Система определения пола

Система определения пола — биологическая система, определяющая развитие половых признаков в организме . [1] Большинство организмов , создающих потомство путем полового размножения, имеют два общих пола и несколько менее распространенных интерсексуальных вариаций.
У некоторых видов встречаются гермафродиты . [2] Есть также некоторые виды, у которых временно или постоянно присутствует только один пол из-за партеногенеза , акта размножения самки без оплодотворения .
У некоторых видов определение пола является генетическим: самцы и самки имеют разные аллели или даже разные гены , определяющие их половую морфологию . У животных это часто сопровождается хромосомными различиями, обычно за счет комбинаций хромосом XY , ZW , XO , ZO или гаплодиплоидии . Половая дифференциация обычно запускается основным геном («половым локусом»), за которым следует множество других генов, создавая эффект домино .
В других случаях пол плода определяется переменными окружающей среды (например, температурой ). Детали некоторых систем определения пола еще не до конца изучены. Надежды [ ВОЗ? ] для будущего анализа биологической системы плода включают в себя инициализированные сигналы полной системы воспроизводства, которые можно измерить во время беременности, чтобы более точно определить, является ли определенный пол плода мужским или женским. [ нужна ссылка ] Такой анализ биологических систем может также указать, является ли плод гермафродитом, который включает в себя полностью или частично как мужские, так и женские органы воспроизводства.
Некоторые виды, такие как различные растения и рыбы, не имеют фиксированного пола, а вместо этого проходят жизненные циклы и меняют пол на основе генетических сигналов на соответствующих стадиях жизни своего типа. Это может быть связано с факторами окружающей среды, такими как время года и температура. У некоторых гонохорных видов некоторые особи могут иметь половые признаки обоих полов — состояние, называемое интерсекс . [3]
Хотя разнообразие систем определения пола характерно для различных биологических систем, системы, выходящие за рамки XY/XX/XO у млекопитающих, часто оставляют на более продвинутых курсах для тех, чьи исследования специализируются на генетике других организмов.
Открытие [ править ]
Этот раздел нуждается в расширении . Вы можете помочь, добавив к нему . ( июнь 2021 г. ) |
Определение пола было обнаружено у мучного червя американским генетиком Нетти Стивенс в 1903 году. [4] [5] [6]
В 1694 году Дж. Р. Камерариус провел первые эксперименты по опылению и сообщил о существовании мужских и женских признаков у растений (кукуруза).
В 1866 году Грегор Мендель опубликовал работу о наследовании генетических признаков. Это известно как менделевское наследование , и оно в конечном итоге установило современное понимание наследования от двух гамет .
В 1902 г. К.Э. МакКлюнг определил половые хромосомы у насекомых.
В 1917 г. К. Э. Аллен открыл механизмы определения пола у растений.
В 1922 году С. Б. Бриджес выдвинул теорию генного баланса определения пола.
Хромосомные системы [ править ]
/ Половые XY хромосомы XX


Система определения пола XX/XY является наиболее распространенной, поскольку она встречается у людей. Система XX/XY встречается у большинства других млекопитающих , а также у некоторых насекомых. В этой системе у женщин есть две половые хромосомы одного типа (XX), а у мужчин - две разные половые хромосомы (XY). Половые хромосомы X и Y отличаются друг от друга по форме и размеру, в отличие от остальных хромосом ( аутосом ), и иногда их называют аллосомами . У некоторых видов, например у человека, организмы какое-то время во время развития остаются безразличными к полу ( эмбриогенез ); однако у других, например у плодовых мух, половая дифференциация происходит сразу после оплодотворения яйцеклетки. [7]
пола по Y Определение - центру
имеется ген SRY У некоторых видов (включая человека) на Y-хромосоме , определяющий мужскую принадлежность . Представители видов, зависящих от SRY, могут иметь необычные хромосомные комбинации XY, такие как XXY , и все еще жить. [7] Пол человека определяется наличием или отсутствием Y-хромосомы с функциональным геном SRY. После активации гена SRY клетки вырабатывают тестостерон и антимюллеров гормон , которые обычно обеспечивают развитие единственной мужской репродуктивной системы. [7] У типичных XX эмбрионов клетки секретируют эстроген , который направляет организм по женскому пути.
При Y-центрированном определении пола ген SRY является основным геном, определяющим мужские характеристики, но для развития семенников требуется несколько генов. У мышей XY отсутствие гена DAX1 на Х-хромосоме приводит к бесплодию, а у человека - к врожденной гипоплазии надпочечников . [8] Однако, когда дополнительный ген DAX1 помещается в Х-хромосому, в результате появляется женщина, несмотря на существование SRY, поскольку он подавляет эффекты SRY. [9] Даже если у XX женщин имеются нормальные половые хромосомы, дупликация или экспрессия SOX9 вызывает развитие семенников. [10] [11] Постепенное изменение пола гена FOXL2 у самок. у развитых мышей также может происходить при удалении [12] Несмотря на то, что ген DMRT1 используется птицами в качестве полового локуса, виды, имеющие XY-хромосомы, также полагаются на DMRT1, содержащийся в хромосоме 9, для половой дифференциации на определенном этапе своего формирования. [7]
Определение пола по Х-центру [ править ]
Некоторые виды, например плодовые мухи , используют наличие двух Х-хромосом для определения женской принадлежности . [13] Виды, которые используют количество X для определения пола, нежизнеспособны с дополнительной Х-хромосомой.
