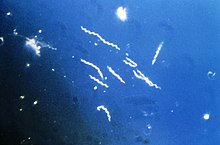
Боррелия бургдорфери
| Боррелия бургдорфери | |
|---|---|
 | |
| Боррелия бургдорфери | |
| Научная классификация | |
| Домен: | Бактерии |
| Тип: | Спирохетата |
| Сорт: | Спирохета |
| Заказ: | Спирохеты |
| Семья: | Боррелиевые |
| Род: | Боррелия |
| Разновидность: | Б. бургдорфери |
| Биномиальное имя | |
| Боррелия бургдорфери Джонсон и др. 1984 г. Бартон и др. 1992 год | |
Borrelia burgdorferi — бактерий вид класса спирохет рода Borrelia , один из возбудителей болезни Лайма у человека. [1] [2] Наряду с несколькими сходными геновидами, некоторые из которых также вызывают болезнь Лайма, он составляет комплекс видов Borrelia burgdorferi sensu lato . В настоящее время комплекс включает 20 признанных и 3 предложенных геновида. [2] B. burgdorferi sensu stricto существует в Северной Америке и Евразии и до 2016 года был единственной известной причиной болезни Лайма в Северной Америке. [3] [4] [2] Borrelia Виды являются грамотрицательными . [5]
Микробиология
[ редактировать ]Borrelia burgdorferi названа в честь исследователя Вилли Бургдорфера , который впервые выделил бактерию в 1982 году. [6]
| Тип теста | Тест | Характеристики |
| Персонажи колонии | Размер | Маленький [7] |
| Тип | Круглый [7] | |
| Цвет | Белый [7] | |
| Форма | Поднятый [7] | |
| Морфологические символы | Форма | Спирохета [8] |
| Физиологические признаки | Подвижность | + [8] |
| Рост при 6,5% NaCl | + [8] | |
| Биохимические признаки | Окраска по Граму | - |
| Оксидаза | - [9] | |
| Каталаза | - [9] | |
| Окислительно-ферментативный | Ферментативный [10] | |
| β-галактозидаза | + [11] | |
| Использование | Глицерин | + [12] |
| Галактоза | + [13] | |
| D-глюкоза | + [13] | |
| D-манноза | + [13] |
Borrelia burgdorferi — микроаэрофил, которому требуется небольшое количество кислорода для гликолиза и выживания. Как и все другие виды Borrelia , эта бактерия также является грамотрицательной и является спирохетой. Колонии боррелий часто меньшего размера, округлой формы, белого цвета с приподнятым центром. [7] B. burgdorferi обладает жгутиками , которые обеспечивают ему подвижность. Он может быть оксидазо-отрицательным, но B. burgdorferi обладает геном, кодирующим супероксиддисмутазу . Этот белок ингибирует накопление активных форм кислорода (АФК). [9] Бактерия, по-видимому, способна использовать множество различных моносахаридов для производства энергии. [13]
Морфология
[ редактировать ]B. burgdorferi напоминает другие спирохеты тем, что имеет внешнюю и внутреннюю мембраны с тонким слоем пептидогликана между ними. Он характеризуется наличием гибкой клеточной лунки и длинными цилиндрическими клетками шириной примерно 1 микрон. Однако внешняя мембрана лишена липополисахарида . Его форма — плоская волна. Его ширина составляет около 0,3 мкм, а длина от 5 до 20 мкм. [14]
B. burgdorferi представляет собой микроаэробную подвижную спирохету с семью-11 пучками периплазматических жгутиков, расположенных на каждом конце, которые позволяют бактерии перемещаться как в средах с низкой, так и с высокой вязкостью, что связано с ее высоким фактором вирулентности. [15]
Метаболизм
[ редактировать ]B. burgdorferi представляет собой медленнорастущую микроаэрофильную спирохету со временем удвоения от 24 до 48 часов. [16]
