон, Жан; Коссе, Франсуа-Лоик (2014). «Вирусология и клеточная биология жизненного цикла вируса гепатита С – обновленная информация» . Журнал гепатологии . 61 (1): С3–С13. дои : 10.1016/j.jhep.2014.06.031 . hdl : 20.500.12210/28720 . ПМИД 25443344 .
- ^ Jump up to: а б с Кайто, Масахико; Исида, Сатоши; Танака, Хидеаки; Хориике, Шиничиро; Фудзита, Наоки; Адачи, Юкихико; Кохара, Мичинори; Кониси, Масаеши; Ватанабэ, Сёдзо (июнь 2006 г.). «Морфология частиц вируса гепатита С и гепатита В, обнаруженная с помощью электронной микроскопии иммунозолота» . Медицинская молекулярная морфология . 39 (2): 63–71. дои : 10.1007/s00795-006-0317-8 . ISSN 1860-1480 . ПМИД 16821143 . S2CID 24668769 .
- ^ Оп Де Бек А, Дюбюиссон Дж (2003). «Топология гликопротеинов оболочки вируса гепатита С». Преподобный Мед. Вирол . 13 (4): 233–41. дои : 10.1002/rmv.391 . ПМИД 12820185 . S2CID 22280227 .
- ^ Jump up to: а б с Кастелли, Маттео; Клементи, Никола; Пфафф, Дженнифер; Саутто, Джузеппе А.; Диотти, Роберта А.; Буриони, Роберто ; Доранц, Бенджамин Дж.; Даль Пераро, Маттео; Клементи, Массимо (16 марта 2017 г.). «Биологически подтвержденная структурная модель гетеродимера E1E2 HCV» . Научные отчеты . 7 (1): 214. Бибкод : 2017НатСР...7..214С . дои : 10.1038/s41598-017-00320-7 . ISSN 2045-2322 . ПМЦ 5428263 . ПМИД 28303031 .
- ^ Басу, Арнаб; Бейене, Астер; Мейер, Кейт; Рэй, Ранджит (май 2004 г.). «Гипервариабельный участок 1 гликопротеина Е2 вируса гепатита С связывается с гликозаминогликанами, но это связывание не приводит к инфекции в псевдотипической системе» . Журнал вирусологии . 78 (9): 4478–4486. doi : 10.1128/JVI.78.9.4478-4486.2004 . ISSN 0022-538X . ПМЦ 387685 . ПМИД 15078928 .
- ^ Като Н (2000). «Геном вируса гепатита С человека (ВГС): организация генов, разнообразие последовательностей и вариации». Микроб. Комп. Геном . 5 (3): 129–51. дои : 10.1089/mcg.2000.5.129 . ПМИД 11252351 .
- ^ Джубин Р. (2001). «IRES гепатита С: перевод перевода в терапевтическую цель». Курс. Мнение. Мол. Там . 3 (3): 278–87. ПМИД 11497352 .
- ^ Берри К.Э., Вагрей С., Мортимер С.А., Бай Ю., Дудна Дж.А. (октябрь 2011 г.). «Кристаллическая структура центрального домена IRES вируса гепатита С раскрывает стратегию позиционирования стартовых кодонов» . Структура . 19 (10): 1456–66. дои : 10.1016/j.str.2011.08.002 . ПМК 3209822 . ПМИД 22000514 .
- ^ Дюбюиссон Дж (2007). «Белки вируса гепатита С» . Мир Дж. Гастроэнтерол . 13 (17): 2406–15. дои : 10.3748/wjg.v13.i17.2406 . ПМК 4146758 . ПМИД 17552023 .
- ^ Де Франческо Р. (1999). «Молекулярная вирусология вируса гепатита С». Дж Гепатол . 31 (Приложение 1): 47–53. дои : 10.1016/S0168-8278(99)80374-2 . ПМИД 10622560 .
