Эухроматин

Эухроматин (также называемый « открытым хроматином ») представляет собой слегка упакованную форму хроматина ( ДНК , РНК и белка ), которая обогащена генами и часто (но не всегда) находится в состоянии активной транскрипции . Эухроматин отличается от гетерохроматина , который плотно упакован и менее доступен для транскрипции. 92% генома человека является эухроматичным. [1]
У эукариот эухроматин составляет наиболее активную часть генома внутри клеточного ядра . У прокариот эухроматин является единственной присутствующей формой хроматина; это указывает на то, что структура гетерохроматина развилась позже вместе с ядром , возможно, как механизм, позволяющий справиться с увеличением размера генома.
Структура
[ редактировать ]Эухроматин состоит из повторяющихся субъединиц, известных как нуклеосомы , напоминающих развернутый набор бусинок на нити диаметром примерно 11 нм. [2] В основе этих нуклеосом лежит набор из четырех пар белков- гистонов : H3 , H4 , H2A и H2B . [2] Каждый коровый гистоновый белок имеет структуру «хвоста», которая может варьироваться по-разному; Считается, что эти вариации действуют как «главные переключатели управления» посредством различных состояний метилирования и ацетилирования , которые определяют общее расположение хроматина. [2] Вокруг октамеров гистонов намотано примерно 147 пар оснований ДНК , или чуть менее 2 витков спирали. [3] Нуклеосомы вдоль цепи связаны между собой посредством гистона H1 . [4] и короткий участок открытой линкерной ДНК в диапазоне от 0 до 80 пар оснований. Ключевое различие между структурой эухроматина и гетерохроматина состоит в том, что нуклеосомы в эухроматине расположены гораздо шире, что обеспечивает более легкий доступ различных белковых комплексов к цепи ДНК и, таким образом, увеличивает транскрипцию генов . [2]
Появление
[ редактировать ]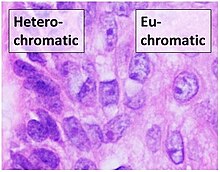
Эухроматин при большом увеличении напоминает набор бусинок на нитке. [2] Издалека он может напоминать клубок запутанных ниток, как, например, на некоторых изображениях, полученных с помощью электронного микроскопа . [5] Как при оптической, так и при электронной микроскопии эухроматин кажется более светлым, чем гетерохроматин , который также присутствует в ядре и выглядит темным. [6] - из-за менее компактной структуры. [5] При визуализации хромосом , например, на кариограмме , цитогенетическое кольцевание для окрашивания хромосом используется . Цитогенетическое кольцевание позволяет нам увидеть, какие части хромосомы состоят из эухроматина или гетерохроматина, чтобы дифференцировать хромосомные участки, нарушения или перестройки. [7] Одним из таких примеров является окрашивание G , иначе известное как окрашивание по Гимзе , при котором эухроматин кажется светлее, чем гетерохроматин. [8]
| Гимза (G-) Бандинг | Обратная (R-) полоса | Конститутивные полосы гетерохроматина (C-) | Квинакриновые (Q-) полосы | Теломерные полосы R (T-) | |
|---|---|---|---|---|---|
| Эухроматин | Зажигалка | Темнее | Зажигалка | Тупой | Свет |
| Гетерохроматин | Темнее | Зажигалка | Темнее | Яркий (флуоресцентный) | Темнее (Слабый) |
Функция
[ редактировать ]
Транскрипция
[ редактировать ]Эухроматин участвует в активной транскрипции ДНК продуктов до мРНК . Развернутая структура позволяет регуляторным белкам генов и комплексам РНК-полимеразы связываться с последовательностью ДНК, что затем может инициировать процесс транскрипции. [2] Хотя не весь эухроматин обязательно транскрибируется, поскольку эухроматин разделен на транскрипционно активные и неактивные домены. [13] эухроматин по-прежнему обычно связан с активной транскрипцией генов. Таким образом, существует прямая связь с тем, насколько активно продуктивна клетка и количеством эухроматина, который можно найти в ее ядре.