Трансформация
[ редактировать ]Бактериальная трансформация использовалась исследователями для выделения специфических патогенных генов среди Borrelia burgdorferi . Штаммы B. burgdorferi кажутся совершенно недостаточными для использования в бактериальной трансформации из-за большого количества ДНК, необходимого для трансформации, времени, необходимого для получения надежных трансформантов, и влияния систем рестрикционной модификации . [17] [18] Фактически, для инфекционности B. burgdorferi часто требуется ген pncA , который присутствует на бактериальной плазмиде, содержащей ген bbe02 , который подвергается тщательному отбору во время трансформации. Поскольку эти гены часто соединяются вместе, при трансформации отбирается инфекционность, что препятствует исследованиям по выявлению конкретных генов, которые участвуют в патогенности Borrelia burgdorferi . [19] Несмотря на это, был достигнут некоторый прогресс в разгадке загадок B. burgdorferi, например, открытие гена cyaB , необходимого для заражения млекопитающих. [20]
Жизненный цикл
[ редактировать ]B. burgdorferi циркулирует между иксодовыми клещами и позвоночным хозяином в рамках энзоотического цикла . [2] B. burgdorferi, живущая в клеще, в основном передается через кровь от инфицированного компетентного позвоночного хозяина. [21] но существуют редкие случаи трансовариальной передачи . [22] После заражения клещ передает B. burgdorferi , питаясь другим позвоночным, чтобы завершить цикл. [23] Клещи могут передавать B. burgdorferi человеку, но люди являются тупиковыми хозяевами и вряд ли смогут продолжить жизненный цикл спирохеты. [24] Нимфы линяют во взрослых клещей, которые обычно питаются более крупными млекопитающими, не способными поддерживать выживание B. burgdorferi . [25]
Болезнь
[ редактировать ]B. burgdorferi является возбудителем болезни Лайма , и именно поэтому эта бактерия так важна и изучается. Чаще всего он передается от клещей человеку. клеща Человек выступает в качестве хозяина для этой бактерии. Болезнь Лайма — зоонозное передающееся трансмиссивное заболевание, иксодовым клещом ( также переносчиком бабезий и анаплазм ). Инфицированный нимфальный клещ передает B. burgdorferi через слюну человеку во время еды с кровью. [25]
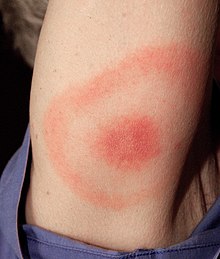
Клиническая картина болезни Лайма наиболее известна характерной сыпью типа «бычий глаз» (также известной как хроническая мигрирующая эритема ), но может также включать миокардит , кардиомиопатию , аритмию , артрит , артралгию , менингит , невропатии и паралич лицевого нерва . [26] в зависимости от стадии заражения.
Инфекции B. burgdorferi были обнаружены в возможной связи с первичными кожными диффузными крупноклеточными В-клеточными лимфомами (PCDLBCL). [27] [28] где обзор первичной литературы по состоянию на 2010 год показал, что большинство исследованных PCBCL «не реагируют» на антибиотики; [28] : 846 следовательно, как и в случае Chlamydophila psittaci ассоциации с лимфомой лимфоидной ткани, ассоциированной со слизистой оболочкой глаза (MALT-лимфомой), рабочий вывод заключался в том, что «если B. burgdorferi действительно связана с PCBCL, то существует широкая географическая изменчивость и другие факторы вероятно, причастен». [28] : 846
Развитие заболевания протекает в три этапа.
