хромосома

| Часть серии о |
| Генетика |
|---|
 |

- хроматида
- Центромера
- Короткая рука
- Длинная рука
Хромосома материал — это пакет ДНК , весь генетический организма содержащий часть или . В большинстве хромосом очень длинные тонкие волокна ДНК покрыты нуклеосомы упаковочными белками, образующими ; в эукариотических клетках наиболее важными из этих белков являются гистоны . Эти белки при помощи белков-шаперонов связываются с молекулой ДНК и конденсируют ее , поддерживая ее целостность. [1] [2] Эти хромосомы имеют сложную трехмерную структуру , которая играет значительную роль в регуляции транскрипции . [3]
Хромосомы обычно видны под световым микроскопом только во время метафазы деления клетки (когда все хромосомы в конденсированной форме располагаются в центре клетки). [4] Прежде чем это произойдет, каждая хромосома дублируется ( S-фаза ), и обе копии соединяются центромерой , в результате чего образуется либо Х-образная структура (на фото выше), если центромера расположена экваториально, либо двуплечая структура, если центромера расположена дистально. Объединенные копии теперь называются сестринскими хроматидами . Во время метафазы Х-образная структура называется метафазной хромосомой, которая сильно конденсируется и поэтому ее легче всего различить и изучить. [5] В клетках животных хромосомы достигают наивысшего уровня уплотнения в анафазе во время сегрегации хромосом . [6]
Хромосомная рекомбинация во время мейоза и последующего полового размножения играет значительную роль в генетическом разнообразии . Если этими структурами манипулировать неправильно, посредством процессов, известных как хромосомная нестабильность и транслокация, клетка может подвергнуться митотической катастрофе . Обычно это приводит к инициированию апоптоза клетки , что приводит к ее собственной гибели, но иногда мутации в клетке препятствуют этому процессу и, таким образом, вызывают прогрессирование рака .
Некоторые используют термин «хромосома» в более широком смысле, чтобы обозначить отдельные части хроматина в клетках, видимые или невидимые под световой микроскопией. Другие используют эту концепцию в более узком смысле, обозначая отдельные части хроматина во время деления клеток, видимые под световой микроскопией из-за высокой конденсации.
Этимология [ править ]
Слово хромосома ( / ˈ k r oʊ m ə ˌ s oʊ m , - ˌ z oʊ m / [7] [8] ) происходит от греческого χρῶμα ( цветность , «цвет») и σῶμα ( сома , «тело»), описывающего их сильное окрашивание определенными красителями . [9] Термин был введен немецким анатомом Генрихом Вильгельмом Вальдейером . [10] Имея в виду термин хроматин , который был введен Вальтером Флеммингом .
Некоторые из ранних кариологических терминов устарели. [11] [12] Например, хроматин (Flemming 1880) и хромосома (Waldeyer 1888) приписывают цвет неокрашенному состоянию. [13]
История открытия [ править ]


Отто Бючли был первым ученым, распознавшим структуры, ныне известные как хромосомы. [14]
В серии экспериментов, начавшихся в середине 1880-х годов, Теодор Бовери внес решающий вклад в объяснение того, что хромосомы являются векторами наследственности, с помощью двух понятий, которые стали известны как «непрерывность хромосом» и «хромосомная индивидуальность». [15]
Вильгельм Ру предположил, что каждая хромосома несет различную генетическую конфигурацию , и Бовери смог проверить и подтвердить эту гипотезу. Благодаря повторному открытию в начале 1900-х годов более ранних работ Грегора Менделя Бовери смог указать на связь между правилами наследственности и поведением хромосом. Бовери оказал влияние на два поколения американских цитологов: Эдмунд Бичер Уилсон , Нетти Стивенс , Уолтер Саттон и Теофил Пейнтер находились под влиянием Бовери (Уилсон, Стивенс и Пейнтер фактически работали с ним). [16]
В своем знаменитом учебнике « Клетка в развитии и наследственности » Уилсон связал воедино независимые работы Бовери и Саттона (оба около 1902 года), назвав хромосомную теорию наследования хромосомной теорией Бовери-Саттона (названия иногда меняются местами). [17] Эрнст Майр отмечает, что эта теория горячо оспаривалась некоторыми известными генетиками: Уильямом Бейтсоном , Вильгельмом Йоханнсеном , Ричардом Гольдшмидтом и Т.Х. Морганом , все они придерживались довольно догматического склада ума. В конце концов, полное доказательство было получено на основе карт хромосом в собственной лаборатории Моргана. [18]
Число хромосом человека было опубликовано в 1923 году Теофилом Пейнтером . Рассматривая их под микроскопом, он насчитал двадцать четыре пары, что означало сорок восемь хромосом. Его ошибку скопировали другие, и только в 1956 году истинное число — сорок шесть — было определено цитогенетиком индонезийского происхождения Джо Хин Тджио . [19]
Прокариоты [ править ]
Прокариоты . – бактерии и археи – обычно имеют одну кольцевую хромосому , но существует множество вариаций [20] Хромосомы большинства бактерий, которые некоторые авторы предпочитают называть генофорами , могут иметь размер всего от 130 000 пар оснований у эндосимбиотических бактерий Candidatus Hodgkinia cicadicola. [21] и кандидат в князи Тремблайи , [22] до более чем 14 000 000 пар оснований у почвенной бактерии Sorangium cellulosum . [23] Спирохеты рода Borrelia Borrelia являются заметным исключением из этого правила: такие бактерии, как burgdorferi , причина болезни Лайма , содержат одну линейную хромосому. [24]
Структура в последовательностях [ править ]
Хромосомы прокариот имеют менее основанную на последовательностях структуру, чем эукариоты. Бактерии обычно имеют одну точку ( начало репликации ), из которой начинается репликация, тогда как некоторые археи содержат несколько точек начала репликации. [25] Гены у прокариот часто организованы в опероны и обычно не содержат интронов , в отличие от эукариот.
ДНК Упаковка
Прокариоты не имеют ядра. Вместо этого их ДНК организована в структуру, называемую нуклеоидом . [26] [27] Нуклеоид представляет собой отдельную структуру и занимает определенную область бактериальной клетки. Однако эта структура динамична и поддерживается и ремоделируется под действием ряда гистоноподобных белков, которые связываются с бактериальной хромосомой. [28] У архей ДНК в хромосомах еще более организована: ДНК упакована в структуры, подобные эукариотическим нуклеосомам. [29] [30]
Некоторые бактерии также содержат плазмиды или другую внехромосомную ДНК . Это кольцевые структуры в цитоплазме , которые содержат клеточную ДНК и играют роль в горизонтальном переносе генов . [5] У прокариот (см. Нуклеоиды ) и вирусов [31] ДНК часто плотно упакована и организована; в случае архей — по гомологии с гистонами эукариот, а в случае бактерий — по гистоноподобным белкам.
Бактериальные хромосомы, как правило, прикреплены к плазматической мембране бактерий. В молекулярной биологии это позволяет выделить его из плазмидной ДНК путем центрифугирования лизированных бактерий и осаждения мембран (и прикрепленной ДНК).
Прокариотические хромосомы и плазмиды, как и эукариотическая ДНК, обычно имеют сверхспиральную структуру . Сначала ДНК должна быть переведена в расслабленное состояние для доступа к транскрипции , регуляции и репликации .
