Холлидей-Джанкшн
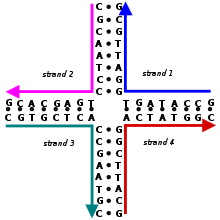
Соединение Холлидея представляет собой разветвленную структуру нуклеиновой кислоты , содержащую четыре соединенных двухцепочечных плеча. Эти плечи могут принимать одну из нескольких конформаций в зависимости от концентрации буферной соли и последовательности нуклеиновых оснований, ближайших к месту соединения. Структура названа в честь Робина Холлидея , молекулярного биолога , который предположил ее существование в 1964 году.
В биологии соединения Холлидея являются ключевым промежуточным звеном во многих типах генетической рекомбинации , а также в восстановлении двухцепочечных разрывов . Эти соединения обычно имеют симметричную последовательность и, таким образом, подвижны, а это означает, что четыре отдельных плеча могут скользить через соединение по определенному шаблону, который в значительной степени сохраняет спаривание оснований . РНК появляются четырехветвевые соединения, подобные соединениям Холлидея Кроме того, в некоторых функциональных молекулах .
Неподвижные соединения Холлидея с асимметричными последовательностями, которые фиксируют нити в определенном положении, были искусственно созданы учеными для изучения их структуры в качестве модели естественных соединений Холлидея. Эти соединения позже также нашли применение в качестве основных структурных строительных блоков в нанотехнологиях ДНК , где многочисленные соединения Холлидея могут быть объединены в определенные геометрические формы, которые обеспечивают молекулам высокую степень структурной жесткости .
Структура
[ редактировать ]

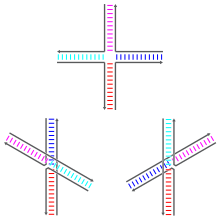
Соединения Холлидея могут существовать в различных конформационных изомерах с различными паттернами коаксиальной укладки между четырьмя двойными спиральными плечами. Коаксиальная укладка — это тенденция тупых концов нуклеиновой кислоты связываться друг с другом за счет взаимодействия между открытыми основаниями. Существует три возможных конформера: несложенная форма (или открытая X) и две сложенные формы. Несложенная форма доминирует в отсутствие двухвалентных катионов, таких как Mg. 2+ , из-за электростатического отталкивания между отрицательно заряженными основами нитей. В присутствии по меньшей мере примерно 0,1 мМ Mg 2+ , электростатическое отталкивание противодействует и преобладают многослойные структуры. По состоянию на 2000 год не было с уверенностью известно, является ли электростатическое экранирование результатом сайт-специфического связывания катионов с соединением или присутствия диффузного скопления ионов в растворе. [1]
Несложенная форма представляет собой почти квадратную плоскую вытянутую конформацию. С другой стороны, пакетированные конформеры имеют два непрерывных двухспиральных домена, разделенных углом около 60° в правом направлении. Две из четырех цепей остаются примерно спиральными, оставаясь внутри каждого из двух двуспиральных доменов, тогда как две другие пересекаются между двумя доменами антипараллельным образом . [1]
Две возможные сложенные формы различаются тем, какие пары рычагов сложены друг с другом; какой из двух доминирует, сильно зависит от последовательностей оснований, ближайших к стыку. Некоторые последовательности приводят к равновесию между двумя конформерами, в то время как другие категорически предпочитают один конформер. В частности, соединения, содержащие последовательность A-CC, соединяющую точку соединения, по-видимому, сильно предпочитают конформер, который позволяет образовывать водородную связь между вторым цитозином и одним из фосфатов в точке соединения. Хотя большинство исследований было сосредоточено на идентичности четырех оснований, ближайших к соединению на каждом плече, очевидно, что основания, расположенные дальше, также могут влиять на наблюдаемые конформации укладки. [1]
В соединениях с симметричными последовательностями точка ветвления подвижна и может мигрировать в процессе случайного блуждания . Скорость миграции ветвей резко варьируется в зависимости от концентрации ионов: время одного шага увеличивается от 0,3 до 0,4 мс при отсутствии ионов до 270–300 мс при 10 мМ Mg. 2+ . Изменение скорости коррелирует с образованием сложенных и неуложенных структур. [1]
