Трепонема бледная
| Трепонема бледная | |
|---|---|
 | |
| Научная классификация | |
| Домен: | Бактерии |
| Тип: | Спирохетата |
| Сорт: | Спирохета |
| Заказ: | Спирохеты |
| Семья: | Трепонемовые |
| Род: | Трепонема |
| Разновидность: | Т. паллидум |
| Биномиальное имя | |
| Трепонема бледная | |
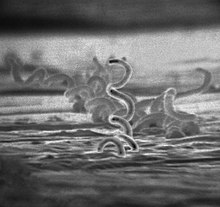
Treponema pallidum , ранее известная как Spirochaeta pallida , представляет собой микроаэрофильную -спирохету бактерию , подвид которой вызывает заболевания сифилис , беджель (также известный как эндемический сифилис) и фрамбезию . Известно, что он передается только среди людей. [1] и бабуины. [2] Это спирально свернутый микроорганизм обычно длиной 6–15 мкм и шириной 0,1–0,2 мкм. [1] T. pallidum как Отсутствие у цикла трикарбоновых кислот , так и процессов окислительного фосфорилирования приводит к минимальной метаболической активности. [3] Трепонемы имеют цитоплазматическую и наружную мембраны . При световой микроскопии трепонемы видны только при темнопольном освещении . T. pallidum состоит из трех подвидов: T. p. паллидум, Т. п. endemicum и Т. р. pertenue, каждый из которых имеет отдельное ассоциированное заболевание. [4]
Подвиды
[ редактировать ]три подвида : T. pallidum Известны [5]
- Бледная трепонема , вызывающая сифилис.
- Т. п. endemicum , вызывающий беджель или эндемический сифилис.
- Т. п. pertenue , вызывающий отклонение от курса
Три подвида, вызывающие фрамбезию , беджель и сифилис , морфологически и серологически неразличимы. [1] Эти бактерии первоначально были классифицированы как представители отдельных видов, но анализ гибридизации ДНК показывает, что они принадлежат к одному и тому же виду. Treponema carateum , возбудитель пинты , остается отдельным видом, поскольку изолята для анализа ДНК не существует. [6] Пути передачи заболевания у подвидов T. p. endemicum и Т. р. pertenue считается невенерическим. [7] Т. п. pallidum — наиболее инвазивный патогенный подвид, а T. carateum — наименее инвазивный из видов. Т. п. endemicum и Т. р. pertenue являются среднеинвазивными. [1]
Микробиология
[ редактировать ]Физиология
[ редактировать ]
Treponema pallidum — бактерия спиралевидной формы с высокой подвижностью, состоящая из наружной мембраны, слоя пептидогликана , внутренней мембраны , протоплазматического цилиндра и периплазматического пространства . [1] Его часто называют грамотрицательным , но в его внешней мембране отсутствует липополисахарид , который содержится во внешней мембране других грамотрицательных бактерий. [8] Он имеет эндофлагеллум (периплазматический жгутик), состоящий из четырех основных полипептидов , сердцевинной структуры и оболочки. [9] Жгутик расположен в периплазматическом пространстве и обертывает протоплазматический цилиндр. Наружная мембрана T. pallidum имеет наибольший контакт с клетками-хозяевами и содержит мало трансмембранных белков , ограничивающих антигенность , тогда как ее цитоплазматическая мембрана покрыта липопротеинами. [3] [10] Основная функция трепонемных лигандов внешней мембраны - прикрепление к клеткам-хозяевам с функциональным и антигенным родством между лигандами. [11] Род Treponema имеет ленты цитоскелетных цитоплазматических нитей, которые проходят по всей длине клетки прямо под цитоплазматической мембраной.