Эукариоты [ править ]
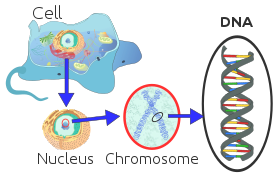
Каждая эукариотическая хромосома состоит из длинной линейной молекулы ДНК , связанной с белками , образующими компактный комплекс белков и ДНК, называемый хроматином . Хроматин содержит подавляющее большинство ДНК в организме, но небольшое количество, унаследованное по материнской линии, можно обнаружить в митохондриях . Он присутствует в большинстве клеток , за некоторыми исключениями, например, в эритроцитах .
Гистоны ответственны за первую и самую основную единицу организации хромосом — нуклеосому .
Эукариоты ( клетки с ядрами, подобными тем, которые встречаются у растений, грибов и животных) обладают множеством крупных линейных хромосом, содержащихся в ядре клетки. Каждая хромосома имеет одну центромеру с одним или двумя плечами, выступающими из центромеры, хотя в большинстве случаев эти плечи как таковые не видны. Кроме того, большинство эукариот имеют небольшой кольцевой митохондриальный геном , а некоторые эукариоты могут иметь дополнительные небольшие кольцевые или линейные цитоплазматические хромосомы.
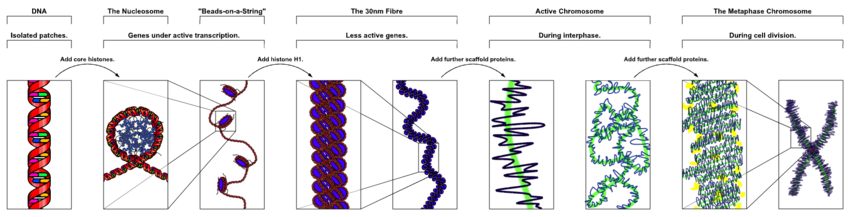
В ядерных хромосомах эукариот неконденсированная ДНК существует в полуупорядоченной структуре, где она обернута вокруг гистонов (структурных белков ), образуя композитный материал, называемый хроматином .
Интерфазный хроматин [ править ]
Упаковка ДНК в нуклеосомы приводит к образованию волокна диаметром 10 нанометров, которое может в дальнейшем конденсировать волокна размером до 30 нм. [32] Большая часть эухроматина в интерфазных ядрах представлена в виде волокон длиной 30 нм. [32] Структура хроматина представляет собой более деконденсированное состояние, т.е. конформация размером 10 нм обеспечивает транскрипцию. [32]
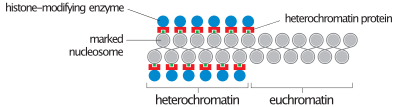
Во время интерфазы (периода клеточного цикла , когда клетка не делится) два типа хроматина можно выделить :
- Эухроматин , который состоит из активной ДНК, например, экспрессируемой в виде белка.
- Гетерохроматин , состоящий в основном из неактивной ДНК. Похоже, что он служит структурным целям на хромосомных стадиях. Гетерохроматин можно разделить на два типа:
- Конститутивный гетерохроматин , который никогда не экспрессируется. Он расположен вокруг центромеры и обычно содержит повторяющиеся последовательности .
- Факультативный гетерохроматин , который иногда выражен.
хроматин деление и Метафазный


На ранних стадиях митоза или мейоза (деление клетки) двойная спираль хроматина становится все более конденсированной. Они перестают функционировать как доступный генетический материал ( транскрипция прекращается) и становятся компактной транспортабельной формой. Считается, что петли тридцатинанометровых волокон хроматина складываются дальше, образуя компактные метафазные хромосомы митотических клеток. Таким образом, ДНК конденсируется примерно в десять тысяч раз. [32]
Хромосомный каркас , состоящий из таких белков, как конденсин , TOP2A и KIF4 , [33] играет важную роль в удержании хроматина в компактных хромосомах. Петли тридцатинанометровой структуры далее конденсируются с каркасом в структуры более высокого порядка. [34]
Эта очень компактная форма делает отдельные хромосомы видимыми, и они образуют классическую четырехплечевую структуру — пару сестринских хроматид, прикрепленных друг к другу в центромере . Более короткие ветви называются p-руками (от французского petit — маленький), а более длинные — q-руками ( q следует за p в латинском алфавите; qg «гранде»; в качестве альтернативы иногда говорят, что q — это сокращение от «очередь», означающее «хвост» в латинском алфавите). Французский [35] ). Это единственный природный контекст, в котором отдельные хромосомы можно увидеть в оптический микроскоп .
Митотические метафазные хромосомы лучше всего описываются линейно организованным, сжатым в продольном направлении массивом последовательных петель хроматина. [36]
Во время митоза микротрубочки растут из центросом, расположенных на противоположных концах клетки, а также прикрепляются к центромере в специализированных структурах, называемых кинетохорами , по одной из которых присутствует на каждой сестринской хроматиде . Особая последовательность оснований ДНК в области кинетохор обеспечивает, наряду со специальными белками, более длительное прикрепление в этом районе. Затем микротрубочки раздвигают хроматиды по направлению к центросомам, так что каждая дочерняя клетка наследует один набор хроматид. После деления клеток хроматиды раскручиваются, и ДНК снова может транскрибироваться. Несмотря на свой внешний вид, хромосомы структурно сильно конденсированы, что позволяет этим гигантским структурам ДНК удерживаться внутри ядра клетки.
Хромосомы человека [ править ]
Хромосомы у человека можно разделить на два типа: аутосомы (хромосома(ы) тела) и аллосомы ( половая хромосома (и)). Определенные генетические черты связаны с полом человека и передаются через половые хромосомы. Аутосомы содержат остальную часть генетической наследственной информации. Все действуют одинаково во время деления клеток. Клетки человека имеют 23 пары хромосом (22 пары аутосом и одна пара половых хромосом), всего 46 на клетку. Помимо этого, клетки человека имеют многие сотни копий митохондриального генома . Секвенирование генома человека предоставило много информации о каждой хромосоме. Ниже представлена таблица, в которой собрана статистика по хромосомам, основанная на Института Сэнгера информации о геноме человека в базе данных аннотаций генома позвоночных (VEGA) . [37] Число генов является приблизительным, поскольку оно частично основано на предсказаниях генов . Общая длина хромосом также является оценкой, основанной на предполагаемом размере несеквенированных областей гетерохроматина .