- ^ Гупта Дж., Цинь Х., Сонг Дж. (2012). «По сути неструктурированный домен 3 вируса гепатита С NS5A образует «нечеткий комплекс» с доменом VAPB-MSP, который несет мутации, вызывающие БАС» . ПЛОС ОДИН . 7 (6): e39261. Бибкод : 2012PLoSO...739261G . дои : 10.1371/journal.pone.0039261 . ПМЦ 3374797 . ПМИД 22720086 .
- ^ Джин, З; Левек, В; Ма, Х; Джонсон, Калифорния; Клампп, К. (2012). «Сборка, очистка и предстационарный кинетический анализ активного РНК-зависимого комплекса элонгации РНК-полимеразы» . Журнал биологической химии . 287 (13): 10674–83. дои : 10.1074/jbc.M111.325530 . ПМК 3323022 . ПМИД 22303022 .
- ^ Морадпур Д., Пенин Ф., Райс К.М. (июнь 2007 г.). «Репликация вируса гепатита С». Нат. Преподобный Микробиол . 5 (6): 453–63. дои : 10.1038/nrmicro1645 . ПМИД 17487147 . S2CID 13176201 .
- ^ Ригат К., Ван Й., Худыма Т.В. и др. (ноябрь 2010 г.). «Лиганд-индуцированные изменения в структуре полимеразы NS5B вируса гепатита С». Противовирусный Рес . 88 (2): 197–206. дои : 10.1016/j.antiviral.2010.08.014 . ПМИД 20813137 .
- ^ Бисвал Б.К., Черней М.М., Ван М. и др. (май 2005 г.). «Кристаллические структуры РНК-зависимой РНК-полимеразы генотипа 2а вируса гепатита С обнаруживают две конформации и предполагают механизмы ингибирования ненуклеозидными ингибиторами» . Ж. Биол. Хим . 280 (18): 18202–10. дои : 10.1074/jbc.M413410200 . ПМИД 15746101 .
- ^ О'Фаррелл Д., Троубридж Р., Роулендс Д., Джагер Дж. (февраль 2003 г.). «Субстратные комплексы РНК-полимеразы вируса гепатита С (HC-J4): структурные доказательства импорта нуклеотидов и инициации de novo». Дж. Мол. Биол . 326 (4): 1025–35. дои : 10.1016/s0022-2836(02)01439-0 . ПМИД 12589751 .
- ^ Бисвал Б.К., Ван М., Черный М.М. и др. (август 2006 г.). «Ненуклеозидные ингибиторы, связывающиеся с полимеразой NS5B вируса гепатита С, открывают новый механизм ингибирования». Дж. Мол. Биол . 361 (1): 33–45. дои : 10.1016/j.jmb.2006.05.074 . ПМИД 16828488 .
- ^ Валевски Дж.Л., Келлер Т.Р., Стамп Д.Д., Филиал А.Д. (2001). «Доказательства существования нового антигена вируса гепатита С, закодированного в перекрывающейся рамке считывания» . РНК . 7 (5): 710–721. дои : 10.1017/S1355838201010111 . ПМК 1370123 . ПМИД 11350035 .
- ^ Багбани-арани Ф., Рухванд Ф., Агасадеги М.Р., Эйди А., Амини С., Мотевалли Ф., Садат С.М., Мемарнеджадиан А., Халили Г. и др. (2012). «Экспрессия и характеристика белка ARFP/F вируса гепатита С, полученного из Escherichia coli». Мол Биол (Москва) . 46 (2): 251–9. дои : 10.1134/S0026893312020033 . ПМИД 22670521 . S2CID 7379944 .
- ^ Бартеншлагер Р., Ломанн В. (июль 2000 г.). «Репликация вируса гепатита С» . Дж. Генерал Вирол . 81 (Часть 7): 1631–48. CiteSeerX 10.1.1.319.8775 . doi : 10.1099/0022-1317-81-7-1631 (неактивен 7 февраля 2024 г.). ПМИД 10859368 . Архивировано из оригинала 3 декабря 2009 г. Проверено 16 июля 2010 г.