Внешняя мембрана
[ редактировать ]Наружная мембрана (ОМ) T. pallidum имеет несколько особенностей, которые исторически затрудняли ее исследование. К ним относятся такие детали, как низкое содержание белка, хрупкость и меньшее количество генных последовательностей, связанных с другими грамотрицательными внешними мембранами. [12] Прогресс был достигнут в использовании геномного секвенирования и передовых вычислительных моделей. Белки внешней мембраны трепонемы являются ключевыми факторами патогенеза, устойчивости и стратегии уклонения от иммунитета бактерий. Относительно низкое содержание белка препятствует распознаванию антигена иммунной системой, а существующие белки выступают из ОМ, обеспечивая его взаимодействие с хозяином. [12] Репутация трепонемы как «скрытого патогена» обусловлена, прежде всего, этой уникальной структурой ОМ, которая позволяет уклоняться от иммунного обнаружения. [12]
ТП0326
[ редактировать ]TP0326 — ортолог станка для сборки β-стволов Bam A. Аппарат BamA вставляет вновь синтезированные и экспортированные белки внешней мембраны во внешнюю мембрану. [13]
ТП0965
[ редактировать ]TP0965 представляет собой белок, который имеет решающее значение для слияния мембран у T. pallidum и расположен в периплазме . [14] TP0965 вызывает дисфункцию эндотелиального барьера, что является признаком поздней стадии патогенеза сифилиса. [15] Это происходит за счет снижения экспрессии белков плотных соединений, что, в свою очередь, увеличивает экспрессию молекул адгезии и проницаемость эндотелиальных клеток, что в конечном итоге приводит к разрушению эндотелиального слоя . [16]
ТП0453
[ редактировать ]TP0453 представляет собой белок из 287 аминокислот, связанный с внутренней мембраной внешней мембраны микроба. [14] У этого белка отсутствует обширная структура бета-листа , характерная для других мембранных белков, и он не пересекает внешнюю мембрану. [17] Предполагается, что функция этого белка связана с контролем потребления питательных веществ. [18]
ТП0624
[ редактировать ]Белки, содержащие домен белка А внешней мембраны (OmpA), необходимы для поддержания структурной целостности грамотрицательных бактерий. Эти домены содержат сайты связывания пептидогликана, которые создают «структурный мост между слоем пептидогликана и внешней мембраной». [19] Было высказано предположение, что белок TP0624, обнаруженный в T. pallidum, облегчает эту структурную связь, а также взаимодействие между белками внешней мембраны и соответствующими доменами на тонком слое пептидогликана . [19]
Семейство белков, повторяющих трепонему
[ редактировать ]Семейство белков повторов трепонемы (Tpr) представляет собой белки, экспрессируемые во время инфекционного процесса. Tprs образованы консервативным N-концевым доменом, аминоконцевым участком длиной около 50 аминокислот, центральной вариабельной областью и консервативным C-концевым доменом. [13] Многие различные типы Tpr включают TprA, TprB, TprC, TprD и TprE, но вариабельность TprK является наиболее актуальной из-за характеристик иммунного ускользания, которые она обеспечивает. [20]
Вариация антигена TprK регулируется конверсией генов. Таким образом, фрагменты семи вариабельных областей (V1–V7), присутствующих в TprK, и 53 донорских сайтов TprD могут быть объединены для получения новых структурированных последовательностей. [21] Вариации антигена TprK могут помочь T. pallidum избежать сильной иммунной реакции хозяина, а также сделать возможным повторное заражение людей. Это возможно, поскольку вновь структурированные белки могут избежать специфического распознавания антител. [20]
Чтобы внести больше фенотипического разнообразия, T. pallidum может подвергаться фазовым изменениям . Этот процесс в основном происходит в TprF, TprI, TprG, TprJ и TprL и состоит из обратимого расширения или сжатия полимерных повторов. Эти различия в размерах могут помочь бактерии быстро адаптироваться к микроокружению, уклониться от иммунного ответа или даже повысить сродство к хозяину. [21]
Культура