Этап 1
[ редактировать ]Стадия 1 известна как ранняя локализованная стадия и возникает примерно через 3 дня – 1 месяц после инокуляции. Он поражает область вокруг укуса и характеризуется локальным отеком и/или красной сыпью в форме «бычьего глаза» (также известной как хроническая мигрирующая эритема ), которая выглядит как эритематозный круг, окружающий определенный центр, который расширяется наружу. В диаметре он может достигать 15 см. [29] : 658 Как только сыпь начнет стихать, первые симптомы могут проявиться как симптомы гриппа. На этом этапе антибиотики наиболее эффективны для предотвращения дальнейшего роста и симптомов заболевания до того, как проявятся основные симптомы. [29] : 659
Этап 2
[ редактировать ]Стадия 2 известна как ранняя диссеминированная стадия и возникает через несколько недель или месяцев после заражения, если ее не лечить. Бактерии распространяются через кровь по организму и поражают органы. Он часто проявляется как общими симптомами, такими как лихорадка, озноб, усталость и лимфаденопатия, так и органоспецифическими симптомами. Это может повлиять на сердце, вызывая миокардит , а также аритмии, такие как атриовентрикулярные блокады (которые, если они достаточно значительны, могут потребовать установки кардиостимулятора). Он может поражать опорно-двигательный аппарат, вызывая невоспалительный преходящий артрит и/или артралгии . Это может повлиять на нервную систему, проявляясь параличом лицевого нерва ( классически двусторонний паралич Белла ), утомляемостью и потерей памяти. [ нужна ссылка ]
Этап 3
[ редактировать ]Стадия 3 известна как поздняя диссеминированная стадия и возникает через несколько месяцев или лет после первоначального заражения. К последствиям 3-й стадии относятся энцефалит или менингит, [29] а также мигрирующие артропатии (чаще всего коленного сустава). [29]
Анаплазмоз и бабезиоз также являются распространенными клещевыми возбудителями, переносимыми иксодовым клещом, которые заражают людей так же, как Borrelia burgdorferi . [30] Следовательно, иксодовый клещ может одновременно заразить хозяина двумя или всеми другими заболеваниями. Когда хозяин коинфицирован, совокупные эффекты болезней действуют синергично, часто вызывая более серьезные симптомы, чем одна инфекция в отдельности. [30] У людей с коинфекцией, как правило, наблюдаются более тяжелые проявления болезни Лайма. Кроме того, они имеют тенденцию приобретать более широкий спектр вторичных симптомов, таких как гриппоподобные симптомы. [30] Необходимо провести дополнительные исследования и исследования, чтобы определить синергетический эффект коинфекции и ее влияние на организм человека.
Изменение степени тяжести
[ редактировать ]На данный момент существует три фактора, которые могут способствовать тяжести клинических проявлений болезни Лайма. Наличие рибосомальных спейсеров, плазмид и белка С внешней поверхности (OspC) являются показателями тяжести инфекции. [31] Кроме того, сами люди по-разному реагируют на инфекцию. [31] Различия в реакции приводят к различным клиническим проявлениям и различным инфекциям разных органов. [ нужна ссылка ]
Молекулярный патогенез
[ редактировать ]После передачи возбудителя он акклиматизируется к условиям обитания млекопитающих. Borrelia burgdorferi меняет гликопротеины и протеазы на плазматической мембране, чтобы облегчить распространение по крови. [31] Во время заражения B. burgdorferi будет экспрессировать белки, которые будут взаимодействовать с эндотелиальными клетками , тромбоцитами , хондроцитами и внеклеточным матриксом . [31] Это взаимодействие подавляет правильное функционирование инфицированных участков, что приводит к патологическим проявлениям болезни Лайма. В ответ хозяин инициирует воспалительную реакцию , пытаясь удалить инфекцию. [31]
Borrelia burgdorferi также экспрессирует по меньшей мере семь белков, связывающих плазминоген, для вмешательства в фактор H на уровне активации. Это часть стратегии уклонения от системы комплемента , которая приводит к последующей блокировке иммунного ответа. [32]
Кроме того, Borrelia burgdorferi обладает стратегией прямого ингибирования классического пути системы комплемента. Боррелиальный липопротеин BBK32, экспрессируемый на поверхности Borrelia burgdorferi , связывает инициирующий протеазный комплекс С1 классического пути. Более конкретно, BBK32 взаимодействует с субъединицей C1r C1. С-концевой домен белка BBK32 опосредует связывание. В результате C1 оказывается в неактивной форме. [33]
Геном
