Метилирование гистонов
Метилирование гистонов — это процесс, при котором метильные группы передаются на аминокислоты белков - гистонов , составляющих нуклеосомы , которые двойная спираль ДНК обвивает с образованием хромосом . Метилирование гистонов может как увеличивать, так и уменьшать транскрипцию генов, в зависимости от того, какие аминокислоты в гистонах метилированы и сколько метильных групп присоединено. События метилирования, которые ослабляют химическое притяжение между хвостами гистонов и ДНК, увеличивают транскрипцию, поскольку они позволяют ДНК раскручиваться от нуклеосом, чтобы белки транскрипционных факторов и РНК-полимераза могли получить доступ к ДНК. Этот процесс имеет решающее значение для регуляции экспрессии генов , что позволяет различным клеткам экспрессировать разные гены.
Функция
[ редактировать ]Метилирование гистонов как механизм модификации структуры хроматина связано со стимуляцией нервных путей, которые, как известно, важны для формирования долговременной памяти и обучения. [1] Метилирование гистонов имеет решающее значение почти для всех фаз эмбрионального развития животных . [2]
Модели на животных показали, что метилирование и другие механизмы эпигенетической регуляции связаны со старением, нейродегенеративными заболеваниями и умственной отсталостью. [1] ( синдром Рубинштейна-Тайби , Х-сцепленная умственная отсталость ). [3] Неправильная регуляция H3K4, H3K27 и H4K20 связана с раком . [4] Эта модификация изменяет свойства нуклеосомы и влияет на ее взаимодействие с другими белками, особенно в отношении процессов транскрипции генов.
- Метилирование гистонов может быть связано либо с репрессией транскрипции , либо с активацией . Например, триметилирование гистона H3 по лизину 4 ( H3K4me3 ) является активным маркером транскрипции и активируется в гиппокампе через час после контекстуального формирования страха у крыс. Однако диметилирование гистона H3 по лизину 9 ( H3K9me2 ), сигнал для подавления транскрипции, усиливается после воздействия либо воздействия страха, либо только новой среды. [5]
- Метилирование некоторых остатков лизина (K) и аргинина (R) гистонов приводит к активации транскрипции. Примеры включают метилирование лизина 4 гистона 3 ( H3K4me1 ) и остатков аргинина (R) на H3 и H4 .
- Добавление метильных групп к гистонам с помощью гистон-метилтрансфераз может либо активировать, либо дополнительно подавлять транскрипцию, в зависимости от метилируемой аминокислоты и присутствия поблизости других метильных или ацетильных групп. [6]
- Чаще всего при метилировании лизина гистонов метилтрансферазы гистонов (которые метилируют) характеризуются как «писатели», тогда как деметилазы характеризуются как «ластики». [2] [4]
Механизм
[ редактировать ]Фундаментальная единица хроматина , называемая нуклеосомой , содержит ДНК, намотанную на белковый октамер . Этот октамер состоит из двух копий каждого из четырех белков-гистонов: H2A , H2B , H3 и H4 . Каждый из этих белков имеет удлиненный хвост, и эти хвосты являются мишенями модификации нуклеосом путем метилирования. Активация или инактивация ДНК во многом зависит от конкретного метилированного хвостового остатка и степени его метилирования. Гистоны могут быть метилированы только по остаткам лизина (K) и аргинина (R), но чаще всего метилирование наблюдается по остаткам лизина хвостов гистонов H3 и H4. [7] Хвостовой конец, наиболее удаленный от ядра нуклеосомы, является N-концевым (остатки нумеруются, начиная с этого конца). Общие сайты метилирования, связанные с активацией генов, включают H3K4, H3K48 и H3K79. Общие сайты инактивации генов включают H3K9 и H3K27. [8] Исследования этих сайтов показали, что метилирование хвостов гистонов по различным остаткам служит маркерами для рекрутирования различных белков или белковых комплексов, которые служат для регуляции активации или инактивации хроматина.