Другие варианты определения пола XX / XY
У некоторых рыб есть варианты системы определения пола XY , а также обычная система. Например, Xiphophorus nezahualcoyotl и X. milleri, имея формат XY , также имеют вторую Y-хромосому, известную как Y', которая создает самок XY' и самцов YY'. [14]
По крайней мере, одна однопроходная , утконос , представляет собой особую схему определения пола, которая в некотором смысле напоминает схему половых хромосом ZW птиц и лишена гена SRY. У утконоса ; десять половых хромосом мужчины имеют образец XYXYXYXYXY, а женщины имеют десять X-хромосом. Хотя это система XY, половые хромосомы утконоса не имеют гомологов с половыми хромосомами платипуса. [15] Вместо этого гомологи с половыми хромосомами эутериев лежат на хромосоме 6 уттерипа, а это означает, что половые хромосомы эутериев были аутосомами в то время, когда однопроходные отделились от терианских млекопитающих (сумчатых и плаунообразных). Однако гомологи птичьего гена DMRT1 на половых хромосомах X3 и X5 утконоса позволяют предположить, что, возможно, ген, определяющий пол у утконоса, является тем же самым, который участвует в определении пола птиц. Необходимо провести дополнительные исследования, чтобы определить точный ген, определяющий пол утконоса. [16]

/ X0 Половые хромосомы XX
В этом варианте системы XY у женщин есть две копии половой хромосомы (XX), а у мужчин — только одна (X0). 0 . означает отсутствие второй половой хромосомы Обычно в этом методе пол определяется количеством генов, выраженных в двух хромосомах. Эта система наблюдается у ряда насекомых, в том числе у кузнечиков и сверчков отряда прямокрылых и у тараканов (отряда Blattodea ). У небольшого числа млекопитающих также отсутствует Y-хромосома. К ним относятся колючая крыса Амами ( Tokudaia osimensis ) и колючая крыса Токуносима ( Tokudaia tokunoshimensis ) и Sorex araneus , вид землероек . Закавказские слепушонки ( Ellobius lutescens ) также обладают формой детерминации ХО, при которой у представителей обоих полов отсутствует вторая половая хромосома. [9] Механизм определения пола еще не изучен. [17]
Нематода ; C. elegans — самец с одной половой хромосомой (X0) с парой хромосом (ХХ) — гермафродит. [18] Его основным половым геном является XOL, который кодирует XOL-1 , а также контролирует экспрессию генов TRA-2 и HER-1. Эти гены уменьшают активацию мужских генов и увеличивают ее соответственно. [19]
/ Половые хромосомы ZW ZZ
Система определения пола ZW встречается у птиц, некоторых рептилий, некоторых насекомых и других организмов. Система определения пола ZW перевернута по сравнению с системой XY: у женщин есть два разных типа хромосом (ZW), а у мужчин - две хромосомы одного и того же типа (ZZ). Было обнаружено, что у кур это зависит от экспрессии DMRT1. [20] У птиц гены FET1 и ASW обнаружены на W-хромосоме самок, аналогично тому, как Y-хромосома содержит SRY. [7] Однако не все виды зависят от W в определении пола. Например, есть мотыльки и бабочки с ZW, но у некоторых были обнаружены самки с ZO, а также самки с ZZW. [18] Кроме того, хотя млекопитающие деактивируют одну из своих дополнительных Х-хромосом у самок, оказывается, что в случае чешуекрылых самцы производят вдвое больше нормального количества ферментов из-за наличия двух Z. [18] Поскольку использование определения пола ZW различно, до сих пор неизвестно, как именно большинство видов определяют свой пол. [18] Однако, как сообщается, шелкопряд Bombyx mori использует одну специфичную для самок пиРНК в качестве основного определения пола. [21] Несмотря на сходство систем ZW и XY, эти половые хромосомы развивались раздельно. В случае с курицей их Z-хромосома больше похожа на аутосому 9 человека. [22] Z-хромосома курицы также, по-видимому, связана с X-хромосомой утконоса. [23] Когда вид ZW, такой как варан Комодо , размножается партеногенетически , обычно производятся только самцы. Это связано с тем, что гаплоидные яйца удваивают свои хромосомы, в результате чего получается ZZ или WW. ЗЗ становятся самцами, а ВВ нежизнеспособны и не доводятся до срока. [24]
В системах определения пола как XY, так и ZW половая хромосома, несущая критические факторы, часто значительно меньше и несет немногим больше генов, необходимых для запуска развития данного пола. [25] [ нужен лучший источник ]
ZZ/ хромосомы Половые Z0
Система определения пола ZZ/Z0 встречается у некоторых бабочек. У этих насекомых одна половая хромосома Z. У самцов две Z-хромосомы, а у самок одна Z. У самцов ZZ, у самок Z0. [26] [27] [28]
хромосомы половые УФ