[ редактировать ]B. burgdorferi (штамм B31) был третьим микробным геномом когда-либо секвенированным после секвенирования Haemophilus influenzae и Mycoplasmagentium в 1995 году. Его линейная хромосома содержит 910 725 пар оснований и 853 гена . [34] В качестве метода секвенирования использовался метод полногеномного секвенирования . Проект секвенирования, опубликованный в журналах Nature в 1997 году и Molecular Microbiology в 2000 году, проводился в Институте геномных исследований . [35] Геном B. burgdorferi состоит из одной мегабазной хромосомы и необычного разнообразия кольцевых и линейных плазмид размером от 9 до 62 килобаз. [23] Мегабазная хромосома, в отличие от многих других эубактерий, не имеет никакого отношения ни к вирулентности бактерий, ни к взаимодействию хозяин-паразит. [34] Некоторые из плазмид необходимы для жизненного цикла B. burgdorferi , но не для размножения бактерий в культуре. [23]
Геномные вариации B. burgdorferi способствуют различной степени заражения и распространения. [36] Каждая геномная группа имеет на своем мембранном рецепторе различные антигены , специфичные для инфекции хозяина. Одним из таких мембранных рецепторов является поверхностный белок OspC. [36] Показано, что поверхностный белок OspC является сильным индикатором идентификации геномной классификации и степени распространения. [36] Различное количество локусов OspC является показателем и определяющим фактором вариаций B. burgdorferi . [36] Поверхностный белок также находится на переднем крае текущих исследований вакцин против болезни Лайма, вызываемых боррелиями . [37]
Бактериофаг
[ редактировать ]относительно небольшое количество бактериофагов Известно , инфицирующих B. burgdorferi . размером 8 т.п.н. дцДНК что они имели геном Было изолировано несколько фаговых частиц, и некоторые данные свидетельствуют о том , Среди наиболее изученных фагов боррелий находится φBB-1, фаг с многогранной головкой и сократительным хвостом длиной 90 нм. [38] [39] φBB-1 был первым бактериофагом, который предоставил доказательства трансдукции латерального переноса генов у видов Borrelia, вызывающих болезнь Лайма. [40] Текущие исследования направлены на использование бактериофагов как способ выявления факторов вирулентности спирохет, которые приводят к болезни Лайма. [ нужна ссылка ]
Эволюция
[ редактировать ]Генетически разнообразные штаммы B. burgdorferi , определяемые последовательностью ospC , сохраняются на северо-востоке США. Балансирующий отбор может воздействовать на ospC или близлежащую последовательность для поддержания генетического разнообразия B. burgdorferi . [41] Балансирующий отбор — это процесс, при котором несколько версий гена сохраняются в генофонде с неожиданно высокой частотой. Две основные модели, которые контролируют баланс отбора B.burgdorferi, - это отрицательный частотно-зависимый отбор и полиморфизм с множеством ниш . [42] Эти модели могут объяснить, как B. burgdorferi диверсифицировался и как отбор мог повлиять на распространение вариантов B. burgdorferi или на вариации конкретных признаков вида в определенных средах. [ нужна ссылка ]
Выбор, зависящий от отрицательной частоты
[ редактировать ]При отрицательном частотно-зависимом отборе редкие и необычные варианты будут иметь селективное преимущество перед вариантами, которые очень распространены в окружающей среде. [42] Для B. burgdorferi низкочастотные варианты будут выгодны, поскольку потенциальные хозяева с меньшей вероятностью будут вызывать иммунологический ответ на специфичный для варианта внешний белок OspC. [42]
Многонишевый полиморфизм
[ редактировать ]Экологические ниши — это все переменные окружающей среды, такие как ресурсы, конкуренты и реакции, которые способствуют приспособленности организма. Полиморфизм с несколькими нишами утверждает, что разнообразие поддерживается внутри популяции из-за различного количества возможных ниш и сред. [42] Следовательно, чем больше разнообразных ниш, тем больше вероятность полиморфизма и разнообразия. Для B. burgdorferi различные ниши позвонков, например, у оленей и мышей, могут влиять на общий балансирующий выбор вариантов. [42]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Радольф Дж.Д., Сэмюэлс Д.С., ред. (2021). Болезнь Лайма и возвратная лихорадка, спирохеты: геномика, молекулярная биология, взаимодействие с хозяином и патогенез заболевания . Кайстер Академик Пресс. ISBN 978-1-913652-61-6 .
- ^ Jump up to: а б с д Уолкотт, Кэтрин А.; Маргос, Габриэле; Фингерл, Волкер; Беккер, Ноэми С. (сентябрь 2021 г.). «Принимающая ассоциация Borrelia burgdorferi sensu lato: обзор». Клещи и клещевые заболевания . 12 (5): 101766. doi : 10.1016/j.ttbdis.2021.101766 . ПМИД 34161868 .
- ^ CDC (08 февраля 2016 г.). «Обнаружены новые виды бактерий, вызывающих болезнь Лайма» . Центры по контролю и профилактике заболеваний . Проверено 18 января 2019 г.