Остатки лизина и аргинина содержат аминогруппы, которые придают основные и гидрофобные характеристики. Лизин может быть моно-, ди- или триметилирован с метильной группой, замещающей каждый водород в его группе NH3+. Имея свободные группы NH2 и NH2+, аргинин может быть моно- или диметилирован. Это диметилирование может происходить асимметрично по группе NH2 или симметрично с одним метилированием по каждой группе. [9] Каждое добавление метильной группы к каждому остатку требует определенного набора белковых ферментов с различными субстратами и кофакторами. Как правило, для метилирования остатка аргинина требуется комплекс, включающий белок аргининметилтрансферазу (PRMT), тогда как для лизина требуется специфическая гистоновая метилтрансфераза (HMT), обычно содержащая эволюционно консервативный домен SET. [10]
Разная степень метилирования остатка может придавать разные функции, как показано на примере метилирования обычно изучаемого остатка H4K20. Монометилированный H4K20 ( H4K20me 1) участвует в уплотнении хроматина и, следовательно, в репрессии транскрипции. Однако H4K20me2 жизненно важен для восстановления поврежденной ДНК. При диметилировании этот остаток обеспечивает платформу для связывания белка 53BP1, участвующего в восстановлении двухцепочечных разрывов ДНК путем негомологичного соединения концов. Наблюдается концентрация H4K20me3 в гетерохроматине, и снижение этого триметилирования наблюдается при прогрессировании рака. Следовательно, H4K20me3 выполняет дополнительную роль в репрессии хроматина. [10] Восстановление двухцепочечных разрывов ДНК в хроматине также происходит путем гомологичной рекомбинации и также включает метилирование гистонов ( H3K9me3 ), чтобы облегчить доступ ферментов репарации к местам повреждения. [11]
Гистон метилтрансфераза
[ редактировать ]Геном плотно конденсирован в хроматин, который необходимо ослабить, чтобы транскрипция произошла . Чтобы остановить транскрипцию гена, ДНК должна быть намотана туже. Это можно сделать путем модификации гистонов в определенных сайтах путем метилирования. Гистоновые метилтрансферазы представляют собой ферменты, которые переносят метильные группы от S-аденозилметионина (SAM) на остатки лизина или аргинина гистонов H3 и H4. Есть случаи, когда основные глобулярные домены гистонов также метилируются.
Гистоновые метилтрансферазы специфичны либо к лизину, либо к аргинину. Лизин-специфичные трансферазы далее подразделяются на наличие у них домена SET или домена без SET. Эти домены точно определяют, как фермент катализирует перенос метила от SAM к белку-переносчику и далее к остатку гистона. [12] Метилтрансферазы могут добавлять 1-3 метила к целевым остаткам.
Эти метилы, добавляемые к гистонам, регулируют транскрипцию, блокируя или поощряя доступ ДНК к факторам транскрипции. Таким образом, целостность генома и эпигенетическое наследование генов находятся под контролем действия гистоновых метилтрансфераз. Метилирование гистонов играет ключевую роль в различении целостности генома и генов, экспрессируемых клетками, что придает клеткам их идентичность.
Метилированные гистоны могут как подавлять, так и активировать транскрипцию. [12] Например, H3K4me2 , H3K4me3 и H3K79me3 обычно связаны с транскрипционной активностью, тогда как H3K9me2 , H3K9me3 , H3K27me2 , H3K27me3 и H4K20me3 связаны с репрессией транскрипции. [13]
Эпигенетика
[ редактировать ]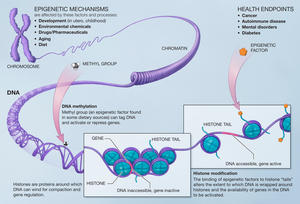
Модификации, внесенные в гистон, влияют на гены, которые экспрессируются в клетке, и это тот случай, когда метилтрансферазы добавляют к остаткам гистонов метилы. [14] Метилирование гистонов играет важную роль в сборке гетерохроматинового механизма и поддержании границ между генами, которые транскрибируются, и генами, которые не транскрибируются. Эти изменения передаются потомству и могут зависеть от окружающей среды, которой подвергаются клетки. Эпигенетические изменения обратимы, что означает, что они могут быть мишенью для терапии.
