Гомологичная рекомбинация

Гомологичная рекомбинация — это тип генетической рекомбинации , при котором происходит обмен генетической информацией между двумя сходными или идентичными молекулами двухцепочечных или одноцепочечных нуклеиновых кислот (обычно ДНК, как в клеточных организмах, но может быть также РНК, как и в вирусах ).
Гомологичная рекомбинация широко используется клетками для точного восстановления вредных разрывов ДНК, которые происходят на обеих цепях ДНК, известных как двухцепочечные разрывы (DSB), в процессе, называемом гомологичной рекомбинационной репарацией (HRR). [1]
Гомологичная рекомбинация также приводит к образованию новых комбинаций последовательностей ДНК во время мейоза — процесса, посредством которого эукариоты создают гаметы , подобно сперматозоидам и яйцеклеткам у животных. Эти новые комбинации ДНК представляют собой генетические вариации потомства, что, в свою очередь, позволяет популяциям адаптироваться в ходе эволюции . [2]
Гомологичная рекомбинация также используется при горизонтальном переносе генов для обмена генетическим материалом между различными штаммами и видами бактерий и вирусов. Горизонтальный перенос генов является основным механизмом распространения устойчивости бактерий к антибиотикам.
Хотя гомологичная рекомбинация широко варьируется среди разных организмов и типов клеток, большинство форм двухцепочечной ДНК ( дцДНК ) включают одни и те же основные этапы. После того, как происходит двухцепочечный разрыв, участки ДНК вокруг 5'-конца разрыва отсекаются в процессе, называемом резекцией . На следующем этапе инвазии цепи выступающий 3'-конец разорванной молекулы ДНК затем «вторгается» в аналогичную или идентичную молекулу ДНК, которая не разорвана. После инвазии цепи дальнейшая последовательность событий может следовать любому из двух основных путей, обсуждаемых ниже (см. Модели ); путь DSBR (восстановление двухцепочечного разрыва) или путь SDSA (синтез-зависимый отжиг цепи). Гомологичная рекомбинация, происходящая во время репарации ДНК, имеет тенденцию приводить к образованию продуктов, не связанных с кроссинговером, фактически восстанавливая поврежденную молекулу ДНК в том виде, в котором она существовала до двухцепочечного разрыва.
Гомологичная рекомбинация сохраняется во всех трех областях жизни, а также в ДНК- и РНК- вирусах , что позволяет предположить, что это почти универсальный биологический механизм. Открытие генов гомологичной рекомбинации у протистов — разнообразной группы эукариотических микроорганизмов — было интерпретировано как свидетельство того, что гомологичная рекомбинация возникла на ранних стадиях эволюции эукариот. Поскольку их дисфункция тесно связана с повышенной восприимчивостью к нескольким типам рака , белки, способствующие гомологичной рекомбинации, являются предметом активных исследований. Гомологичная рекомбинация также используется в генном нацеливании — методе внесения генетических изменений в организмы-мишени. За разработку этой техники Марио Капечки , Мартин Эванс и Оливер Смитис были удостоены Нобелевской премии по физиологии и медицине 2007 года ; Капечки [3] и кузницы [4] независимо обнаружили применение к эмбриональным стволовым клеткам мыши, однако высококонсервативные механизмы, лежащие в основе модели репарации DSB, включая равномерную гомологичную интеграцию трансформированной ДНК (генная терапия), были впервые показаны в экспериментах с плазмидами Орром-Уивером, Шостаком и Ротштейном. [5] [6] [7] Исследование плазмид-индуцированного DSB с использованием γ-облучения [8] в 1970-1980-х годах, привело к более поздним экспериментам с использованием эндонуклеаз (например, I-SceI) для разрезания хромосом для генной инженерии клеток млекопитающих, где негомологичная рекомбинация встречается чаще, чем у дрожжей. [9]
и открытия История

В начале 1900-х годов Уильям Бейтсон и Реджинальд Паннетт обнаружили исключение из одного из принципов наследования, первоначально описанного Грегором Менделем в 1860-х годах. В отличие от идеи Менделя о том, что признаки передаются от родителя к ребенку независимо друг от друга (например, что цвет шерсти кошки и длина ее хвоста наследуются независимо друг от друга), Бейтсон и Паннетт показали, что определенные гены, связанные с физическими признаками, могут наследоваться вместе. или генетически связаны . [10] [11] В 1911 году, заметив, что сцепленные признаки могут иногда наследоваться по отдельности, Томас Хант Морган « кроссинговеры ». предположил, что между сцепленными генами могут происходить [12] где один из связанных генов физически переходит на другую хромосому . Два десятилетия спустя Барбара МакКлинток и Гарриет Крейтон продемонстрировали, что хромосомный кроссинговер происходит во время мейоза . [13] [14] процесс деления клеток, в ходе которого сперматозоиды и яйцеклетки образуются . В том же году, когда было сделано открытие МакКлинтока, Курт Стерн показал, что кроссинговер, позже названный «рекомбинацией», может также происходить в соматических клетках, таких как лейкоциты и клетки кожи , которые делятся посредством митоза . [13] [15]
В 1947 году микробиолог Джошуа Ледерберг показал, что бактерии, которые, как предполагалось, размножаются только бесполым путем путем бинарного деления , способны к генетической рекомбинации, которая больше похожа на половое размножение. Эта работа сделала E. coli модельным организмом в генетике. [16] и помог Ледербергу получить Нобелевскую премию по физиологии и медицине 1958 года . [17] Основываясь на исследованиях грибов , в 1964 году Робин Холлидей предложил модель рекомбинации при мейозе, которая представила ключевые детали того, как может работать этот процесс, включая обмен материалом между хромосомами через соединения Холлидея . [18] В 1983 году Джек Шостак и его коллеги представили модель, теперь известную как путь DSBR , которая учитывала наблюдения, не объясненные моделью Холлидея. [18] [7] В течение следующего десятилетия эксперименты на дрозофиле , почкующихся дрожжах и клетках млекопитающих привели к появлению других моделей гомологичной рекомбинации, называемых путями SDSA , которые не всегда полагаются на соединения Холлидея. [18]
Большая часть более поздних работ по идентификации белков, участвующих в этом процессе, и определению их механизмов была выполнена рядом людей, включая Джеймса Хабера , Патрика Санга , Стивена Ковальчиковски и других.
У эукариотов [ править ]
Гомологичная рекомбинация (HR) необходима для деления клеток у эукариот, таких как растения, животные, грибы и простейшие. Гомологичная рекомбинация восстанавливает двухцепочечные разрывы ДНК, вызванные ионизирующим излучением или химическими веществами, повреждающими ДНК. [19] Если не устранить эти двухцепочечные разрывы, они могут вызвать крупномасштабную перестройку хромосом в соматических клетках . [20] что, в свою очередь, может привести к раку. [21]
Помимо восстановления ДНК, гомологичная рекомбинация также помогает создавать генетическое разнообразие , когда клетки делятся в мейозе и становятся специализированными гаметными клетками — сперматозоидами или яйцеклетками у животных, пыльцой или семяпочками у растений и спорами у грибов . Это происходит за счет облегчения хромосомного кроссинговера обмениваются областями сходной, но не идентичной ДНК , при котором гомологичные хромосомы . [22] [23] Это создает новые, возможно, полезные комбинации генов, которые могут дать потомству эволюционное преимущество. [24] Хромосомный кроссинговер часто начинается, когда белок под названием Spo11 совершает целевой двухцепочечный разрыв ДНК. [25] Эти участки расположены на хромосомах неслучайно; обычно в межгенных промоторных областях и предпочтительно в богатых GC доменах, [26] Эти сайты двухцепочечного разрыва часто возникают в «горячих точках» рекомбинации — участках хромосом, длина которых составляет около 1000–2000 пар оснований и имеют высокие скорости рекомбинации. Отсутствие «горячей точки» рекомбинации между двумя генами в одной и той же хромосоме часто означает, что эти гены будут унаследованы будущими поколениями в равной пропорции. Это представляет собой более сильную связь между двумя генами, чем можно было бы ожидать от генов, которые независимо сортируются во время мейоза. [27]
Сроки митотического цикла клеточного
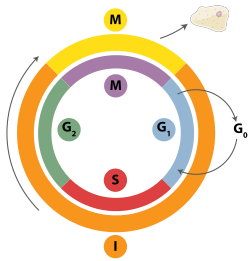
Двухцепочечные разрывы можно восстановить посредством гомологичной рекомбинации, тета-опосредованного соединения концов полимеразы (TMEJ) или негомологичного соединения концов (NHEJ). [28] NHEJ — это механизм репарации ДНК, который, в отличие от гомологичной рекомбинации, не требует длинной гомологичной последовательности для управления репарацией. Используется ли гомологичная рекомбинация или NHEJ для восстановления двухцепочечных разрывов, во многом определяется фазой клеточного цикла . Гомологичная рекомбинация восстанавливает ДНК до того, как клетка вступает в митоз (М-фаза). вскоре после репликации ДНК , в S и G2 фазах Это происходит во время и клеточного цикла, когда сестринские хроматиды более доступны. [29] По сравнению с гомологичными хромосомами, которые похожи на другую хромосому, но часто имеют разные аллели , сестринские хроматиды являются идеальной матрицей для гомологичной рекомбинации, поскольку они представляют собой идентичную копию данной хромосомы. Когда гомологичная матрица недоступна или когда матрица недоступна из-за дефекта гомологичной рекомбинации, разрыв восстанавливается посредством TMEJ в S и G 2 фазах клеточного цикла. В отличие от гомологичной рекомбинации и TMEJ, NHEJ преобладает в клеточного G1 фазе цикла, когда клетка растет, но еще не готова к делению. Это происходит реже после фазы G1 , но сохраняет по крайней мере некоторую активность на протяжении всего клеточного цикла. Механизмы, которые регулируют гомологичную рекомбинацию и NHEJ на протяжении клеточного цикла, широко варьируются у разных видов. [30]
Циклин-зависимые киназы (CDK), которые модифицируют активность других белков путем добавления к ним фосфатных групп (то есть фосфорилирования ), являются важными регуляторами гомологичной рекомбинации у эукариот. [30] Когда у почкующихся дрожжей начинается репликация ДНК, циклинзависимая киназа Cdc28 начинает гомологичную рекомбинацию путем фосфорилирования белка Sae2 . [31] После активации добавлением фосфата Sae2 вызывает чистый разрез вблизи двухцепочечного разрыва ДНК. Неясно, является ли эндонуклеаза, ответственная за этот разрез, самим Sae2 или другим белком, Mre11 . [32] Это позволяет белковому комплексу , включающему Mre11, известному как комплекс MRX , связываться с ДНК и запускать серию управляемых белками реакций, которые обменивают материал между двумя молекулами ДНК. [33]
Роль хроматина [ править ]
Упаковка эукариотической ДНК в хроматин представляет собой барьер для всех процессов, основанных на ДНК, которые требуют привлечения ферментов к местам их действия. Чтобы обеспечить репарацию ДНК гомологичной рекомбинации (HR), хроматин должен быть ремоделирован. У эукариот АТФ-зависимые ремоделирования хроматина комплексы и ферменты, модифицирующие гистоны, являются двумя преобладающими факторами, используемыми для осуществления этого процесса ремоделирования. [34]
Релаксация хроматина быстро происходит в месте повреждения ДНК. [35] На одном из самых ранних этапов активируемая стрессом протеинкиназа, N-концевая киназа c-Jun (JNK) , фосфорилирует SIRT6 по серину 10 в ответ на двухцепочечные разрывы или другие повреждения ДНК. [36] Эта посттрансляционная модификация облегчает мобилизацию SIRT6 к сайтам повреждения ДНК и необходима для эффективного рекрутирования поли(ADP-рибозо)полимеразы 1 (PARP1) к сайтам разрыва ДНК и для эффективного восстановления DSB. [36] Белок PARP1 начинает появляться в местах повреждения ДНК менее чем за секунду, с полумаксимальным накоплением в течение 1,6 секунды после возникновения повреждения. [37] Затем ремодератор хроматина Alc1 быстро присоединяется к продукту действия PARP1 — рибозной цепи поли-АДФ, и Alc1 завершает прибытие к повреждению ДНК в течение 10 секунд после возникновения повреждения. [35] Около половины максимального расслабления хроматина, предположительно за счет действия Alc1, происходит к 10 с. [35] Затем это позволяет задействовать фермент репарации ДНК MRE11 , чтобы инициировать репарацию ДНК в течение 13 секунд. [37]
γH2AX, фосфорилированная форма H2AX, также участвует на ранних этапах, приводящих к деконденсации хроматина после двухцепочечных разрывов ДНК. Вариант гистонов H2AX составляет около 10% гистонов H2A в хроматине человека. [38] γH2AX (H2AX, фосфорилированный по серину 139) можно обнаружить уже через 20 секунд после облучения клеток (с образованием двухцепочечного разрыва ДНК), а накопление γH2AX на половину максимального уровня происходит за одну минуту. [38] Протяженность хроматина с фосфорилированным γH2AX составляет около двух миллионов пар оснований в месте двухцепочечного разрыва ДНК. [38] γH2AX сам по себе не вызывает деконденсации хроматина, но в течение 30 секунд после облучения RNF8 в ассоциации с γH2AX. можно обнаружить белок [39] RNF8 опосредует обширную деконденсацию хроматина посредством его последующего взаимодействия с CHD4 . [40] компонент комплекса ремоделирования нуклеосом и деацетилазы NuRD .