Соединения Холлидея с надрезом или разрывом одной из нитей в точке соединения принимают перпендикулярную ориентацию и всегда отдают предпочтение укладочному конформеру, который помещает надрез на перекрестную нить, а не на спиральную нить. [1]
Соединения РНК Холлидея принимают антипараллельную стопочную конформацию при высоких концентрациях магния, перпендикулярную стопочную конформацию при умеренных концентрациях и вращаются в конформацию параллельной стопки при низких концентрациях, в то время как даже небольшие концентрации ионов кальция благоприятствуют антипараллельному конформеру. [1]
Биологическая функция
[ редактировать ]
Соединение Холлидея является ключевым промежуточным звеном в гомологичной рекомбинации , биологическом процессе, который увеличивает генетическое разнообразие путем перемещения генов между двумя хромосомами , а также сайт-специфической рекомбинации событий с участием интеграз . Они дополнительно участвуют в восстановлении двухцепочечных разрывов . [1] Кроме того, крестообразные структуры, включающие соединения Холлидея, могут возникать для снятия спирального напряжения в симметричных последовательностях суперспиралей ДНК . [2] Хотя четырехветвевые соединения также появляются в функциональных молекулах РНК, таких как сплайсосомальная РНК U1 и шпильковый рибозим вируса кольцевой пятнистости табака , они обычно содержат непарные нуклеотиды между парными двухспиральными доменами и, таким образом, не соответствуют строго структуре Холлидея. . [1]
Соединения Холлидея при гомологичной рекомбинации располагаются между идентичными или почти идентичными последовательностями, что приводит к симметричному расположению последовательностей вокруг центрального соединения. Это позволяет происходить процессу миграции ветвей , когда нити проходят через точку соединения. [1] Расщепление или разрешение соединения Холлидея может происходить двумя способами. Расщепление исходного набора цепей приводит к образованию двух молекул, которые могут демонстрировать генную конверсию , но не хромосомный кроссинговер , в то время как расщепление другого набора из двух нитей приводит к тому, что полученные рекомбинантные молекулы демонстрируют кроссинговер. Все продукты, независимо от спайности, представляют собой гетеродуплексы в области миграции соединения Холлидея. [3]
Многие белки способны распознавать или искажать структуру соединения Холлидея. Один из таких классов содержит ферменты, расщепляющие соединения , которые расщепляют соединения, иногда в зависимости от последовательности. Такие белки искажают структуру соединения различными способами, часто переводя соединение в несложенную конформацию, разрывая центральные пары оснований и/или изменяя углы между четырьмя плечами. Другие классы — это белки миграции ветвей, увеличивающие скорость обмена на порядки, и сайт-специфические рекомбиназы . [1] У прокариот резольвазы соединения Холлидея делятся на два семейства: интегразы и нуклеазы, каждое из которых структурно сходно, хотя их последовательности не консервативны. [3]
У эукариот двумя основными моделями того, как гомологичная рекомбинация восстанавливает двухцепочечные разрывы в ДНК, являются путь восстановления двухцепочечных разрывов (DSBR) (иногда называемый моделью двойного соединения Холлидея ) и путь синтез-зависимого отжига цепей (SDSA). [4] В случае двухцепочечного разрыва 3'-конец разрушается, а более длинный 5'-конец вторгается в соседнюю сестринскую хроматиду, образуя репликационный пузырь. Когда этот пузырь приближается к разорванной ДНК, более длинная 5'-антисмысловая цепь снова вторгается в смысловую цепь этой части ДНК, транскрибируя вторую копию. Когда репликация заканчивается, оба хвоста снова соединяются, образуя два соединения Холлидея, которые затем расщепляются белками по различным схемам. [5] Анимацию этого процесса можно увидеть здесь . [6]