{{cite journal}}: CS1 maint: DOI неактивен по состоянию на февраль 2024 г. ( ссылка ) - ^ Зейзель, М.; Барт, Х.; Шустер, К.; Баумерт, Т. (2009). «Проникновение вируса гепатита С: молекулярные механизмы и мишени противовирусной терапии» . Границы бионауки . 14 (8): 3274–3285. Бибкод : 2009CNSNS..14.3274H . дои : 10.1016/j.cnsns.2008.11.006 . ПМК 3235086 . ПМИД 19273272 .
- ^ Кохаар, И.; Плосс, А.; Король, Э.; Му, К.; Шоггинс, Дж.; О'Брайен, Т.; Райс, К.; Прокунина-Олссон, Л. (2010). «Разнообразие сплайсинга человеческого гена OCLN и его биологическое значение для проникновения вируса гепатита С» . Журнал вирусологии . 84 (14): 6987–6994. дои : 10.1128/JVI.00196-10 . ПМЦ 2898237 . ПМИД 20463075 .
- ^ Линденбах Б., Райс С. (2005). «Распутывание репликации вируса гепатита С из генома в функцию» . Природа . 436 (7053): 933–8. Бибкод : 2005Natur.436..933L . дои : 10.1038/nature04077 . ПМИД 16107832 .
- ^ Филиал, АД; Стамп, Д.Д.; Гутьеррес, Дж.А.; Энг, Ф.; Валевски, Дж. Л. (2005). «Альтернативная рамка считывания вируса гепатита С (ARF) и семейство ее новых продуктов: белок альтернативной рамки считывания/F-белок, белок с двойным сдвигом рамки и другие». Семинары по заболеваниям печени . 25 (1): 105–117. дои : 10.1055/s-2005-864786 . ПМИД 15732002 . S2CID 260316882 .
- ^ Дюбюиссон Дж., Пенин Ф., Морадпур Д. (2002). «Взаимодействие белков вируса гепатита С с мембранами и липидами клеток-хозяев». Тенденции клеточной биологии . 12 (11): 517–523. дои : 10.1016/S0962-8924(02)02383-8 . ПМИД 12446113 .
- ^ Эггер Д., Вёльк Б., Госерт Р., Бьянки Л., Блюм Х.Э., Морадпур Д., Биенц К. (2002). «Экспрессия белков вируса гепатита С вызывает отчетливые изменения мембран, включая потенциальный комплекс репликации вируса» . Дж Вирол . 76 (12): 5974–84. doi : 10.1128/JVI.76.12.5974-5984.2002 . ПМК 136238 . ПМИД 12021330 .
- ^ Булан С., Дуглас М.В., Муди Л., Будковска А., Таргетт-Адамс П., Маклаучлан Дж. (2008). «Коровой белок вируса гепатита С индуцирует перераспределение липидных капель зависимым от микротрубочек и динеина способом» . Трафик . 9 (8): 1268–82. дои : 10.1111/j.1600-0854.2008.00767.x . ПМИД 18489704 . S2CID 20609887 .
- ^ Сайед Г.Х., Амако Ю., Сиддики А. (2010). «Вирус гепатита С нарушает липидный обмен хозяина» . Тенденции Метаб. эндокринолов . 21 (1): 33–40. дои : 10.1016/j.tem.2009.07.005 . ПМК 2818172 . ПМИД 19854061 .
- ^ Симмондс П., Холмс ЕС, Ча Т.А. и др. (ноябрь 1993 г.). «Классификация вируса гепатита С на шесть основных генотипов и ряд подтипов с помощью филогенетического анализа региона NS-5» (PDF) . Дж. Генерал Вирол . 74 (Часть 11): 2391–9. CiteSeerX 10.1.1.325.7888 . дои : 10.1099/0022-1317-74-11-2391 . PMID 8245854 . S2CID 17597460 . Проверено 10 июля 2020 г.
- ^ Накано, Тацунори; Лау, Джиллиан М.Г.; Лау, Грейс М.Л.; Сугияма, Масая; Мизоками, Масаси (9 октября 2011 г.). «Обновленный анализ генотипов и подтипов вируса гепатита С на основе полной кодирующей области». Печень Интернационал . 32 (2): 339–45. дои : 10.1111/j.1478-3231.2011.02684.x . ПМИД 22142261 . S2CID 23271017 .