| хромосома | Гены [38] | Всего пар оснований | % оснований | Секвенированные пары оснований [39] | % секвенированных пар оснований |
|---|---|---|---|---|---|
| 1 | 2000 | 247,199,719 | 8.0 | 224,999,719 | 91.02% |
| 2 | 1300 | 242,751,149 | 7.9 | 237,712,649 | 97.92% |
| 3 | 1000 | 199,446,827 | 6.5 | 194,704,827 | 97.62% |
| 4 | 1000 | 191,263,063 | 6.2 | 187,297,063 | 97.93% |
| 5 | 900 | 180,837,866 | 5.9 | 177,702,766 | 98.27% |
| 6 | 1000 | 170,896,993 | 5.5 | 167,273,993 | 97.88% |
| 7 | 900 | 158,821,424 | 5.2 | 154,952,424 | 97.56% |
| 8 | 700 | 146,274,826 | 4.7 | 142,612,826 | 97.50% |
| 9 | 800 | 140,442,298 | 4.6 | 120,312,298 | 85.67% |
| 10 | 700 | 135,374,737 | 4.4 | 131,624,737 | 97.23% |
| 11 | 1300 | 134,452,384 | 4.4 | 131,130,853 | 97.53% |
| 12 | 1100 | 132,289,534 | 4.3 | 130,303,534 | 98.50% |
| 13 | 300 | 114,127,980 | 3.7 | 95,559,980 | 83.73% |
| 14 | 800 | 106,360,585 | 3.5 | 88,290,585 | 83.01% |
| 15 | 600 | 100,338,915 | 3.3 | 81,341,915 | 81.07% |
| 16 | 800 | 88,822,254 | 2.9 | 78,884,754 | 88.81% |
| 17 | 1200 | 78,654,742 | 2.6 | 77,800,220 | 98.91% |
| 18 | 200 | 76,117,153 | 2.5 | 74,656,155 | 98.08% |
| 19 | 1500 | 63,806,651 | 2.1 | 55,785,651 | 87.43% |
| 20 | 500 | 62,435,965 | 2.0 | 59,505,254 | 95.31% |
| 21 | 200 | 46,944,323 | 1.5 | 34,171,998 | 72.79% |
| 22 | 500 | 49,528,953 | 1.6 | 34,893,953 | 70.45% |
| Х (половая хромосома) | 800 | 154,913,754 | 5.0 | 151,058,754 | 97.51% |
| Y (половая хромосома) | 200 [40] | 57,741,652 | 1.9 | 25,121,652 | 43.51% |
| Общий | 21,000 | 3,079,843,747 | 100.0 | 2,857,698,560 | 92.79% |
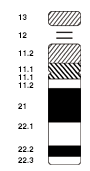
На основании микрографических характеристик размера, положения центромеры и иногда наличия хромосомного спутника хромосомы человека классифицируются на следующие группы: [41] [42]
| Группа | Хромосомы | Функции |
|---|---|---|
| А | 1–3 | Большие, метацентрические или субметацентрические |
| Б | 4–5 | Большой, субметацентрический |
| С | 6–12, Х | Среднего размера, субметацентрический |
| Д | 13–15 | Среднего размера, акроцентрическая, со сателлитом. |
| И | 16–18 | Маленькие, метацентрические или субметацентрические |
| Ф | 19–20 | Очень маленький, метацентрический |
| Г | 21–22 и | Очень маленький, акроцентрический (и 21, 22 со сателлитом ) |
Кариотип [ править ]


В общем, кариотип — это характерный хромосомный набор эукариот видов . [43] Получение и изучение кариотипов является частью цитогенетики .
Хотя репликация и транскрипция ДНК . высоко стандартизированы у эукариот , этого нельзя сказать об их кариотипах, которые часто сильно варьируют Между видами могут быть различия в количестве хромосом и в детальной организации.В некоторых случаях наблюдаются значительные различия внутри вида. Часто бывает:
- 1. различия между двумя полами
- 2. различия между зародышевой линией и сомой (между гаметами и остальным телом)
- 3. изменчивость между членами популяции из-за сбалансированного генетического полиморфизма.
- 4. географические различия между расами
- 5. мозаики или иные ненормальные особи.
Кроме того, изменения кариотипа могут возникать во время развития оплодотворенной яйцеклетки.
Методика определения кариотипа обычно называется кариотипированием . Клетки можно заблокировать на полпути деления (в метафазе) in vitro (в реакционном флаконе) с помощью колхицина . Затем эти клетки окрашивают, фотографируют и формируют кариограмму с расположенным набором хромосом, аутосомами в порядке длины и половыми хромосомами (здесь X/Y) в конце.
Как и у многих видов, размножающихся половым путем, у человека имеются особые гоносомы (половые хромосомы, в отличие от аутосом ). Это XX у женщин и XY у мужчин.
и методы анализа История
Исследования кариотипа человека заняли много лет, чтобы решить самый основной вопрос: сколько хромосом содержит нормальная диплоидная клетка человека? В 1912 году Ганс фон Винивартер сообщил о 47 хромосомах в сперматогониях и 48 в оогониях , сделав вывод о XX/XO механизме определения пола . [44] В 1922 году Пейнтер не был уверен, равно ли диплоидное число человека 46 или 48, сначала отдавая предпочтение 46. [45] Позже он пересмотрел свое мнение с 46 на 48 и правильно настаивал на том, что у людей есть система XX/XY . [46]
Для окончательного решения проблемы потребовались новые методы:
- Использование клеток в культуре
- Задержка митоза в метафазе раствором колхицина
- Предварительная обработка клеток гипотоническим раствором 0,075 М KCl, который набухает и распределяет хромосомы.
- Сдавливание препарата на предметном стекле, выравнивание хромосом в одной плоскости
- Разрезание микрофотографии и преобразование результата в бесспорную кариограмму.
Прошло до 1954 года, прежде чем диплоидное число человека было подтверждено как 46. [47] [48] Учитывая методы Винивартера и Пейнтера, их результаты были весьма замечательными. [49] Шимпанзе , ближайшие ныне живущие родственники современного человека, имеют 48 хромосом, как и другие человекообразные обезьяны : у человека две хромосомы сливаются, образуя хромосому 2 .
Аберрации [ править ]
Хромосомные аберрации – это нарушения нормального хромосомного состава клетки. Они могут вызывать генетические заболевания у людей, такие как синдром Дауна , [50] хотя большинство аберраций практически не имеют эффекта. , не вызывают заболеваний у носителей Некоторые хромосомные аномалии, такие как транслокации или хромосомные инверсии , хотя они могут привести к более высокой вероятности рождения ребенка с хромосомным нарушением. [ нужна ссылка ] Аномальное количество хромосом или наборов хромосом, называемое анеуплоидией , может быть смертельным или привести к генетическим нарушениям. [51] Генетическое консультирование предлагается семьям, в которых может быть перестройка хромосом.
Приобретение или потеря ДНК из хромосом может привести к множеству генетических нарушений . [52] Человеческие примеры включают в себя:
- Cri du Chat , вызванный делецией части короткого плеча хромосомы 5. «Cri du Chat» на французском языке означает «кошачий крик»; Состояние было названо так потому, что пострадавшие младенцы издают пронзительный крик, похожий на кошачий. Заболевшие люди имеют широко посаженные глаза, маленькую голову и челюсть, психические расстройства от умеренных до тяжелых и очень низкий рост.
- Синдром Дауна , наиболее распространенная трисомия, обычно вызываемая лишней копией хромосомы 21 ( трисомия 21 ). Характеристики включают снижение мышечного тонуса, более коренастое телосложение, асимметричный череп, раскосые глаза и отставание в развитии от легкой до умеренной степени. [53]
- Синдром Эдвардса , или трисомия-18, вторая по распространенности трисомия. [54] Симптомы включают задержку моторики, отставание в развитии и многочисленные врожденные аномалии, вызывающие серьезные проблемы со здоровьем. Девяносто процентов заболевших умирают в младенчестве. У них характерные сжатые руки и переплетенные пальцы.
- Изодицентрическая 15 , также называемая idic(15), частичная тетрасомия 15q или инвертированная дупликация 15 (inv dup 15).