[ редактировать ]В прошлом столетии, с момента первого открытия, культивирование бактерий in vitro было затруднено. [22] Без возможности выращивать и поддерживать бактерии в лабораторных условиях открытия, касающиеся их метаболизма и антимикробной чувствительности, были значительно затруднены. [12] об успешном долгосрочном культивировании T. pallidum in vitro. { Однако в 2017 г. сообщалось [22] Это было достигнуто с использованием эпителиальных клеток Sf1Ep кроликов, которые были необходимым условием для дальнейшего размножения и выживания системы. [23] Использовалась среда TpCM-2, представляющая собой замену более простой среды, которая ранее обеспечивала рост культуры всего лишь в течение нескольких недель. [23] Этот успех стал результатом замены минимальной необходимой среды (MEM) на CMRL 1066, сложную среду для культуры тканей. [22] По мере развития могут произойти новые открытия о потребностях T. pallidum для роста и экспрессии генов, что, в свою очередь, приведет к исследованиям, полезным для лечения и профилактики сифилиса вне хозяина. [24] Однако постоянные попытки вырастить T. pallidum в аксенической культуре оказались безуспешными, что указывает на то, что он не удовлетворяет постулатам Коха . [25] Проблема, вероятно, связана с сильной адаптацией организма к проживанию в тканях млекопитающих, что приводит к уменьшению генома и значительным нарушениям метаболических и биосинтетических функций. [23]
Геном
[ редактировать ]Хромосомы вида T. pallidum небольшие, около 1,14 Мпн. Их последовательности ДНК идентичны более чем на 99,7%. [26] Было установлено, что около 92,9% ДНК представляют собой открытые рамки считывания , 55% из которых предсказывают биологические функции. [3] Геном T. pallidum был впервые секвенирован в 1998 году. [27] T. pallidum невозможно получить в чистой культуре, а это означает, что это секвенирование сыграло важную роль в заполнении пробелов в понимании функций микробов. Было обнаружено, что T. pallidum зависит от своего хозяина во многих молекулах, обычно образующихся в результате биосинтетических путей, и у него отсутствуют гены, ответственные за кодирование ключевых ферментов окислительного фосфорилирования и цикла трикарбоновых кислот. [28] Группа T. pallidum и ее уменьшенный геном, вероятно, являются результатом различных адаптаций, в результате которых она больше не обладает способностью синтезировать жирные кислоты, нуклеиновые кислоты и аминокислоты и вместо этого полагается на своих хозяев-млекопитающих в получении этих материалов. [24] Недавнее секвенирование геномов нескольких спирохет позволило тщательно проанализировать сходства и различия внутри этого бактериального типа и внутри вида. [29] [30] [31] T. pallidum имеет один из самых маленьких бактериальных геномов и ограниченные метаболические возможности, что отражает его адаптацию посредством редукции генома к богатой среде тканей млекопитающих. T. pallidum отличается спиральной, спиралевидной формой. [32] Чтобы избежать атаки антител, клетка имеет мало белков, находящихся на внешней мембранной оболочке. [33] Его хромосома имеет длину около 1000 пар оснований и имеет круглую форму со средним содержанием G + C 52,8%. [27] Секвенирование выявило набор из 12 белков и некоторые предполагаемые гемолизины, которые являются потенциальными факторами вирулентности T. pallidum. [34] Считается, что эти факторы вирулентности способствуют способности бактерии уклоняться от иммунной системы и вызывать заболевание. [34]
Клиническое значение
[ редактировать ]Клинические проявления сифилиса, фрамбезии и беджеля протекают в несколько стадий и поражают кожу. Поражения кожи, наблюдаемые на ранней стадии, сохраняются в течение недель или месяцев. Поражения кожи высокозаразны, а спирохеты в них передаются контактным путем. Поражения регрессируют по мере развития иммунного ответа против T. pallidum . Возникающая в результате латентная стадия во многих случаях может длиться всю жизнь. В некоторых случаях заболевание выходит из латентного периода и переходит в третичную фазу, в ходе которой возникают деструктивные поражения кожи, костей и хрящей. В отличие от фрамбезии и беджеля, сифилис на третичной стадии часто поражает также сердце, глаза и нервную систему. [6]
Сифилис
[ редактировать ]Treponema pallidum pallidum представляет собой подвижную спирохету, которая обычно передается при тесном половом контакте и попадает в организм хозяина через повреждения плоского или столбчатого эпителия . Возбудитель также может передаваться плоду трансплацентарным путем на более поздних стадиях беременности, вызывая врожденный сифилис. [35] Спиральная структура Т. р. pallidum позволяет ему двигаться штопором через слизистые оболочки или проникать в мельчайшие повреждения кожи. У женщин начальное поражение обычно локализуется на половых губах, стенках влагалища или шейке матки; у мужчин он находится на стержне или головке полового члена. [1] Он получает доступ к кровеносной и лимфатической системам хозяина через ткани и слизистые оболочки. В более тяжелых случаях он может получить доступ к хозяину, заразив кости скелета и центральную нервную систему организма. [1]