- ^ Оно О, Мизоками М, Ву РР, Салех М.Г., Охба К., Орито Э., Мукайде М., Уильямс Р., Лау Дж.И. и др. (2007). «Новая система генотипирования вируса гепатита С (ВГС), которая позволяет идентифицировать генотипы ВГС 1a, 1b, 2a, 2b, 3a, 3b, 4, 5a и 6a» . J Clin Микробиол . 35 (1): 201–7. doi : 10.1128/JCM.35.1.201-207.1997 . ПМК 229539 . ПМИД 8968908 .
- ^ Симмондс П; Бух Дж; Комбет С; Делеаж Ж; Эномото Н; Фейнстон С; Халфон П; Инчауспе Г; Куйкен С; Мартенс Г; Мизоками М; Мерфи, генеральный директор; Окамото, Х; Павлоцкий, Дж. М.; Пенин, Ф; Саблон, Э; Шин-И, Т; Стайвер, LJ; Тиль, Х.Дж.; Вязов С; Вайнер, Эй Джей; Уайделл, А. (2005). «Консенсусные предложения по единой системе номенклатуры генотипов вируса гепатита С» . Гепатология . 42 (4): 962–73. дои : 10.1002/hep.20819 . ПМИД 16149085 . S2CID 21393716 .
- ^ Ю М.Л., Чуанг В.Л. (2009). «Лечение хронического гепатита С в Азии: когда Восток встречается с Западом» . J Гастроэнтерол Гепатол . 24 (3): 336–345. дои : 10.1111/j.1440-1746.2009.05789.x . ПМИД 19335784 . S2CID 27333980 .
- ^ Мьюир, Эй Джей; Борнштейн, доктор медицинских наук; Килленберг, П.Г.; Группа по лечению гепатита Атлантического побережья (2004 г.). «Пегинтерферон альфа-2b и рибавирин для лечения хронического гепатита С у чернокожих и белых неиспаноязычных народов» . N Engl J Med . 350 (22): 2265–71. doi : 10.1056/NEJMoa032502 . ПМИД 15163776 . Ошибка: дои : 10.1056/nejm200409163511229
- ^ Ге, Д; Феллай, Дж; Томпсон, Эй Джей; Саймон, С.Дж.; Шианна, КВ; Урбан, Техас; Хейнцен, Эль; и др. (2009). «Генетическая вариация IL28B предсказывает клиренс вируса, вызванный лечением гепатита С». Природа . 461 (7262): 399–401. Бибкод : 2009Natur.461..399G . дои : 10.1038/nature08309 . ПМИД 19684573 . S2CID 1707096 .
- ^ Роуз, Р; Марков П.В.; Лам, ТТ; Пибус, О.Г. (2013). «Эволюция вируса объясняет связь между генотипом вируса гепатита С, клиническими исходами и генетическими вариациями человека». Заразить Генет Эвол . 20 : 418–21. дои : 10.1016/j.meegid.2013.09.029 . hdl : 10722/221827 . ПМИД 24140473 .
- ^ Ласкус Т., Ван Л.Ф., Радковски М., Варгас Х., Новицкий М., Уилкинсон Дж., Ракела Дж. (2001). «Воздействие РНК-положительных реципиентов вируса гепатита С (ВГС) на РНК-положительных доноров крови ВГС приводит к быстрому преобладанию одного донорского штамма и исключению и/или подавлению штамма-реципиента» . Журнал вирусологии . 75 (5): 2059–66. doi : 10.1128/JVI.75.5.2059-2066.2001 . ПМК 114790 . ПМИД 11160710 .
- ^ Jump up to: а б Гонсалес-Канделас Ф., Лопес-Лабрадор Ф.Х., Брачо М.А. (октябрь 2011 г.). «Рекомбинация вируса гепатита С» . Вирусы . 3 (10): 2006–24. дои : 10.3390/v3102006 . ПМК 3205392 . ПМИД 22069526 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Jump up to: а б Шепард, CW; Финелли, Л; Альтер, MJ (сентябрь 2005 г.). «Глобальная эпидемиология инфекции вируса гепатита С» . Ланцет Инфекционный Дис . 5 (9): 558–67. дои : 10.1016/S1473-3099(05)70216-4 . ПМИД 16122679 .