- ^ Тилли, Кит; Роза, Патрисия А.; Стюарт, Филип Э. (2008). «Биология заражения Borrelia burgdorferi» . Клиники инфекционных заболеваний Северной Америки . 22 (2): 217–234. дои : 10.1016/j.idc.2007.12.013 . ПМК 2440571 . ПМИД 18452798 .
- ^ Сэмюэлс Д.С.; Радольф, JD, ред. (2010). «Глава 6, Структура, функции и биогенез клеточной оболочки боррелий ». Боррелии : молекулярная биология, взаимодействие с хозяином и патогенез . Кайстер Академик Пресс. ISBN 978-1-904455-58-5 .
- ^ Бургдорфер В., Барбур А.Г., Хейс С.Ф., Бенах Дж.Л., Грюнвальдт Э., Дэвис Дж.П. (июнь 1982 г.). «Болезнь Лайма — клещевой спирохетоз?» . Наука . 216 (4552): 1317–9. Бибкод : 1982Sci...216.1317B . дои : 10.1126/science.7043737 . ПМИД 7043737 .
- ^ Jump up to: а б с д и Куртти, Ти Джей; Мандерло, Юга; Джонсон, Р.К.; Альстранд, Г.Г. (ноябрь 1987 г.). «Формирование и морфология колоний Borrelia burgdorferi» . Журнал клинической микробиологии . 25 (11): 2054–2058. doi : 10.1128/jcm.25.11.2054-2058.1987 . ISSN 0095-1137 . ПМК 269410 . ПМИД 3693538 .
- ^ Jump up to: а б с Ши, Вэньюань; Ян, Чжаоминь; Гэн, Юнчжи; Волински, Лоуренс Э.; Ловетт, Майкл А. (январь 1998 г.). «Хемотаксис у Borrelia burgdorferi» . Журнал бактериологии . 180 (2): 231–235. дои : 10.1128/JB.180.2.231-235.1998 . ISSN 0021-9193 . ПМЦ 106876 . ПМИД 9440510 .
- ^ Jump up to: а б с Трокселл, Брайан; Сюй, Хайцзюнь; Ян, X. Франк (1 июня 2012 г.). «Borrelia burgdorferi, патоген, которому не хватает железа, кодирует марганцево-зависимую супероксиддисмутазу, необходимую для устойчивости к стрептонигрину» . Журнал биологической химии . 287 (23): 19284–19293. дои : 10.1074/jbc.M112.344903 . ISSN 0021-9258 . ПМК 3365960 . ПМИД 22500025 .
- ^ Корона, Арианна; Шварц, Ира (04 августа 2015 г.), « Borrelia burgdorferi : метаболизм углерода и энзоотический цикл клещей и млекопитающих» , Метаболизм и бактериальный патогенез , том. 3, нет. 3, Вашингтон, округ Колумбия, США: ASM Press, стр. 167–184, doi : 10.1128/9781555818883.ch8 , ISBN. 9781683670926 , PMC 7942402 , PMID 26185064 , получено 21 апреля 2022 г.
- ^ Василюк, Анна; Вашкевич, Наполеон; Шайда, Славомир Дариуш; Воеводска-Железнякович, Мажена; Кемпка, Алина; Минаровская, Алина; Звеж, Збигнев Войцех; Панцевич, Славомир; Приятно, Джордж Роберт; Зверц, Кшиштоф (5 июля 2012 г.). «Альфа-фукозидаза и бета-галактозидаза в сыворотке крови пациентов с болезнью Лайма как возможный маркер ускоренного старения - предварительное исследование» . Folia Histochemica et Cytobiologica . 50 (2): 270–274. дои : 10.5603/fhc.2012.0036 . ISSN 1897-5631 . ПМИД 22763966 .
- ^ Паппас, Кристофер Дж.; Айер, Радха; Пецке, Мэри М.; Каймано, Мелисса Дж.; Радольф, Джастин Д.; Шварц, Ира (7 июля 2011 г.). «Borrelia burgdorferi требует глицерина для максимальной работоспособности во время клещевой фазы энзоотического цикла» . ПЛОС Патогены . 7 (7): е1002102. дои : 10.1371/journal.ppat.1002102 . ISSN 1553-7366 . ПМК 3131272 . ПМИД 21750672 .