Инкубационный период Т. р. pallidum обычно длится около 21 дня, но может варьироваться от 10 до 90 дней. [36]
Лабораторная идентификация
[ редактировать ]
Treponema pallidum была впервые микроскопически обнаружена в сифилитических шанкрах Фрицем Шаудином и Эрихом Хоффманном в Шарите в Берлине в 1905 году. [37] Эту бактерию можно обнаружить с помощью специальных красителей, например, красителя Дитерле . T. pallidum также выявляется с помощью серологии , включая нетрепонемный VDRL , быстрый плазменный реагин , тесты на трепонемные антитела ( FTA-ABS ), реакцию иммобилизации T. pallidum и тест TPHA на сифилис . [38]
Уход
[ редактировать ]В начале 1940-х годов модели кроликов в сочетании с препаратом пенициллином позволяли проводить длительное лечение препаратом. Эти эксперименты заложили основу, которую современные ученые используют для терапии сифилиса. Пенициллин может ингибировать T. pallidum через 6–8 часов, хотя клетки все еще остаются в лимфатических узлах и регенерируют. Пенициллин — не единственный препарат, который можно использовать для ингибирования T. pallidum ; любые β-лактамные антибиотики или макролиды . можно использовать [39] Штамм T. pallidum 14 обладает встроенной устойчивостью к некоторым макролидам, включая эритромицин и азитромицин . Считается, что устойчивость к макролидам у штамма T. pallidum 14 обусловлена одноточечной мутацией, которая увеличила жизнеспособность организма. [40] Многие методы лечения сифилиса приводят только к бактериостатическим действия не используются более высокие концентрации пенициллина результатам, если только для бактерицидного . [39] [40] Пенициллин в целом является наиболее рекомендуемым антибиотиком Центрами по контролю за заболеваниями, поскольку он показывает наилучшие результаты при длительном применении. Он может ингибировать и даже уничтожать T. pallidum при низких и высоких дозах, причем каждое увеличение концентрации становится более эффективным. [40]
Вакцина
[ редактировать ]против сифилиса не По состоянию на 2024 год вакцины существует. Наружная мембрана T. pallidum имеет слишком мало поверхностных белков, чтобы антитела были эффективными. Усилия по разработке безопасной и эффективной вакцины против сифилиса были затруднены неопределенностью относительно относительной важности гуморальных и клеточных механизмов для защитного иммунитета. [41] и потому, что белки внешней мембраны T. pallidum не были однозначно идентифицированы. [42] [43] Напротив, некоторые из известных антигенов являются внутриклеточными, и антитела против них неэффективны для устранения инфекции. [44] [45] В прошлом веке было разработано несколько прототипов, и хотя ни один из них не обеспечивал защиты от инфекции, некоторые предотвращали распространение бактерий в дистальные органы и способствовали ускорению заживления. [46]
Ссылки
[ редактировать ]- ^ Jump up to: а б с д и ж г Радольф Дж.Д. (1996). Барон С. (ред.). Трепонема (4-е изд.). Галвестон (Техас): Медицинский филиал Техасского университета в Галвестоне. ISBN 978-0-9631172-1-2 . ПМИД 21413263 . НБК7716 . Проверено 13 февраля 2019 г.
- ^ Харпер К.Н., Фьюмагва Р.Д., Хоар Р., Вамбура П.Н., Коппенхавер Д.Х., Сапольски Р.М., Альбертс С.К., Тунг Дж., Роджерс Дж., Килево М., Батамузи Е.К., Леендерц Ф.Х., Армелагос Г.Дж., Кнауф С. (2012). « Инфекция Treponema pallidum у диких бабуинов Восточной Африки: распространение и генетическая характеристика ответственных штаммов» . ПЛОС ОДИН . 7 (12): e50882. Бибкод : 2012PLoSO...750882H . дои : 10.1371/journal.pone.0050882 . ПМЦ 3527465 . ПМИД 23284649 .
- ^ Jump up to: а б с Норрис С.Дж., Кокс Д.Л., генеральный директор Weinstock (2001). «Биология Treponema pallidum : корреляция функциональной активности с данными последовательности генома» (PDF) . Обзор JMMB . 3 (1): 37–62. ПМИД 11200228 .
- ^ Центурион-Лара А., Молини Б.Дж., Годорнес С., Сан Э., Хевнер К., Вурхис В.К., Люкхарт С.А. (1 сентября 2006 г.). «Молекулярная дифференциация подвидов Treponema pallidum » . Журнал клинической микробиологии . 44 (9): 3377–80. дои : 10.1128/JCM.00784-06 . ПМК 1594706 . ПМИД 16954278 .