Двухцепочечные разрывы ДНК у бактерий восстанавливаются с помощью RecBCD пути гомологичной рекомбинации . Считается, что разрывы, возникающие только на одной из двух цепей ДНК, известные как одноцепочечные разрывы, восстанавливаются с помощью пути RecF . Оба пути RecBCD и RecF включают серию реакций, известных как миграция ветвей, при которой отдельные цепи ДНК обмениваются между двумя перекрещенными молекулами дуплексной ДНК, и резолюция , при которой эти две перекрещенные молекулы ДНК разрезаются на части и восстанавливаются до нормального состояния. двухцепочечное состояние. [7] Гомологичная рекомбинация происходит у нескольких групп вирусов. В ДНК-вирусах, таких как вирус герпеса , рекомбинация происходит по механизму разрыва и воссоединения, как у бактерий и эукариот. [8] У бактерий миграции ветвей способствует комплекс RuvABC или белок RecG , молекулярные моторы, которые используют энергию гидролиза АТФ для перемещения соединения. Затем соединение должно быть разделено на два отдельных дуплекса, восстанавливая либо родительскую конфигурацию, либо перекрестную конфигурацию. Разрешение может происходить как горизонтально, так и вертикально во время гомологичной рекомбинации, образуя патч-продукты (если они имеют одинаковую ориентацию во время восстановления двухцепочечного разрыва) или продукты сплайсинга (если в разных ориентациях во время восстановления двухцепочечного разрыва). [9] [10] RuvA и RuvB представляют собой белки миграции ветвей, а RuvC представляет собой фермент, расщепляющий соединения. [1]
Имеются доказательства рекомбинации у некоторых РНК-вирусов , особенно у вирусов с оцРНК с положительным смыслом, таких как ретровирусы , пикорнавирусы и коронавирусы . Существуют разногласия по поводу того, происходит ли гомологичная рекомбинация у вирусов оцРНК с отрицательным смыслом, таких как грипп . [11]
Разрешение
[ редактировать ]У почкующихся дрожжей Saccharomyces cerevisiae соединения Холлидея могут разрешаться четырьмя различными путями, которые отвечают практически за все разрешение соединений Холлидея in vivo . [12] Путь, который вызывает большинство кроссинговеров у почкующихся дрожжей S. cerevisiae и, возможно, у млекопитающих, включает белки EXO1 , гетеродимер MLH1 - MLH3 (называемый MutL гамма) и SGS1 (ортолог геликазы синдрома Блума ). [12] Гетеродимер MLH1-MLH3 преимущественно связывается с соединениями Холлидея. [13] Это эндонуклеаза, которая делает одноцепочечные разрывы в суперскрученной двухцепочечной ДНК. [13] [14] Гетеродимер MLH1-MLH3 способствует образованию перекрестных рекомбинантов . [15] В то время как другие три пути, включающие белки MUS81 -MMS4, SLX1 и YEN1, соответственно, могут способствовать разрешению соединений Холлидея in vivo, отсутствие всех трех нуклеаз оказывает лишь умеренное влияние на образование продуктов кроссинговера.
Двойные мутанты, удаленные как по MLH3 (основной путь), так и по MMS4 (второстепенный путь), показали резкое снижение кроссинговера по сравнению с диким типом (в 6-17 раз); однако жизнеспособность спор была достаточно высокой (62%), а расхождение хромосом оказалось в основном функциональным. [15]
Хотя MUS81 является компонентом незначительного пути кроссовера в мейозе почкующихся дрожжей, растений и позвоночных, [16] у простейших Tetrahymena thermophila MUS81, по-видимому, является частью важного, если не преобладающего пути кроссинговера. [16] Путь MUS81 также, по-видимому, является преобладающим путем кроссовера у делящихся дрожжей Schizosaccharomyces pombe . [16]
Белки MSH4 и MSH5 образуют гетероолигомерную структуру (гетеродимер) у дрожжей и человека. [17] [18] [19] В дрожжах Saccharomyces cerevisiae MSH4 и MSH5 действуют специфически, облегчая кроссовер между гомологичными хромосомами во время мейоза. [17] Комплекс MSH4/MSH5 связывает и стабилизирует двойные соединения Холлидея и способствует их расщеплению в продукты кроссовера. Гипоморфный (частично функциональный) мутант MSH4 S. cerevisiae показал 30%-ное снижение числа кроссинговеров по всему геному и большое количество мейозов с необменными хромосомами. [20] Тем не менее, этот мутант дал начало моделям жизнеспособности спор, что позволяет предположить, что сегрегация необменных хромосом происходит эффективно. Таким образом, у S. cerevisiae правильная сегрегация, по-видимому, не полностью зависит от кроссоверов между гомологичными парами.