Считается, что клетка использует трансформацию из эухроматина в гетерохроматин как метод управления генов экспрессией и репликацией , поскольку на плотно уплотненном хроматине такие процессы ведут себя иначе. Это известно как «гипотеза доступности». [14] Одним из примеров конститутивного эухроматина, который «всегда включен», являются гены «домашнего хозяйства », которые кодируют белки, необходимые для основных функций выживания клеток. [15]
Эпигенетика
[ редактировать ]Эпигенетика предполагает изменения фенотипа , которые могут передаваться по наследству без изменения последовательности ДНК. Это может происходить посредством многих типов взаимодействий с окружающей средой. [16] Что касается эухроматина, посттрансляционные модификации гистонов могут изменить структуру хроматина, что приводит к изменению экспрессии генов без изменения ДНК. [17] Кроме того, было показано, что потеря гетерохроматина и увеличение эухроматина коррелируют с ускоренным процессом старения , особенно при заболеваниях, напоминающих преждевременное старение . [18] Исследования показали наличие эпигенетических маркеров на гистонах ряда дополнительных заболеваний. [19] [20]
Регулирование
[ редактировать ]Эухроматин в первую очередь регулируется посредством посттрансляционных модификаций его нуклеосом гистонов , осуществляемых многими ферментами, модифицирующими гистоны . хвостах гистонов Эти модификации происходят на N-концевых , которые выступают из структуры нуклеосомы, и, как полагают, привлекают ферменты для сохранения хроматина либо в открытой форме, как эухроматин, либо в закрытой форме, как гетерохроматин . [21] ацетилирование гистонов Например, обычно связано со структурой эухроматина, тогда как метилирование гистонов способствует ремоделированию гетерохроматина. [22] Ацетилирование делает группу гистонов более отрицательно заряженной, что, в свою очередь, нарушает ее взаимодействие с цепью ДНК, по сути «открывая» цепь для облегчения доступа. [21] Ацетилирование может происходить на нескольких остатках лизина хвоста гистона N-концевого и в различных гистонах одной и той же нуклеосомы, что, как полагают, еще больше увеличивает доступность ДНК для факторов транскрипции . [21]
Фосфорилирование гистонов — еще один метод регуляции эухроматина. [21] Обычно это происходит на N-концевых хвостах гистонов, однако некоторые участки присутствуют и в ядре. [21] Фосфорилирование контролируется киназами и фосфатазами , которые добавляют и удаляют фосфатные группы соответственно. Это может произойти при остатках серина , треонина или тирозина , присутствующих в эухроматине. [21] [22] Поскольку фосфатные группы, добавленные в структуру, будут иметь отрицательный заряд, это будет способствовать более расслабленной «открытой» форме, подобно ацетилированию. [22] Что касается функциональности, фосфорилирование гистонов участвует в экспрессии генов, восстановлении повреждений ДНК и ремоделировании хроматина . [22]
Другим методом регуляции, включающим отрицательный заряд и тем самым отдающим предпочтение «открытой» форме, является рибозилирование АДФ . [22] Этот процесс добавляет АДФ-рибозы к гистону одну или несколько единиц и участвует в пути реакции на повреждение ДНК . [22]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Международный консорциум по секвенированию генома человека (октябрь 2004 г.). «Завершение эухроматической последовательности генома человека» . Природа . 431 (7011): 931–945. Бибкод : 2004Natur.431..931H . дои : 10.1038/nature03001 . ПМИД 15496913 . S2CID 186242248 .
- ^ Перейти обратно: а б с д и ж г Бабу А., Верма Р.С. (январь 1987 г.). Борн Г.Х., Чон К.В., Фридлендер М. (ред.). «Структура хромосомы: эухроматин и гетерохроматин». Международный обзор цитологии . 108 . Академическая пресса: 1–60. дои : 10.1016/s0074-7696(08)61435-7 . ISBN 978-0-12-364508-1 . ПМИД 2822591 .
