Биология
| Часть серии о |
| Биология |
|---|
Биология – это научное исследование жизни . [1] [2] [3] Это естественная наука с широким охватом, но имеющая несколько объединяющих тем, которые связывают ее в единую, последовательную область. [1] [2] [3] Например, все организмы состоят из клеток , которые обрабатывают наследственную информацию, закодированную в генах , которая может передаваться будущим поколениям. Другая важная тема — эволюция , объясняющая единство и разнообразие жизни. [1] [2] [3] Обработка энергии также важна для жизни, поскольку она позволяет организмам двигаться , расти и размножаться . [1] [2] [3] Наконец, все организмы способны регулировать свою внутреннюю среду . [1] [2] [3] [4] [5]
Биологи способны изучать жизнь на нескольких уровнях организации . [1] от молекулярной биологии клетки до анатомии и физиологии растений и животных, а также эволюции популяций. [1] [6] Следовательно, в биологии существует множество субдисциплин , каждая из которых определяется характером исследовательских вопросов и инструментами , которые они используют. [7] [8] [9] Как и другие ученые, биологи используют научный метод для наблюдения , постановки вопросов, выдвижения гипотез , проведения экспериментов и формирования выводов об окружающем мире. [1]
Жизнь на Земле, возникшая более 3,7 миллиардов лет назад, [10] чрезвычайно разнообразен. Биологи стремились изучить и классифицировать различные формы жизни: от прокариотических организмов, таких как археи и бактерии, до эукариотических организмов, таких как протисты , грибы, растения и животные. Эти различные организмы вносят вклад в биоразнообразие экосистемы круговороте , где они играют специализированную роль в питательных веществ и энергии в биофизической среде .
История

Самые ранние корни науки, включая медицину, можно проследить в Древнем Египте и Месопотамии примерно в 3000–1200 годах до нашей эры . [11] [12] Их вклад сформировал древнегреческую натурфилософию . [11] [12] [13] [14] Древнегреческие философы, такие как Аристотель (384–322 до н.э.), внесли большой вклад в развитие биологических знаний. Он исследовал биологическую причинность и разнообразие жизни. Его преемник Теофраст начал научное изучение растений. [15] Среди ученых средневекового исламского мира, писавших по биологии, были аль-Джахиз (781–869), аль-Динавари (828–896), писавшие по ботанике, [16] и Разес (865–925), писавший по анатомии и физиологии . Медицина особенно хорошо изучалась исламскими учеными, работавшими в традициях греческих философов, а естествознание во многом опиралось на аристотелевскую мысль.
Биология начала быстро развиваться после ван Левенгуком радикального усовершенствования микроскопа Антоном . Именно тогда учёные открыли сперматозоиды , бактерии, инфузории и разнообразие микроскопической жизни. Исследования Яна Сваммердама привели к новому интересу к энтомологии и помогли разработать методы микроскопического вскрытия и окрашивания . [17] Достижения микроскопии оказали глубокое влияние на биологическое мышление. В начале 19 века биологи указали на центральную важность клетки . В 1838 году Шлейден и Шванн начали продвигать ныне универсальные идеи о том, что (1) основной единицей организмов является клетка и (2) что отдельные клетки обладают всеми характеристиками жизни, хотя они и выступали против идеи, что (3) все клетки происходят из от деления других клеток, продолжая поддерживать спонтанное зарождение . Однако Роберт Ремак и Рудольф Вирхов смогли воплотить третий принцип, и к 1860-м годам большинство биологов приняли все три принципа, которые объединились в клеточную теорию . [18] [19]
Тем временем таксономия и классификация оказались в центре внимания естествоиспытателей. Карл Линней опубликовал базовую таксономию мира природы в 1735 году, а в 1750-х годах ввел научные названия для всех своих видов. [20] Жорж-Луи Леклерк, граф де Бюффон , рассматривал виды как искусственные категории, а живые формы как податливые, даже предполагая возможность общего происхождения . [21]

Серьезное эволюционное мышление зародилось в работах Жана-Батиста Ламарка , который представил последовательную теорию эволюции. [23] Британский натуралист Чарльз Дарвин , объединив биогеографический подход Гумбольдта , униформистскую геологию Лайелла , труды Мальтуса о росте населения, а также свои собственные морфологические знания и обширные естественные наблюдения, выработал более успешную эволюционную теорию, основанную на естественном отборе ; аналогичные рассуждения и доказательства привели Альфреда Рассела Уоллеса к независимым выводам. [24] [25]