- ^ Jump up to: а б с д Гулинска, Д.; Вольф, П.; Грубхоффер, Л. (апрель 1992 г.). «Характеристика гликоконъюгатов Borrelia burgdorferi и поверхностных углеводов» . Централблатт по бактериологии . 276 (4): 473–480. дои : 10.1016/s0934-8840(11)80672-9 . ISSN 0934-8840 . ПМИД 1611204 .
- ^ Моталеб М.А., Лю Дж., Вутен Р.М. (2015). «Подвижность спирохет и хемотаксис в естественном энзоотическом цикле и развитии болезни Лайма» . Современное мнение в микробиологии . 28 : 106–13. дои : 10.1016/j.mib.2015.09.006 . ПМЦ 4688064 . ПМИД 26519910 .
- ^ Моталеб, Мохаммед; Корум, Линда; Боно, Джеймс; Элиас, Абдалла; Роза, Патрисия; Сэмюэлс, Д. Скотт; Харон, Найлс (2000). «Периплазматические жгутики Borrelia burgdorferi обладают как скелетной, так и двигательной функцией» . Труды Национальной академии наук Соединенных Штатов Америки . 97 (20): 10899–10904. Бибкод : 2000PNAS...9710899M . дои : 10.1073/pnas.200221797 . ПМК 27121 . ПМИД 10995478 .
- ^ Цюкерт, Вольфрам Р. (2007). «Лабораторное содержание Borrelia burgdorferi ». Современные протоколы в микробиологии . Том. 4. стр. 12C.1.1–12C.1.10. дои : 10.1002/9780471729259.mc12c01s4 . ISBN 978-0-471-72925-9 . ПМИД 18770608 .
- ^ Бриссон, Дастин; Дыхейзен, Дэниел Э (01 октября 2004 г.). «Разнообразие ospC в Borrelia burgdorferi» . Генетика . 168 (2): 713–722. doi : 10.1534/genetics.104.028738 . ISSN 1943-2631 . ПМЦ 1448846 . ПМИД 15514047 .
- ^ Хайд, Дженни А.; Вининг, Эрик Х.; Скаре, Джон Т. (февраль 2011 г.). «Генетические манипуляции с Borrelia burgdorferi» . Современные протоколы в микробиологии . ГЛАВА: Раздел – 12C.4. дои : 10.1002/9780471729259.mc12c04s20 . ISSN 1934-8525 . ПМЦ 3561735 . ПМИД 21400675 .
- ^ Бриссон, Дастин; Дректра, Дэн; Эггерс, Кристиан Х.; Сэмюэлс, Д. Скотт (2012). «Генетика Borrelia burgdorferi» . Ежегодный обзор генетики . 46 : 10.1146/аннурев–генет–011112-112140. doi : 10.1146/annurev-genet-011112-112140 . ISSN 0066-4197 . ПМЦ 3856702 . ПМИД 22974303 .
- ^ Анте, Ванесса М.; Фаррис, Лорен С.; Сапутра, Элизабет П.; Холл, Элли Дж.; О'Биер, Натаниэль С.; Олива Чавес, Адела С.; Маркони, Ричард Т.; Любекер, Меган С.; Хайд, Дженни А. (25 мая 2021 г.). «Аденилатциклаза Borrelia burgdorferi, CyaB, важна для производства фактора вирулентности и инфекции млекопитающих» . Границы микробиологии . 12 : 676192. doi : 10.3389/fmicb.2021.676192 . ISSN 1664-302X . ПМЦ 8186283 . ПМИД 34113333 .
- ^ Эйзен, Ребекка Дж.; Эйзен, Ларс (апрель 2018 г.). «Черноногий клещ, Ixodes Scapularis: растущая проблема общественного здравоохранения» . Тенденции в паразитологии . 34 (4): 295–6. дои : 10.1016/j.pt.2017.12.006 . ПМК 5879012 . ПМИД 29336985 .
- ^ Ролленд, Линдси; Рыба, Дурланд; Чайлдс, Джеймс Э. (февраль 2013 г.). «Трансовариальная передача спирохет Borrelia Ixodes scapularis: обзор литературы и недавние наблюдения». Клещи и клещевые заболевания . 4 (1–2): 46–51. дои : 10.1016/j.ttbdis.2012.06.008 . ПМИД 23238242 .