- ^ Маркс М., Соломон А.В., Мэби, округ Колумбия (октябрь 2014 г.). «Эндемические трепонемные болезни» . Труды Королевского общества тропической медицины и гигиены . 108 (10): 601–7. дои : 10.1093/trstmh/tru128 . ПМК 4162659 . ПМИД 25157125 .
- ^ Jump up to: а б Джакани Л., Люкхарт С.А. (январь 2014 г.). «Эндемические трепонематозы» . Обзоры клинической микробиологии . 27 (1): 89–115. дои : 10.1128/CMR.00070-13 . ПМЦ 3910905 . ПМИД 24396138 .
- ^ «Другие инфекции, вызванные бледной трепонемой » . Здоровье иммигрантов и беженцев . Центры по контролю и профилактике заболеваний. 26 февраля 2019 года . Проверено 12 ноября 2019 г. .
- ^ Пилинг Р.В., Мэби Д., Камб М.Л., Чен XS, Радольф Дж.Д., Бензакен А.С. (октябрь 2017 г.). «Сифилис» . Обзоры природы. Праймеры по болезням . 3 : 17073. дои : 10.1038/nrdp.2017.73 . ПМК 5809176 . ПМИД 29022569 .
- ^ Сан-Мартин Ф, Фюле Л, Ираола Дж, Бускьяццо А, Пикардо М (1 марта 2023 г.). «Погружение в сложность спирохетального эндофлагеллума» . Тенденции в микробиологии . 31 (3): 294–307. дои : 10.1016/j.tim.2022.09.010 . ISSN 0966-842X . ПМИД 36244923 . S2CID 252916923 .
- ^ Лю Дж., Хауэлл Дж.К., Брэдли С.Д., Чжэн Ю., Чжоу Чж., Норрис С.Дж. (ноябрь 2010 г.). «Клеточная архитектура Treponema pallidum : новый жгутик, периплазматический конус и клеточная оболочка, выявленные с помощью криоэлектронной томографии» . Журнал молекулярной биологии . 403 (4): 546–61. дои : 10.1016/j.jmb.2010.09.020 . ПМЦ 2957517 . ПМИД 20850455 .
- ^ Альдерете Дж. Ф., игрок с низов Дж. Б. (декабрь 1980 г.). «Поверхностная характеристика вирулентной бледной трепонемы » . Инфекция и иммунитет . 30 (3): 814–823. дои : 10.1128/iai.30.3.814-823.1980 . ПМК 551388 . ПМИД 7014451 .
- ^ Jump up to: а б с д Радольф Дж.Д., Кумар С. (2018). « Наружная мембрана бледной трепонемы ». Биология спирохет: постгеномная эра . Актуальные темы микробиологии и иммунологии. Том. 415. стр. 1–38. дои : 10.1007/82_2017_44 . ISBN 978-3-319-89637-3 . ISSN 0070-217X . ПМЦ 5924592 . ПМИД 28849315 .
- ^ Jump up to: а б Хоули К.Л., Монтесума-Руска Дж.М., Дельгадо К.Н., Сингх Н., Уверски В.Н., Каймано М.Дж., Радольф Дж.Д., Лутра А. (8 июля 2021 г.). Гальперин М.Ю. (ред.). «Структурное моделирование репертуара белков внешней мембраны Treponema pallidum : дорожная карта для деконволюции патогенеза сифилиса и разработки вакцины против сифилиса» . Журнал бактериологии . 203 (15): e0008221. дои : 10.1128/JB.00082-21 . ПМЦ 8407342 . ПМИД 33972353 .
- ^ Jump up to: а б Чэнь Дж, Хуан Дж, Лю Цз, Се Ю (27 сентября 2022 г.). « Белки наружной мембраны Treponema pallidum : современное состояние и перспективы» . Патогены и болезни . 80 (1). doi : 10.1093/femspd/ftac023 . ПМИД 35869970 .