- Синдром Якобсена , который встречается очень редко. Его также называют синдромом делеции терминала 11q. [55] Пострадавшие имеют нормальный интеллект или легкую инвалидность в развитии, а также плохие навыки выразительной речи. У большинства из них имеется нарушение свертываемости крови, называемое синдромом Парижа-Труссо .
- Синдром Клайнфельтера (XXY). Мужчины с синдромом Клайнфельтера обычно бесплодны, обычно выше ростом и имеют более длинные руки и ноги, чем их сверстники. Мальчики с этим синдромом часто застенчивы и тихи, у них чаще наблюдается задержка речи и дислексия . Без лечения тестостероном у некоторых может развиться гинекомастия в период полового созревания.
- Синдром Патау , также называемый D-синдромом или трисомией-13. Симптомы чем-то похожи на симптомы трисомии-18, но без характерной сложенной руки.
- Малая нештатная маркерная хромосома . Это означает, что существует дополнительная, аномальная хромосома. Особенности зависят от происхождения дополнительного генетического материала. Синдром кошачьего глаза и синдром изодицентрической хромосомы 15 (или Idic15) вызваны нештатной маркерной хромосомой, как и синдром Паллистера-Киллиана .
- Синдром тройного Х (ХХХ). Девочки XXX, как правило, высокие и худые и чаще страдают дислексией.
- Синдром Тернера (X вместо XX или XY). При синдроме Тернера женские половые признаки присутствуют, но недостаточно развиты. Женщины с синдромом Тернера часто имеют невысокий рост, низкую линию роста волос, аномальные черты глаз и развитие костей, а также «прогнутую» грудь.
- Синдром Вольфа-Хиршхорна , вызванный частичной делецией короткого плеча хромосомы 4. Он характеризуется задержкой роста, задержкой развития двигательных навыков, чертами лица «греческого шлема» и легкими или глубокими проблемами психического здоровья.
- Синдром XYY . Мальчики XYY обычно выше своих братьев и сестер. Как и XXY-мальчики и XXX-девочки, они чаще испытывают трудности с обучением.
спермы Анеуплоидия
Воздействие на мужчин определенного образа жизни, окружающей среды и/или профессиональных опасностей может увеличить риск образования анеуплоидных сперматозоидов. [56] В частности, риск анеуплоидии увеличивается при курении табака. [57] [58] и профессиональное воздействие бензола, [59] инсектициды, [60] [61] и перфторированные соединения. [62] Повышенная анеуплоидия часто связана с повышенным повреждением ДНК сперматозоидов.
Численность в различных организмах [ править ]
У эукариотов [ править ]
Число хромосом у эукариот сильно варьирует (см. таблицу). Фактически, хромосомы могут сливаться или разрываться и таким образом превращаться в новые кариотипы. Хромосомы также могут быть слиты искусственно. Например, 16 хромосом дрожжей были слиты в одну гигантскую хромосому, и клетки все еще были жизнеспособны, лишь с несколько сниженной скоростью роста. [63]
В таблицах ниже указано общее количество хромосом (включая половые хромосомы) в ядре клетки. Например, большинство эукариот диплоидны люди , как и , у которых есть 22 различных типа аутосом , каждая из которых представлена в виде двух гомологичных пар и двух половых хромосом . Всего это дает 46 хромосом. Другие организмы имеют более двух копий своих типов хромосом, например мягкая пшеница , которая является гексаплоидной и имеет шесть копий семи различных типов хромосом – всего 42 хромосомы.
|
|
|
Все нормальные представители определенного вида эукариот имеют одинаковое количество ядерных хромосом (см. таблицу). Другие эукариотические хромосомы, т.е. митохондриальные и плазмидоподобные небольшие хромосомы, гораздо более изменчивы по количеству, и их могут быть тысячи копий на клетку.

Виды , размножающиеся бесполым путем, имеют один набор хромосом, одинаковый во всех клетках тела. Однако бесполые виды могут быть как гаплоидными, так и диплоидными.
Виды, размножающиеся половым путем, имеют соматические клетки (клетки тела), которые являются диплоидными [2n] и имеют два набора хромосом (23 пары у человека), один набор от матери и один от отца. Гаметы , репродуктивные клетки, гаплоидны [n]: они имеют один набор хромосом. Гаметы образуются в результате мейоза диплоидной зародышевой клетки. Во время мейоза совпадающие хромосомы отца и матери могут обмениваться небольшими частями друг друга ( кроссинговер ) и, таким образом, создавать новые хромосомы, которые не наследуются исключительно от одного из родителей. При слиянии мужской и женской гамет ( оплодотворении ) образуется новый диплоидный организм.
Некоторые виды животных и растений являются полиплоидными [Xn]: они имеют более двух наборов гомологичных хромосом . Растения, важные в сельском хозяйстве, такие как табак или пшеница, часто являются полиплоидными по сравнению с их предковыми видами. Пшеница имеет гаплоидный набор из семи хромосом, который до сих пор наблюдается у некоторых сортов, а также у диких прародителей. Более распространенные виды макаронных изделий и мягкой пшеницы являются полиплоидными, имеющими 28 (тетраплоидные) и 42 (гексаплоидные) хромосомы по сравнению с 14 (диплоидными) хромосомами у дикой пшеницы. [89]
У прокариотов [ править ]
прокариот Виды обычно имеют по одной копии каждой основной хромосомы, но большинство клеток могут легко выжить, имея несколько копий. [90] Например, Buchnera , симбионт тли , имеет несколько копий своей хромосомы в диапазоне от 10 до 400 копий на клетку. [91] Однако у некоторых крупных бактерий, таких как Epulopiscium fishelsoni, может присутствовать до 100 000 копий хромосомы. [92] Плазмиды и плазмидоподобные малые хромосомы, как и у эукариот, сильно различаются по числу копий. Количество плазмид в клетке почти полностью определяется скоростью деления плазмиды – быстрое деление приводит к высокому числу копий.
См. также [ править ]
- Анеуплоидия
- Хромомера
- Расхождение хромосом
- Когезин
- Конденсин
- ДНК
- Генетическая делеция
- Эпигенетика
- Информацию о хромосомах в генетических алгоритмах см. в разделе хромосома (генетический алгоритм).
- Генетическая генеалогия
- Хромосома ламповой щетки
- Список числа хромосом различных организмов
- Локус (объясняет номенклатуру расположения генов)
- Влияние матери на определение пола
- Микрохромосома
- Минихромосома
- Нерасхождение
- Вторичная хромосома
- Система определения пола
- Политенная хромосома
- Протамин
- неохромосома
- Паразитарная хромосома
Примечания и ссылки [ править ]
- ^ Хаммонд CM, Стрёмме CB, Хуан Х, Патель DJ, Грот А (март 2017 г.). «Сети гистонов-шаперонов, формирующие функцию хроматина» . Обзоры природы. Молекулярно-клеточная биология . 18 (3): 141–158. дои : 10.1038/номер.2016.159 . ПМК 5319910 . ПМИД 28053344 .
- ^ Уилсон, Джон (2002). Молекулярная биология клетки: проблемный подход . Нью-Йорк: Garland Science. ISBN 978-0-8153-3577-1 .