После релаксации после повреждения ДНК с последующей репарацией ДНК хроматин восстанавливается до состояния уплотнения, близкого к уровню до повреждения, примерно через 20 мин. [35]
Гомологичная рекомбинация во время мейоза [ править ]
У позвоночных места, в которых происходит рекомбинация, определяются местами связывания PRDM9 , белка, который распознает специфический мотив последовательности с помощью своего массива цинковых пальцев. [41] В этих сайтах другой белок, SPO11, катализирует двухцепочечные разрывы, инициирующие рекомбинацию (DSB), часть которых восстанавливается путем рекомбинации с гомологичной хромосомой. PRDM9 откладывает метки метилирования гистонов H3K4me3 и H3K36me3 в сайтах, с которыми он связывается, и эта активность метилтрансферазы важна для его роли в позиционировании DSB. После их формирования сайты DSB подвергаются резекции, в результате чего образуется одноцепочечная ДНК (оцДНК), которая декорируется DMC1. От средней зиготены до ранней пахитены, как часть процесса рекомбинационной репарации, DMC1 диссоциирует от оцДНК и количество уменьшается до тех пор, пока все разрывы (кроме тех, что на хромосомах XY) не будут восстановлены в поздней пахитене. В этом процессе участвуют несколько других белков, в том числе ZCWPW1, [42] первый белок, непосредственно позиционируемый по двойным гистоновым меткам PRDM9. ZCWPW1 важен для гомологичной репарации DSB, а не для позиционирования.
Модели [ править ]
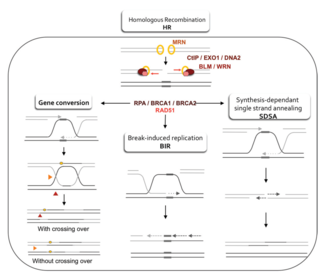
Двумя основными моделями того, как гомологичная рекомбинация восстанавливает двухцепочечные разрывы в ДНК, являются путь восстановления двухцепочечных разрывов (DSBR) (иногда называемый моделью двойного соединения Холлидея ) и путь синтез-зависимого отжига цепей (SDSA). [43] Оба пути схожи на первых этапах. После того, как происходит двухцепочечный разрыв, комплекс MRX ( комплекс MRN у человека) связывается с ДНК по обе стороны от разрыва. Затем происходит резекция, при которой ДНК вокруг 5'-конца разрыва обрезается. Это происходит в два отдельных этапа: сначала комплекс MRX рекрутирует белок Sae2, и эти два белка обрезают 5'-концы по обе стороны от разрыва, создавая короткие 3'-выступы одноцепочечной ДНК; на втором этапе резекцию 5'→3' продолжают хеликазы Sgs1 и нуклеазы Exo1 и Dna2 . Будучи хеликазой , Sgs1 «расстегивает» двухцепочечную ДНК, тогда как нуклеазная активность Exo1 и Dna2 позволяет им разрезать одноцепочечную ДНК, продуцируемую Sgs1. [31]
Белок RPA , обладающий высоким сродством к одноцепочечной ДНК, затем связывает 3'-концы. [44] С помощью нескольких других белков, которые опосредуют этот процесс, белок Rad51 (и Dmc1 в мейозе) затем образует нить из нуклеиновой кислоты и белка на единственной цепи ДНК, покрытой RPA. Затем эта нуклеопротеиновая нить начинает поиск последовательностей ДНК, аналогичных последовательностям 3'-конца. После обнаружения такой последовательности одноцепочечная нуклеопротеиновая нить перемещается (вторгается) в аналогичный или идентичный дуплекс ДНК реципиента в процессе, называемом инвазией цепи . В клетках, которые делятся посредством митоза, дуплекс ДНК-реципиента обычно представляет собой сестринскую хроматиду, которая идентична поврежденной молекуле ДНК и обеспечивает матрицу для восстановления. Однако при мейозе ДНК-реципиент обычно происходит из сходной, но не обязательно идентичной гомологичной хромосомы. [43] Петля смещения ( D-петля ) образуется во время инвазии цепи между вторгающейся 3'-концевой цепью и гомологичной хромосомой. После инвазии цепи ДНК-полимераза удлиняет конец вторгшейся 3'-цепи, синтезируя новую ДНК. Это превращает D-петлю в крестообразную структуру, известную как соединение Холлидея . После этого на вторгающейся цепи происходит больший синтез ДНК (т.е. на одном из исходных 3'-концевиков), эффективно восстанавливая цепь гомологичной хромосомы, которая была смещена во время инвазии цепи. [43]
Путь DSBR [ править ]

После стадий резекции, инвазии цепи и синтеза ДНК пути DSBR и SDSA становятся разными. [43] Путь DSBR уникален тем, что второй 3'-выступ (который не участвовал в инвазии цепи) также образует соединение Холлидея с гомологичной хромосомой. Двойные соединения Холлидея затем превращаются в продукты рекомбинации с помощью разрывающих эндонуклеаз , типа эндонуклеазы рестрикции , которая разрезает только одну цепь ДНК. Путь DSBR обычно приводит к кроссоверу, хотя иногда он может приводить к образованию продуктов без кроссовера; способность сломанной молекулы ДНК собирать последовательности из отдельных донорских локусов была показана на митотических почкующихся дрожжах с использованием плазмид или эндонуклеазной индукции хромосомных событий. [45] [46] Из-за этой тенденции к хромосомному кроссоверу путь DSBR является вероятной моделью того, как происходит гомологичная рекомбинация кроссовера во время мейоза. [22]
Приведет ли рекомбинация по пути DSBR к хромосомному кроссоверу, определяется тем, как двойное соединение Холлидея разрезается или «разрешается». Хромосомный кроссинговер произойдет, если одно соединение Холлидея перерезать на пересекающейся цепи, а другое соединение Холлидея перерезать на непересекающейся цепи (на рисунке 5 вдоль горизонтальных фиолетовых стрелок на одном соединении Холлидея и вдоль вертикальных оранжевых стрелок на другом). ). Альтернативно, если два соединения Холлидея разрезать на пересекающихся цепях (вдоль горизонтальных фиолетовых стрелок на обоих соединениях Холлидея на рисунке 5), то будут получены хромосомы без кроссинговера. [47]
Путь SDSA [ править ]
Гомологичная рекомбинация по пути SDSA происходит в клетках, которые делятся посредством митоза и мейоза и приводят к образованию продуктов без кроссинговера. В этой модели вторгающаяся 3'-цепь удлиняется вдоль дуплекса ДНК реципиента с помощью ДНК-полимеразы и высвобождается, когда соединение Холлидея между молекулами ДНК донора и реципиента скользит в процессе, называемом миграцией ветвей . Вновь синтезированный 3'-конец вторгшейся цепи затем способен отжигаться с другим 3'-концом поврежденной хромосомы посредством комплементарного спаривания оснований. После отжига нитей иногда может оставаться небольшой лоскут ДНК. Любые такие лоскуты удаляются, и путь SDSA заканчивается повторным запечатыванием, также известным как лигирование , любых оставшихся одноцепочечных разрывов. [48]
Во время митоза основным путем гомологичной рекомбинации для восстановления двухцепочечных разрывов ДНК, по-видимому, является путь SDSA (а не путь DSBR). [49] Путь SDSA приводит к образованию некроссинговерных рекомбинантов (рис. 5). Во время мейоза также часто встречаются некроссинговерные рекомбинанты, которые, по-видимому, также возникают в основном по пути SDSA. [49] [50] События некроссинговерной рекомбинации, происходящие во время мейоза, вероятно, отражают случаи восстановления двухцепочечных повреждений ДНК или других типов повреждений ДНК.
Путь SSA [ править ]

Путь гомологичной рекомбинации одноцепочечного отжига (SSA) восстанавливает двухцепочечные разрывы между двумя повторяющимися последовательностями . Путь SSA уникален тем, что он не требует отдельной подобной или идентичной молекулы ДНК, как пути гомологичной рекомбинации DSBR или SDSA. Вместо этого путь SSA требует только одного дуплекса ДНК и использует повторяющиеся последовательности как идентичные последовательности, необходимые для репарации гомологичной рекомбинации. Концепция этого пути относительно проста: после того, как две цепи одного и того же дуплекса ДНК разрезаются вокруг места двухцепочечного разрыва, два образующихся 3'-выступа затем выравниваются и отжигаются друг с другом, восстанавливая ДНК как непрерывный дуплекс. . [48] [51]
Поскольку ДНК вокруг двухцепочечного разрыва сокращается, образующиеся одноцепочечные 3'-концы покрываются белком RPA , который предотвращает прилипание 3'-концов к самим себе. [52] Белок под названием Rad52 затем связывает каждую из повторяющихся последовательностей по обе стороны от разрыва и выравнивает их, позволяя двум комплементарным повторяющимся последовательностям соединиться. [52] После завершения отжига оставшиеся негомологичные лоскуты 3'-концов отсекаются набором нуклеаз, известных как Rad1/Rad10 , которые доставляются к лоскутам белками Saw1 и Slx4 . [52] [53] Синтез новой ДНК заполняет любые пробелы, а лигирование восстанавливает дуплекс ДНК в виде двух непрерывных цепей. [54] Последовательность ДНК между повторами всегда теряется, как и один из двух повторов. Путь SSA считается мутагенным , поскольку он приводит к таким делециям генетического материала. [48]
Путь BIR [ править ]
Во время репликации ДНК иногда можно встретить двухцепочечные разрывы в репликационных вилках, поскольку ДНК-хеликаза расстегивает цепь матрицы. Эти дефекты устраняются путем индуцированной разрывом репликации гомологичной рекомбинации, (BIR). Точные молекулярные механизмы пути BIR остаются неясными. Три предложенных механизма включают инвазию цепи в качестве начального этапа, но они различаются тем, как моделируют миграцию D-петли и более поздние фазы рекомбинации. [55]
Путь BIR также может помочь поддерживать длину теломер (участков ДНК на концах эукариотических хромосом) в отсутствие теломеразы (или во взаимодействии с ней) . Без рабочих копий фермента теломеразы теломеры обычно укорачиваются с каждым циклом митоза, что в конечном итоге блокирует деление клеток и приводит к старению . В почкующихся дрожжевых клетках, где теломераза была инактивирована в результате мутаций, было обнаружено два типа «выживших» клеток, которые избегают старения дольше, чем ожидалось, за счет удлинения своих теломер посредством путей BIR. [55]
Поддержание длины теломер имеет решающее значение для иммортализации клеток , ключевой особенности рака. Большинство видов рака поддерживают теломеры за счет активации теломеразы. Однако при некоторых типах рака человека BIR-подобный путь помогает поддерживать некоторые опухоли, действуя в качестве альтернативного механизма поддержания теломер. [56] Этот факт побудил ученых исследовать, могут ли такие механизмы поддержания теломер, основанные на рекомбинации, помешать лечению противораковыми препаратами, такими как ингибиторы теломеразы . [57]
У бактерий [ править ]

Гомологичная рекомбинация является основным процессом репарации ДНК у бактерий. Это также важно для создания генетического разнообразия в бактериальных популяциях, хотя этот процесс существенно отличается от мейотической рекомбинации, которая восстанавливает повреждения ДНК и приводит к разнообразию в эукариот геномах . Гомологичная рекомбинация наиболее изучена и лучше всего понята для Escherichia coli . [59] Двухцепочечные разрывы ДНК у бактерий восстанавливаются с помощью RecBCD пути гомологичной рекомбинации . Считается, что разрывы, возникающие только на одной из двух цепей ДНК, известные как одноцепочечные разрывы, восстанавливаются с помощью пути RecF . [60] Оба пути RecBCD и RecF включают в себя серию реакций, известных как миграция ветвей , при которой отдельные цепи ДНК обмениваются между двумя перекрещивающимися молекулами дуплексной ДНК, и резолюция , при которой эти две перекрещенные молекулы ДНК разрезаются на части и восстанавливаются до нормального состояния. двухцепочечное состояние.