У некоторых видов мохообразных и некоторых водорослей видов гаметофитная стадия жизненного цикла не является гермафродитной, а возникает в виде отдельных мужских или женских особей, которые производят мужские и женские гаметы соответственно. Когда мейоз происходит в поколении спорофитов жизненного цикла, половые хромосомы, известные как U и V, сортируются в споры, которые несут либо U-хромосому и дают начало женским гаметофитам, либо V-хромосому и дают начало мужским гаметофитам. [29] [30]
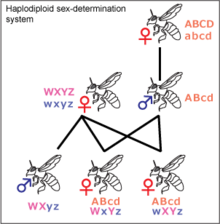
Гаплодиплоидия [ править ]
Гаплодиплоидия обнаружена у насекомых, принадлежащих к перепончатокрылым , таких как муравьи и пчелы . Определение пола контролируется зиготностью дополнительного локуса определения пола ( csd ). Из неоплодотворенных яиц развиваются гаплоидные особи, которые имеют единственную гемизиготную копию локуса csd и, следовательно, являются самцами. Из оплодотворенных яиц развиваются диплоидные особи, которые из-за высокой изменчивости локуса csd обычно представляют собой гетерозиготных самок. В редких случаях диплоидные особи могут быть гомозиготными, из них развиваются стерильные самцы.Ген, действующий как локус csd, был идентифицирован у медоносных пчел , и несколько генов-кандидатов были предложены в качестве локуса csd для других перепончатокрылых. [31] [32] [33] Большинство самок отряда перепончатокрылых могут определять пол своего потомства, удерживая полученную сперму в своей сперматеке и либо выпуская ее в яйцевод, либо нет. Это позволяет им создавать больше рабочих, в зависимости от статуса колонии. [34]
Другие хромосомные системы [ править ]
Другие необычные системы включают системы зеленого меченосца (полифакториальная система с генами, определяющими пол, в нескольких хромосомах). [14] Chironomus мошки [ нужны разъяснения ] [ нужна ссылка ] ; ювенильный гермафродитизм рыбок данио с неизвестным возбудителем; [14] и утконос , имеющий W, X и Y-хромосомы. Это позволяет женщинам WY, WX или XX и мужчинам YY или XY. [14]
Тип спаривания у микроорганизмов аналогичен полу у многоклеточных организмов и иногда описывается с использованием этих терминов, хотя они не обязательно коррелируют с физическими структурами тела. Некоторые виды имеют более двух типов спаривания. Тетрахимена , разновидность инфузорий , имеет семь типов спаривания. Schizophyllum commune , разновидность гриба, насчитывает 23 328.
Экологические системы
Зависит от температуры [ править ]

Существует множество других систем определения пола. У некоторых видов рептилий, включая аллигаторов , некоторых черепах и туатару , пол определяется температурой, при которой яйцо инкубируется в период, чувствительный к температуре. Примеров температурно-зависимого определения пола (ТЗД) у птиц нет. мегаподов наблюдается это явление, но оказалось, что на самом деле у них разные уровни смертности эмбрионов в зависимости от температуры для каждого пола. Раньше считалось, что у [35] У некоторых видов с TSD определение пола достигается воздействием более высоких температур, в результате чего потомство оказывается одного пола, а более низкие температуры приводят к другому полу. Этот тип TSD называется Pattern I. Для других видов, использующих TSD, воздействие обеих крайних температур приводит к появлению потомства одного пола, а воздействие умеренных температур приводит к появлению потомков противоположного пола, что называется модели II TSD . Конкретные температуры, необходимые для образования каждого пола, известны как температура, способствующая развитию самок, и температура, способствующая развитию самцов. [36] Когда температура остается вблизи порога в течение периода чувствительности к температуре, соотношение полов варьируется между двумя полами. [37] Температурные стандарты некоторых видов основаны на том, когда создается тот или иной фермент. Эти виды, у которых определение пола зависит от температуры, не имеют гена SRY , но имеют другие гены, такие как DAX1 , DMRT1 и SOX9 , которые экспрессируются или не экспрессируются в зависимости от температуры. [36] Пол некоторых видов, таких как нильская тилапия , австралийский сцинк и австралийская ящерица-дракон , имеет первоначальную предвзятость, задаваемую хромосомами, но позже может быть изменен под действием температуры инкубации. [14]
Неизвестно, как именно эволюционировало определение пола в зависимости от температуры. [38] Это могло произойти благодаря тому, что определенные полы стали более приспособлены к определенным местам, соответствующим температурным требованиям. Например, более теплая территория может быть более подходящей для гнездования, поэтому будет произведено больше самок, чтобы увеличить количество гнездящихся в следующем сезоне. [38] У амниот детерминация пола в окружающей среде предшествовала генетически детерминированным системам птиц и млекопитающих; Считается, что температурозависимые амниоты были общим предком амниот с половыми хромосомами. [39]