Использование в нанотехнологиях ДНК
[ редактировать ]
Нанотехнология ДНК — это разработка и производство структур искусственных нуклеиновых кислот в качестве инженерных материалов для нанотехнологий, а не носителей генетической информации в живых клетках. В этой области разветвленные структуры ДНК используются в качестве фундаментальных компонентов для создания более сложных, рационально спроектированных структур. Таким образом, соединения Холлидея являются компонентами многих таких структур ДНК. Поскольку изолированные комплексы соединений Холлидея слишком гибки, чтобы их можно было собрать в большие упорядоченные массивы, структурные мотивы с несколькими соединениями Холлидея используются для создания жестких « плиток », которые затем можно собрать в более крупные «массивы». [22] [23]

Наиболее распространенным таким мотивом является комплекс двойного кроссовера (DX), который содержит два соединения Холлидея, расположенные в непосредственной близости друг от друга, что приводит к образованию жесткой структуры, которая может самособираться в более крупные массивы. Структура молекулы DX заставляет соединения Холлидея принимать конформацию с двухспиральными доменами, расположенными рядом друг с другом, в отличие от их предпочтительного угла около 60 °. Комплекс может быть спроектирован таким образом, чтобы соединения переходили в параллельную или антипараллельную ориентацию, но на практике антипараллельный вариант ведет себя более хорошо, а параллельная версия используется редко. [22] [23]
Структурный мотив DX является фундаментальным строительным блоком метода ДНК-оригами , который используется для создания более крупных двух- и трехмерных структур произвольной формы. Вместо использования отдельных плиток DX одна длинная прядь каркаса сгибается до желаемой формы с помощью нескольких коротких штапельных прядей. При сборке нить каркаса непрерывна через двухспиральные домены, тогда как штапельные нити участвуют в соединениях Холлидея как кроссоверные нити. [25]
Были продемонстрированы некоторые типы плитки, которые сохраняют исходный угол развязки Холлидей в 60 °. В одном из таких массивов используются плитки, содержащие четыре соединения Холлидея в виде параллелограмма. Преимущество этой структуры заключалось в том, что угол перехода можно было непосредственно визуализировать с помощью атомно-силовой микроскопии . Плитки из трех контактов Холлидея в треугольной форме использовались для создания периодических трехмерных массивов для использования в рентгеновской кристаллографии биомолекул. Эти структуры названы в честь их сходства со структурными единицами, основанными на принципе тенсегрити , который использует элементы как при растяжении, так и при сжатии. [22] [23]
История
[ редактировать ]Робин Холлидей предложил структуру соединения, которая теперь носит его имя, как часть своей модели гомологичной рекомбинации в 1964 году, основываясь на своих исследованиях организмов Ustilago maydis и Saccharomyces cerevisiae . Модель предоставила молекулярный механизм, который объяснил как конверсию генов , так и хромосомный кроссинговер . Холлидей понял, что предложенный путь позволит создать гетеродуплексные сегменты ДНК с несоответствием оснований между различными версиями одного гена. Он предсказал, что в клетке будет механизм исправления несоответствий, который был позже обнаружен. [3] До модели Холлидея принятая модель включала механизм выбора копирования. [26] где новая цепь синтезируется непосредственно из частей разных родительских цепей. [27]
В исходной модели гомологичной рекомбинации Холлидея однонитевые разрывы происходят в одной и той же точке на одной цепи каждой родительской ДНК. Свободные концы каждой разорванной цепи затем мигрируют к другой спирали ДНК. Там вторгающиеся нити соединяются со свободными концами, с которыми они сталкиваются, образуя соединение Холлидея. По мере того как каждая кроссоверная цепь повторно отжигается со своей исходной партнерской цепью, она вытесняет исходную комплементарную цепь впереди себя. Это приводит к миграции соединения Холлидея, создавая гетеродуплексные сегменты. В зависимости от того, какая цепь использовалась в качестве матрицы для восстановления другой, четыре клетки, образующиеся в результате мейоза, могут оказаться с тремя копиями одного аллеля и только одной копией другого вместо обычных двух копий каждого. Это свойство известно как генная конверсия. . [3]