Путь RecBCD [ править ]


Путь RecBCD является основным путем рекомбинации, используемым у многих бактерий для восстановления двухцепочечных разрывов ДНК, а белки обнаружены у широкого спектра бактерий. [63] [64] [65] Эти двухцепочечные разрывы могут быть вызваны ультрафиолетовым излучением и другим излучением , а также химическими мутагенами . Двухцепочечные разрывы также могут возникать в результате репликации ДНК через одноцепочечный разрыв или разрыв. Такая ситуация вызывает так называемую коллапс репликационной вилки и фиксируется несколькими путями гомологичной рекомбинации, включая путь RecBCD. [66]
В этом пути трехсубъединичный ферментный комплекс , называемый RecBCD, инициирует рекомбинацию, связываясь с тупым или почти тупым концом разрыва двухцепочечной ДНК. RecB и RecD После того, как RecBCD связывается с концом ДНК, субъединицы начинают распаковывать дуплекс ДНК посредством активности геликазы . Субъединица RecB также имеет нуклеазный домен, который разрезает одну цепь ДНК, возникающую в процессе распаковки. Это разархивирование продолжается до тех пор, пока RecBCD не встретит определенную нуклеотидную последовательность (5'-GCTGGTGG-3'), известную как сайт Chi . [65]
При встрече с сайтом Chi активность фермента RecBCD резко меняется. [64] [61] [67] Раскручивание ДНК приостанавливается на несколько секунд, а затем возобновляется примерно на половине начальной скорости. Вероятно, это связано с тем, что более медленная хеликаза RecB раскручивает ДНК после Chi, а не более быстрая хеликаза RecD, которая раскручивает ДНК до Chi. [68] [69] Распознавание сайта Chi также изменяет фермент RecBCD, так что он разрезает цепь ДНК с Chi и начинает загрузку нескольких белков RecA в одноцепочечную ДНК с вновь образовавшимся 3'-концом. нить , покрытая RecA Полученная в результате нуклеопротеиновая , затем ищет аналогичные последовательности ДНК на гомологичной хромосоме. Процесс поиска вызывает растяжение дуплекса ДНК, что усиливает распознавание гомологии (механизм, называемый конформационной корректурой). [70] [71] [72] ). При обнаружении такой последовательности одноцепочечная нуклеопротеиновая нить перемещается в гомологичный дуплекс ДНК реципиента в процессе, называемом инвазией цепи . [73] Вторжение 3'-выступа приводит к смещению одной из нитей дуплекса ДНК-реципиента с образованием D-петли. Если D-петлю разрезать, то еще одна замена нитей местами образует крестообразную структуру, называемую соединением Холлидея . [65] Разрешение соединения Холлидея с помощью некоторой комбинации RuvABC или RecG может привести к образованию двух рекомбинантных молекул ДНК с взаимными генетическими типами, если две взаимодействующие молекулы ДНК различаются генетически. Альтернативно, вторгающийся 3'-конец рядом с Chi может запускать синтез ДНК и образовывать репликационную вилку. Этот тип разрешения дает только один тип рекомбинанта (невзаимный).
Путь RecF [ править ]
Бактерии, по-видимому, используют путь гомологичной рекомбинации RecF для восстановления одноцепочечных разрывов в ДНК. Когда путь RecBCD инактивируется мутациями, а дополнительные мутации инактивируют нуклеазы SbcCD и ExoI, путь RecF также может восстанавливать двухцепочечные разрывы ДНК. [74] В пути RecF хеликаза RecQ раскручивает ДНК, а нуклеаза RecJ разрушает цепь с 5'-концом, оставляя цепь с 3'-концом неповрежденной. Белок RecA связывается с этой цепью и либо поддерживается белками RecF, RecO и RecR, либо стабилизируется ими. Затем нуклеопротеиновая нить RecA ищет гомологичную ДНК и обменивается местами с идентичной или почти идентичной цепью гомологичной ДНК.
Хотя белки и специфические механизмы, участвующие в их начальных фазах, различаются, эти два пути схожи в том, что для инвазии цепи им обоим требуется одноцепочечная ДНК с 3'-концом и белок RecA. Пути также схожи по фазам миграции ветвей , при которой соединение Холлидея скользит в одном направлении, и по фазам разрешения , при котором соединения Холлидея расщепляются ферментами. [75] [76] Альтернативный, нереципрокный тип разрешения также может происходить любым путем.
Миграция филиала [ править ]
Сразу после инвазии цепи соединение Холлидея перемещается вдоль связанной ДНК во время процесса миграции ветвей. Именно при этом движении соединения Холлидея происходит обмен парами оснований между двумя гомологичными дуплексами ДНК. Чтобы катализировать миграцию ветвей, белок RuvA сначала распознает соединение Холлидея, связывается с ним и рекрутирует белок RuvB для формирования комплекса RuvAB. Два набора белков RuvB, каждый из которых образует кольцевую АТФазу , загружаются на противоположные стороны соединения Холлидея, где они действуют как двойные насосы, обеспечивающие силу миграции ветвей. Между этими двумя кольцами RuvB два набора белков RuvA собираются в центре соединения Холлидея, так что ДНК в месте соединения оказывается зажатой между каждым набором RuvA. Нити обоих дуплексов ДНК — дуплексов «донора» и «реципиента» — раскручиваются на поверхности RuvA, поскольку они направляются белком от одного дуплекса к другому. [77] [78]
Разрешение [ править ]
В фазе разрешения рекомбинации любые соединения Холлидея, образовавшиеся в результате процесса инвазии цепи, разрезаются, тем самым восстанавливая две отдельные молекулы ДНК. Это расщепление осуществляется комплексом RuvAB, взаимодействующим с RuvC, которые вместе образуют комплекс RuvABC . RuvC представляет собой эндонуклеазу , разрезающую вырожденную последовательность 5'-(A/T)TT(G/C)-3'. Последовательность часто встречается в ДНК, примерно один раз на каждые 64 нуклеотида. [78] Перед разрезанием RuvC, вероятно, получает доступ к соединению Холлидея, вытесняя один из двух тетрамеров RuvA, покрывающих там ДНК. [77] Рекомбинация приводит к образованию продуктов «сращивания» или «заплаток», в зависимости от того, как RuvC расщепляет соединение Холлидея. [78] Продукты сплайсинга — это продукты кроссинговера, при которых происходит перестановка генетического материала вокруг места рекомбинации. С другой стороны, патч-продукты - это продукты без кроссовера, в которых такой перегруппировки нет, а в продукте рекомбинации есть только «участок» гибридной ДНК. [79]
переносу генетическому Содействие
Гомологичная рекомбинация — важный метод интеграции донорской ДНК в геном организма-реципиента при горизонтальном переносе генов — процессе, посредством которого организм включает чужеродную ДНК из другого организма, не являясь потомком этого организма. Гомологичная рекомбинация требует, чтобы входящая ДНК была очень похожа на геном реципиента, поэтому горизонтальный перенос генов обычно ограничивается сходными бактериями. [80] Исследования на нескольких видах бактерий установили, что существует лог-линейное снижение частоты рекомбинации с увеличением разницы в последовательностях между ДНК хозяина и реципиента. [81] [82] [83]
При бактериальной конъюгации , когда ДНК передается между бактериями посредством прямого контакта между клетками, гомологичная рекомбинация помогает интегрировать чужеродную ДНК в геном хозяина по пути RecBCD. Фермент RecBCD способствует рекомбинации после того, как ДНК преобразуется из одноцепочечной ДНК (в той форме, в которой она первоначально поступает в бактерию) в двухцепочечную ДНК во время репликации. Путь RecBCD также важен для заключительной фазы трансдукции , типа горизонтального переноса генов, при котором ДНК переносится от одной бактерии к другой с помощью вируса . Чужеродная бактериальная ДНК иногда ошибочно включается в капсидную головку вирусных частиц бактериофага, поскольку ДНК упаковывается в новые бактериофаги во время репликации вируса. Когда эти новые бактериофаги заражают другие бактерии, ДНК предыдущей бактерии-хозяина вводится в новую бактерию-хозяина в виде двухцепочечной ДНК. Фермент RecBCD затем включает эту двухцепочечную ДНК в геном нового бактериального хозяина. [65]
Бактериальная трансформация [ править ]
Естественная бактериальная трансформация включает перенос ДНК от бактерии-донора к бактерии-реципиенту, при этом донор и реципиент обычно принадлежат к одному и тому же виду . Трансформация, в отличие от бактериальной конъюгации и трансдукции, зависит от многочисленных продуктов бактериальных генов, которые специфически взаимодействуют, осуществляя этот процесс. [84] Таким образом, трансформация явно является бактериальной адаптацией к переносу ДНК. Чтобы бактерия могла связать, захватить и интегрировать донорскую ДНК в свою резидентную хромосому путем гомологичной рекомбинации, она должна сначала войти в особое физиологическое состояние, называемое компетентностью . Семейство генов RecA / Rad51 / DMC1 играет центральную роль в гомологичной рекомбинации во время бактериальной трансформации, а также во время эукариотического мейоза и митоза. Например, белок RecA необходим для трансформации Bacillus subtilis и Streptococcus pneumoniae . [85] и экспрессия гена RecA индуцируется во время развития способности к трансформации у этих организмов.
В рамках процесса трансформации белок RecA взаимодействует с входящей одноцепочечной ДНК (оцДНК) с образованием нуклеофиламентов RecA/оцДНК, которые сканируют резидентную хромосому на предмет областей гомологии и доставляют вступающую оцДНК в соответствующую область, где происходит обмен цепей и гомологичность. происходит рекомбинация. [86] Таким образом, процесс гомологичной рекомбинации при бактериальной трансформации имеет фундаментальное сходство с гомологичной рекомбинацией во время мейоза .