Активность гистон-метилтрансфераз компенсируется активностью гистон-деметилаз. Это позволяет включать или выключать транскрипцию путем отмены ранее существовавших модификаций. Необходимо жестко регулировать активность как гистонметилтрансфераз, так и гистондеметилаз. Неправильная регуляция любого из них может привести к экспрессии генов, что приводит к повышенной восприимчивости к болезням. Многие виды рака возникают из-за неадекватных эпигенетических эффектов неправильной регуляции метилирования. [15] Однако, поскольку эти процессы иногда обратимы, существует интерес к использованию их активности в сочетании с противораковой терапией. [15]
При инактивации Х-хромосомы
[ редактировать ]В женских организмах сперматозоид, содержащий Х-хромосому, оплодотворяет яйцеклетку, давая эмбриону две копии Х-хромосомы. Однако женщинам изначально не требуются обе копии Х-хромосомы, поскольку это только удвоит количество транскрибируемых белковых продуктов, как показывает гипотеза дозовой компенсации. Отцовская Х-хромосома быстро инактивируется в течение первых нескольких делений. [16] Эта неактивная Х-хромосома (Xi) упакована в невероятно плотную форму хроматина, называемую гетерохроматином . [17] Эта упаковка происходит из-за метилирования различных остатков лизина, которые помогают образовывать разные гистоны. У человека инактивация X представляет собой случайный процесс, опосредованный некодирующей РНК XIST. [18]
Хотя метилирование остатков лизина происходит на многих различных гистонах, наиболее характерным для Xi является девятый лизин третьего гистона (H3K9). Хотя однократное метилирование этой области позволяет связанным генам оставаться транскрипционно активными, [19] в гетерохроматине этот остаток лизина часто метилируется дважды или трижды, H3K9me2 или H3K9me3 соответственно, чтобы гарантировать неактивность связанной с ДНК. Более поздние исследования показали, что H3K27me3 и H4K20me1 также часто встречаются у ранних эмбрионов. Другие метки метилирования, связанные с транскрипционно активными областями ДНК, H3K4me2 и H3K4me3, отсутствуют в Xi-хромосоме вместе со многими метками ацетилирования. Хотя было известно, что определенные метки метилирования гистонов Xi оставались относительно постоянными между видами, недавно было обнаружено, что разные организмы и даже разные клетки внутри одного организма могут иметь разные метки для инактивации Xi. [20] Посредством метилирования гистонов происходит генетический импринтинг , так что один и тот же гомолог X остается инактивированным в ходе репликации хромосом и деления клеток.