- ^ Альтер, MJ (ноябрь 2011 г.). «Пути передачи ВГС: что происходит, то и происходит». Семин Дис печени . 31 (4): 340–6. дои : 10.1055/s-0031-1297923 . ПМИД 22189974 . S2CID 260310609 .
- ^ Пибус, Огайо; Марков П.В.; Ву, А; Татем, AJ (июль 2007 г.). «Исследование эндемической передачи вируса гепатита С». Инт Дж Паразитол . 37 (8–9): 839–49. дои : 10.1016/j.ijpara.2007.04.009 . ПМИД 17521655 .
- ^ Салеми М., Вандам А.М. (2002). «Эволюционные закономерности вируса гепатита С изучены посредством анализа полногеномных последовательностей» . Джей Мол Эвол . 54 (1): 62–70. Бибкод : 2002JMolE..54...62S . дои : 10.1007/s00239-001-0018-9 . ПМИД 11734899 . S2CID 35899454 .
- ^ Сарвар М.Т. и др. (2011). «Белок NS4A как маркер истории ВГС позволяет предположить, что различные генотипы ВГС первоначально произошли от генотипа 1b» . Вирол. Дж . 8 : 317. дои : 10.1186/1743-422X-8-317 . ПМК 3145594 . ПМИД 21696641 .
- ^ Jump up to: а б Пайбус О.Г., Барнс Э., Таггарт Р., Леми П., Марков П.В., Расачак Б., Сихавонг Б., Петсувана Р., Шеридан И. и др. (2009). «Генетическая история вируса гепатита С в Восточной Азии» . Дж Вирол . 83 (2): 1071–82. дои : 10.1128/JVI.01501-08 . ПМЦ 2612398 . ПМИД 18971279 .
- ^ Като Н., Уэда Ю., Седзима Х., Гу В., Сато С., Дансако Х., Икеда М., Шимотохно К. (2019) Исследование множественных генетических вариаций, вызванных стойкой репликацией вируса гепатита С в долгосрочной культуре клеток. Арка Вирол
- ^ Симмондс П., Смит Д.Б. (1997). «Исследование характера разнообразия вируса гепатита С в зависимости от времени передачи». Журнал вирусного гепатита . 4 (Приложение 1): 69–74. дои : 10.1111/j.1365-2893.1997.tb00163.x . ПМИД 9097281 . S2CID 41594303 .
- ^ Магиоркинис Г., Магиоркинис Е., Параскевис Д. и др. (декабрь 2009 г.). «Глобальное распространение вируса гепатита С 1а и 1b: филодинамический и филогеографический анализ» . ПЛОС Мед . 6 (12): e1000198. doi : 10.1371/journal.pmed.1000198 . ПМЦ 2795363 . ПМИД 20041120 .
- ^ Марков П.В., Пепин Дж., Фрост Э., Десландес С., Лаббе А.С., Пибус О.Г. (сентябрь 2009 г.). «Филогеография и молекулярная эпидемиология генотипа 2 вируса гепатита С в Африке» . Дж. Генерал Вирол . 90 (Часть 9): 2086–96. дои : 10.1099/vir.0.011569-0 . ПМИД 19474244 .
- ^ Марков П.В.; ван де Лаар, Ти Джей; Томас, XV; Аронсон, С.Дж.; Вигинк, CJ; ван ден Берк, GE; Принс, М.; и др. (2012). «Колониальная история и современная передача формируют генетическое разнообразие генотипа 2 вируса гепатита С в Амстердаме» . Дж Вирол . 86 (14): 7677–7687. дои : 10.1128/JVI.06910-11 . ПМК 3416291 . ПМИД 22573865 .