В вирусах [ править ]
Гомологичная рекомбинация происходит у нескольких групп вирусов. В ДНК-вирусах, таких как вирус герпеса , рекомбинация происходит по механизму разрыва и воссоединения, как у бактерий и эукариот. [87] Существуют также доказательства рекомбинации у некоторых РНК-вирусов , особенно у вирусов с оцРНК с положительным смыслом, таких как ретровирусы , пикорнавирусы и коронавирусы . [88] Существуют разногласия по поводу того, происходит ли гомологичная рекомбинация у вирусов оцРНК с отрицательным смыслом, таких как грипп . [89]
У РНК-вирусов гомологичная рекомбинация может быть точной или неточной. В точном типе рекомбинации РНК-РНК нет разницы между двумя родительскими последовательностями РНК и полученной кроссоверной областью РНК. Из-за этого часто бывает трудно определить местонахождение событий кроссовера между двумя рекомбинирующими последовательностями РНК. При неточной гомологичной рекомбинации РНК область кроссовера имеет некоторые различия с последовательностями родительской РНК, вызванные добавлением, делецией или другой модификацией нуклеотидов. Уровень точности кроссовера контролируется контекстом последовательности двух рекомбинирующих цепей РНК: последовательности, богатые аденином и урацилом, снижают точность кроссовера. [88] [90]
Гомологичная рекомбинация важна для облегчения вирусной эволюции . [88] [91] Например, если геномы двух вирусов с разными невыгодными мутациями подвергнутся рекомбинации, то они смогут регенерировать полностью функциональный геном. Альтернативно, если два похожих вируса заразили одну и ту же клетку-хозяина, гомологичная рекомбинация может позволить этим двум вирусам поменяться генами и тем самым развить более мощные вариации самих себя. [91]
Гомологичная рекомбинация - это предполагаемый механизм, посредством которого ДНК вируса герпеса человека-6 интегрируется в теломеры человека. [92]
Когда два или более вируса, каждый из которых содержит летальные геномные повреждения, заражают одну и ту же клетку-хозяина, геномы вируса часто могут спариваться друг с другом и подвергаться гомологичной рекомбинационной репарации с образованием жизнеспособного потомства. Этот процесс, известный как реактивация множественности, был изучен на нескольких бактериофагах , включая фаг Т4 . [93] Ферменты, используемые в рекомбинационной репарации фага Т4, функционально гомологичны ферментам, участвующим в рекомбинационной репарации бактерий и эукариот. [94] В частности, что касается гена, необходимого для реакции обмена цепи, ключевого этапа гомологичной рекомбинационной репарации, существует функциональная гомология вирусов и человека (т.е. uvsX в фаге Т4; RecA в E. coli и других бактериях, а также rad51 и dmc1 у дрожжей и других эукариот, включая человека). [95] Реактивация множественности также была продемонстрирована у многих патогенных вирусов. [96]
[ править ]
Коронавирусы способны к генетической рекомбинации как минимум два вирусных генома , когда в одной зараженной клетке присутствуют . Рекомбинация РНК , по-видимому, является основной движущей силой в определении (1) генетической изменчивости внутри вида CoV, (2) способности вида CoV переходить от одного хозяина к другому и (3) редко появления новых CoV. [97] Механизм рекомбинации CoV, вероятно, включает переключение шаблона во время репликации генома. [97] Рекомбинация РНК-вирусов, по-видимому, является адаптацией, позволяющей справиться с повреждением генома. [98]
был введен в результате рекомбинации коронавирусов по-видимому , панголинов Весь мотив связывания рецептора пандемического SARS-CoV-2 , . [99] Такое событие рекомбинации могло стать решающим шагом в развитии способности SARS-CoV-2 заражать людей. [99] События рекомбинации, вероятно, являются ключевыми шагами в эволюционном процессе, который приводит к появлению новых коронавирусов человека. [100]
Во время пандемии COVID-19 в 2020 году многие геномные последовательности австралийских изолятов SARS-CoV-2 имели делеции или мутации (29742G>A или 29742G>U; «G19A» или «G19U») в 3'-стебельной петле II-контура коронавируса . подобный мотив (s2m) — мотив РНК в 3'-нетранслируемой области вирусного генома, что позволяет предположить, что события рекомбинации РНК могли происходить в s2m SARS-CoV-2. На основе компьютерного анализа 1319 последовательностей австралийского SARS-CoV-2 с использованием алгоритма Recco ( https://recco.bioinf.mpi-inf.mpg.de/ ), 29742G("G19"), 29744G("G21") и 29751G («G28») были предсказаны как горячие точки рекомбинации. [101]
Вспышка SARS-CoV-2 на круизном лайнере Diamond Princess , скорее всего, возникла либо от одного человека, инфицированного вариантом вируса, идентичного изоляту WIV04 в Ухане, либо одновременно с другим первичным случаем, инфицированным вирусом, содержащим мутацию 11083G > T. Анализ неравновесия по сцеплению подтвердил, что рекомбинация РНК с мутацией 11083G > T также способствует увеличению количества мутаций среди вирусного потомства. Результаты показывают, что мутация 11083G > T SARS-CoV-2 распространилась во время карантина на корабле и возникла в результате рекомбинации РНК de novo под положительным давлением отбора. Кроме того, у трех пациентов в этом круизе две мутации 29736G > T и 29751G > T («G13» и «G28») также были локализованы в 3'-подобном мотиве «стебель-петля II» коронавируса (s2m) , как «G28». был предсказан как горячие точки рекомбинации у австралийских мутантов SARS-CoV-2. Хотя s2m считается высококонсервативным мотивом РНК среди многих видов коронавирусов, этот результат также предполагает, что s2m SARS-CoV-2 скорее является мотивом РНК. РНК . Горячая точка рекомбинации/мутации [102]
дисфункции Последствия
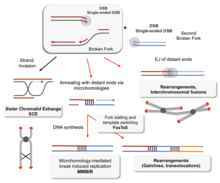
Без правильной гомологичной рекомбинации хромосомы часто неправильно выравниваются на первой фазе деления клеток в мейозе . Это приводит к тому, что хромосомы не могут правильно разделиться в процессе, называемом нерасхождением . В свою очередь, нерасхождение может привести к тому, что сперматозоиды и яйцеклетки будут иметь слишком мало или слишком много хромосом. Синдром Дауна , вызванный дополнительной копией хромосомы 21 , является одним из многих нарушений, возникающих в результате такого нарушения гомологичной рекомбинации в мейозе. [78] [103]
Нарушения гомологичной рекомбинации тесно связаны с образованием рака у людей. Например, каждое из связанных с раком заболеваний синдром Блума , синдром Вернера и синдром Ротмунда-Томсона вызваны неисправными копиями генов хеликаз RecQ, участвующих в регуляции гомологичной рекомбинации: BLM , WRN и RECQL4 соответственно. [104] В клетках больных синдромом Блума, у которых отсутствует рабочая копия белка BLM, наблюдается повышенная скорость гомологичной рекомбинации. [105] Эксперименты на мышах с дефицитом BLM показали, что эта мутация приводит к раку из-за потери гетерозиготности, вызванной усилением гомологичной рекомбинации. [106] Утрата гетерозиготности означает потерю одной из двух версий (или аллелей ) гена. Если один из утраченных аллелей помогает подавлять опухоли, как, например, ген белка ретинобластомы , то потеря гетерозиготности может привести к раку. [107] : 1236
Снижение скорости гомологичной рекомбинации приводит к неэффективной репарации ДНК. [107] : 310 что также может привести к раку. [108] Так обстоит дело с BRCA1 и BRCA2 , двумя схожими генами-супрессорами опухолей , чья неисправность связана со значительно повышенным риском развития рака груди и яичников . Клетки, в которых отсутствуют BRCA1 и BRCA2, имеют пониженную скорость гомологичной рекомбинации и повышенную чувствительность к ионизирующему излучению , что позволяет предположить, что снижение гомологичной рекомбинации приводит к повышенной восприимчивости к раку. [108] Поскольку единственная известная функция BRCA2 — помогать инициировать гомологичную рекомбинацию, исследователи предположили, что более детальное знание роли BRCA2 в гомологичной рекомбинации может быть ключом к пониманию причин рака молочной железы и яичников. [108]
Опухоли с дефицитом гомологичной рекомбинации (включая дефекты BRCA) описываются как HRD-положительные. [109]
сохранение Эволюционное

Хотя пути могут механически различаться, способность организмов осуществлять гомологичную рекомбинацию универсально сохраняется во всех сферах жизни. [110] Основываясь на сходстве их аминокислотных последовательностей, гомологи ряда белков можно обнаружить во многих сферах жизни, что указывает на то, что они эволюционировали давным-давно и с тех пор разошлись с общими предковыми белками. [110]
Члены семейства рекомбиназ RecA обнаружены почти во всех организмах с RecA у бактерий, Rad51 и DMC1 у эукариот, RadA у архей и UvsX у фага Т4 . [111]
Родственные одноцепочечные связывающие белки, которые важны для гомологичной рекомбинации и многих других процессов, также обнаруживаются во всех сферах жизни. [112]
Rad54, Mre11 , Rad50 и ряд других белков также обнаружены как у архей, так и у эукариот. [110] [111] [113]
Семейство RecA рекомбиназ
Считается, что белки семейства рекомбиназ RecA произошли от общей предковой рекомбиназы. [110] Семейство рекомбиназ RecA содержит белок RecA из бактерий , белки Rad51 и Dmc1 из эукариот, RadA из архей и белки-паралоги рекомбиназы. Исследования, моделирующие эволюционные взаимоотношения между белками Rad51, Dmc1 и RadA, показывают, что они монофилетичны или имеют общего молекулярного предка. [110] Внутри этого семейства белков Rad51 и Dmc1 сгруппированы в отдельную от RadA кладу . Одной из причин объединения этих трех белков в одну группу является то, что все они обладают модифицированным мотивом спираль-поворот-спираль , который помогает белкам связываться с ДНК по направлению к их N-концевым концам . [110] Древнее событие дупликации эукариотического гена RecA и последующая мутация были предложены в качестве вероятного происхождения современных генов RAD51 и DMC1. [110]
Белки обычно имеют длинную консервативную область, известную как домен RecA/Rad51. Внутри этого белкового домена имеются два мотива последовательности : мотив Уокера А и мотив Уокера В. Мотивы Уокера A и B позволяют членам семейства белков RecA/Rad51 участвовать в связывании АТФ и гидролизе АТФ . [110] [114]
Белки, мейоза специфичные для
Открытие Dmc1 у нескольких видов Giardia , одного из первых простейших , разошедшихся в эукариоты, предполагает, что мейотическая гомологичная рекомбинация – и, следовательно, сам мейоз – возникла очень рано в эволюции эукариот. [115] Помимо исследований Dmc1, исследования белка Spo11 предоставили информацию о происхождении мейотической рекомбинации. [116] Spo11, топоизомераза типа II , может инициировать гомологичную рекомбинацию в мейозе, создавая целевые двухцепочечные разрывы в ДНК. [25] Филогенетические деревья, основанные на последовательности генов, сходных с SPO11 у животных, грибов, растений, протистов и архей, привели ученых к мысли, что версия Spo11, существующая в настоящее время у эукариот, возникла у последнего общего предка эукариот и архей. [116]
приложения Технологические
Нацеливание на гены [ править ]

Многие методы введения последовательностей ДНК в организмы для создания рекомбинантной ДНК и генетически модифицированных организмов используют процесс гомологичной рекомбинации. [117] Этот метод, также называемый генным нацеливанием , особенно распространен в генетике дрожжей и мышей . Метод нацеливания на гены у нокаутных мышей использует эмбриональные стволовые клетки мыши для доставки искусственного генетического материала (в основном имеющего терапевтический интерес), который подавляет целевой ген мыши по принципу гомологичной рекомбинации. Таким образом, мышь выступает в качестве рабочей модели для понимания эффектов конкретного гена млекопитающих. В знак признания открытия того, как гомологичная рекомбинация может быть использована для введения генетических модификаций мышам посредством эмбриональных стволовых клеток, Марио Капечки , Мартин Эванс и Оливер Смитис были удостоены Нобелевской премии по физиологии и медицине 2007 года . [118]
Достижения в области технологий генного таргетинга, которые захватывают механизм гомологичной рекомбинации клеток, в настоящее время приводят к разработке новой волны более точных изогенных моделей заболеваний человека . Считается, что эти сконструированные модели человеческих клеток более точно отражают генетику заболеваний человека, чем их предшественники на мышиных моделях. Во многом это связано с тем, что представляющие интерес мутации вводятся в эндогенные гены точно так же, как они происходят у реальных пациентов, и потому, что они основаны на геномах человека, а не геномах крыс. Более того, некоторые технологии позволяют нокаутировать конкретную мутацию, а не просто нокаутировать, как это было со старыми технологиями нацеливания на гены.
Белковая инженерия [ править ]
Белковая инженерия с гомологичной рекомбинацией создает химерные белки путем замены фрагментов между двумя родительскими белками. Эти методы используют тот факт, что рекомбинация может обеспечить высокую степень разнообразия последовательностей , сохраняя при этом способность белка сворачиваться в его третичную структуру или трехмерную форму. [119] Это контрастирует с другими методами белковой инженерии, такими как случайный точечный мутагенез , при котором вероятность сохранения функции белка снижается экспоненциально с увеличением аминокислотных замен. [120] Химеры, полученные методами рекомбинации, способны сохранять свою способность сворачиваться, поскольку их замененные родительские фрагменты структурно и эволюционно консервативны. Эти рекомбинируемые «строительные блоки» сохраняют структурно важные взаимодействия, например, точки физического контакта между различными аминокислотами в структуре белка. Вычислительные методы, такие как SCHEMA и статистический анализ связи, могут использоваться для идентификации структурных субъединиц, подходящих для рекомбинации. [121] [122] [123]
Методы, основанные на гомологичной рекомбинации, использовались для создания новых белков. [121] В исследовании, опубликованном в 2007 году, исследователям удалось создать химеры двух ферментов, участвующих в биосинтезе изопреноидов , разнообразного класса соединений, включая гормоны , зрительные пигменты и некоторые феромоны . Химерные белки приобрели способность катализировать важную реакцию биосинтеза изопреноидов — одного из самых разнообразных путей биосинтеза , встречающихся в природе, — которая отсутствовала в исходных белках. [124] Белковая инженерия посредством рекомбинации также привела к созданию химерных ферментов с новой функцией в составе группы белков, известных как семейство цитохромов P450 . [125] который у человека участвует в детоксикации чужеродных соединений, таких как лекарства, пищевые добавки и консерванты. [22]
рака Терапия
Раковые клетки, способные к гомологичной рекомбинации (HRP), способны восстанавливать повреждения ДНК, вызванные химиотерапией, такой как цисплатин. Таким образом, рак HRP трудно поддается лечению. Исследования показывают, что гомологичную рекомбинацию можно нацелить посредством ингибирования c-Abl. [126] [127] Раковые клетки с мутациями BRCA имеют недостатки в гомологичной рекомбинации, и лекарства, использующие эти недостатки, были разработаны и успешно используются в клинических испытаниях. [128] [129] Олапариб , ингибитор PARP1, уменьшал или останавливал рост опухолей молочной железы , яичников и простаты, вызванных мутациями в генах BRCA1 или BRCA2 , которые необходимы для HR. Когда BRCA1 или BRCA2 отсутствуют, другие типы механизмов репарации ДНК должны компенсировать дефицит HR, такие как репарация с вырезанием оснований (BER) для остановленных вилок репликации или негомологическое соединение концов (NHEJ) для двухцепочечных разрывов. [128] Ингибируя BER в клетках с дефицитом HR, олапариб применяет концепцию синтетической летальности для специфического воздействия на раковые клетки. Хотя ингибиторы PARP1 представляют собой новый подход к терапии рака, исследователи предупреждают, что они могут оказаться недостаточными для лечения метастатического рака на поздних стадиях. [128] Раковые клетки могут стать устойчивыми к ингибитору PARP1, если в них будут удалены мутации в BRCA2, что подрывает синтетическую летальность препарата за счет восстановления способности раковых клеток восстанавливать ДНК с помощью HR. [130]
См. также [ править ]
Ссылки [ править ]
- ^ Томпсон Л.Х., Шильд Д. (июнь 2001 г.). «Гомологичная рекомбинационная репарация ДНК обеспечивает стабильность хромосом млекопитающих». Мутационные исследования . 477 (1–2): 131–53. дои : 10.1016/S0027-5107(01)00115-4 . ПМИД 11376695 .