Мутации
[ редактировать ]В связи с тем, что метилирование гистонов регулирует большую часть транскрибируемых генов, даже небольшие изменения в характере метилирования могут иметь тяжелые последствия для организма. Мутации, которые происходят, чтобы увеличить или уменьшить метилирование, имеют большие изменения в регуляции генов, в то время как мутации таких ферментов, как метилтрансфераза и деметилтрансфераза, могут полностью изменить то, какие белки транскрибируются в данной клетке. Чрезмерное метилирование хромосомы может привести к инактивации определенных генов, необходимых для нормального функционирования клеток. В определенном штамме дрожжей Saccharomyces cerevisiae мутация, вызывающая метилирование трех остатков лизина в третьем гистоне, H3K4, H3K36 и H3K79, вызывает задержку митотического клеточного цикла, поскольку многие гены, необходимые для этого прогрессирования, инактивируются. Эта крайняя мутация приводит к гибели организма. Было обнаружено, что удаление генов, которые в конечном итоге позволят производить гистон-метилтрансферазу, позволяет этому организму жить, поскольку его остатки лизина не метилируются. [21]
В последние годы внимание исследователей привлекло то, что многие виды рака вызываются в основном эпигенетическими факторами. Рак может быть вызван различными способами из-за дифференциального метилирования гистонов. С момента открытия онкогенов , а также генов-супрессоров опухолей стало известно, что главный фактор, вызывающий и подавляющий рак, находится внутри нашего собственного генома. Если области вокруг онкогенов станут неметилированными, эти гены, вызывающие рак, могут транскрибироваться с угрожающей скоростью. Противоположностью этому является метилирование генов-супрессоров опухолей. В тех случаях, когда области вокруг этих генов были сильно метилированы, ген-супрессор опухоли не был активен и, следовательно, вероятность возникновения рака была выше. Эти изменения в характере метилирования часто происходят из-за мутаций метилтрансферазы и деметилтрансферазы. [22] Другие типы мутаций в белках, таких как изоцитратдегидрогеназа 1 (IDH1) и изоцитратдегидрогеназа 2 (IDH2), могут вызывать инактивацию гистондеметилтрансферазы, что, в свою очередь, может привести к различным видам рака, глиомам и лейкозам, в зависимости от того, в каких клетках произошла мутация. происходит. [23]
Одноуглеродный метаболизм изменяет метилирование гистонов
[ редактировать ]При одноуглеродном метаболизме аминокислоты глицин и серин превращаются посредством фолатного и метионинового циклов в предшественники нуклеотидов и SAM. Многочисленные питательные вещества поддерживают одноуглеродный метаболизм, включая глюкозу , серин, глицин и треонин . Высокие уровни SAM-донора метила влияют на метилирование гистонов, что может объяснить, как высокие уровни SAM предотвращают злокачественную трансформацию. [24]
См. также
[ редактировать ]- Гистоновый код
- Ацетилирование и деацетилирование гистонов
- Гистон метилтрансфераза
- Метилирование
- Метиллизин
- Генетический импринтинг
- Метилирование ДНК
Ссылки
[ редактировать ]- ^ Jump up to: а б Крамер, Джейми М. (февраль 2013 г.). «Эпигенетическая регуляция памяти: последствия когнитивных расстройств человека» . Биомолекулярные концепции . 4 (1): 1–12. дои : 10.1515/bmc-2012-0026 . hdl : 2066/118379 . ПМИД 25436561 . S2CID 6195513 .
- ^ Jump up to: а б Джамбхекар А., Дхалл А., Ши Ю (2019). «Роль и регуляция метилирования гистонов в развитии животных» . Nature Reviews Молекулярно-клеточная биология . 20 (10): 625–641. дои : 10.1038/s41580-019-0151-1 . ПМК 6774358 . ПМИД 31267065 .
- ^ Лилья, Тобиас; Нина Хелдринг; Ола Хермансон (февраль 2013 г.). «Как вращающийся гистон: эпигенетическая регуляция нервных стволовых клеток и развитие мозга с помощью факторов, контролирующих ацетилирование и метилирование гистонов» . Biochimica et Biophysical Acta (BBA) – Общие предметы . 1830 (2): 2354–2360. дои : 10.1016/j.bbagen.2012.08.011 . ПМИД 22986149 .
- ^ Jump up to: а б Хён К., Чон Дж., Пак К., Ким Дж. (2017). «Запись, стирание и чтение метилирования лизина гистонов» . Экспериментальная и молекулярная медицина . 49 (4): е324. дои : 10.1038/emm.2017.11 . ПМК 6130214 . ПМИД 28450737 .