экологические Другие системы
Существуют и другие системы определения пола в окружающей среде, в том числе системы определения местоположения, зависящие от местоположения, как это видно у морского червя Bonellia viridis : личинки становятся самцами, если они вступают в физический контакт с самкой, и самками, если они оказываются на голом морском дне. Это вызвано присутствием химического вещества, вырабатываемого самками, бонеллина . [40] Некоторые виды, например улитки , практикуют смену пола : взрослые особи сначала становятся самцами, а затем становятся самками. У тропических рыб-клоунов доминирующей особью в группе становится самка, а остальные — самцы, а у синеголовых губанов ( Thalassoma bifasciatum ) — наоборот. Однако у некоторых видов нет системы определения пола. К гермафродитным видам относятся обыкновенный дождевой червь и некоторые виды улиток. Некоторые виды рыб, рептилий и насекомых размножаются партеногенезом и вообще являются самками. Есть некоторые рептилии, такие как удав и варан Комодо , которые могут размножаться как половым, так и бесполым путем, в зависимости от наличия партнера. [41]
Эволюция [ править ]
Системы определения пола, возможно, произошли от типа спаривания , который является особенностью микроорганизмов .
Хромосомное определение пола, возможно, возникло на ранних этапах истории эукариот. [42] Но у растений было высказано предположение, что он развился недавно. [43]
Принятая гипотеза эволюции половых хромосом XY и ZW у амниот состоит в том, что они развивались одновременно, в двух разных ветвях. [44] [45]
Никакие гены не являются общими между хромосомами ZW птиц и XY млекопитающих. [46] а Z-хромосома курицы похожа на аутосомную хромосому 9 человека, а не на X или Y. Это предполагает не то, что системы определения пола ZW и XY имеют общее происхождение, а то, что половые хромосомы произошли от аутосомных хромосом предка общего птицы и млекопитающие. У утконоса , однопроходного , хромосома X1 имеет гомологию с терианскими млекопитающими, тогда как хромосома X5 содержит птичий ген определения пола, что еще раз указывает на эволюционную связь. [47]
Однако есть некоторые свидетельства, позволяющие предположить, что могли быть переходы между ZW и XY, например, у Xiphophorus maculatus , у которых есть системы ZW и XY в одной и той же популяции, несмотря на то, что ZW и XY имеют разные местоположения генов. [48] [49] Недавняя теоретическая модель открывает возможность как перехода между системами XY/XX и ZZ/ZW, так и определения пола в окружающей среде. [50] Гены утконоса также подтверждают возможную эволюционную связь между XY и ZW, поскольку в их Х-хромосомах имеется ген DMRT1 , которым обладают птицы. [51] Тем не менее, XY и ZW идут по схожему маршруту. Все половые хромосомы возникли как исходные аутосомы исходного амниота, пол потомства которых определялся по температуре. После разделения млекопитающих ветвь рептилий далее разделилась на Lepidosauria и Archosauromorpha . Обе эти две группы развили систему ZW отдельно, о чем свидетельствует существование разных локализаций половых хромосом. [45] У млекопитающих одна из пар аутосом, теперь Y, мутировала свой ген SOX3 в ген SRY , в результате чего эта хромосома стала обозначать пол. [45] [51] [52] После этой мутации хромосома, содержащая SRY, инвертировалась и больше не была полностью гомологичной своему партнеру. Области X и Y-хромосом , которые все еще гомологичны друг другу, известны как псевдоаутосомные области . [53] Как только Y-хромосома инвертировалась, она стала неспособна лечить вредные мутации и, таким образом, деградировала . [45] Есть некоторые опасения, что Y-хромосома еще больше сократится и перестанет функционировать через десять миллионов лет: но Y-хромосома строго законсервирована после первоначальной быстрой потери генов. [54] [55]
Есть некоторые виды позвоночных, такие как рыба медака , у которых половые хромосомы развились отдельно; их Y-хромосома никогда не инвертируется и все еще может обмениваться генами с X. Половые хромосомы этих видов относительно примитивны и неспециализированы. Поскольку Y не имеет генов, специфичных для самцов, и может взаимодействовать с X, могут образоваться самки XY и YY, а также самцы XX. [14] Неинвертированные Y-хромосомы с долгой историей обнаружены у питонов и эму , возраст каждой системы превышает 120 миллионов лет, что позволяет предположить, что инверсии не обязательно являются случайностью. [56] Определение пола XO может развиться из определения пола XY примерно через 2 миллиона лет. [ нужны разъяснения ] [57]
См. также [ править ]
- Кларенс Эрвин МакКлунг , открывший роль хромосом в определении пола.