- ^ Симмондс П. (ноябрь 2004 г.). «Генетическое разнообразие и эволюция вируса гепатита С — 15 лет спустя» . Дж. Генерал Вирол . 85 (Часть 11): 3173–88. дои : 10.1099/vir.0.80401-0 . ПМИД 15483230 .
- ^ Симмондс П. (2001). «Реконструкция происхождения вирусов гепатита человека» . Philos Trans R Soc Lond B Biol Sci . 356 (1411): 1013–26. дои : 10.1098/rstb.2001.0890 . ПМЦ 1088496 . ПМИД 11516379 .
- ^ Ю КИ, Чан Б.Л. (2010). «Новый взгляд на разработку вакцины против гепатита С» . Дж. Биомед. Биотехнология . 2010 : 1–12. дои : 10.1155/2010/548280 . ПМЦ 2896694 . ПМИД 20625493 .
- ^ Раух, А.; Гаудиери, С.; Тио, К.; Бочуд, П.Я. (2009). «Генетические детерминанты спонтанного клиренса гепатита С». Фармакогеномика . 10 (11): 1819–1837. дои : 10.2217/стр.09.121 . ПМИД 19891557 .
- ^ Мейер В., Рамадори Дж. (апрель 2009 г.). «Вирусология вируса гепатита С и новые цели лечения». Expert Rev Anti Infect Ther . 7 (3): 329–50. дои : 10.1586/eri.09.12 . ПМИД 19344246 . S2CID 38411966 .
- ^ Маннс М.П., Фостер Г.Р., Рокстро Дж.К., Зейзем С., Зулим Ф., Хоутон М. (декабрь 2007 г.). «Путь вперед в лечении ВГС – поиск правильного пути» . Nat Rev Drug Discov . 6 (12): 991–1000. дои : 10.1038/nrd2411 . ПМИД 18049473 . S2CID 52874660 .
- ^ Ахмед, Али Махмуд; Дохайм, Мохамед Фахми; Маттар, Омар Мохамед; Шериф, Нурин Али; Труонг, Дуй Хиеу; Фам ТЛ, Хоа; Хираяма, Кендзи; Хай, Нгуен Тьен (май 2018 г.). «Беклабувир в сочетании с асунапревиром и даклатасвиром при инфекции генотипа 1 вируса гепатита С: систематический обзор и метаанализ». Джей Мед Вирол . 90 (5): 907–918. дои : 10.1002/jmv.24947 . ПМИД 28892235 . S2CID 3829214 .
- ^ «FDA одобрило препарат Виктрелис для лечения гепатита С» (пресс-релиз) . FDA. 13 мая 2011 г.
- ^ Гутьеррес Х.А., Парих Н., Филиал AD (2011). «Классическая и новая роль витамина D при инфекции вируса гепатита С» . Семин Дис печени . 31 (4): 387–398. дои : 10.1055/s-0031-1297927 . ПМК 4107414 . ПМИД 22189978 .
- ^ Ланге С.М., Боджунга Дж., Рамос-Лопес Е., фон Вагнер М., Хасслер А., Вермерен Дж., Херрманн Е., Баденхуп К., Зейзем С. и др. (2011). «Дефицит витамина D и полиморфизм промотора CYP27B1-1260 связаны с хроническим гепатитом С и плохим ответом на терапию на основе интерферона-альфа» . Дж Гепатол . 54 (5): 887–893. дои : 10.1016/j.jhep.2010.08.036 . ПМИД 21145801 . Проверено 10 июля 2020 г.
- ^ Баур К., Мертенс Дж.К., Шмитт Дж. и др. (2012). «Гаплотип гена рецептора витамина D bAt (CCA) ухудшает ответ на терапию на основе пегилированного интерферона / рибавирина у пациентов с хроническим гепатитом С» . Антивирь. Там . 17 (3): 541–7. дои : 10.3851/IMP2018 . ПМИД 22300961 . S2CID 32175340 .