- ^ Бонев, Боян; Кавалли, Джакомо (14 октября 2016 г.). «Организация и функции 3D-генома». Обзоры природы Генетика . 17 (11): 661–678. дои : 10.1038/nrg.2016.112 . hdl : 2027.42/151884 . ПМИД 27739532 . S2CID 31259189 .
- ^ Альбертс Б., Брэй Д., Хопкин К., Джонсон А., Льюис Дж., Рафф М., Робертс К., Уолтер П. (2014). Основная клеточная биология (Четвертое изд.). Нью-Йорк, Нью-Йорк, США: Garland Science. стр. 621–626. ISBN 978-0-8153-4454-4 .
- ↑ Перейти обратно: Перейти обратно: а б Шлейден, MJ (1847). Микроскопические исследования соответствия строения и роста животных и растений . Напечатано для Сиденхемского общества.
- ^ Антонин В., Нойманн Х. (июнь 2016 г.). «Конденсация и деконденсация хромосом во время митоза» (PDF) . Современное мнение в области клеточной биологии . 40 : 15–22. дои : 10.1016/j.ceb.2016.01.013 . ПМИД 26895139 .
- ^ Джонс, Дэниел (2003) [1917], Питер Роуч; Джеймс Хартманн; Джейн Сеттер (ред.), Словарь английского произношения , Кембридж: Издательство Кембриджского университета, ISBN 978-3-12-539683-8
- ^ «Хромосома» . Словарь Merriam-Webster.com .
- ^ Кокс, HJ (1925). Биологические пятна - Справочник по природе и использованию красителей, используемых в биологической лаборатории . Комиссия по стандартизации биологических красителей.
- ^ Вальдейер-Харц (1888 г.). «О кариокинезе и его связи с процессами оплодотворения». Архив микроскопической анатомии и механики развития . 32:27 .
- ^ Гарбари Ф, Бедини Дж, Перуцци Л (2012). «Хромосомное число итальянской флоры. От фонда Caryologia до наших дней» (PDF) . Кариология – Международный журнал цитологии, цитосистематики и цитогенетики . 65 (1): 65–66. дои : 10.1080/00087114.2012.678090 . S2CID 83748967 .
- ^ Перуцци Л., Гарбари Ф., Бедини Дж. (2012). «Новые тенденции в цитогенетике и цитоэмбриологии растений: Памяти Эмилио Баттальи посвящается» . Биосистемы растений . 146 (3): 674–675. Бибкод : 2012PBios.146..674P . дои : 10.1080/11263504.2012.712553 . S2CID 83749502 .
- ^ Батталья, Эмилио (2009). «Карионема, альтернатива хромосоме, и новая кариологическая номенклатура» (PDF) . Кариология – Международный журнал цитологии, цитосистематики . 62 (4): 1–80 . Проверено 6 ноября 2017 г.
- ^ Фокин С.И. (2013). «Отто Бючли (1848–1920). Где мы преклоним колени?» (PDF) . Протистология . 8 (1): 22–35.
- ^ Мадерспехер, Флориан (2008). «Теодор Бовери и естественный эксперимент» . Современная биология . 18 (7): Р279–Р286. дои : 10.1016/j.cub.2008.02.061 . ПМИД 18397731 . S2CID 15479331 .
- ^ Карлсон, Элоф А. (2004). Наследие Менделя: происхождение классической генетики (PDF) . Колд-Спринг-Харбор, Нью-Йорк: Лабораторное издательство Колд-Спринг-Харбор. п. 88. ИСБН 978-087969675-7 .
- ^ Уилсон, Э.Б. (1925). Клетка в развитии и наследственности / Под ред. 3. Макмиллан, Нью-Йорк. п. 923.
- ^ Майр, Э. (1982). Рост биологической мысли . Гарвард. п. 749. ISBN 9780674364462
- ^ Гартлер, Стэнли М. (1 августа 2006 г.). «Число хромосом у человека: краткая история». Обзоры природы Генетика . 7 (8): 655–660. дои : 10.1038/nrg1917 . ПМИД 16847465 . S2CID 21365693 .
- ^ Танбихлер М., Шапиро Л. (ноябрь 2006 г.). «Организация хромосом и сегрегация у бактерий». Журнал структурной биологии . 156 (2): 292–303. дои : 10.1016/j.jsb.2006.05.007 . ПМИД 16860572 .
- ^ Ван Левен Дж.Т., Мейстер Р.К., Саймон С., Маккатчеон Дж.П. (сентябрь 2014 г.). «Симпатрическое видообразование у бактериального эндосимбионта приводит к образованию двух геномов с функциональностью одного» . Клетка . 158 (6): 1270–1280. дои : 10.1016/j.cell.2014.07.047 . ПМИД 25175626 . S2CID 11839535 .
- ^ Маккатчеон Дж. П., фон Долен CD (август 2011 г.). «Взаимозависимое метаболическое одеяло во гнездовом симбиозе мучнистых червецов» . Современная биология . 21 (16): 1366–72. дои : 10.1016/j.cub.2011.06.051 . ПМК 3169327 . ПМИД 21835622 .
- ^ Хан К., Ли ЗФ, Пэн Р., Чжу ЛП, Чжоу Т, Ван ЛГ, Ли С.Г., Чжан С.Б., Ху В, Ву Чж., Цинь Н., Ли Ю.З. (2013). «Необычайное расширение генома Sorangium cellulosum из щелочной среды» . Научные отчеты . 3 : 2101. Бибкод : 2013NatSR...3E2101H . дои : 10.1038/srep02101 . ПМЦ 3696898 . ПМИД 23812535 .
- ^ Хиннебуш Дж., Тилли К. (декабрь 1993 г.). «Линейные плазмиды и хромосомы бактерий» . Молекулярная микробиология . 10 (5): 917–22. дои : 10.1111/j.1365-2958.1993.tb00963.x . ПМИД 7934868 . S2CID 23852021 .
- ^ Кельман Л.М., Кельман З. (сентябрь 2004 г.). «Множественные источники репликации у архей». Тенденции в микробиологии . 12 (9): 399–401. дои : 10.1016/j.tim.2004.07.001 . ПМИД 15337158 .
- ^ Танбихлер М., Ван С.К., Шапиро Л. (октябрь 2005 г.). «Бактериальный нуклеоид: высокоорганизованная и динамичная структура» . Журнал клеточной биохимии . 96 (3): 506–21. дои : 10.1002/jcb.20519 . ПМИД 15988757 . S2CID 25355087 .
- ^ Ле ТБ, Имакаев М.В., Мирный Л.А., Лауб М.Т. (ноябрь 2013 г.). «Картирование пространственной организации бактериальной хромосомы высокого разрешения» . Наука . 342 (6159): 731–4. Бибкод : 2013Sci...342..731L . дои : 10.1126/science.1242059 . ПМЦ 3927313 . ПМИД 24158908 .
- ^ Сэндман К., Перейра С.Л., Рив Дж.Н. (декабрь 1998 г.). «Разнообразие прокариотических хромосомных белков и происхождение нуклеосомы» . Клеточные и молекулярные науки о жизни . 54 (12): 1350–64. дои : 10.1007/s000180050259 . ПМЦ 11147202 . ПМИД 9893710 . S2CID 21101836 .