- Фактор, определяющий яички
- Влияние матери на определение пола
- Последовательный гермафродитизм
- Определение и дифференциация пола (человек)
Ссылки [ править ]
- ^ Шнебли, Риса Ария (2021). «Определение пола у человека» . Энциклопедия проекта «Эмбрион» . Проверено 6 июля 2022 г.
- ^ Розенфилд, К.А. (2018). «Гермафродит». В Вонк Дж, Шакелфорд Т. (ред.). Энциклопедия познания и поведения животных . Чам: Международное издательство Springer. стр. 1–2. дои : 10.1007/978-3-319-47829-6_329-1 . ISBN 978-3-319-47829-6 .
- ^ Минелли А. , Фуско Дж. (2019). Биология размножения . Издательство Кембриджского университета. стр. 116–117. ISBN 978-1108499859 . Архивировано из оригинала 11 октября 2020 года . Проверено 11 октября 2020 г.
- ^ «Нетти Стивенс: первооткрывательница половых хромосом | Изучайте науку в Scitable» . www.nature.com . Архивировано из оригинала 7 апреля 2019 года . Проверено 7 июня 2018 г.
- ^ Огилви М.Б., Чокетт СиДжей (август 1981 г.). «Нетти Мария Стивенс (1861–1912): ее жизнь и вклад в цитогенетику». Труды Американского философского общества . 125 (4): 292–311. JSTOR 986332 . ПМИД 11620765 .
- ^ «Нетти Мария Стивенс (1861–1912) | Энциклопедия проекта «Эмбрион»» . «эмбрион.asu.edu ». Архивировано из оригинала 8 апреля 2019 года . Проверено 7 июня 2018 г.
- ^ Jump up to: Перейти обратно: а б с д и Хейк Л. (2008). «Генетические механизмы определения пола» . Природное образование . 1 (1). Архивировано из оригинала 19 августа 2017 года . Проверено 8 декабря 2011 г.
- ^ Гудфеллоу П.Н., Камерино Дж. (июнь 1999 г.). «DAX-1, ген «антитестис»» . Клеточные и молекулярные науки о жизни . 55 (6–7): 857–863. дои : 10.1007/PL00013201 . ПМЦ 11147076 . ПМИД 10412368 . S2CID 19764423 .
- ^ Jump up to: Перейти обратно: а б Чандра, HS (25 апреля 1999 г.). «Другой способ взглянуть на загадку определения пола у Ellobius lutescens». Современная наука . 76 (8): 1072.
- ^ Кокс Дж.Дж., Уиллатт Л., Хомфрей Т., Вудс К.Г. (январь 2011 г.). «Дупликация SOX9 и семейное нарушение развития яичек 46,XX» . Медицинский журнал Новой Англии . 364 (1): 91–93. дои : 10.1056/NEJMc1010311 . ПМИД 21208124 .
- ^ Хуан Б., Ван С., Нин Ю., Лэмб А.Н., Бартли Дж. (декабрь 1999 г.). «Аутосомное изменение пола XX, вызванное дупликацией SOX9». Американский журнал медицинской генетики . 87 (4): 349–353. doi : 10.1002/(SICI)1096-8628(19991203)87:4<349::AID-AJMG13>3.0.CO;2-N . PMID 10588843 .
- ^ Уленхаут Н.Х., Якоб С., Анлаг К., Айзенбергер Т., Секидо Р., Кресс Дж. и др. (декабрь 2009 г.). «Соматическое половое перепрограммирование яичников взрослых в семенники путем абляции FOXL2» . Клетка . 139 (6): 1130–1142. дои : 10.1016/j.cell.2009.11.021 . ПМИД 20005806 .
- ^ Пенальва ЛО, Санчес Л (сентябрь 2003 г.). «РНК-связывающий секс-летальный белок (Sxl) и контроль определения пола дрозофилы и компенсация дозы» . Обзоры микробиологии и молекулярной биологии . 67 (3): 343–59, оглавление. дои : 10.1128/MMBR.67.3.343-359.2003 . ЧВК 193869 . ПМИД 12966139 .
- ^ Jump up to: Перейти обратно: а б с д и ж Шартл М. (июль 2004 г.). «Сравнительный взгляд на определение пола в Медаке» . Механизмы развития . 121 (7–8): 639–645. дои : 10.1016/j.mod.2004.03.001 . ПМИД 15210173 . S2CID 17401686 .
- ^ Уоррен В.К., Хиллиер Л.В., Маршалл Грейвс Дж.А., Бирни Э., Понтинг К.П., Грюцнер Ф. и др. (май 2008 г.). «Анализ генома утконоса выявил уникальные признаки эволюции» . Природа . 453 (7192): 175–183. Бибкод : 2008Natur.453..175W . дои : 10.1038/nature06936 . ПМК 2803040 . ПМИД 18464734 .