- ^ Битетто Д., Фаттович Г., Фабрис С., Чериани Е., Фаллети Е., Форнасьере Е., Пасино М., Иелузи Д., Кассиг А. и др. (2011). «Дополнительная роль дефицита витамина D и полиморфизма интерлейкина-28B rs12979860 C/T в прогнозировании противовирусного ответа при хроническом гепатите С» . Гепатология . 53 (4): 1118–26. дои : 10.1002/hep.24201 . ПМИД 21480318 . S2CID 5329252 .
- ^ Гал-Танами М, Бахметов Л, Равид А, Корен Р, Эрман А, Тур-Каспа Р, Земель Р (2011). «Витамин D: врожденный противовирусный агент, подавляющий вирус гепатита С в гепатоцитах человека» . Гепатология . 54 (5): 1570–9. дои : 10.1002/hep.24575 . ПМИД 21793032 . S2CID 10090454 .
- ^ Абу-Муш С., Пожарный З., Ярховский Дж., Зейна А.Р., Асси Н. (2011). «Добавка витамина D улучшает устойчивый вирусологический ответ у пациентов с хроническим гепатитом С (генотип 1), ранее не болевших» . Мир Дж Гастроэнтерол . 17 (47): 5184–90. дои : 10.3748/wjg.v17.i47.5184 . ПМЦ 3243885 . ПМИД 22215943 .
- ^ Битетто Д., Фабрис С., Форнасьер Е., Пипан С., Фумоло Е., Кассиг А., Бигнулин С., Cmet С., Фонтанини Е. и др. (2011). «Добавка витамина D улучшает ответ на противовирусное лечение рецидивирующего гепатита С». Транспл Инт . 24 (1): 43–50. дои : 10.1111/j.1432-2277.2010.01141.x . ПМИД 20649944 . S2CID 22124427 .
- ^ Чолонгитас Э., Теохариду Э., Гулис Дж., Цочацис Э., Акривиадис Э., Берроуз К. (март 2012 г.). «Обзорная статья: внескелетные эффекты витамина D при хроническом гепатите С» . Питание. Фармакол. Там . 35 (6): 634–46. дои : 10.1111/j.1365-2036.2012.05000.x . ПМИД 22316435 . S2CID 25534747 .
- ^ Какопардо Б, Камма С, Петта С, Пинцоне М.Р., Каппеллани А, Занги А, Николози А, Нуннари Г (2012). «Диагностическая и терапевтическая роль витамина D при хронической инфекции вируса гепатита С». Фронт Биосци . 1 (4): 1276–1286. дои : 10.2741/e458 . ПМИД 22201953 .
- ^ Jump up to: а б с Халегуа-Де Марцио, Дина; Фенкель, Джонатан (27 января 2014 г.). «Альтернативные лекарства при инфекции гепатита С» . Всемирный журнал гепатологии . 6 (1): 9–16. дои : 10.4254/wjh.v6.i1.9 . ПМЦ 3953807 . ПМИД 24653790 .
- ^ Саррацин С., Хезод С., Зейзем С., Павлоцкий Дж. М. (2012). «Противовирусные стратегии при инфекции вируса гепатита С». Дж. Гепатол . 56 (Приложение 1): С88–100. дои : 10.1016/S0168-8278(12)60010-5 . ПМИД 22300469 .
- ^ «Сообщение для прессы, FDA, 6 декабря 2013 г.» . Управление по контролю за продуктами и лекарствами .
- ^ «FDA одобряет новое лечение вируса гепатита С» . Управление по контролю за продуктами и лекарствами . 22 ноября 2013 г.
- ^ «Медивир: Симепревир был одобрен в Японии для лечения хронического гепатита С генотипа 1» . Уолл Стрит Джорнал. 27 сентября 2013 г.
- ^ «Нобелевская премия по физиологии и медицине 2020» . NobelPrize.org .
Внешние ссылки
[ редактировать ]- Научные статьи о шести генотипах ВГС. Архивировано 11 октября 2020 г. в Wayback Machine Clodovero Ferri.
- Базы данных последовательностей ВГС и иммунологии в Национальной лаборатории Лос-Аламоса
- База данных и аналитический ресурс вирусных патогенов (ViPR): Flaviviridae
| Базы данных органов управления : Национальные |
|---|