- ^ «Определение: нуклеосома/нуклеосомы» . Воспитательное природное образование . Проверено 6 октября 2021 г.
- ^ Mobley AS (январь 2019 г.). «Глава 4 - Индуцированные плюрипотентные стволовые клетки». В Mobley AS (ред.). Нейронные стволовые клетки и взрослый нейрогенез . Академическая пресса. стр. 67–94. ISBN 978-0-12-811014-0 .
- ^ Перейти обратно: а б «Клетка. 4. Ядро. Хроматин. Атлас гистологии растений и животных» . mmegias.webs.uvigo.es . Проверено 2 декабря 2021 г.
- ^ Енукашвили Н.И. (январь 2013 г.). «Глава вторая - Спутниковая ДНК млекопитающих: говорящий немой». В Донев Р., Пономарцев Н.В. (ред.). Достижения в области химии белков и структурной биологии . Организация хромосом. Том. 90. Академическая пресса. стр. 31–65. дои : 10.1016/B978-0-12-410523-2.00002-X . ISBN 978-0-12-410523-2 . ПМИД 23582201 .
- ^ Шен Ч. (январь 2019 г.). «Глава 13 – Молекулярная диагностика хромосомных заболеваний». Ин Шен Ч. (ред.). Диагностическая молекулярная биология . Академическая пресса. стр. 331–358. дои : 10.1016/B978-0-12-802823-0.00013-4 . ISBN 978-0-12-802823-0 . S2CID 131915096 .
- ^ Перейти обратно: а б «Полосы Гимзы» . Статьи, учебные пособия и онлайн-словари по биологии . 07.10.2019 . Проверено 2 декабря 2021 г.
- ^ «Обратные полосы — Определение и примеры — Биологический онлайн-словарь» . Статьи, учебные пособия и онлайн-словари по биологии . 18 сентября 2020 г. Проверено 2 декабря 2021 г.
- ^ «Конститутивные полосы гетерохроматина» . Статьи, учебные пособия и онлайн-словари по биологии . 07.10.2019 . Проверено 2 декабря 2021 г.
- ^ «Хинакриновый бандаж» . Статьи, учебные пособия и онлайн-словари по биологии . 07.10.2019 . Проверено 2 декабря 2021 г.
- ^ «Т-образный бандаж» . Статьи, учебные пособия и онлайн-словари по биологии . 07.10.2019 . Проверено 2 декабря 2021 г.
- ^ Вершуре П.Дж., ван дер Краан И., Мандерс Э.М., ван Дрил Р. (октябрь 1999 г.). «Пространственные взаимоотношения между сайтами транскрипции и территориями хромосом» . Журнал клеточной биологии . 147 (1): 13–24. дои : 10.1083/jcb.147.1.13 . ПМК 2164981 . ПМИД 10508851 .
- ^ Мюгге К. (1 апреля 2003 г.). «Модификации ядер и хвостов гистонов при рекомбинации V (D) J» . Геномная биология . 4 (4): 211. doi : 10.1186/gb-2003-4-4-211 . ПМК 154571 . ПМИД 12702201 .
- ^ Айзенберг Э., Леванон Э.Ю. (октябрь 2013 г.). «Человеческие гены домашнего хозяйства, еще раз». Тенденции в генетике . 29 (10): 569–574. дои : 10.1016/j.tig.2013.05.010 . ПМИД 23810203 .
- ^ Арни К.Л., Fisher AG (сентябрь 2004 г.). «Эпигенетические аспекты дифференцировки». Журнал клеточной науки . 117 (Часть 19): 4355–4363. дои : 10.1242/jcs.01390 . ПМИД 15331660 . S2CID 24376600 .
- ^ Сингх Н.П., Мадабхуши С.Р., Шривастава С., Сентилкумар Р., Нираджа С., Хосла С., Мишра Р.К. (май 2011 г.). «Эпигенетический профиль эухроматической области Y-хромосомы человека» . Исследования нуклеиновых кислот . 39 (9): 3594–3606. дои : 10.1093/nar/gkq1342 . ПМК 3089472 . ПМИД 21252296 .