- ^ Грюцнер Ф., Эшли Т., Роуэлл Д.М., Маршалл Грейвс Дж.А. (апрель 2006 г.). «Как утконос получил цепочку половых хромосом? Сравнение мейотических множественных чисел и половых хромосом у растений и животных». Хромосома . 115 (2): 75–88. дои : 10.1007/s00412-005-0034-4 . ПМИД 16344965 . S2CID 23603889 .
- ^ Куроива А., Ханда С., Нисияма С., Чиба Е., Ямада Ф., Абэ С., Мацуда Ю. (июль 2011 г.). «Дополнительные копии CBX2 в геномах самцов млекопитающих, лишенных SRY, колючей крысы Амами (Tokudaia osimensis) и колючей крысы Токуносима (Tokudaia tokunoshimensis)». Хромосомные исследования . 19 (5): 635–644. дои : 10.1007/s10577-011-9223-6 . ПМИД 21656076 . S2CID 23311263 .
- ^ Jump up to: Перейти обратно: а б с д Маджерус М.Э. (2003). Сексуальные войны: гены, бактерии и предвзятое соотношение полов . Издательство Принстонского университета. п. 250. ИСБН 978-0-691-00981-0 . Проверено 4 ноября 2011 г.
- ^ Кувабара П.Е., Оккема П.Г., Кимбл Дж. (апрель 1992 г.). «tra-2 кодирует мембранный белок и может опосредовать клеточную связь в пути определения пола Caenorhabditis elegans» . Молекулярная биология клетки . 3 (4): 461–473. дои : 10.1091/mbc.3.4.461 . ПМК 275596 . ПМИД 1498366 .
- ^ Смит К.А., Рёслер К.Н., Онесорг Т., Камминс Д.М., Фарли П.Г., Доран Т.Дж., Синклер А.Х. (сентябрь 2009 г.). «Птичий Z-связанный ген DMRT1 необходим для определения мужского пола у кур». Природа . 461 (7261): 267–271. Бибкод : 2009Natur.461..267S . дои : 10.1038/nature08298 . ПМИД 19710650 . S2CID 4413389 .
- ^ Киучи Т., Кога Х., Кавамото М., Сёдзи К., Сакаи Х., Араи Ю. и др. (май 2014 г.). «Одна специфичная для самки пиРНК является основным определяющим фактором пола тутового шелкопряда». Природа . 509 (7502): 633–636. Бибкод : 2014Natur.509..633K . дои : 10.1038/nature13315 . ПМИД 24828047 . S2CID 205238635 .
- ^ Стиглек Р., Эзаз Т., Грейвс Дж. А. (2007). «Новый взгляд на эволюцию половых хромосом птиц». Цитогенетические и геномные исследования . 117 (1–4): 103–109. дои : 10.1159/000103170 . ПМИД 17675850 . S2CID 12932564 .
- ^ Грюцнер Ф., Ренс В., Ценд-Аюш Э., Эль-Могарбель Н., О'Брайен ПК, Джонс Р.К. и др. (декабрь 2004 г.). «У утконоса мейотическая цепь из десяти половых хромосом разделяет гены с Z-хромосомами птиц и X-хромосомами млекопитающих». Природа . 432 (7019): 913–917. Бибкод : 2004Natur.432..913G . дои : 10.1038/nature03021 . ПМИД 15502814 . S2CID 4379897 .
- ^ «Непорочное зачатие для гигантских ящериц» . Новости Би-би-си . 20 декабря 2006 г. Архивировано из оригинала 4 ноября 2014 г. . Проверено 13 марта 2008 г.
- ^ «Эволюция Y-хромосомы» . Анненберг Медиа . Архивировано из оригинала 4 ноября 2004 года . Проверено 1 апреля 2008 г.
- ^ Траут В., Сахара К., Марек Ф. (2007). «Половые хромосомы и определение пола у чешуекрылых». Сексуальное развитие . 1 (6): 332–346. дои : 10.1159/000111765 . ПМИД 18391545 . S2CID 6885122 .
- ^ «Генетические механизмы определения пола — изучайте науку в Scitable» . www.nature.com . Архивировано из оригинала 19 августа 2017 года . Проверено 8 декабря 2011 г.
- ^ Справочник по зоологии . Вальтер де Грюйтер. 1925. ISBN 9783110162103 – через Google Книги.
- ^ Бахтрог Д., Киркпатрик М., Мэнк Дж.Э., МакДэниел С.Ф., Пирес Дж.К., Райс В., Валенсуэла Н. (сентябрь 2011 г.). «Все ли половые хромосомы созданы равными?». Тенденции в генетике . 27 (9): 350–357. дои : 10.1016/j.tig.2011.05.005 . ПМИД 21962970 .