- ^ Сэндман К., Рив Дж. Н. (март 2000 г.). «Структура и функциональные взаимоотношения архейных и эукариальных гистонов и нуклеосом». Архив микробиологии . 173 (3): 165–9. Бибкод : 2000ArMic.173..165S . дои : 10.1007/s002039900122 . ПМИД 10763747 . S2CID 28946064 .
- ^ Перейра С.Л., Грейлинг Р.А., Лурц Р., Рив Дж.Н. (ноябрь 1997 г.). «Архейные нуклеосомы» . Труды Национальной академии наук Соединенных Штатов Америки . 94 (23): 12633–7. Бибкод : 1997PNAS...9412633P . дои : 10.1073/pnas.94.23.12633 . ПМК 25063 . ПМИД 9356501 .
- ^ Джонсон Дж. Э., Чиу В. (апрель 2000 г.). «Структуры вирусов и вирусоподобных частиц». Современное мнение в области структурной биологии . 10 (2): 229–35. дои : 10.1016/S0959-440X(00)00073-7 . ПМИД 10753814 .
- ↑ Перейти обратно: Перейти обратно: а б с д Купер, генеральный менеджер (2019). Клетка (8-е изд.). Издательство Оксфордского университета . ISBN 978-1605357072 .
- ^ Пунперм, Равин; Таката, Хидеаки; Хамано, Тору; Мацуда, Ацуши; Утияма, Сусуму; Хираока, Ясуши; Фукуи, Киичи (1 июля 2015 г.). «Хромосомный каркас представляет собой двухцепочечную сборку каркасных белков» . Научные отчеты . 5 (1): 11916. Бибкод : 2015НатСР...511916П . дои : 10.1038/srep11916 . ПМЦ 4487240 . ПМИД 26132639 .
- ^ Лодиш, UH; Лодиш, Х.; Берк, А.; Кайзер, Калифорния; Кайзер, К.; Кайзер, УЦА; Кригер, М.; Скотт, член парламента; Бретчер, А.; Пло, Х.; другие (2008). Молекулярно-клеточная биология . У. Х. Фриман. ISBN 978-0-7167-7601-7 .
- ^ « Картирование хромосом: идиограммы » Nature Education - 13 августа 2013 г.
- ^ Наумова Н., Имакаев М., Фуденберг Г., Жан Ю., Лажуа Б.Р., Мирный Л.А., Деккер Дж. (ноябрь 2013 г.). «Организация митотической хромосомы» . Наука . 342 (6161): 948–53. Бибкод : 2013Sci...342..948N . дои : 10.1126/science.1236083 . ПМК 4040465 . ПМИД 24200812 .
- ^ Vega.sanger.ad.uk , все данные в этой таблице взяты из этой базы данных, 11 ноября 2008 г.
- ^ «Обозреватель генома Ensembl 71: Homo sapiens - Сводка хромосом - Хромосома 1: 1–1 000 000» . апрель2013.archive.ensembl.org . Проверено 11 апреля 2016 г.
- ^ Процентные значения секвенирования основаны на доле части эухроматина, поскольку цели проекта «Геном человека» предусматривали определение только эухроматиновой части генома. Теломеры , центромеры и другие гетерохроматиновые области остались неопределенными, как и небольшое количество неклонируемых пробелов. Дополнительную информацию о проекте «Геном человека» см. «Секвенирование генома» . Национальный центр биотехнологической информации . Архивировано из оригинала 1 апреля 2005 года.
- ^ «Хромосомная карта». Гены и болезни . Бетесда, Мэриленд: Национальный центр биотехнологической информации. 1998.
- ^ Цвета каждой строки соответствуют цветам кариограммы (см. раздел «Кариотип»).
- ^ Эрвинсия, Р.; Рианди; Нуржани, М. (2017). «Актуальность анализа хромосом человека в сравнении с концепцией мутаций в курсе генетики. Серия конференций IOP» . Материаловедение и инженерия . дои : 10.1088/1757-899x/180/1/012285 . S2CID 90739754 .
- ^ Уайт, MJD (1973). Хромосомы (6-е изд.). Лондон: Чепмен и Холл, распространяется Halsted Press, Нью-Йорк. п. 28. ISBN 978-0-412-11930-9 .
- ^ фон Винивартер Х (1912). «Исследования сперматогенеза человека». Архивы биологии . 27 (93): 147–9.
- ^ Художник Т.С. (1922). «Сперматогенез человека». Анат. Рез . 23 :129.
- ^ Художник Теофил С. (апрель 1923 г.). «Исследования сперматогенеза млекопитающих. II. Сперматогенез человека». Журнал экспериментальной зоологии . 37 (3): 291–336. Бибкод : 1923JEZ....37..291P . дои : 10.1002/jez.1400370303 .
- ^ Тио Дж. Х., Леван А. (1956). «Число хромосом человека» . Эредитас . 42 (1–2): 723–4. дои : 10.1111/j.1601-5223.1956.tb03010.x . hdl : 10261/15776 . ПМИД 345813 .
- ^ Форд CE, Хамертон Дж.Л. (ноябрь 1956 г.). «Хромосомы человека». Природа . 178 (4541): 1020–3. Бибкод : 1956Natur.178.1020F . дои : 10.1038/1781020a0 . ПМИД 13378517 . S2CID 4155320 .
- ^ Hsu TC (1979) Цитогенетика человека и млекопитающих: историческая перспектива . Спрингер-Верлаг, Нью-Йорк ISBN 9780387903644 с. 10: «Удивительно, что он [Пейнтер] даже приблизился!»
- ^ «Хромосомные аномалии» , «Понимание генетики: Нью-Йорк», Среднеатлантическое руководство для пациентов и медицинских работников , Genetic Alliance, 8 июля 2009 г. , получено 27 сентября 2023 г.
- ^ Сантагуида С., Амон А. (август 2015 г.). «Краткосрочные и долгосрочные последствия неправильной сегрегации хромосом и анеуплоидии» (PDF) . Обзоры природы. Молекулярно-клеточная биология . 16 (8): 473–85. дои : 10.1038/nrm4025 . hdl : 1721.1/117201 . ПМИД 26204159 . S2CID 205495880 .
- ^ «Генетические нарушения» . medlineplus.gov . Проверено 27 апреля 2022 г.
- ^ Миллер КР (2000). «Глава 9-3». Биология (5-е изд.). Река Аппер-Сэддл, Нью-Джерси: Прентис-Холл. стр. 194–5 . ISBN 978-0-13-436265-6 .
- ^ «Что такое трисомия 18?» . Фонд трисомии 18 . Архивировано из оригинала 30 января 2017 года . Проверено 4 февраля 2017 г.
- ^ «Удаление терминала» . Европейская сеть хромосом 11 . Проверено 20 февраля 2023 г.
- ^ Темпландо С, Уроз Л, Эстоп А (октябрь 2013 г.). «Новые сведения о происхождении и значении анеуплоидии в сперматозоидах человека». Молекулярная репродукция человека . 19 (10): 634–43. дои : 10.1093/моль/gat039 . ПМИД 23720770 .
- ^ Ши Кью, Ко Э, Барклай Л, Хоанг Т, Радемейкер А, Мартин Р (август 2001 г.). «Курение сигарет и анеуплоидия человеческой спермы». Молекулярное воспроизводство и развитие . 59 (4): 417–21. дои : 10.1002/mrd.1048 . ПМИД 11468778 . S2CID 35230655 .