Первоначальная модель Холлидея предполагала, что гетеродуплексная ДНК будет присутствовать на обеих хромосомах, но экспериментальные данные на дрожжах опровергли это. Обновленная модель Мэтта Мезельсона и Чарли Рэддинга в 1975 году представила идею миграции ветвей. [26] Дальнейшие наблюдения в 1980-х годах привели к предложению альтернативных механизмов рекомбинации, таких как модель двухцепочечного разрыва (Джека Шостака , Фрэнка Сталя и других) и модель одноцепочечного отжига. Третья модель, модель отжига цепей, зависящая от синтеза, не включала контакты Холлидея. [3]
Первые экспериментальные доказательства структуры соединения Холлидея были получены в результате исследований электронной микроскопии в конце 1970-х годов, когда четырехветвевая структура была четко видна на изображениях ДНК плазмид и бактериофагов . Позже, в 1980-х годах, были идентифицированы ферменты, ответственные за инициацию образования соединений Холлидея и связывание с ними, хотя по состоянию на 2004 год идентификация резольваз соединений Холлидея у млекопитающих оставалась неуловимой (однако более подробную информацию см. в разделе «Разрешение соединений Холлидея» выше). последняя информация). сконструировал искусственные молекулы соединения Холлидея из синтетических олигонуклеотидов впервые В 1983 году Надриан Симан , что позволило более непосредственно изучить их физические свойства. Большая часть раннего анализа структуры соединения Холлидея была сделана на основе гель-электрофореза , FRET , а также гидроксильных радикалов и нуклеазного исследований следа. В 1990-х годах стали доступны методы кристаллографии и ЯМР нуклеиновых кислот , а также компьютерное молекулярное моделирование. инструменты. [1] [3] [28]
Первоначально генетики предполагали, что соединение примет параллельную, а не антипараллельную конформацию, поскольку это приведет к более тесному выравниванию гомологичных дуплексов друг к другу. [1] Химический анализ 1980-х годов показал, что соединение на самом деле предпочитает антипараллельную конформацию, и это открытие считалось спорным, и сам Робин Холлидей поначалу сомневался в этих результатах. [1] [3] Антипараллельная структура позже получила широкое признание благодаря данным рентгеновской кристаллографии молекул in vitro , хотя по состоянию на 2004 год значение структуры in vivo оставалось неясным, особенно структура соединений часто изменяется связанными с ней белками. [3]
Концептуальная основа нанотехнологии ДНК была впервые заложена Надрианом Зееманом в начале 1980-х годов. [29] В то время был известен ряд природных разветвленных структур ДНК, включая репликационную вилку ДНК и мобильное соединение Холлидея, но идея Симана заключалась в том, что неподвижные соединения нуклеиновых кислот могут быть созданы путем правильного проектирования последовательностей цепей для устранения симметрии в собранной молекуле. и что эти неподвижные соединения в принципе можно объединить в жесткие кристаллические решетки. Первая теоретическая статья, предлагающая эту схему, была опубликована в 1982 году, а первая экспериментальная демонстрация неподвижного соединения ДНК была опубликована в следующем году. [23] [30] двойного пересечения (DX) Симан разработал более жесткий мотив , подходящий для формирования двумерных решеток, продемонстрированный в 1998 году им и Эриком Уинфри . [22] В 2006 году Пол Ротемунд впервые продемонстрировал технику ДНК-оригами, позволяющую легко и надежно создавать свернутые структуры ДНК произвольной формы. Этот метод позволил создавать гораздо более крупные структуры, чем это было возможно ранее, и которые менее технически сложны в проектировании и синтезе. [31] Синтез трехмерной решетки был наконец опубликован Симаном в 2009 году, почти через тридцать лет после того, как он намеревался достичь этого. [32]
Ссылки
[ редактировать ]- ^ Jump up to: а б с д и ж г час я дж к л м н Лилли, Дэвид MJ (2000). «Структуры спиральных соединений нуклеиновых кислот». Ежеквартальные обзоры биофизики . 33 (2): 109–159. дои : 10.1017/S0033583500003590 . ПМИД 11131562 . S2CID 40501795 .