- ^ Маккевитт М., Бринкман М.Б., Маклафлин М., Перес С., Хауэлл Дж.К., Вайнсток Г.М., Норрис С.Дж., Палцкилл Т. (июль 2005 г.). «Идентификация антигенов Treponema pallidum в масштабе генома » . Инфекция и иммунитет . 73 (7): 4445–50. дои : 10.1128/iai.73.7.4445-4450.2005 . ПМЦ 1168556 . ПМИД 15972547 .
- ^ Чжан Р.Л., Чжан Дж.П., Ван QQ (16 декабря 2014 г.). «Рекомбинантный белок Treponema pallidum Tp0965 активирует эндотелиальные клетки и увеличивает проницаемость монослоя эндотелиальных клеток» . ПЛОС ОДИН . 9 (12): е115134. Бибкод : 2014PLoSO...9k5134Z . дои : 10.1371/journal.pone.0115134 . ПМЦ 4267829 . ПМИД 25514584 .
- ^ Хэзлетт К.Р., Кокс Д.Л., Декаффмейер М., Беннетт М.П., Дерозье Д.С., Ла Вак С.Дж., Ла Вак М.Э., Бурелл К.В., Робинсон Э.Дж., Брассер Р., Радольф Дж.Д. (сентябрь 2005 г.). «TP0453, скрытый белок внешней мембраны Treponema pallidum , повышает проницаемость мембраны» . Журнал бактериологии . 187 (18): 6499–6508. дои : 10.1128/JB.187.18.6499-6508.2005 . ПМЦ 1236642 . ПМИД 16159783 .
- ^ Лутра А., Жу Дж., Дерозье Д.С., Эггерс CH, Мулай В., Ананд А., МакАртур Ф.А., Романо Ф.Б., Каймано М.Дж., Хойк А.П., Малковски М.Г., Радольф Дж.Д. (2 декабря 2011 г.). «Переход от закрытой к открытой конформации Treponema pallidum липопротеина TP0453, связанного с внешней мембраной , включает мембранное зондирование и интеграцию с помощью двух амфипатических спиралей» . Журнал биологической химии . 286 (48): 41656–68. дои : 10.1074/jbc.M111.305284 . ISSN 1083-351X . ПМЦ 3308875 . ПМИД 21965687 .
- ^ Jump up to: а б Паркер М.Л., Хьюстон С., Уэзерелл К., Кэмерон С.Э., Буланже М.Дж. (10 ноября 2016 г.). «Структура Treponema pallidum Tp0624 демонстрирует модульную сборку дивергентно функционализированных и ранее нехарактерных доменов» . ПЛОС ОДИН . 11 (11): e0166274. Бибкод : 2016PLoSO..1166274P . дои : 10.1371/journal.pone.0166274 . ПМК 5104382 . ПМИД 27832149 .
- ^ Jump up to: а б Центурион-Лара А., Кастро С., Барретт Л., Кэмерон С., Мостоуфи М., Ван Вурхис В.К., Люкхарт С.А. (15 февраля 1999 г.). « Гомолог белка основной оболочки Treponema pallidum Tpr K является мишенью опсонических антител и защитного иммунного ответа» . Журнал экспериментальной медицины . 189 (4): 647–656. дои : 10.1084/jem.189.4.647 . ПМК 2192927 . ПМИД 9989979 .
- ^ Jump up to: а б Тан Ю, Чжоу Ю, Хэ Б, Цао Т, Чжоу Х, Нин Л, Чен Э, Ли Ю, Се Х, Пэн Б, Ху Ю, Лю С (19 октября 2022 г.). «Исследование механизма иммунного ускользания бледной трепонемы ». Инфекция . 51 (2): 305–321. дои : 10.1007/s15010-022-01939-z . ПМИД 36260281 . S2CID 252994863 .
- ^ Jump up to: а б с Эдмондсон Д.Г., Ху Б., Норрис С.Дж. (июнь 2018 г.). «Долговременное культивирование in vitro сифилиса спирохеты Treponema pallidum subsp. pallidum» . мБио . 9 (3). дои : 10.1128/mBio.01153-18 . ПМК 6020297 . ПМИД 29946052 .
- ^ Jump up to: а б с Эдмондсон Д.Г., ДеЛэй Б.Д., Ковис Л.Е., Норрис С.Дж. (23 февраля 2021 г.). «Параметры, влияющие на непрерывное культивирование штаммов Treponema pallidum in vitro » . мБио . 12 (1): 10.1128/mbio.03536–20. дои : 10.1128/mbio.03536-20 . ПМЦ 8545124 . ПМИД 33622721 .