- ^ Реннер, СС; Генрихс, Дж.; Соуза, А. (2017). «Половые хромосомы мохообразных: последние открытия, открытые вопросы и повторные исследования Frullania dilatata и Plagiochila asplenioides» . Журнал систематики и эволюции . 55 (4): 333–339. дои : 10.1111/jse.12266 .
- ^ Бей М., Хассельман М., Фондрк М.К., Пейдж Р.Э., Омхольт С.В. (август 2003 г.). «Ген csd является основным сигналом полового развития медоносной пчелы и кодирует белок SR-типа» . Клетка . 114 (4): 419–429. дои : 10.1016/S0092-8674(03)00606-8 . ПМИД 12941271 .
- ^ Привман Э., Вурм Ю., Келлер Л. (май 2013 г.). «Дублирование и согласованная эволюция главного определителя пола в условиях балансирующего отбора» . Слушания. Биологические науки . 280 (1758): 20122968. doi : 10.1098/rspb.2012.2968 . ПМЦ 3619454 . ПМИД 23466984 .
- ^ Миякава М.О., Цучида К., Миякава Х. (март 2018 г.). «Ген двойного пола объединяет мультилокусные комплементарные сигналы определения пола у японского муравья Vollenhovia emeryi». Биохимия насекомых и молекулярная биология . 94 : 42–49. дои : 10.1016/j.ibmb.2018.01.006 . ПМИД 29408414 .
- ^ ван Вильгенбург Э., Дриссен Г., Бёкебум Л.В. (январь 2006 г.). «Однолокусное комплементарное определение пола у перепончатокрылых: «неразумный» замысел?» . Границы в зоологии . 3 (1): 1. дои : 10.1186/1742-9994-3-1 . ПМК 1360072 . ПМИД 16393347 .
- ^ Гёт А., Бут Д.Т. (март 2005 г.). «Температурно-зависимое соотношение полов у птиц» . Письма по биологии . 1 (1): 31–33. дои : 10.1098/rsbl.2004.0247 . ПМК 1629050 . ПМИД 17148121 .
- ^ Jump up to: Перейти обратно: а б Торрес Мальдонадо LC, Ланда Пьедра А, Морено Мендоса Н, Мармолехо Валенсия А, Меса Мартинес А, Торговец Лариос Х (октябрь 2002 г.). «Профили экспрессии Dax1, Dmrt1 и Sox9 при определении пола по температуре в гонадах морской черепахи Lepidochelys olivacea». Общая и сравнительная эндокринология . 129 (1): 20–26. дои : 10.1016/s0016-6480(02)00511-7 . ПМИД 12409092 .
- ^ Булл Джей-Джей (март 1980 г.). «Определение пола у рептилий». Ежеквартальный обзор биологии . 55 (1): 3–21. дои : 10.1086/411613 . JSTOR 2826077 . S2CID 85177125 .
- ^ Jump up to: Перейти обратно: а б Валенсуэла Н., Янзен Ф.Дж. (2001). «Филопатрия гнездового участка и эволюция определения пола в зависимости от температуры» (PDF) . Исследования в области эволюционной экологии . 3 : 779–794. Архивировано (PDF) из оригинала 4 мая 2013 года . Проверено 7 декабря 2011 г.
- ^ Янзен Ф.Дж., ПК Phillips (ноябрь 2006 г.). «Изучение эволюции определения пола в окружающей среде, особенно у рептилий». Журнал эволюционной биологии . 19 (6): 1775–1784. дои : 10.1111/j.1420-9101.2006.01138.x . ПМИД 17040374 . S2CID 15485510 .
- ^ Гилберт С. (2006). Биология развития (8-е изд.). Сандерленд, Массачусетс: Издательство Sinauer Associates, Inc. стр. 550–553 . ISBN 9780878932504 .
- ^ Уоттс ПК, Бьюли К.Р., Сандерсон С., Бордман В., Чиофи С., Гибсон Р. (декабрь 2006 г.). «Партеногенез у варанов Комодо». Природа . 444 (7122): 1021–1022. Бибкод : 2006Natur.444.1021W . дои : 10.1038/4441021a . ПМИД 17183308 . S2CID 4311088 .
- ^ Лехтонен Дж., Паркер Г.А. (декабрь 2014 г.). «Конкуренция гамет, ограничение гамет и эволюция двух полов» . Молекулярная репродукция человека . 20 (12): 1161–1168. дои : 10.1093/моль/гау068 . ПМИД 25323972 .
- ^ Кларк Р., Мерлин М. (28 июня 2016 г.). Каннабис: эволюция и этноботаника . Университет Калифорнии Пресс. п. 359. ИСБН 978-0-520-29248-2 .
- ^ Намекава С.Х., Ли Дж.Т. (май 2009 г.). «XY и ZW: является ли мейотическая инактивация половых хромосом правилом эволюции?» . ПЛОС Генетика . 5 (5): e1000493. дои : 10.1371/journal.pgen.1000493 . ПМК 2679206 . ПМИД 19461890 .