Основа современной генетики заложена работой Грегора Менделя в 1865 году. [26] Это изложило принципы биологической наследственности. [27] Однако значение его работы не было осознано до начала 20 века, когда эволюция стала единой теорией, поскольку современный синтез примирил дарвиновскую эволюцию с классической генетикой . [28] В 1940-х и начале 1950-х годов серия экспериментов Альфреда Херши и Марты Чейз указала на ДНК как на компонент хромосом , содержащий несущие признаки единицы, которые стали известны как гены . Сосредоточение внимания на новых видах модельных организмов, таких как вирусы в 1953 году двойной спиральной структуры ДНК и бактерии, наряду с открытием Джеймсом Уотсоном и Фрэнсисом Криком ознаменовало переход к эпохе молекулярной генетики . Начиная с 1950-х годов биология значительно расширилась в молекулярной области. Генетический код был взломан Хар Гобиндом Хораной , Робертом У. Холли и Маршаллом Уорреном Ниренбергом после того, как выяснилось, что ДНК содержит кодоны . Проект «Геном человека» человека был запущен в 1990 году с целью картирования генома . [29]
Химическая основа
Атомы и молекулы
Все организмы состоят из химических элементов ; [30] кислород , углерод , водород и азот составляют большую часть (96%) массы всех организмов, а кальций , фосфор , сера , натрий , хлор и магний составляют практически всю остальную часть. Различные элементы могут объединяться, образуя соединения, такие как вода, которая имеет основополагающее значение для жизни. [30] Биохимия – это изучение химических процессов и связанных с ними внутри живых организмов . Молекулярная биология — это раздел биологии, который стремится понять молекулярные основы биологической активности внутри клеток и между ними, включая молекулярный синтез, модификацию, механизмы и взаимодействия.
Вода
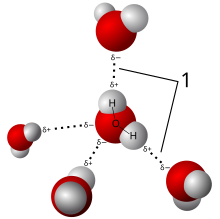
Жизнь возникла из первого океана Земли, образовавшегося около 3,8 миллиарда лет назад. [31] С тех пор вода продолжает оставаться самой распространенной молекулой в каждом организме. Вода важна для жизни, поскольку она является эффективным растворителем , способным растворять растворенные вещества, такие как ионы натрия и хлорида , или другие небольшие молекулы с образованием водного раствора . Растворившись в воде, эти растворенные вещества с большей вероятностью вступят в контакт друг с другом и, следовательно, примут участие в химических реакциях , поддерживающих жизнь. [31] По своей молекулярной структуре вода представляет собой небольшую полярную молекулу изогнутой формы, образованную полярными ковалентными связями двух атомов водорода (Н) с одним атомом кислорода (О) (H 2 O). [31] Поскольку связи O–H полярны, атом кислорода имеет небольшой отрицательный заряд, а два атома водорода имеют небольшой положительный заряд. [31] Это полярное свойство воды позволяет ей притягивать другие молекулы воды посредством водородных связей, что делает воду когезионной . [31] Поверхностное натяжение возникает в результате силы сцепления, возникающей из-за притяжения между молекулами на поверхности жидкости. [31] Вода также является клейкой , поскольку она способна прилипать к поверхности любых полярных или заряженных молекул, не являющихся водой. [31] Вода состоянии плотнее в жидком , чем в твердом состоянии (или льду). [31] Это уникальное свойство воды позволяет льду плавать над жидкой водой, такой как пруды, озера и океаны, тем самым изолируя жидкость внизу от холодного воздуха вверху. [31] Вода обладает способностью поглощать энергию, что придает ей более высокую удельную теплоемкость , чем у других растворителей, таких как этанол . [31] Таким образом, необходимо большое количество энергии, чтобы разорвать водородные связи между молекулами воды и превратить жидкую воду в водяной пар . [31] Как молекула вода не является полностью стабильной, поскольку каждая молекула воды постоянно диссоциирует на ионы водорода и гидроксила, прежде чем снова превратиться в молекулу воды. [31] В чистой воде количество ионов водорода уравновешивает (или равно) количеству гидроксильных ионов, в результате чего pH становится нейтральным.