- ^ Гупта, Свати; Се Ю. Ким; Соня Артис; Дэвид Л. Молфезе; Армин Шумахер; Дж. Дэвид Суэтт; Ричард Э. Пэйлор; Фара Д. Любин (10 марта 2010 г.). «Метилирование гистонов регулирует формирование памяти» . Журнал неврологии . 30 (10): 3589–3599. doi : 10.1523/JNEUROSCI.3732-09.2010 . ПМЦ 2859898 . ПМИД 20219993 .
- ^ Грир, Эрик Л.; Ши, Ян (2012). «Метилирование гистонов: динамический показатель здоровья, болезней и наследственности» . Обзоры природы Генетика . 13 (5): 343–57. дои : 10.1038/nrg3173 . ПМЦ 4073795 . ПМИД 22473383 .
- ^ Цзя, Сонгтао; Ван, Ю (2009). «Градусы имеют решающее значение» (PDF) . Эпигенетика . 4 (5): 1–4.
- ^ Гилберт, Сан-Франциско (2010). Биология развития. (9-е изд.). Sinauer Associates, Inc. 35-37.
- ^ Блан, Ромео С; Ричард, Стефан (5 января 2017 г.). «Метилирование аргинина: достижение совершеннолетия» . Молекулярная клетка . 65 (1): 8–24. дои : 10.1016/j.molcel.2016.11.003 . ПМИД 28061334 .
- ^ Jump up to: а б Рейнберг, Дэнни; Чжан, Йи (2001). «Регуляция транскрипции посредством метилирования гистонов: взаимодействие между различными ковалентными модификациями основных хвостов гистонов» . Гены и развитие . 15 (18): 2343–2360. дои : 10.1101/gad.927301 . ПМИД 11562345 .
- ^ Вэй С., Ли С., Инь Цз., Вэнь Дж., Мэн Х., Сюэ Л., Ван Дж. (2018). «Метилирование гистонов в репарации ДНК и клиническая практика: новые открытия за последние 5 лет» . Джей Рак . 9 (12): 2072–2081. дои : 10.7150/jca.23427 . ПМК 6010677 . ПМИД 29937925 .
- ^ Jump up to: а б Райс, Джадд К.; Бриггс, Скотт Д.; Юберхайде, Беатрикс; Барбер, Синтия М.; Шабановиц, Джеффри; Хант, Дональд Ф.; Синкай, Ёичи; Эллис, К.Дэвид (2003). «Гистоновые метилтрансферазы управляют различными степенями метилирования для определения различных доменов хроматина» . Молекулярная клетка . 12 (6): 1591–1598. дои : 10.1016/S1097-2765(03)00479-9 . ПМИД 14690610 .
- ^ Куистра, Сюзанна Марие; Хелин, Кристиан (4 апреля 2012 г.). «Молекулярные механизмы и потенциальные функции деметилаз гистонов» . Nature Reviews Молекулярно-клеточная биология . 13 (5): 297–311. дои : 10.1038/nrm3327 . ISSN 1471-0072 . ПМИД 22473470 . S2CID 26181430 .
- ^ Чунг, Питер; Лау, Присцилла (2005). «Эпигенетическая регуляция посредством метилирования гистонов и вариантов гистонов» . Молекулярная эндокринология . 19 (3): 563–73. дои : 10.1210/me.2004-0496 . ПМИД 15677708 .
- ^ Jump up to: а б Альберт, Марейке; Хелин, Кристиан (2010). «Гистон-метилтрансферазы при раке» . Семинары по клеточной биологии и биологии развития . 21 (2): 209–220. дои : 10.1016/j.semcdb.2009.10.007 . ПМИД 19892027 .
- ^ Акаги, Нобуо; Сасаки, Мотомичи (1975). «Преимущественная инактивация Х-хромосомы отцовского происхождения во внеэмбриональных мембранах мыши» . Природа . 256 (5519): 640–42. Бибкод : 1975Natur.256..640T . дои : 10.1038/256640a0 . ПМИД 1152998 . S2CID 4190616 .