- ^ Рубс Дж., Лоу Х., Мур Д., Перро С., Слотт В., Эвенсон Д., Селеван С.Г., Выробек А.Дж. (октябрь 1998 г.). «Курение сигарет связано с увеличением дисомии сперматозоидов у мужчин-подростков» . Фертильность и бесплодие . 70 (4): 715–23. дои : 10.1016/S0015-0282(98)00261-1 . ПМИД 9797104 .
- ^ Син С., Маркетти Ф., Ли Г., Уэлдон Р.Х., Куртович Э., Янг С., Шмид Т.Э., Чжан Л., Раппапорт С., Вайдьянатха С., Выробек А.Дж., Эскенази Б (июнь 2010 г.). «Воздействие бензола, близкое к допустимому пределу в США, связано с анеуплоидией сперматозоидов» . Перспективы гигиены окружающей среды . 118 (6): 833–9. дои : 10.1289/ehp.0901531 . ПМЦ 2898861 . ПМИД 20418200 .
- ^ Ся Ю, Бянь Ц, Сюй Л, Ченг С, Сун Л, Лю Дж, Ву В, Ван С, Ван X (октябрь 2004 г.). «Генотоксическое воздействие на сперматозоиды человека среди рабочих пестицидных заводов, подвергшихся воздействию фенвалерата». Токсикология . 203 (1–3): 49–60. дои : 10.1016/j.tox.2004.05.018 . ПМИД 15363581 . S2CID 36073841 .
- ^ Ся Ю, Ченг С, Бянь Ц, Сюй Л, Коллинз МД, Чанг ХК, Сун Л, Лю Дж, Ван С, Ван X (май 2005 г.). «Генотоксическое воздействие на сперматозоиды рабочих, подвергшихся воздействию карбарила» . Токсикологические науки . 85 (1): 615–23. дои : 10.1093/toxsci/kfi066 . ПМИД 15615886 .
- ^ Говернини Л., Герранти С., Де Лео В., Боски Л., Лудди А., Гори М., Орвието Р., Пьомбони П. (ноябрь 2015 г.). «Хромосомные анеуплоидии и фрагментация ДНК сперматозоидов человека от пациентов, подвергшихся воздействию перфторированных соединений» . Андрология . 47 (9): 1012–9. дои : 10.1111/and.12371 . hdl : 11365/982323 . ПМИД 25382683 . S2CID 13484513 .
- ^ Шао, Янъян; Ву, Чжэньфан; Ван, Лин-Ли; Сяо, Шицзюнь; Цзэн, Сяофэй (август 2018 г.). Создание функциональных однохромосомных дрожжей» . Nature . 560 ): 331–335. : 2018Natur.560..331S doi « : 10.1038 s41586-018-0382- . 5 ( 7718 Bibcode / x . S2CID 51894920 .
- ^ Армстронг С.Дж., Джонс Г.Х. (январь 2003 г.). «Мейотическая цитология и поведение хромосом у Arabidopsis thaliana дикого типа» . Журнал экспериментальной ботаники . 54 (380): 1–10. дои : 10.1093/jxb/54.380.1 . ПМИД 12456750 .
- ^ Гилл Б.С., Кимбер Дж. (апрель 1974 г.). «С-полосный кариотип ржи по Гимзе» . Труды Национальной академии наук Соединенных Штатов Америки . 71 (4): 1247–9. Бибкод : 1974PNAS...71.1247G . дои : 10.1073/pnas.71.4.1247 . ПМЦ 388202 . ПМИД 4133848 .
- ↑ Перейти обратно: Перейти обратно: а б с Дубцовский Дж., Луо М.К., Чжун Г.И., Бранштейн Р., Десаи А., Килиан А., Кляйнхофс А., Дворак Дж. (июнь 1996 г.). «Генетическая карта диплоидной пшеницы Triticum monococcum L. и ее сравнение с картами Hordeum vulgare L» . Генетика . 143 (2): 983–99. дои : 10.1093/генетика/143.2.983 . ПМК 1207354 . ПМИД 8725244 .
- ^ Като А., Лэмб Дж. К., Бирчлер Дж. А. (сентябрь 2004 г.). «Окрашивание хромосом с использованием повторяющихся последовательностей ДНК в качестве зондов для идентификации соматических хромосом у кукурузы» . Труды Национальной академии наук Соединенных Штатов Америки . 101 (37): 13554–9. Бибкод : 2004PNAS..10113554K . дои : 10.1073/pnas.0403659101 . ПМК 518793 . ПМИД 15342909 .
- ^ Кентон А., Пароконный А.С., Глеба Ю.Ю., Беннетт, доктор медицинских наук (август 1993 г.). «Характеристика генома Nicotiana tabacum L. с помощью молекулярной цитогенетики». Молекулярная и общая генетика . 240 (2): 159–69. дои : 10.1007/BF00277053 . ПМИД 8355650 . S2CID 6953185 .
- ^ Лейтч И.Дж., Солтис Д.Э., Солтис П.С. , Беннетт, доктор медицинских наук (январь 2005 г.). «Эволюция количества ДНК у наземных растений (эмбриофит)» . Анналы ботаники . 95 (1): 207–17. дои : 10.1093/aob/mci014 . ПМЦ 4246719 . ПМИД 15596468 .
- ^ Амбариш, Китай; Шридхар, КР (2014). «Цитологические и кариологические наблюдения за двумя эндемичными гигантскими многоножками-таблетками Arthrosphaera (Pocock 1895) (Diplopoda: Sphaerotheriida) Западных Гат Индии». Кариология . 67 (1): 49–56. дои : 10.1080/00087114.2014.891700 . S2CID 219554731 .
- ^ Виттури Р., Коломба М.С., Пирроне А.М., Мандриоли М. (2002). «Колокализация рДНК (18S–28S и 5S) и связь между рибосомными генами и теломерной последовательностью (TTAGGG) (n) у дождевого червя Octodrilus complanatus (Annelida: Oligochaeta: Lumbricidae), выявленные с помощью одно- и двухцветного FISH» . Журнал наследственности . 93 (4): 279–82. дои : 10.1093/jhered/93.4.279 . ПМИД 12407215 .
- ^ Не В., Ван Дж., О'Брайен ПК, Фу Б., Ин Т., Фергюсон-Смит М.А., Ян Ф. (2002). «Филогения генома домашней кошки, красной панды и пяти видов куньих, выявленная с помощью сравнительной окраски хромосом и G-бэндинга». Хромосомные исследования . 10 (3): 209–22. дои : 10.1023/А:1015292005631 . ПМИД 12067210 . S2CID 9660694 .
- ↑ Перейти обратно: Перейти обратно: а б Романенко С.А., Перельман П.Л., Сердюкова Н.А., Трифонов В.А., Билтуева Л.С., Ван Дж, Ли Т, Ни В, О'Брайен П.С., Волобуев В.Т., Станьон Р., Фергюсон-Смит М.А., Ян Ф., Графодатский А.С. (декабрь 2006 г.). «Взаимная окраска хромосом между тремя видами лабораторных грызунов». Геном млекопитающих . 17 (12): 1183–92. дои : 10.1007/s00335-006-0081-z . ПМИД 17143584 . S2CID 41546146 .