- ^ Блумфилд, Виктор А.; Кротерс, Дональд М.; Тиноко-младший, Игнасио (2000). Нуклеиновые кислоты: структура, свойства и функции . Саусалито, Калифорния: Университетские научные книги. п. 468 . ISBN 0935702490 .
- ^ Jump up to: а б с д и ж г час Лю Ю, Вест С (2004). «Счастливых праздников: 40-летие перекрестка Холлидей». Nature Reviews Молекулярно-клеточная биология . 5 (11): 937–44. дои : 10.1038/nrm1502 . ПМИД 15520813 . S2CID 24520723 .
- ^ Сун, П; Кляйн, Х. (октябрь 2006 г.). «Механизм гомологичной рекомбинации: медиаторы и хеликазы берут на себя регуляторные функции». Nature Reviews Молекулярно-клеточная биология . 7 (10): 739–750. дои : 10.1038/nrm2008 . ПМИД 16926856 . S2CID 30324005 .
- ^ Хартель, Дэниел Л.; Джонс, Элизабет В. (2009). «Глава 6: Молекулярная биология репликации и рекомбинации ДНК» . Генетика: анализ генетики и геномов . Берлингтон: Джонс и Бартлетт. ISBN 9780763758684 .
- ^ Хелледей, Т. (20 ноября 2018 г.). «Ремонт двухнитевого разрыва с помощью двойных соединений Холлидея (модель Шостака)» . Анимация . Массачусетский технологический институт.
- ^ Роша, EPC; Корнет, Э; Мишель, Б. (август 2005 г.). «Сравнительный и эволюционный анализ бактериальных гомологичных рекомбинационных систем» . ПЛОС Генетика . 1 (2): е15. дои : 10.1371/journal.pgen.0010015 . ПМК 1193525 . ПМИД 16132081 .

- ^ Флейшманн-младший, WR (1996). «Глава 43» . Медицинская микробиология (4-е изд.). Медицинский филиал Техасского университета в Галвестоне. ISBN 0-9631172-1-1 .
- ^ Западный СК (2003). «Молекулярные взгляды на рекомбинационные белки и их контроль». Nature Reviews Молекулярно-клеточная биология . 4 (6): 435–45. дои : 10.1038/nrm1127 . ПМИД 12778123 . S2CID 28474965 .
- ^ Ковальчиковский СК (2000). «Инициация генетической рекомбинации и рекомбинационно-зависимой репликации». Тенденции биохимических наук . 25 (4): 156–65. дои : 10.1016/S0968-0004(00)01569-3 . ПМИД 10754547 .
- ^ Бони, МФ; де Йонг, доктор медицины; ван Доорн, HR; Холмс, ЕС; Мартин, Даррен П. (3 мая 2010 г.). Мартин, Даррен П. (ред.). «Руководство по выявлению событий гомологичной рекомбинации в вирусе гриппа А» . ПЛОС ОДИН . 5 (5): е10434. Бибкод : 2010PLoSO...510434B . дои : 10.1371/journal.pone.0010434 . ПМЦ 2862710 . ПМИД 20454662 .