- ^ Jump up to: а б с Бриссон Д., Дректра Д., Эггерс Ч.С., Сэмюэлс Д.С. (2012). «Генетика B. burgdorferi » . Ежегодный обзор генетики . 46 : 515–36. doi : 10.1146/annurev-genet-011112-112140 . ПМЦ 3856702 . ПМИД 22974303 .
- ^ Радольф Дж.Д., Каймано М.Дж., Стивенсон Б., Ху Л.Т. (2012). «О клещах, мышах и людях: понимание образа жизни спирохет болезни Лайма с двумя хозяевами» . Обзоры природы. Микробиология . 10 (2): 87–99. дои : 10.1038/nrmicro2714 . ПМЦ 3313462 . ПМИД 22230951 .
- ^ Jump up to: а б Тилли К., Роза П.А., Стюарт П.Е. (2008). «Биология заражения Borrelia burgdorferi» . Клиники инфекционных заболеваний Северной Америки . 22 (2): 217–34, v. doi : 10.1016/j.idc.2007.12.013 . ПМК 2440571 . ПМИД 18452798 .
- ^ «Признаки и симптомы болезни Лайма» . Центры по контролю заболеваний. 4 марта 2015 г. Проверено 16 июля 2015 г.
- ^ Гвидобони М., Феррери А.Дж., Понцони М., Дольони С., Дольчетти Р. (январь 2006 г.). «Инфекционные агенты при лимфомах лимфоидной ткани, ассоциированных со слизистой оболочкой: патогенная роль и терапевтические перспективы». Клиническая лимфома и миелома . 6 (4): 289–300. дои : 10.3816/CLM.2006.n.003 . ПМИД 16507206 .
- ^ Jump up to: а б с Чанг, А.Х.; Парсоннет, Дж. (2010). «Роль бактерий в онкогенезе» . Обзоры клинической микробиологии . 23 (4): 837–857. дои : 10.1128/CMR.00012-10 . ISSN 0893-8512 . ПМЦ 2952975 . ПМИД 20930075 .
- ^ Jump up to: а б с д Тортора, Джерард Дж.; Функе, Берделл Р.; Кейс, Кристин Л. (2013). Микробиология: Введение . Соединенные Штаты Америки: Pearson Education, Inc., стр. 658–659. ISBN 978-0-321-73360-3 .
- ^ Jump up to: а б с Суонсон, Стивен Дж.; Нейцель, Дэвид; Рид, Курт Д.; Белонгия, Эдвард А. (1 октября 2006 г.). «Коинфекции, полученные от иксодовых клещей» . Обзоры клинической микробиологии . 19 (4): 708–727. дои : 10.1128/CMR.00011-06 . ISSN 0893-8512 . ПМЦ 1592693 . ПМИД 17041141 .
- ^ Jump up to: а б с д и Вайс, Джанет (2011). «Критические потребности и пробелы в понимании профилактики, улучшения и лечения болезни Лайма и других клещевых заболеваний: краткосрочные и долгосрочные результаты: отчет о семинаре» . Национальные академии : 97–101.
- ^ Зипфель П., Халльстрем Т., Рисбек К. (2013). «Контроль комплемента человека и уклонение от комплемента патогенными микробами – нарушение баланса» . Молекулярная иммунология . 56 (3): 152–160. дои : 10.1016/j.molimm.2013.05.222 . ПМИД 23810413 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Гарсия, Б.Л., Чжи, Х., Вагер, Б., Хёк, М. и Скаре, Дж.Т. 2016, «Borrelia burgdorferi BBK32 ингибирует классический путь, блокируя активацию комплекса комплемента C1», PLoS Pathogens, vol. 12, нет. 1
- ^ Jump up to: а б Фрейзер К.М., Касьенс С., Хуанг В.М. и др. (декабрь 1997 г.). «Геномная последовательность спирохеты болезни Лайма, B. burgdorferi» . Природа . 390 (6660): 580–6. Бибкод : 1997Natur.390..580F . дои : 10.1038/37551 . ПМИД 9403685 . S2CID 4388492 .