- ^ Jump up to: Перейти обратно: а б с д Валлендер Э.Дж., Лан Б.Т. (ноябрь 2006 г.). «Множественное независимое происхождение половых хромосом у амниот» . Труды Национальной академии наук Соединенных Штатов Америки . 103 (48): 18031–18032. Бибкод : 2006PNAS..10318031V . дои : 10.1073/pnas.0608879103 . ПМЦ 1838700 . ПМИД 17116892 .
- ^ Стиглек Р., Эзаз Т., Грейвс Дж. А. (2007). «Новый взгляд на эволюцию половых хромосом птиц». Цитогенетические и геномные исследования . 117 (1–4): 103–109. дои : 10.1159/000103170 . ПМИД 17675850 . S2CID 12932564 .
- ^ Вейрунес Ф., Уотерс П.Д., Митке П., Ренс В., Макмиллан Д., Олсоп А.Е. и др. (июнь 2008 г.). «Птичьи половые хромосомы утконоса предполагают недавнее происхождение половых хромосом млекопитающих» . Геномные исследования . 18 (6): 965–973. дои : 10.1101/гр.7101908 . ПМК 2413164 . ПМИД 18463302 .
- ^ Маршалл Грейвс, JA (сентябрь 2000 г.). «Y-хромосома человека, определение пола и сперматогенез – феминистский взгляд» . Биология размножения . 63 (3): 667–676. дои : 10.1095/biolreprod63.3.667b . ПМИД 10952906 .
- ^ Эзаз Т., Стиглек Р., Вейрунес Ф., Маршалл Грейвс Дж. А. (сентябрь 2006 г.). «Взаимоотношения между системами половых хромосом ZW и XY позвоночных». Современная биология . 16 (17): Р736–Р743. Бибкод : 2006CBio...16.R736E . дои : 10.1016/j.cub.2006.08.021 . hdl : 1885/37887 . ПМИД 16950100 . S2CID 18864471 .
- ^ Куинн А.Е., Сарр С.Д., Эзаз Т., Маршалл Грейвс Дж.А., Жорж А. (июнь 2011 г.). «Эволюционные переходы между механизмами определения пола у позвоночных» . Письма по биологии . 7 (3): 443–448. дои : 10.1098/rsbl.2010.1126 . ПМК 3097877 . ПМИД 21212104 .
- ^ Jump up to: Перейти обратно: а б Грейвс Дж. А. (март 2006 г.). «Специализация и дегенерация половых хромосом у млекопитающих» . Клетка . 124 (5): 901–914. дои : 10.1016/j.cell.2006.02.024 . ПМИД 16530039 . S2CID 8379688 .
- ^ «Эволюция половых хромосом: Шаг за шагом» (Пресс-релиз). Медицинский центр Чикагского университета. 28 октября 1999 года . Проверено 23 октября 2011 г.
- ^ Чарльзуорт Б. (14 августа 2003 г.). «Организация и эволюция Y-хромосомы человека» . Геномная биология . 4 (9): 226. doi : 10.1186/gb-2003-4-9-226 . ЧВК 193647 . ПМИД 12952526 .
- ^ Грейвс Дж. А. (22 июля 2004 г.). «Вырожденная Y-хромосома – может ли конверсия спасти ее?». Воспроизводство, рождаемость и развитие . 16 (5): 527–534. дои : 10.1071/RD03096 . ПМИД 15367368 . S2CID 23740483 .
- ^ Хьюз Дж. Ф., Скалецкий Х., Браун Л. Г., Пынтикова Т., Грейвс Т., Фултон Р. С. и др. (февраль 2012 г.). «Строгая эволюционная консервация последовала за быстрой потерей генов в Y-хромосомах человека и резус» . Природа . 483 (7387): 82–86. Бибкод : 2012Natur.483...82H . дои : 10.1038/nature10843 . ПМЦ 3292678 . ПМИД 22367542 .
- ^ Бахтрог Д., Манк Дж.Э., Пайчел К.Л., Киркпатрик М., Отто С.П., Ашман Т.Л. и др. (июль 2014 г.). «Определение пола: почему так много способов сделать это?» . ПЛОС Биология . 12 (7): e1001899. дои : 10.1371/journal.pbio.1001899 . ПМК 4077654 . ПМИД 24983465 .
- ^ Ней М (2 мая 2013 г.). Эволюция, управляемая мутациями . ОУП Оксфорд. п. 168. ИСБН 978-0-19-163781-0 .
Дальнейшее чтение [ править ]
- Бекебум Л., Перрин Н. (2014). Эволюция определения пола . Издательство Оксфордского университета. ISBN 978-0-19-163140-5 .
пола Определение и дифференциация | |
|---|---|
| Обзор | |
| Генетическая основа |
|
| См. также | |
| Базы данных органов управления : Национальные |
|---|