- ^ Альбертс Б., Джонсон А., Льюис Дж., Рафф М., Робертс К., Уолтер П. и др. (2002). «Глава 5: Репликация, репарация и рекомбинация ДНК» . Молекулярная биология клетки (4-е изд.). Нью-Йорк: Garland Science. п. 845. ИСБН 978-0-8153-3218-3 . OCLC 145080076 .
- ^ Капечки М.Р. (июнь 1989 г.). «Изменение генома путем гомологичной рекомбинации». Наука . 244 (4910): 1288–92. Бибкод : 1989Sci...244.1288C . дои : 10.1126/science.2660260 . ПМИД 2660260 .
- ^ Смитис О, Грегг Р.Г., Боггс С.С., Коралевски М.А., Кучерлапати Р.С. (19 сентября 1985 г.). «Вставка последовательностей ДНК в хромосомный локус бета-глобина человека путем гомологичной рекомбинации». Природа . 317 (6034): 230–4. Бибкод : 1985Natur.317..230S . дои : 10.1038/317230a0 . ПМИД 2995814 . S2CID 30212766 .
- ^ Орр-Уивер Т.Л., Шостак Дж.В., Ротштейн Р.Дж. (октябрь 1981 г.). «Трансформация дрожжей: модельная система для изучения рекомбинации» . Труды Национальной академии наук Соединенных Штатов Америки . 78 (10): 6354–8. Бибкод : 1981PNAS...78.6354O . дои : 10.1073/pnas.78.10.6354 . ПМК 349037 . ПМИД 6273866 .
- ^ Орр-Уивер Т.Л., Шостак Дж.В. (июль 1983 г.). «Дрожжевая рекомбинация: связь между восстановлением двухцепочечной щели и кроссинговером» . Труды Национальной академии наук Соединенных Штатов Америки . 80 (14): 4417–21. Бибкод : 1983PNAS...80.4417O . дои : 10.1073/pnas.80.14.4417 . ПМЦ 384049 . ПМИД 6308623 .
- ^ Jump up to: Перейти обратно: а б Шостак Дж.В., Орр-Уивер Т.Л., Ротштейн Р.Дж., Шталь Ф.В. (май 1983 г.). «Модель репарации двухцепочечного разрыва для рекомбинации». Клетка . 33 (1): 25–35. дои : 10.1016/0092-8674(83)90331-8 . ПМИД 6380756 . S2CID 39590123 .
- ^ Резник М.А. (июнь 1976 г.). «Восстановление двухцепочечных разрывов ДНК; модель рекомбинации». Журнал теоретической биологии . 59 (1): 97–106. Бибкод : 1976JThBi..59...97R . дои : 10.1016/s0022-5193(76)80025-2 . ПМИД 940351 .
- ^ Джасин М., Ротштейн Р. (ноябрь 2013 г.). «Ремонт разрывов нитей путем гомологичной рекомбинации» . Перспективы Колд-Спринг-Харбор в биологии . 5 (11): а012740. doi : 10.1101/cshperspect.a012740 . ПМК 3809576 . ПМИД 24097900 .
- ^ Бейтсон П. (август 2002 г.). «Уильям Бейтсон: биолог, опередивший свое время» (PDF) . Журнал генетики . 81 (2): 49–58. дои : 10.1007/BF02715900 . ПМИД 12532036 . S2CID 26806110 .
- ^ «Реджинальд Крандалл Паннетт» . NAHSTE, Эдинбургский университет . Проверено 3 июля 2010 г.
- ^ Лобо I, Шоу К. (2008). «Томас Хант Морган, генетическая рекомбинация и картирование генов» . Природное образование . 1 (1).
- ^ Jump up to: Перейти обратно: а б Коу Э., Касс Л.Б. (май 2005 г.). «Доказательство физического обмена генами в хромосомах» . Труды Национальной академии наук Соединенных Штатов Америки . 102 (19): 6641–6. Бибкод : 2005PNAS..102.6641C . дои : 10.1073/pnas.0407340102 . ПМЦ 1100733 . ПМИД 15867161 .
- ^ Крейтон Х.Б., МакКлинток Б. (август 1931 г.). «Корреляция цитологического и генетического кроссинговера у Zea Mays» . Труды Национальной академии наук Соединенных Штатов Америки . 17 (8): 492–7. Бибкод : 1931PNAS...17..492C . дои : 10.1073/pnas.17.8.492 . ПМЦ 1076098 . ПМИД 16587654 .
- ^ Стерн, К. (1931). «Цитолого-генетические исследования как доказательство теории обмена факторами Моргана». Центральный биологический журнал . 51 :547-587.
- ^ «Развитие генетики бактерий» . Национальная медицинская библиотека США . Проверено 3 июля 2010 г.
- ^ «Нобелевская премия по физиологии и медицине 1958 года» . Нобелевская премия.org . Проверено 3 июля 2010 г.
- ^ Jump up to: Перейти обратно: а б с Хабер Дж.Э., Ира Г., Малкова А., Сугавара Н. (январь 2004 г.). «Восстановление двухцепочечного разрыва хромосомы путем гомологичной рекомбинации: пересмотр модели Робина Холлидея» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 359 (1441): 79–86. дои : 10.1098/rstb.2003.1367 . ПМК 1693306 . ПМИД 15065659 .
- ^ Лодиш Х., Берк А., Зипурски С.Л., Мацудайра П., Балтимор Д., Дарнелл Дж. (2000). «12.5: Рекомбинация между гомологичными участками ДНК: двухцепочечные разрывы в ДНК инициируют рекомбинацию». Молекулярно-клеточная биология (4-е изд.). WH Фриман и компания. ISBN 978-0-7167-3136-8 .
- ^ Гриффитс А. и др. (1999). «8: Хромосомные мутации: хромосомные перестройки» . Современный генетический анализ . WH Фриман и компания. ISBN 978-0-7167-3118-4 .
- ^ Ханна К.К., Джексон С.П. (март 2001 г.). «Двухцепочечные разрывы ДНК: передача сигналов, восстановление и связь с раком». Природная генетика . 27 (3): 247–54. дои : 10.1038/85798 . ПМИД 11242102 . S2CID 3012823 .
- ^ Jump up to: Перейти обратно: а б с Нельсон Д.Л., Кокс М.М. (2005). Принципы биохимии (4-е изд.). Фриман. стр. 980–981 . ISBN 978-0-7167-4339-2 .
- ^ Маркон Э., Моенс П.Б. (август 2005 г.). «Эволюция мейоза: рекрутирование и модификация соматических белков, восстанавливающих ДНК». Биоэссе . 27 (8): 795–808. doi : 10.1002/bies.20264 . ПМИД 16015600 . S2CID 27658497 .
- ^ Альбертс Б., Джонсон А., Льюис Дж., Рафф М., Робертс К., Уолтер П. (2008). Молекулярная биология клетки (5-е изд.). Гирляндная наука. п. 305 . ISBN 978-0-8153-4105-5 .
- ^ Jump up to: Перейти обратно: а б Кини С., Жиру С.Н., Клекнер Н. (февраль 1997 г.). «Специфические для мейоза двухцепочечные разрывы ДНК катализируются Spo11, членом широко консервативного семейства белков» . Клетка . 88 (3): 375–84. дои : 10.1016/S0092-8674(00)81876-0 . ПМИД 9039264 . S2CID 8294596 .
- ^ Член парламента Лонгезе, Бонетти Д., Герини И., Манфрини Н., Клеричи М. (сентябрь 2009 г.). «Двухцепочечные разрывы ДНК при мейозе: проверка их образования, процессинга и восстановления». Восстановление ДНК . 8 (9): 1127–38. дои : 10.1016/j.dnarep.2009.04.005 . ПМИД 19464965 .
- ^ Кэхилл Л.П., Мариана Дж.К., Молеон П. (январь 1979 г.). «Общее количество фолликулов у овец с высоким и низким уровнем овуляции» . Журнал репродукции и фертильности . 55 (1): 27–36. дои : 10.1530/jrf.0.0550027 . ПМИД 423159 .
- ^ Шиммель Дж., ван Шендель Р., ден Даннен Дж. Т., Тейстерман М. (сентябрь 2019 г.). «Шаблонные вставки: дымящийся пистолет для тета-опосредованного полимеразным соединением концов» . Тенденции в генетике . 35 (9): 632–644. дои : 10.1016/j.tig.2019.06.001 . ПМИД 31296341 . S2CID 195892718 .
- ^ Альбертс Б., Джонсон А., Льюис Дж., Рафф М., Робертс К., Уолтер П. (2008). Молекулярная биология клетки (5-е изд.). Гирляндная наука. п. 303 . ISBN 978-0-8153-4105-5 .
- ^ Jump up to: Перейти обратно: а б Шривастав М., Де Аро Л.П., Николофф Дж.А. (январь 2008 г.). «Регуляция выбора пути восстановления двухцепочечного разрыва ДНК» . Клеточные исследования . 18 (1): 134–47. дои : 10.1038/cr.2007.111 . ПМИД 18157161 .
- ^ Jump up to: Перейти обратно: а б Мимиту Э.П., Симингтон Л.С. (май 2009 г.). «Нуклеазы и геликазы занимают центральное место в гомологичной рекомбинации». Тенденции биохимических наук . 34 (5): 264–72. дои : 10.1016/j.tibs.2009.01.010 . ПМИД 19375328 .
- ^ Андрес, Сара Н.; Уильямс, Р. Скотт (август 2017 г.). «CtIP/Ctp1/Sae2, молекулярная форма, подходящая для выполнения функции» . Восстановление ДНК . 56 : 109–117. дои : 10.1016/j.dnarep.2017.06.013 . ПМЦ 5543718 . ПМИД 28623092 .
- ^ Уэртас П., Кортес-Ледесма Ф, Сартори А.А., Агилера А., Джексон С.П. (октябрь 2008 г.). «CDK нацелен на Sae2 для контроля резекции концов ДНК и гомологичной рекомбинации» . Природа . 455 (7213): 689–92. Бибкод : 2008Natur.455..689H . дои : 10.1038/nature07215 . ПМЦ 2635538 . ПМИД 18716619 .
- ^ Лю Б, Ип РК, Чжоу Цз (2012). «Ремоделирование хроматина, восстановление повреждений ДНК и старение» . Курс. Геномика . 13 (7): 533–47. дои : 10.2174/138920212803251373 . ПМЦ 3468886 . ПМИД 23633913 .