- ^ Касьенс С., Палмер Н., ван Вугт Р., Хуанг В.М., Стивенсон Б., Роза П., Латигра Р., Саттон Г., Петерсон Дж., Додсон Р.Дж., Хафт Д., Хики Э., Гвинн М., Уайт О., Фрейзер СМ (2000). «Бактериальный геном в движении: двенадцать линейных и девять кольцевых внехромосомных ДНК в инфекционном изоляте спирохеты B. burgdorferi, вызывающей болезнь Лайма ». Молекулярная микробиология . 35 (3): 490–516. дои : 10.1046/j.1365-2958.2000.01698.x . ПМИД 10672174 .
- ^ Jump up to: а б с д Тайзен, М.; Борре, М.; Мэтисен, MJ; Миккельсен, Б.; Лебех, AM; Хансен, К. (1 июня 1995 г.). «Эволюция белка внешней поверхности OspC Borrelia burgdorferi» . Журнал бактериологии . 177 (11): 3036–3044. дои : 10.1128/jb.177.11.3036-3044.1995 . ISSN 0021-9193 . ПМК 176990 . PMID 7768799 .
- ^ Эмберс, Моника Э.; Нарасимхан, Суканья (12 февраля 2013 г.). «Вакцинация против болезни Лайма: прошлое, настоящее и будущее» . Границы клеточной и инфекционной микробиологии . 3 :6. дои : 10.3389/fcimb.2013.00006 . ISSN 2235-2988 . ПМЦ 3569838 . ПМИД 23407755 .
- ^ Эггерс, Швейцария; Касьенс, С.; Хейс, Сан-Франциско; Гарон, CF; Дамман, CJ; Оливер, Д.Б.; Сэмюэлс, DS (октябрь 2000 г.). «Бактериофаги спирохет» . Журнал молекулярной микробиологии и биотехнологии . 2 (4): 365–373. ISSN 1464-1801 . ПМИД 11075907 .
- ^ Спирохеты: молекулярная и клеточная биология . Милтон Х. Сайер, Хорхе Гарсиа-Лара. Уаймондхэм: Horizon Scientific. 2001. ISBN 1-898486-27-1 . OCLC 47901873 .
{{cite book}}: CS1 maint: другие ( ссылка ) - ^ Эггерс, Кристиан Х.; Киммел, Бетси Дж.; Боно, Джеймс Л.; Элиас, Абдалла Ф.; Роза, Патрисия; Сэмюэлс, Д. Скотт (15 августа 2001 г.). «Трансдукция с помощью φBB-1, бактериофага Borrelia burgdorferi» . Журнал бактериологии . 183 (16): 4771–4778. дои : 10.1128/JB.183.16.4771-4778.2001 . ISSN 0021-9193 . ПМК 99531 . ПМИД 11466280 .
- ^ Куртенбах К., Ханинцова К., Цао Дж.И., Маргос Г., Фиш Д., Огден Н.Х. (2006). «Фундаментальные процессы в эволюционной экологии Лайм-боррелиоза» . Обзоры природы. Микробиология . 4 (9): 660–9. дои : 10.1038/nrmicro1475 . ПМИД 16894341 . S2CID 10877654 .
- ^ Jump up to: а б с д и Сэмюэлс, Д. Скотт (1 января 2010 г.). Боррелии: молекулярная биология, взаимодействие с хозяином и патогенез . Горизонт Научная Пресса. ISBN 9781904455585 .
Дальнейшее чтение
[ редактировать ]- Веласкес, Энкарна; Пейш, Альваро; Гомес-Алонсо, Альберто (март 2011 г.). «Микроорганизмы и рак: научные данные и новые гипотезы». Испанская хирургия (на испанском языке). 89 (3): 136–144. дои : 10.1016/j.ciresp.2010.08.006 . ПМИД 21292247 .
- Кочи, Юрай; Биста, Сандхья; Чирания, Паял; Ян, Сюли; Кицу, Хрисула; Рана, Випин Сингх; Яс, Озлем Буюктанир; Соненшайн, Дэниел Э.; Пал, Утпал (17 марта 2021 г.). «Антитела против EGF-подобных доменов в ортологах Ixodes scapularis BM86 влияют на питание клещей и выживаемость Borrelia burgdorferi» . Научные отчеты . 11 (1): 6095. Бибкод : 2021НатСР..11.6095К . дои : 10.1038/s41598-021-85624-5 . ПМЦ 7971074 . ПМИД 33731754 .
Внешние ссылки
[ редактировать ]| Базы данных органов управления : Национальные |
|---|