- ^ Элгин, Сара CR; Гревал, Шив И.С. (2003). «Гетерохроматин: молчание — золото» . Современная биология . 13 (23): 895–898 рэндов. дои : 10.1016/j.cub.2003.11.006 . ПМИД 14654010 . S2CID 9728548 .
- ^ Ан, Дж.; Ли, Дж. (2008). «Х-хромосома: инактивация Х». Природное образование . 1 (1): 24.
- ^ Барский, А.; Каддапа, С.; Цюи, К.; Ро, Т.; Шонес, Д.; Ван, З.; Вэй, Г.; Чепелев И.; Чжао, К. (2007). «Профилирование метилирования гистонов в геноме человека с высоким разрешением» . Клетка . 129 (4): 823–37. дои : 10.1016/j.cell.2007.05.009 . ПМИД 17512414 . S2CID 6326093 .
- ^ Чоу, Дженнифер; Херд, Эдит (2009). «Инактивация X и сложности подавления половой хромосомы» . Современное мнение в области клеточной биологии . 21 (3): 359–66. дои : 10.1016/j.ceb.2009.04.012 . ПМИД 19477626 .
- ^ Джин, Ю.; Родригес, AM; Стэнтон, доктор медицинских наук; Китазоно, А.А.; Вайрик, Джей-Джей (2007). «Одновременная мутация остатков метилированного лизина в гистоне H3 вызывает усиленное молчание генов, дефекты клеточного цикла и гибель клеток у Saccharomyces Cerevisiae» . Молекулярная и клеточная биология . 27 (19): 6832–841. дои : 10.1128/mcb.00745-07 . ПМК 2099221 . ПМИД 17664279 .
- ^ Эстеллер, М (2006). «Эпигенетика открывает новое поколение онкогенов и генов-супрессоров опухолей» . Британский журнал рака . 94 (2): 179–83. дои : 10.1038/sj.bjc.6602918 . ПМК 2361113 . ПМИД 16404435 .
- ^ Лу, Чао; Уорд, Патрик С.; Капур, Гурприт С.; Роул, Дэн; Цуркан, Севин; Абдель-Вахаб, Омар; Эдвардс, Кристофер Р.; Ханин, Рая; Фигероа, Мэри Э.; Мельник, король; Веллен, Кэтрин Э.; О'Рурк, Дональд М.; Бергер, Шелли Л.; Чан, Тимоти А.; Левин, Росс Л.; Меллингхофф, Инго К.; Томпсон, Крейг Б. (2012). «Мутация DH ухудшает деметилирование гистонов и приводит к блокированию дифференцировки клеток» . Природа 483 (7390): 474–78. Бибкод : 2012Nature.483..474L . дои : 10.1038/nature10860 . ПМЦ 3478770 . ПМИД 22343901 .
- ^ Ван Ю.П., Лей Цюй (2018). «Метаболическая перекодировка эпигенетики рака» . Раковая коммуна (Лондон) . 38 (1): 1–8. дои : 10.1186/s40880-018-0302-3 . ПМЦ 5993135 . ПМИД 29784032 .
Дальнейшее чтение
[ редактировать ]- Оруджи, Элиас и Утикал, Йохен. (2018). Эпигенетическая борьба со злокачественной меланомой: метилирование лизина гистонов. Клиническая эпигенетика 2018 10:145 https://clinicalepigeneticsjournal.biomedcentral.com/articles/10.1186/s13148-018-0583-z
- Гозани О. и Ши Ю. (2014). Метилирование гистонов в передаче сигналов хроматина. В: Основы хроматина (стр. 213–256). Спрингер Нью-Йорк. дои : 10.1007/978-1-4614-8624-4_5
- Ши, Ян; Уэтстайн, Джонатан Р. (12 января 2007 г.). «Динамическая регуляция метилирования лизина гистонов деметилазами» . Молекулярная клетка . 25 (1): 1–14. doi : 10.1016/j.molcel.2006.12.010 . ПМИД 17218267 .