- ^ Ван Дж, Цзя С.Т., Цзя С. (май 2016 г.). «Новый взгляд на регуляцию гетерохроматина» . Тенденции в генетике . 32 (5): 284–294. дои : 10.1016/j.tig.2016.02.005 . ПМЦ 4842111 . ПМИД 27005444 .
- ^ Симмонс Д. (2008). «Эпигенетические влияния и болезни» . Природное образование . 1 (1):6 . Проверено 2 декабря 2021 г.
- ^ Аласхар Альхамве Б., Халайла Р., Вольф Дж., фон Бюлов В., Харб Х., Алхамдан Ф. и др. (23 мая 2018 г.). «Модификации гистонов и их роль в эпигенетике атопических и аллергических заболеваний» . Аллергия, астма и клиническая иммунология . 14 (1): 39. дои : 10.1186/s13223-018-0259-4 . ПМЦ 5966915 . ПМИД 29796022 .
- ^ Перейти обратно: а б с д и ж Баннистер А.Дж., Кузаридес Т. (март 2011 г.). «Регуляция хроматина модификациями гистонов» . Клеточные исследования . 21 (3): 381–395. дои : 10.1038/cr.2011.22 . ПМК 3193420 . ПМИД 21321607 .
- ^ Перейти обратно: а б с д и ж Сингх Д., Ниши К., Хамбата К., Баласинор Н.Х. (январь 2020 г.). «Введение в эпигенетику: основные концепции и достижения в этой области». В Толлефсболе Т (ред.). Эпигенетика и репродуктивное здоровье . Трансляционная эпигенетика. Том. 21. Академическая пресса. стр. xxv – xliv. дои : 10.1016/B978-0-12-819753-0.02001-8 . ISBN 978-0-12-819753-0 . S2CID 235031860 .
Дальнейшее чтение
[ редактировать ]- Формирование гетерохроматина включает изменения модификаций гистонов на протяжении нескольких поколений клеток. Катан-Хайкович Ю., Струль К. (июнь 2005 г.). «Формирование гетерохроматина включает изменения модификаций гистонов на протяжении нескольких поколений клеток» . Журнал ЭМБО . 24 (12): 2138–2149. дои : 10.1038/sj.emboj.7600692 . ПМЦ 1150886 . ПМИД 15920479 .
- Chromatin Velocity выявляет эпигенетическую динамику путем профилирования одиночных клеток гетерохроматина и эухроматина. Тедеско М., Джаннезе Ф., Лазаревич Д., Джансанти В., Розано Д., Монзани С. и др. (октябрь 2021 г.). «Скорость хроматина раскрывает эпигенетическую динамику путем профилирования гетерохроматина и эухроматина в отдельных клетках». Природная биотехнология . 40 (2): 235–244. дои : 10.1038/s41587-021-01031-1 . HDL : 11368/3007419 . ПМИД 34635836 . S2CID 238637962 .
- Эпигенетическая наследственность и недостающая наследственность – Треротола М., Релли В., Симеоне П., Альберти С. (июль 2015 г.). «Эпигенетическая наследственность и недостающая наследственность» . Геномика человека . 9 (1): 17. дои : 10.1186/s40246-015-0041-3 . ПМК 4517414 . ПМИД 26216216 .
- Эпигенетические метки гистонов в гетерохроматине и эухроматине переносчика болезни Шагаса, Triatoma infestans – Альваренга Э.М., Родригес В.Л., Мораес А.С., Навес Л.С., Мондин М., Фелисбино М.Б., Мелло М.Л. (май 2016 г.). «Эпигенетические метки гистонов в гетерохроматине и эухроматине вектора болезни Шагаса, Triatoma infestans». Акта гистохимика . 118 (4): 401–412. дои : 10.1016/j.acthis.2016.04.002 . ПМИД 27079857 .