- ^ Jump up to: а б Эдмондсон Д.Г., Норрис С.Дж. (февраль 2021 г.). «Культивирование in vitro сифилисной спирохеты Treponema pallidum » . Текущие протоколы . 1 (2): е44. дои : 10.1002/cpz1.44 . ПМЦ 7986111 . ПМИД 33599121 .
- ^ Прескотт Дж., Фельдманн Х., Сафронец Д. (январь 2017 г.). «Поправки к постулатам Коха о вирусных заболеваниях: когда «рост в чистой культуре» приводит к потере вирулентности» . Противовирусные исследования . 137 : 1–5. дои : 10.1016/j.antiviral.2016.11.002 . ПМК 5182102 . ПМИД 27832942 .
- ^ Шмайс Д., Струхаль М., Кнауф С. (июль 2018 г.). «Генетика некультивируемых трепонемных патогенов человека и животных». Инфекция, генетика и эволюция . 61 : 92–107. Бибкод : 2018InfGE..61...92S . дои : 10.1016/j.meegid.2018.03.015 . ПМИД 29578082 . S2CID 4826749 .
- ^ Jump up to: а б Фрейзер К.М., Норрис С.Дж., Вайнсток Г.М., Уайт О., Саттон Г.Г., Додсон Р. и др. (июль 1998 г.). «Полная последовательность генома бледной трепонемы , спирохеты сифилиса». Наука . 281 (5375): 375–88. Бибкод : 1998Sci...281..375F . дои : 10.1126/science.281.5375.375 . ПМИД 9665876 . S2CID 8641048 .
- ^ Уилли Дж. М. (2020). Микробиология Прескотта (11-е изд.). Макгроу-Хилл Образование. п. 436. ИСБН 978-1-260-21188-7 .
- ^ Зобаникова М, Миколка П, Чейкова Д, Поспишилова П, Чен Л, Струхал М, Цинь Х, Вайншток ГМ, Смайс Д (октябрь 2012 г.). «Полная последовательность генома штамма Treponema pallidum DAL-1» . Стандарты в геномных науках . 7 (1): 12–21. doi : 10.4056/sigs.2615838 . ПМЦ 3570794 . ПМИД 23449808 .
- ^ Тонг МЛ, Чжао Ц, Лю ЛЛ, Чжу XZ, Гао К, Чжан ХЛ, Линь ЛР, Ню Дж.Дж., Цзи ЗЛ, Ян ТК (2017). «Полная последовательность генома штамма Treponema pallidum subsp. pallidum Amoy: азиатский изолят, очень похожий на SS14» . ПЛОС ОДИН . 12 (8): e0182768. Бибкод : 2017PLoSO..1282768T . дои : 10.1371/journal.pone.0182768 . ПМЦ 5546693 . ПМИД 28787460 .
- ^ Сешадри Р., Майерс Г.С., Теттелин Х., Эйзен Дж.А., Гейдельберг Дж.Ф., Додсон Р.Дж. и др. (апрель 2004 г.). «Сравнение генома орального возбудителя Treponema denticola с геномами других спирохет» . Труды Национальной академии наук Соединенных Штатов Америки . 101 (15): 5646–51. Бибкод : 2004PNAS..101.5646S . дои : 10.1073/pnas.0307639101 . ПМЦ 397461 . ПМИД 15064399 .
- ^ Кларк Д.П., Данлэп П.В., Мэдиган Дж.Т., Мартинко Дж.М. (2009). Брок Биология микроорганизмов . Сан-Франциско: Пирсон. п. 79.
- ^ Вилли 2020 , с. 499
- ^ Jump up to: а б Генеральный директор Вайнсток, Хардхэм Дж. М., Маклеод, член парламента, Содергрен Э. Дж., Норрис С. Дж. (1 октября 1998 г.). «Геном бледной трепонемы : новый взгляд на возбудителя сифилиса» . Обзоры микробиологии FEMS . 22 (4): 323–332. дои : 10.1111/j.1574-6976.1998.tb00373.x . ПМИД 9862125 .
- ^ Арора Н., Садовский Ю., Дермоди Т.С., Койн CB (май 2017 г.). «Вертикальная передача микробов во время беременности» . Микроб-хозяин клетки . 21 (5): 561–7. дои : 10.1016/j.chom.2017.04.007 . ПМК 6148370 . ПМИД 28494237 .