- ^ Jump up to: Перейти обратно: а б с д Селлу Х., Лебопен Т., Шапюи С., Смит Р., Хегеле А., Сингх Х.Р., Козловски М., Бультманн С., Ладурнер А.Г., Тимински Г., Уэт С. (2016). «Поли(АДФ-рибоза)-зависимый ремодератор хроматина Alc1 вызывает локальную релаксацию хроматина при повреждении ДНК» . Мол. Биол. Клетка . 27 (24): 3791–3799. дои : 10.1091/mbc.E16-05-0269 . ПМК 5170603 . ПМИД 27733626 .
- ^ Jump up to: Перейти обратно: а б Ван Метер М., Саймон М., Томблайн Г., Мэй А., Морелло Т.Д., Хаббард Б.П., Бредбеннер К., Парк Р., Синклер Д.А., Бор В.А., Горбунова В., Селуанов А. (2016). «JNK фосфорилирует SIRT6, чтобы стимулировать восстановление двухцепочечных разрывов ДНК в ответ на окислительный стресс путем привлечения PARP1 к разрывам ДНК» . Представитель ячейки . 16 (10): 2641–50. дои : 10.1016/j.celrep.2016.08.006 . ПМК 5089070 . ПМИД 27568560 .
- ^ Jump up to: Перейти обратно: а б Хейнс Дж. Ф., Макдональд Д., Родриг А., Дери У., Массон Дж. Ю., Хендзель М. Дж., Пуарье Г. Г. (2008). «PARP1-зависимая кинетика привлечения белков MRE11 и NBS1 к множественным сайтам повреждения ДНК» . Ж. Биол. Хим . 283 (2): 1197–208. дои : 10.1074/jbc.M706734200 . ПМИД 18025084 .
- ^ Jump up to: Перейти обратно: а б с Рогаков Е.П., Пильч Д.Р., Орр А.Х., Иванова В.С., Боннер В.М. (1998). «Двухцепочечные разрывы ДНК индуцируют фосфорилирование гистона H2AX по серину 139» . Ж. Биол. Хим . 273 (10): 5858–68. дои : 10.1074/jbc.273.10.5858 . ПМИД 9488723 .
- ^ Майланд Н., Беккер-Йенсен С., Фауструп Х., Меландер Ф., Бартек Дж., Лукас С., Лукас Дж. (2007). «RNF8 убиквитилирует гистоны в местах двухцепочечных разрывов ДНК и способствует сборке репарационных белков» . Клетка . 131 (5): 887–900. дои : 10.1016/j.cell.2007.09.040 . ПМИД 18001824 . S2CID 14232192 .
- ^ Луистербург М.С., Акс К., Акерманн Л., Вигант В.В., Беккер-Йенсен С., Ларсен Д.Х., Ханна К.К., ван Аттикум Х., Майланд Н., Дантума Н.П. (2012). «Новая некаталитическая роль убиквитинлигазы RNF8 в разворачивании структуры хроматина высшего порядка» . ЭМБО Дж . 31 (11): 2511–27. дои : 10.1038/emboj.2012.104 . ПМЦ 3365417 . ПМИД 22531782 .
- ^ Бода Ф., Буард Дж., Грей С., Фледель-Алон А., Обер С., Пшеворски М. и др. (февраль 2010 г.). «PRDM9 является основным детерминантом горячих точек мейотической рекомбинации у людей и мышей» . Наука . 327 (5967): 836–40. Бибкод : 2010Sci...327..836B . дои : 10.1126/science.1183439 . ПМК 4295902 . ПМИД 20044539 .
- ^ Уэллс Д., Битоун Э., Моралли Д., Чжан Г., Хинч А., Янковска Дж. и др. (август 2020 г.). «ZCWPW1 рекрутируется в горячие точки рекомбинации с помощью PRDM9 и необходим для мейотического восстановления двухцепочечных разрывов» . электронная жизнь . 9 : е53392. дои : 10.7554/eLife.53392 . ПМЦ 7494361 . ПМИД 32744506 .
- ^ Jump up to: Перейти обратно: а б с д Сунг П., Кляйн Х. (октябрь 2006 г.). «Механизм гомологичной рекомбинации: медиаторы и хеликазы берут на себя регуляторные функции». Nature Reviews Молекулярно-клеточная биология . 7 (10): 739–50. дои : 10.1038/nrm2008 . ПМИД 16926856 . S2CID 30324005 .
- ^ Уолд М.С. (1997). «Репликационный белок А: гетеротримерный одноцепочечный ДНК-связывающий белок, необходимый для метаболизма эукариотической ДНК». Ежегодный обзор биохимии . 66 : 61–92. doi : 10.1146/annurev.biochem.66.1.61 . ПМИД 9242902 .
- ^ МакМахилл М.С., Шам К.В., Бишоп Д.К. (ноябрь 2007 г.). «Синтез-зависимый отжиг цепи при мейозе» . ПЛОС Биология . 5 (11): е299. doi : 10.1371/journal.pbio.0050299 . ПМК 2062477 . ПМИД 17988174 .

- ^ Берч С., Канг Л.Е., Симингтон Л.С. (февраль 2000 г.). «RAD51 необходим для восстановления разрывов двухцепочечной ДНК плазмиды из плазмидной или хромосомной матрицы» . Молекулярная и клеточная биология . 20 (4): 1194–205. дои : 10.1128/MCB.20.4.1194-1205.2000 . ПМЦ 85244 . ПМИД 10648605 .
- ^ Альбертс Б., Джонсон А., Льюис Дж., Рафф М., Робертс К., Уолтер П. (2008). Молекулярная биология клетки (5-е изд.). Гирляндная наука. стр. 312–313. ISBN 978-0-8153-4105-5 .
- ^ Jump up to: Перейти обратно: а б с Хелледей Т., Ло Джей, ван Гент, округ Колумбия, Энгельвард Б.П. (июль 2007 г.). «Восстановление двухцепочечных разрывов ДНК: от механистического понимания к лечению рака». Восстановление ДНК . 6 (7): 923–35. дои : 10.1016/j.dnarep.2007.02.006 . ПМИД 17363343 .
- ^ Jump up to: Перейти обратно: а б Андерсен С.Л., Секельски Дж. (декабрь 2010 г.). «Мейотическая и митотическая рекомбинация: два разных пути восстановления двухцепочечного разрыва: разные функции мейотической и митотической репарации DSB отражаются в разном использовании путей и разных результатах» . Биоэссе . 32 (12): 1058–66. doi : 10.1002/bies.201000087 . ПМК 3090628 . ПМИД 20967781 .
- ^ Аллерс Т., Лихтен М. (июль 2001 г.). «Дифференциальное время и контроль некроссоверной и кроссоверной рекомбинации во время мейоза» . Клетка . 106 (1): 47–57. дои : 10.1016/s0092-8674(01)00416-0 . ПМИД 11461701 . S2CID 1878863 .
- ^ Лаборатория Хабера. «Однониточный отжиг» . Университет Брандейса. Архивировано из оригинала 19 января 2015 года . Проверено 3 июля 2010 г.
- ^ Jump up to: Перейти обратно: а б с Линдакер А.М., Алани Э. (март 2009 г.). «Сказка о хвостах: понимание координации обработки 3'-конца во время гомологичной рекомбинации» . Биоэссе . 31 (3): 315–21. doi : 10.1002/bies.200800195 . ПМЦ 2958051 . ПМИД 19260026 .
- ^ Мимиту Э.П., Симингтон Л.С. (сентябрь 2009 г.). «Резекция концов ДНК: многие нуклеазы облегчают работу» . Восстановление ДНК . 8 (9): 983–95. дои : 10.1016/j.dnarep.2009.04.017 . ПМК 2760233 . ПМИД 19473888 .
- ^ Пакес Ф., Хабер Дж.Э. (июнь 1999 г.). «Множественные пути рекомбинации, индуцированные двухцепочечными разрывами у Saccharomyces cerevisiae» . Обзоры микробиологии и молекулярной биологии . 63 (2): 349–404. дои : 10.1128/MMBR.63.2.349-404.1999 . ПМК 98970 . ПМИД 10357855 .
- ^ Jump up to: Перейти обратно: а б Макихерн М.Дж., Хабер Дж.Э. (2006). «Репликация, индуцированная разрывом, и рекомбинационное удлинение теломер у дрожжей». Ежегодный обзор биохимии . 75 : 111–35. doi : 10.1146/annurev.biochem.74.082803.133234 . ПМИД 16756487 .
- ^ Морриш Т.А., Грейдер К.В. (январь 2009 г.). Хабер Дж. Э. (ред.). «Короткие теломеры инициируют рекомбинацию теломер в первичных и опухолевых клетках» . ПЛОС Генетика . 5 (1): e1000357. дои : 10.1371/journal.pgen.1000357 . ПМЦ 2627939 . ПМИД 19180191 .
- ^ Мунтони А., Реддел Р.Р. (октябрь 2005 г.). «Первые молекулярные подробности АЛТ в опухолевых клетках человека». Молекулярная генетика человека . 14 Спецификация № 2 (выпуск обзора 2): R191–6. дои : 10.1093/hmg/ddi266 . ПМИД 16244317 .
- ^ PDB : 3 см ; Чен З, Ян Х, Павлетич Н.П. (май 2008 г.). «Механизм гомологичной рекомбинации структур RecA-ssDNA/dsDNA». Природа . 453 (7194): 489–4. Бибкод : 2008Natur.453..489C . дои : 10.1038/nature06971 . ПМИД 18497818 . S2CID 4416531 .
- ^ Ковальчиковски С.К., Диксон Д.А., Эгглстон А.К., Лаудер С.Д., Рехауэр В.М. (сентябрь 1994 г.). «Биохимия гомологичной рекомбинации у Escherichia coli» . Микробиологические обзоры . 58 (3): 401–65. дои : 10.1128/MMBR.58.3.401-465.1994 . ПМК 372975 . ПМИД 7968921 .
- ^ Роча EP, Корнет Э, Мишель Б (август 2005 г.). «Сравнительный и эволюционный анализ бактериальных гомологичных рекомбинационных систем» . ПЛОС Генетика . 1 (2): е15. дои : 10.1371/journal.pgen.0010015 . ПМК 1193525 . ПМИД 16132081 .
- ^ Jump up to: Перейти обратно: а б Амундсен С.К., Тейлор А.Ф., Редди М., Смит Г.Р. (декабрь 2007 г.). «Межсубъединичная передача сигналов в ферменте RecBCD, сложной белковой машине, регулируемой горячими точками Чи» . Гены и развитие . 21 (24): 3296–307. дои : 10.1101/gad.1605807 . ПМК 2113030 . ПМИД 18079176 .
- ^ Синглтон М.Р., Диллингем М.С., Годье М., Ковальчиковски С.К., Вигли Д.Б. (ноябрь 2004 г.). «Кристаллическая структура фермента RecBCD раскрывает механизм обработки разрывов ДНК» (PDF) . Природа . 432 (7014): 187–93. Бибкод : 2004Natur.432..187S . дои : 10.1038/nature02988 . ПМИД 15538360 . S2CID 2916995 . Архивировано из оригинала (PDF) 25 мая 2004 г.
- ^ Кроми, Джорджия (август 2009 г.). «Филогенетическая повсеместность и перетасовка бактериальных рекомбинационных комплексов RecBCD и AddAB» . Журнал бактериологии . 191 (16): 5076–84. дои : 10.1128/JB.00254-09 . ПМЦ 2725590 . ПМИД 19542287 .
- ^ Jump up to: Перейти обратно: а б Смит Г.Р. (июнь 2012 г.). «Как фермент RecBCD и Chi способствуют восстановлению и рекомбинации разрывов ДНК: взгляд молекулярного биолога» . Обзоры микробиологии и молекулярной биологии . 76 (2): 217–28. дои : 10.1128/MMBR.05026-11 . ПМЦ 3372252 . ПМИД 22688812 .
- ^ Jump up to: Перейти обратно: а б с д Диллингем М.С., Ковальчиковски СК (декабрь 2008 г.). «Фермент RecBCD и восстановление двухцепочечных разрывов ДНК» . Обзоры микробиологии и молекулярной биологии . 72 (4): 642–71, Оглавление. дои : 10.1128/MMBR.00020-08 . ПМЦ 2593567 . ПМИД 19052323 .
- ^ Мишель Б., Бубакри Х., Бахароглу З., ЛеМассон М., Лестини Р. (июль 2007 г.). «Рекомбинационные белки и спасение арестованных репликационных вилок». Восстановление ДНК . 6 (7): 967–80. дои : 10.1016/j.dnarep.2007.02.016 . ПМИД 17395553 .