- ^ Jump up to: а б Захарьевич, К; Тан, С; Может; Хантер, Н. (апрель 2012 г.). «Определение путей разрешения совместных молекул в мейозе идентифицирует резольвазу, специфичную для кроссовера» . Клетка . 149 (2): 334–47. дои : 10.1016/j.cell.2012.03.023 . ПМЦ 3377385 . ПМИД 22500800 .
- ^ Jump up to: а б Ранджа, Л; Ананд, Р; Цейка, П. (2014). «Гетеродимер Saccharomyces cerevisiae Mlh1-Mlh3 представляет собой эндонуклеазу, которая преимущественно связывается с соединениями Холлидея» . Ж. Биол. Хим . 289 (9): 5674–86. дои : 10.1074/jbc.M113.533810 . ПМЦ 3937642 . ПМИД 24443562 .
- ^ Рогачева М.В., Манхарт С.М., Чен С., Гуарн А., Сёртиз Дж., Алани Э. (2014). «Mlh1-Mlh3, фактор мейотического кроссинговера и репарации несоответствия ДНК, представляет собой эндонуклеазу, стимулируемую Msh2-Msh3» . Ж. Биол. Хим . 289 (9): 5664–73. дои : 10.1074/jbc.M113.534644 . ПМЦ 3937641 . ПМИД 24403070 .
- ^ Jump up to: а б Зоннтаг Браун М., Лим Э., Чен С., Нишант К.Т., Алани Э. (2013). «Генетический анализ мутаций mlh3 выявляет взаимодействие между факторами, способствующими кроссинговеру, во время мейоза у пекарских дрожжей» . G3: Гены, геномы, генетика . 3 (1): 9–22. дои : 10.1534/g3.112.004622 . ПМК 3538346 . ПМИД 23316435 .
- ^ Jump up to: а б с Лукашевич А., Ховард-Тилль Р.А., Лойдл Дж. (2013). «Нуклеаза Mus81 и геликаза Sgs1 необходимы для мейотической рекомбинации у простейших, лишенных синаптонемного комплекса» . Нуклеиновые кислоты Рез . 41 (20): 9296–309. дои : 10.1093/нар/gkt703 . ПМЦ 3814389 . ПМИД 23935123 .
- ^ Jump up to: а б Почарт П., Уолтеринг Д., Холлингсворт Н.М. (1997). «Консервативные свойства функционально различных гомологов MutS у дрожжей» . Ж. Биол. Хим . 272 (48): 30345–9. дои : 10.1074/jbc.272.48.30345 . ПМИД 9374523 .
- ^ Винанд, Нью-Джерси, Panzer JA, Колоднер Р.Д. (1998). «Клонирование и характеристика гомологов человека и Caenorhabditis elegans гена Saccharomyces cerevisiae MSH5» . Геномика . 53 (1): 69–80. дои : 10.1006/geno.1998.5447 . ПМИД 9787078 .
- ^ Бокер Т., Барусевичус А., Сноуден Т., Расио Д., Герретт С., Роббинс Д., Шмидт С., Бурчак Дж., Кроче С.М., Коупленд Т., Коватич А.Дж., Фишел Р. (1999). «hMSH5: гомолог MutS человека, который образует новый гетеродимер с hMSH4 и экспрессируется во время сперматогенеза». Рак Рез . 59 (4): 816–22. ПМИД 10029069 .
- ^ Кришнапрасад Г.Н., Ананд М.Т., Лин Г., Теккедил М.М., Стейнмец Л.М., Нишант К.Т. (2015). «Изменения в частоте кроссовера нарушают уверенность кроссовера, не влияя на мейотическую сегрегацию хромосом у Saccharomyces cerevisiae» . Генетика . 199 (2): 399–412. дои : 10.1534/genetics.114.172320 . ПМК 4317650 . ПМИД 25467183 .