- ^ «Факты о ЗППП - Сифилис (подробно)» . Центры по контролю заболеваний (CDC) . Проверено 19 апреля 2017 г.
- ^ Шаудинн Ф.Р. , Хоффманн Э. (1905). «Предварительный отчет о встречаемости спирохет в продуктах сифилитической болезни и папилломах » . Работа от Имперского управления здравоохранения . 22 :527-534.
- ^ Фишер Б., Харви Р.П., Чамп ПК (2007). Иллюстрированные обзоры Липпинкотта: Микробиология . Серия иллюстрированных обзоров Липпинкотта. Хагерстаун, Мэриленд: Липпинкотт Уильямс и Уилкинс. ISBN 978-0-7817-8215-9 .
- ^ Jump up to: а б Fantry LE, Tramont EC. «Бледная трепонема (сифилис)» . Инфекционные болезни и противомикробные средства . Проверено 12 ноября 2019 г. - через www.antimicrobe.org.
- ^ Jump up to: а б с Штамм Л.В. (1 февраля 2010 г.). «Глобальная проблема устойчивой к антибиотикам бледной трепонемы » . Антимикробные средства и химиотерапия . 54 (2): 583–9. дои : 10.1128/AAC.01095-09 . ПМК 2812177 . ПМИД 19805553 .
- ^ Епископ Н.Х., Миллер Дж.Н. (июль 1976 г.). «Гуморальный иммунитет при экспериментальном сифилисе. I. Проявление резистентности, вызываемой пассивной иммунизацией» . Журнал иммунологии . 117 (1): 191–6. doi : 10.4049/jimmunol.117.1.191 . ПМИД 778261 . S2CID 255333392 .
- ^ Томсон Флорида, Конли П.Г., Норгард М.В., Хэгман К.Э. (сентябрь 2007 г.). «Оценка воздействия на клеточную поверхность и вакциногенного потенциала Treponema pallidum белков-кандидатов внешней мембраны » . Микробы и инфекции . 9 (11): 1267–75. doi : 10.1016/j.micinf.2007.05.018 . ПМК 2112743 . ПМИД 17890130 .
- ^ Кэмерон CE, Lukehart SA (март 2014 г.). «Современное состояние разработки вакцины против сифилиса: необходимость, проблемы, перспективы» . Вакцина . 32 (14): 1602–9. doi : 10.1016/j.vaccine.2013.09.053 . ПМЦ 3951677 . ПМИД 24135571 .
- ^ Пенн К.В., Бейли М.Дж., Кокейн А. (апрель 1985 г.). «Аксиальный филаментный антиген Treponema pallidum » . Иммунология . 54 (4): 635–41. ПМЦ 1453562 . ПМИД 3884491 .
- ^ Норрис С.Дж. (сентябрь 1993 г.). «Полипептиды Treponema pallidum : прогресс в понимании их структурных, функциональных и иммунологических ролей. Исследовательская группа полипептидов Treponema Pallidum» . Микробиологические обзоры . 57 (3): 750–79. дои : 10.1128/MMBR.57.3.750-779.1993 . ПМЦ 372934 . ПМИД 8246847 .
- ^ Авила-Ньето К, Педреньо-Лопес Н, Митха О, Клоте Б, Бланко Х, Каррильо Х (2023). «Вакцина от сифилиса: проблемы, противоречия и возможности» . Границы в иммунологии . 14 : 1126170. дои : 10.3389/fimmu.2023.1126170 . ISSN 1664-3224 . ПМЦ 10118025 . ПМИД 37090699 .
Дальнейшее чтение
[ редактировать ]- Альтхаус Б.М., Эбер-Дюфрен Л. (октябрь 2014 г.). «Эпидемические циклы, обусловленные поведением хозяина» . Журнал Королевского общества, Интерфейс . 11 (99): 20140575. doi : 10.1098/rsif.2014.0575 . ПМЦ 4235258 . ПМИД 25100316 .
Внешние ссылки
[ редактировать ]- «Сифилис» (информационный бюллетень CDC). Центры по контролю и профилактике заболеваний. Май 2004 года.
| Базы данных органов управления : Национальные |
|---|