- ^ Спайс М., Бьянко П.Р., Диллингем М.С., Ханда Н., Баскин Р.Дж., Ковальчиковски С.К. (сентябрь 2003 г.). «Молекулярный дроссель: горячая точка рекомбинации chi контролирует транслокацию ДНК с помощью геликазы RecBCD» . Клетка . 114 (5): 647–54. дои : 10.1016/S0092-8674(03)00681-0 . PMID 13678587 . S2CID 16662983 .
- ^ Тейлор А.Ф., Смит Г.Р. (июнь 2003 г.). «Фермент RecBCD представляет собой ДНК-геликазу с быстрым и медленным моторами противоположной полярности». Природа . 423 (6942): 889–93. Бибкод : 2003Natur.423..889T . дои : 10.1038/nature01674 . ПМИД 12815437 . S2CID 4302346 .
- ^ Спайс М., Амитани И., Баскин Р.Дж., Ковальчиковски С.К. (ноябрь 2007 г.). «Ферментные переключатели RecBCD приводят моторные субъединицы в ответ на распознавание ци» . Клетка . 131 (4): 694–705. дои : 10.1016/j.cell.2007.09.023 . ПМК 2151923 . ПМИД 18022364 .
- ^ Савир Ю., Тлусти Т. (ноябрь 2010 г.). «Поиск гомологии, опосредованный RecA, как почти оптимальная система обнаружения сигналов» (PDF) . Молекулярная клетка . 40 (3): 388–96. arXiv : 1011.4382 . Бибкод : 2010arXiv1011.4382S . doi : 10.1016/j.molcel.2010.10.020 . ПМИД 21070965 . S2CID 1682936 . Архивировано из оригинала (PDF) 7 октября 2012 г. Проверено 31 августа 2011 г.
- ^ Рэмбо Р.П., Уильямс Дж.Дж., Тайнер Дж.А. (ноябрь 2010 г.). «Достижение точности гомологичной рекомбинации, несмотря на чрезвычайную сложность: обоснованные решения с помощью молекулярного профилирования» (PDF) . Молекулярная клетка . 40 (3): 347–8. doi : 10.1016/j.molcel.2010.10.032 . ПМК 3003302 . ПМИД 21070960 . Архивировано из оригинала (PDF) 7 октября 2012 г. Проверено 31 августа 2011 г.
- ^ Де Вламинк И., ван Лёнхаут М.Т., Цвайфель Л., ден Бланкен Дж., Хунинг К., Хаге С. и др. (июнь 2012 г.). «Механизм распознавания гомологии при рекомбинации ДНК в экспериментах с двумя молекулами» . Молекулярная клетка . 46 (5): 616–24. doi : 10.1016/j.molcel.2012.03.029 . ПМИД 22560720 .
- ^ Альбертс Б., Джонсон А., Льюис Дж., Рафф М., Робертс К., Уолтер П. (2008). Молекулярная биология клетки (5-е изд.). Гирляндная наука. п. 307 . ISBN 978-0-8153-4105-5 .
- ^ Моримацу К., Ковальчиковски СК (май 2003 г.). «Белки RecFOR загружают белок RecA в ДНК с разрывами, чтобы ускорить обмен цепей ДНК: универсальный этап рекомбинационной репарации» . Молекулярная клетка . 11 (5): 1337–47. дои : 10.1016/S1097-2765(03)00188-6 . ПМИД 12769856 .
- ^ Хиом К. (июль 2009 г.). «Репарация ДНК: общие подходы к исправлению двухцепочечных разрывов» . Современная биология . 19 (13): R523–5. дои : 10.1016/j.cub.2009.06.009 . ПМИД 19602417 . S2CID 2221866 .
- ^ Ханда Н., Моримацу К., Ловетт С.Т., Ковальчиковски СК (май 2009 г.). «Восстановление начальных этапов восстановления разрыва дцДНК по пути RecF E. coli» . Гены и развитие . 23 (10): 1234–45. дои : 10.1101/gad.1780709 . ПМЦ 2685532 . ПМИД 19451222 .
- ^ Jump up to: Перейти обратно: а б Западный СК (июнь 2003 г.). «Молекулярные взгляды на рекомбинационные белки и их контроль». Nature Reviews Молекулярно-клеточная биология . 4 (6): 435–45. дои : 10.1038/nrm1127 . ПМИД 12778123 . S2CID 28474965 .
- ^ Jump up to: Перейти обратно: а б с д Уотсон Дж.Д., Бейкер Т.А., Белл С.П., Ганн А., Левин М., Лосик Р. (2003). Молекулярная биология гена (5-е изд.). Пирсон/Бенджамин Каммингс. стр. 259–291 . ISBN 978-0-8053-4635-0 .
- ^ Гумбинер-Руссо Л.М., Розенберг С.М. (28 ноября 2007 г.). Сэндлер С. (ред.). «Физический анализ продуктов гетеродуплексной рекомбинации E. coli in vivo: о распространенности 5'- и 3'-патчей» . ПЛОС ОДИН . 2 (11): е1242. Бибкод : 2007PLoSO...2.1242G . дои : 10.1371/journal.pone.0001242 . ПМК 2082072 . ПМИД 18043749 .
- ^ Томас К.М., Нильсен К.М. (сентябрь 2005 г.). «Механизмы и препятствия горизонтального переноса генов между бактериями» (PDF) . Обзоры природы. Микробиология . 3 (9): 711–21. дои : 10.1038/nrmicro1234 . ПМИД 16138099 . S2CID 1231127 . Архивировано из оригинала (PDF) 1 июня 2010 г.
- ^ Вулич М., Дионисио Ф., Таддей Ф., Радман М. (сентябрь 1997 г.). «Молекулярные ключи к видообразованию: полиморфизм ДНК и контроль генетического обмена у энтеробактерий» . Труды Национальной академии наук Соединенных Штатов Америки . 94 (18): 9763–7. Бибкод : 1997PNAS...94.9763V . дои : 10.1073/pnas.94.18.9763 . ПМК 23264 . ПМИД 9275198 .
- ^ Маевски Дж., Кохан FM (январь 1998 г.). «Влияние восстановления несоответствия и образования гетеродуплексов на сексуальную изоляцию у Bacillus» . Генетика . 148 (1): 13–8. дои : 10.1093/генетика/148.1.13 . ПМЦ 1459767 . ПМИД 9475717 .
- ^ Маевски Дж., Завадски П., Пикерилл П., Кохан Ф.М., Доусон К.Г. (февраль 2000 г.). «Барьеры для генетического обмена между видами бактерий: трансформация Streptococcus pneumoniae» . Журнал бактериологии . 182 (4): 1016–23. дои : 10.1128/JB.182.4.1016-1023.2000 . ПМК 94378 . ПМИД 10648528 .
- ^ Чен И, Дубнау Д (март 2004 г.). «Поглощение ДНК во время бактериальной трансформации». Обзоры природы. Микробиология . 2 (3): 241–9. дои : 10.1038/nrmicro844 . ПМИД 15083159 . S2CID 205499369 .
- ^ Клаверис Дж.П., Мартин Б., Полард П. (май 2009 г.). «Машина генетической трансформации: состав, локализация и механизм». Обзоры микробиологии FEMS . 33 (3): 643–56. дои : 10.1111/j.1574-6976.2009.00164.x . ПМИД 19228200 .
- ^ Кидане Д., Грауманн П.Л. (июль 2005 г.). «Внутриклеточная динамика белков и ДНК в компетентных клетках Bacillus subtilis» . Клетка . 122 (1): 73–84. дои : 10.1016/j.cell.2005.04.036 . ПМИД 16009134 . S2CID 17272331 .
- ^ Флейшманн-младший WR (1996). «43» . Медицинская микробиология (4-е изд.). Медицинский филиал Техасского университета в Галвестоне. ISBN 978-0-9631172-1-2 .
- ^ Jump up to: Перейти обратно: а б с Надь П.Д., Буярски Дж.Дж. (январь 1996 г.). «Гомологичная рекомбинация РНК в вирусе бромной мозаики: последовательности, богатые AU, снижают точность кроссоверов» . Журнал вирусологии . 70 (1): 415–26. doi : 10.1128/JVI.70.1.415-426.1996 . ЧВК 189831 . ПМИД 8523555 .
- ^ Бони М.Ф., де Йонг, доктор медицинских наук, ван Доорн, Х.Р., Холмс Э.К. (3 мая 2010 г.). Мартин Д.П. (ред.). «Руководство по выявлению событий гомологичной рекомбинации в вирусе гриппа А» . ПЛОС ОДИН . 5 (5): е10434. Бибкод : 2010PLoSO...510434B . дои : 10.1371/journal.pone.0010434 . ПМЦ 2862710 . ПМИД 20454662 .
- ^ Четверин А.Б. (октябрь 1999 г.). «Загадка рекомбинации РНК» . Письма ФЭБС . 460 (1): 1–5. дои : 10.1016/S0014-5793(99)01282-X . ПМК 7163957 . ПМИД 10571050 .
- ^ Jump up to: Перейти обратно: а б Руссинк MJ (сентябрь 1997 г.). «Механизмы эволюции растительных вирусов». Ежегодный обзор фитопатологии . 35 : 191–209. doi : 10.1146/annurev.phyto.35.1.191 . ПМИД 15012521 .
- ^ Арбакл Дж. Х., Медвечки П. Г. (август 2011 г.). «Молекулярная биология латентного периода вируса герпеса-6 человека и интеграции теломер» . Микробы и инфекция / Институт Пастера . 13 (8–9): 731–41. дои : 10.1016/j.micinf.2011.03.006 . ПМК 3130849 . ПМИД 21458587 .
- ^ Бернштейн С. (март 1981 г.). «Репарация дезоксирибонуклеиновой кислоты в бактериофаге» . Микробиологические обзоры . 45 (1): 72–98. дои : 10.1128/ММБР.45.1.72-98.1981 . ПМК 281499 . ПМИД 6261109 .
- ^ Бернштейн С, Бернштейн Х (2001). Репарация ДНК в бактериофаге. В: Николофф Дж.А., Хоекстра М.Ф. (ред.) Повреждение и восстановление ДНК, Том 3. Прогресс от фага к человеку. Humana Press, Тотова, Нью-Джерси, стр. 1–19. ISBN 978-0896038035
- ^ История Р.М., Бишоп Д.К., Клекнер Н., Стейц Т.А. (март 1993 г.). «Структурная связь бактериальных белков RecA с рекомбинационными белками бактериофага Т4 и дрожжей». Наука . 259 (5103): 1892–6. Бибкод : 1993Sci...259.1892S . дои : 10.1126/science.8456313 . ПМИД 8456313 .
- ^ Мишод Р.Э., Бернштейн Х., Недельку А.М. (май 2008 г.). «Адаптационное значение пола у микробных патогенов». Инфекция, генетика и эволюция . 8 (3): 267–85. дои : 10.1016/j.meegid.2008.01.002 . ПМИД 18295550 . http://www.hummingbirds.arizona.edu/Faculty/Michod/Downloads/IGE%20review%20sex.pdf
- ^ Jump up to: Перейти обратно: а б Су, Шуо; Вонг, Гэри; Ши, Вэйфэн; Лю, Цзюнь; Лай, Александр СК; Чжоу, Цзиюнь; Лю, Вэньцзюнь; Би, Юхай; Гао, Джордж Ф. (2016). «Эпидемиология, генетическая рекомбинация и патогенез коронавирусов» . Тенденции в микробиологии . 24 (6): 490–502. дои : 10.1016/j.tim.2016.03.003 . ПМК 7125511 . ПМИД 27012512 .
- ^ Барр, Дж. Н.; Фернс, Р. (2010). «Как РНК-вирусы сохраняют целостность своего генома» . Журнал общей вирусологии . 91 (6): 1373–1387. дои : 10.1099/vir.0.020818-0 . ПМИД 20335491 .
- ^ Jump up to: Перейти обратно: а б Ли, Сяоцзюнь; Георгий, Елена Евгеньевна; Маричаннегоуда, Манукумар Хонаяканахалли; Фоли, Брайан; Сяо, Чуан; Конг, Сян-Пэн; Чен, Юэ; Гнанакаран, С.; Корбер, Бетт; Гао, Фэн (2020). «Появление SARS-CoV-2 в результате рекомбинации и сильного очищающего отбора» . Достижения науки . 6 (27). Бибкод : 2020SciA....6.9153L . дои : 10.1126/sciadv.abb9153 . ПМЦ 7458444 . ПМИД 32937441 .