- ^ Мао, Чэндэ (декабрь 2004 г.). «Возникновение сложности: уроки ДНК» . ПЛОС Биология . 2 (12): 2036–2038. дои : 10.1371/journal.pbio.0020431 . ПМЦ 535573 . ПМИД 15597116 .
- ^ Jump up to: а б с д Симан, Надриан К. (июнь 2004 г.). «Нанотехнологии и двойная спираль». Научный американец . 290 (6): 64–75. Бибкод : 2004SciAm.290f..64S . doi : 10.1038/scientificamerican0604-64 . ПМИД 15195395 .
- ^ Jump up to: а б с д Симан, Надриан К. (2010). «Наноматериалы на основе ДНК» . Ежегодный обзор биохимии . 79 : 65–87. doi : 10.1146/annurev-biochem-060308-102244 . ПМЦ 3454582 . ПМИД 20222824 .
- ^ Пан, Кяо; Ким, До-Нюн; Чжан, Фэй; Адендорф, Мэтью Р.; Ян, Хао; Бат, Марк (3 декабря 2014 г.). «Безрешеточное предсказание трехмерной структуры запрограммированных сборок ДНК» . Природные коммуникации . 5 : 5578. Бибкод : 2014NatCo...5.5578P . дои : 10.1038/ncomms6578 . ПМК 4268701 . ПМИД 25470497 .
- ^ Сакка, Барбара; Нимейер, Кристоф М. (2012). «ДНК-оригами: искусство складывать ДНК» (PDF) . Angewandte Chemie, международное издание . 51 (1): 58–66. дои : 10.1002/anie.201105846 . ПМИД 22162047 . S2CID 8014597 . Проверено 25 февраля 2015 г.
- ^ Jump up to: а б Шталь FW (1 октября 1994 г.). «Перекресток Холлидей в честь своего тридцатилетия» ( PDF ) . Генетика . 138 (2): 241–246. дои : 10.1093/генетика/138.2.241 . ПМК 1206142 . ПМИД 7828807 .
- ^ Достижения генетики . Академическая пресса. 1971. ISBN 9780080568027 .
- ^ Хейс Ф.А., Уотсон Дж., Хо PS (2003). «Осторожно! Пересечение ДНК: кристаллические структуры соединений Холлидея» . J Биол Хим . 278 (50): 49663–49666. дои : 10.1074/jbc.R300033200 . ПМИД 14563836 .
- ^ Пелеско, Джон А. (2007). Самосборка: наука о вещах, которые соединяются воедино . Нью-Йорк: Чепмен и Холл/CRC. стр. 201, 242, 259. ISBN. 978-1-58488-687-7 .
- ^ Пиньейру, А.В.; Рука.; Ши, ВМ; Ян, Х. (декабрь 2011 г.). «Проблемы и возможности структурной ДНК-нанотехнологии» . Природные нанотехнологии . 6 (12): 763–772. Бибкод : 2011НатНа...6..763П . дои : 10.1038/nnano.2011.187 . ПМЦ 3334823 . ПМИД 22056726 .
- ^ Ротемунд, Пол В.К. (2006). «Оригами на каркасах ДНК: от обобщенных мультикроссоверов к полигональным сетям». В Чене, Чунхуэй; Йоноска, Наташа; Розенберг, Гжегож (ред.). Нанотехнологии: наука и вычисления . Серия естественных вычислений. Нью-Йорк: Спрингер. стр. 3–21. дои : 10.1007/3-540-30296-4_1 . ISBN 978-3-540-30295-7 .
- ^ Сервис, Роберт Ф. (3 июня 2011 г.). «ДНК-нанотехнологии растут». Наука . 332 (6034): 1140–1143. Бибкод : 2011Sci...332.1140S . дои : 10.1126/science.332.6034.1140 . ПМИД 21636754 .
Внешние ссылки
[ редактировать ]- Holliday + Junctions в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)
- Конформационное изменение соединения Холлидея
- Анализ ветвевой миграционной активности белков с использованием синтетических ДНК-субстратов (протокол)