- ↑ Перейти обратно: Перейти обратно: а б Художник Т.С. (март 1928). «Сравнение хромосом крысы и мыши применительно к вопросу гомологии хромосом у млекопитающих» . Генетика . 13 (2): 180–9. дои : 10.1093/генетика/13.2.180 . ПМК 1200977 . ПМИД 17246549 .
- ^ Хейс Х., Рогель-Гайяр С., Зийлстра С., Де Хаан Н.А., Уриен С., Буржо Н., Берто М., Босма А.А. (2002). «Создание номенклатуры кариотипов кроликов с R-диапазонами с помощью FISH-локализации 23 хромосомно-специфичных генов как на G-, так и на R-хромосомах». Цитогенетические и геномные исследования . 98 (2–3): 199–205. дои : 10.1159/000069807 . ПМИД 12698004 . S2CID 29849096 .
- ^ «Генетика популярного аквариумного питомца – рыбок гуппи» . Архивировано из оригинала 31 мая 2023 года . Проверено 6 декабря 2009 г.
- ↑ Перейти обратно: Перейти обратно: а б Де Груши Дж. (август 1987 г.). «Хромосомная филогения человека, человекообразных обезьян и обезьян Старого Света». Генетика . 73 (1–2): 37–52. дои : 10.1007/bf00057436 . ПМИД 3333352 . S2CID 1098866 .
- ^ Робинсон Т.Дж., Ян Ф., Харрисон В.Р. (2002). «Хромосомная окраска уточняет историю эволюции генома зайцев и кроликов (отряд зайцеобразных)». Цитогенетические и геномные исследования . 96 (1–4): 223–7. дои : 10.1159/000063034 . ПМИД 12438803 . S2CID 19327437 .
- ^ Чепмен Дж.А., Флакс Дж.Е. (1990), «раздел 4.W4», Кролики, зайцы и пищухи. Обзор состояния и план действий по сохранению , стр. 61–94, ISBN. 9782831700199
- ^ Виттури Р., Либертини А., Синео Л., Спарацио И., Ланнино А., Грегорини А., Коломба М. (2005). «Цитогенетика наземных улиток Cantareus aspersus и C. mazzullii (Mollusca: Gastropoda: Pulmonata)». Микроны . 36 (4): 351–7. дои : 10.1016/j.micron.2004.12.010 . ПМИД 15857774 .
- ^ Ясукочи Ю., Ашакумари Л.А., Баба К., Ёсидо А., Сахара К. (июль 2006 г.). «Интегрированная карта тутового шелкопряда второго поколения показывает синтению и консервативный порядок генов между чешуекрылыми насекомыми» . Генетика . 173 (3): 1319–28. doi : 10.1534/genetics.106.055541 . ПМЦ 1526672 . ПМИД 16547103 .
- ^ Хоук М.Л., Кумамото А.Т., Галлахер Д.С., Бениршке К. (2001). «Сравнительная цитогенетика африканского слона (Loxodonta africana) и азиатского слона (Elephas maximus)». Цитогенетика и клеточная генетика . 93 (3–4): 249–52. дои : 10.1159/000056992 . ПМИД 11528120 . S2CID 23529399 .
- ^ Семба У, Умеда Ю, Сибуя Ю, Окабе Х, Танасе С, Ямамото Т (октябрь 2004 г.). «Первичные структуры высокомолекулярных и низкомолекулярных кининогенов морских свинок». Международная иммунофармакология . 4 (10–11): 1391–400. дои : 10.1016/j.intimp.2004.06.003 . ПМИД 15313436 .
- ^ Уэйн Р.К., Острандер Э.А. (март 1999 г.). «Происхождение, генетическое разнообразие и структура генома домашней собаки». Биоэссе . 21 (3): 247–57. doi : 10.1002/(SICI)1521-1878(199903)21:3<247::AID-BIES9>3.0.CO;2-Z . ПМИД 10333734 . S2CID 5547543 .
- ^ Сьюдад Дж., Сид Э., Веласко А., Лара Х.М., Айон Дж., Орфао А. (май 2002 г.). «Измерение содержания ДНК диплоидных клеток G0/G1 с помощью проточной цитометрии трех различных видов костистых рыб». Цитометрия . 48 (1): 20–5. дои : 10.1002/cyto.10100 . ПМИД 12116377 .
- ^ Берт Д.В. (2002). «Происхождение и эволюция птичьих микрохромосом». Цитогенетические и геномные исследования . 96 (1–4): 97–112. дои : 10.1159/000063018 . ПМИД 12438785 . S2CID 26017998 .
- ^ Ито М., Икеучи Т., Симба Х., Мори М., Сасаки М., Макино С. (1969). «Сравнительное исследование кариотипа четырнадцати видов птиц» . Японский журнал генетики . 44 (3): 163–170. дои : 10.1266/jjg.44.163 .
- ^ Смит Дж., Берт Д.В. (август 1998 г.). «Параметры генома курицы (Gallus Gallus)». Генетика животных . 29 (4): 290–4. дои : 10.1046/j.1365-2052.1998.00334.x . ПМИД 9745667 .
- ^ Сакамура, Тецу (1918). «Краткая справка о числе хромосом и взаимоотношениях видов Triticum» . Сёкубуцугаку Засси . 32 (379): 150–3. дои : 10.15281/jplantres1887.32.379_150 .
- ^ Шарлебуа Р.Л. (редактор) 1999. Организация генома прокариот . ASM Press, Вашингтон, округ Колумбия.
- ^ Комаки К., Исикава Х (март 2000 г.). «Количество геномных копий внутриклеточных бактериальных симбионтов тлей варьируется в зависимости от стадии развития и морфологии их хозяина». Биохимия насекомых и молекулярная биология . 30 (3): 253–8. дои : 10.1016/S0965-1748(99)00125-3 . ПМИД 10732993 .
- ^ Менделл Дж. Э., Клементс К. Д., Чоат Дж. Х., Ангерт Э. Р. (май 2008 г.). «Чрезвычайная полиплоидия у крупной бактерии» . Труды Национальной академии наук Соединенных Штатов Америки . 105 (18): 6730–4. Бибкод : 2008PNAS..105.6730M . дои : 10.1073/pnas.0707522105 . ПМЦ 2373351 . ПМИД 18445653 .
Внешние ссылки [ править ]
- Введение в ДНК и хромосомы от HOPES : информационно-пропагандистский проект Хантингтона в области образования в Стэнфорде
- Хромосомные аномалии в AtlasGeneticsOncology
- Он-лайн выставка хромосом и генома (СИБ)
- О чем могут рассказать нам наши хромосомы? , из Учебного центра генетических наук Университета Юты.
- Попробуйте самостоятельно сделать кариотип в Учебном центре генетики Университета Юты.
- Страницы хромосом Кимбалла
- Хромосомные новости от Genome News Network
- Eurochromnet , Европейская сеть Интернет по редким хромосомным заболеваниям
- Ensembl.org , Ensembl проект , графически представляющий хромосомы, их гены и синтенные локусы через Интернет.
- Генографический проект. Архивировано 12 июля 2007 г. в Wayback Machine.
- Домашний справочник по хромосомам из Национальной медицинской библиотеки США.
- Визуализация хромосом человека и сравнение с другими видами
- Уникальный – Группа поддержки редких хромосомных заболеваний. Поддержка людей с редкими хромосомными заболеваниями.