- ^ Рехман, Саиф ур; Шафик, Лайба; Ихсан, Авайс; Лю, Цинъю (2020). «Эволюционная траектория появления нового коронавируса SARS-CoV-2» . Патогены . 9 (3): 240. doi : 10.3390/pathogens9030240 . ПМЦ 7157669 . ПМИД 32210130 .
- ^ Да, Тай, Гран-при Контрераса (июль 2020 г.). «Появление вирусных мутантов в Австралии предполагает событие рекомбинации РНК в геноме SARS-CoV-2» . Медицинский журнал Австралии . 213 (1): 44–44.д1. дои : 10.5694/mja2.50657 . ПМК 7300921 . ПМИД 32506536 .
- ^ Да, Тай, Контрерас GP (1 июля 2021 г.). «Передача вируса и динамика развития SARS-CoV-2 в условиях корабельного карантина» . Бык. Всемирный орган здравоохранения . 99 (7): 486–495. дои : 10.2471/BLT.20.255752 . ПМЦ 8243027 . ПМИД 34248221 .
- ^ Лэмб Н.Э., Ю.К., Шаффер Дж., Фейнгольд Э., Шерман С.Л. (январь 2005 г.). «Связь между возрастом матери и мейотической рекомбинацией при трисомии 21» . Американский журнал генетики человека . 76 (1): 91–9. дои : 10.1086/427266 . ПМК 1196437 . ПМИД 15551222 .
- ^ Лаборатория Колд-Спринг-Харбор (2007). «Человеческие геликазы RecQ, гомологичная рекомбинация и геномная нестабильность» . ScienceDaily . Проверено 3 июля 2010 г.
- ^ Модести М, Канаар Р (2001). «Гомологичная рекомбинация: от модельных организмов к болезням человека» . Геномная биология . 2 (5): ОБЗОРЫ1014. doi : 10.1186/gb-2001-2-5-reviews1014 . ПМЦ 138934 . ПМИД 11387040 .
- ^ Луо Г., Санторо И.М., МакДэниел Л.Д., Нисидзима И., Миллс М., Юсуфян Х., Фогель Х., Шульц Р.А., Брэдли А. (декабрь 2000 г.). «Предрасположенность к раку, вызванная повышенной митотической рекомбинацией у мышей Блума». Природная генетика . 26 (4): 424–9. дои : 10.1038/82548 . ПМИД 11101838 . S2CID 21218975 .
- ^ Jump up to: Перейти обратно: а б Альбертс Б., Джонсон А., Льюис Дж., Рафф М., Робертс К., Уолтер П. (2007). Молекулярная биология клетки (5-е изд.). Гирляндная наука. ISBN 978-0-8153-4110-9 .
- ^ Jump up to: Перейти обратно: а б с Пауэлл С.Н., Качник Л.А. (сентябрь 2003 г.). «Роли BRCA1 и BRCA2 в гомологичной рекомбинации, точности репликации ДНК и клеточном ответе на ионизирующее излучение». Онкоген . 22 (37): 5784–91. дои : 10.1038/sj.onc.1206678 . ПМИД 12947386 .
- ^ «Использование показателя дефицита гомологичной рекомбинации (HRD) для выявления чувствительных к нирапарибу опухолей яичников высокой степени злокачественности» . Архивировано из оригинала 30 апреля 2017 г. Проверено 30 декабря 2016 г.
- ^ Jump up to: Перейти обратно: а б с д и ж г час Лин З, Конг Х, Ней М, Ма Х (июль 2006 г.). «Происхождение и эволюция семейства генов RecA/RAD51: доказательства древней дупликации генов и эндосимбиотического переноса генов» . Труды Национальной академии наук Соединенных Штатов Америки . 103 (27): 10328–33. Бибкод : 2006PNAS..10310328L . дои : 10.1073/pnas.0604232103 . ПМК 1502457 . ПМИД 16798872 .
- ^ Jump up to: Перейти обратно: а б Хазелтин, Калифорния, Ковальчиковски СК (май 2009 г.). «Архейный белок Rad54 реконструирует ДНК и стимулирует обмен цепей ДНК с помощью RadA» . Исследования нуклеиновых кислот . 37 (8): 2757–70. дои : 10.1093/нар/gkp068 . ПМЦ 2677860 . ПМИД 19282450 .
- ^ Рольфсмайер М.Л., Хазелтин, Калифорния (март 2010 г.). «Одноцепочечный ДНК-связывающий белок Sulfolobus solfataricus действует на пресинаптической стадии гомологичной рекомбинации». Журнал молекулярной биологии . 397 (1): 31–45. дои : 10.1016/j.jmb.2010.01.004 . ПМИД 20080104 .
- ^ Хуан Ц, Лю Л, Лю Дж, Ни Дж, Ше Ц, Шен Ю (2015). «Эффективная резекция концов 5'-3' ДНК с помощью HerA и NurA необходима для жизнеспособности клеток кренархеона Sulfolobus Islandicus» . BMC Молекулярная биология . 16 :2. дои : 10.1186/s12867-015-0030-z . ПМЦ 4351679 . ПМИД 25880130 .
- ^ Джайн С.К., Кокс М.М., Инман Р.Б. (август 1994 г.). «О роли гидролиза АТФ в обмене цепей ДНК, опосредованном белком RecA. III. Однонаправленная миграция ветвей и обширное образование гибридной ДНК» . Журнал биологической химии . 269 (32): 20653–61. дои : 10.1016/S0021-9258(17)32043-4 . ПМИД 8051165 .
- ^ Рамеш М.А., Малик С.Б., Логсдон Дж.М. (январь 2005 г.). «Филогеномный перечень мейотических генов; доказательства пола у лямблий и раннего эукариотического происхождения мейоза» . Современная биология . 15 (2): 185–91. дои : 10.1016/j.cub.2005.01.003 . ПМИД 15668177 . S2CID 17013247 .
- ^ Jump up to: Перейти обратно: а б Малик С.Б., Рамеш М.А., Халстранд А.М., Логсдон Дж.М. (декабрь 2007 г.). «Гомологи протистов мейотического гена Spo11 и топоизомеразы VI демонстрируют эволюционную историю дупликации генов и специфичных для линии потерь» . Молекулярная биология и эволюция . 24 (12): 2827–41. дои : 10.1093/molbev/msm217 . ПМИД 17921483 .
- ^ Лодиш Х., Берк А., Зипурски С.Л., Мацудайра П., Балтимор Д., Дарнелл Дж. (2000). «Глава 8.5: Замена генов и трансгенные животные: ДНК переносится в эукариотические клетки различными способами». Молекулярно-клеточная биология (4-е изд.). WH Фриман и компания. ISBN 978-0-7167-3136-8 .
- ^ «Нобелевская премия по физиологии и медицине 2007 года» . Нобелевский фонд . Проверено 15 декабря 2008 г.
- ^ Драммонд Д.А., Силберг Дж.Дж., Мейер М.М., Уилке К.О., Арнольд Ф.Х. (апрель 2005 г.). «О консервативной природе внутригенной рекомбинации» . Труды Национальной академии наук Соединенных Штатов Америки . 102 (15): 5380–5. Бибкод : 2005PNAS..102.5380D . дои : 10.1073/pnas.0500729102 . ПМК 556249 . ПМИД 15809422 .
- ^ Блум Дж.Д., Силберг Дж.Дж., Уилке К.О., Драммонд Д.А., Адами С., Арнольд Ф.Х. (январь 2005 г.). «Термодинамическое предсказание нейтральности белка» . Труды Национальной академии наук Соединенных Штатов Америки . 102 (3): 606–11. arXiv : q-bio/0409013 . Бибкод : 2005PNAS..102..606B . дои : 10.1073/pnas.0406744102 . ПМК 545518 . ПМИД 15644440 .
- ^ Jump up to: Перейти обратно: а б Карбоне М.Н., Арнольд Ф.Х. (август 2007 г.). «Инженерия путем гомологичной рекомбинации: исследование последовательности и функции внутри консервативной складки». Современное мнение в области структурной биологии . 17 (4): 454–9. дои : 10.1016/j.sbi.2007.08.005 . ПМИД 17884462 .
- ^ Оти Ч.Р., Ландвер М., Эндельман Дж.Б., Хирага К., Блум Дж.Д., Арнольд Ф.Х. (май 2006 г.). «Структурно-ориентированная рекомбинация создает искусственное семейство цитохромов P450» . ПЛОС Биология . 4 (5): е112. дои : 10.1371/journal.pbio.0040112 . ПМЦ 1431580 . ПМИД 16594730 .
- ^ Соколич М., Локлесс С.В., Расс В.П., Ли Х., Гарднер К.Х., Ранганатан Р. (сентябрь 2005 г.). «Эволюционная информация для определения складки белка». Природа . 437 (7058): 512–8. Бибкод : 2005Natur.437..512S . дои : 10.1038/nature03991 . ПМИД 16177782 . S2CID 4363255 .
- ^ Туласирам Х.В., Эриксон Х.К., Поултер К.Д. (апрель 2007 г.). «Химеры двух изопреноидсинтаз катализируют все четыре реакции сочетания в биосинтезе изопреноидов». Наука . 316 (5821): 73–6. Бибкод : 2007Sci...316...73T . дои : 10.1126/science.1137786 . ПМИД 17412950 . S2CID 43516273 .
- ^ Ландвер М., Карбон М., Оти Ч.Р., Ли Ю., Арнольд Ф.Х. (март 2007 г.). «Диверсификация каталитической функции синтетического семейства химерных цитохромов p450» . Химия и биология . 14 (3): 269–78. doi : 10.1016/j.chembiol.2007.01.009 . ЧВК 1991292 . ПМИД 17379142 .
- ^ Чоудхури, Ананья; Чжао, Хелен; Джалали, Фарид; Рашид, Шахназ А.Л.; Ран, Джейн; Супиот, Стефан; Килти, Энн Э.; Бристоу, Роберт Г. (1 января 2009 г.). «Нацеливание на гомологичную рекомбинацию с помощью иматиниба приводит к повышению химиочувствительности и радиочувствительности опухолевых клеток» (PDF) . Молекулярная терапия рака . 8 (1): 203–213. doi : 10.1158/1535-7163.MCT-08-0959 . ISSN 1535-7163 . ПМИД 19139130 . S2CID 6493434 .
- ^ Сиддики, Арафат; Тумиати, Мануэла; Либо, Алия; Сандхольм, Джоко; Реринг, Пиа; Аакко, София; Вайнионпяя, Реетта; Кайпио, Катя; Хухтинен, Кайса; Кауппи, Лийза; Туомела, Йоханна; Хиетанен, Сакари (2021). «Нацеливание на способность гомологичной репарации ДНК при одновременном ингибировании топоизомеразы II и c-Abl» . Границы онкологии . 11 : 3666. doi : 10.3389/fonc.2021.733700 . ISSN 2234-943X . ПМЦ 8488401 . ПМИД 34616682 .
- ^ Jump up to: Перейти обратно: а б с Иглхарт JD, Silver DP (июль 2009 г.). «Синтетическая летальность - новое направление в разработке лекарств от рака». Медицинский журнал Новой Англии . 361 (2): 189–91. дои : 10.1056/NEJMe0903044 . ПМИД 19553640 .
- ^ Фонг ПК, Босс Д.С., Яп Т.А., Тутт А., Ву П., Мерги-Рулвинк М., Мортимер П., Суэйсленд Х., Лау А., О'Коннор М.Дж., Эшворт А., Кармайкл Дж., Кэй С.Б., Шелленс Дж.Х., де Боно Дж.С. ( июль 2009 г.). «Ингибирование поли(АДФ-рибозы)-полимеразы в опухолях от носителей мутации BRCA» . Медицинский журнал Новой Англии . 361 (2): 123–34. doi : 10.1056/NEJMoa0900212 . ПМИД 19553641 .
- ^ Эдвардс С.Л., Бро Р., Лорд С.Дж., Натраян Р., Ватчева Р., Левин Д.А., Бойд Дж., Рейс-Фильо Дж.С., Эшворт А. (февраль 2008 г.). «Сопротивление терапии, вызванное внутригенной делецией BRCA2». Природа . 451 (7182): 1111–5. Бибкод : 2008Natur.451.1111E . дои : 10.1038/nature06548 . ПМИД 18264088 . S2CID 205212044 .
Внешние ссылки [ править ]
- Анимации – гомологичная рекомбинация : Анимации, показывающие несколько моделей гомологичной рекомбинации.
- Гомологичная рекомбинация: Tempy & Trun : анимация бактериального пути гомологичной рекомбинации RecBCD.