Органические соединения

Органические соединения — это молекулы, содержащие углерод, связанный с другим элементом, например водородом. [31] За исключением воды, почти все молекулы, составляющие каждый организм, содержат углерод. [31] [32] Углерод может образовывать ковалентные связи с четырьмя другими атомами, что позволяет ему образовывать разнообразные, большие и сложные молекулы. [31] [32] Например, один атом углерода может образовывать четыре одинарные ковалентные связи, например, в метане , две двойные ковалентные связи, например, в диоксиде углерода (CO 2 ), или тройную ковалентную связь, например, в моноксиде углерода (CO). Более того, углерод может образовывать очень длинные цепи соединяющихся углерод-углеродных связей, таких как октан , или кольцевые структуры, такие как глюкоза .
Простейшей формой органической молекулы является углеводород , который представляет собой большое семейство органических соединений, состоящих из атомов водорода, связанных с цепочкой атомов углерода. Углеводородная основная цепь может быть заменена другими элементами, такими как кислород (O), водород (H), фосфор (P) и сера (S), которые могут изменить химическое поведение этого соединения. [31] Группы атомов, которые содержат эти элементы (O-, H-, P- и S-) и связаны с центральным атомом углерода или скелетом, называются функциональными группами . [31] В организмах можно обнаружить шесть основных функциональных групп: аминогруппу , карбоксильную группу , карбонильную группу , гидроксильную группу , фосфатную группу и сульфгидрильную группу . [31]
В 1953 году эксперимент Миллера-Юри показал, что органические соединения могли быть синтезированы абиотически в закрытой системе, имитирующей условия ранней Земли , что позволило предположить, что сложные органические молекулы могли возникнуть спонтанно на ранней Земле (см. Абиогенез ). [33] [31]
Макромолекулы
Макромолекулы — это большие молекулы, состоящие из более мелких субъединиц или мономеров . [34] Мономеры включают сахара, аминокислоты и нуклеотиды. [35] Углеводы включают мономеры и полимеры сахаров. [36] Липиды — единственный класс макромолекул, не состоящих из полимеров. Они включают стероиды , фосфолипиды и жиры. [35] преимущественно неполярные и гидрофобные (водоотталкивающие) вещества. [37] Белки являются наиболее разнообразными макромолекулами. К ним относятся ферменты , транспортные белки , большие сигнальные молекулы, антитела и структурные белки . Основной единицей (или мономером) белка является аминокислота . [34] Двадцать аминокислот используются в белках. [34] Нуклеиновые кислоты представляют собой полимеры нуклеотидов . [38] Их функция — хранить, передавать и выражать наследственную информацию. [35]
Клетки
Клеточная теория утверждает, что клетки являются фундаментальными единицами жизни, что все живые существа состоят из одной или нескольких клеток и что все клетки возникают из ранее существовавших клеток посредством клеточного деления . [39] Большинство клеток очень малы, их диаметр составляет от 1 до 100 микрометров , поэтому их можно увидеть только под световым или электронным микроскопом . [40] Обычно существует два типа клеток: эукариотические клетки, содержащие ядро , и прокариотические клетки, которые его не имеют. Прокариоты — это одноклеточные организмы , такие как бактерии , тогда как эукариоты могут быть одноклеточными или многоклеточными. У многоклеточных организмов каждая клетка организма в конечном итоге происходит из одной клетки оплодотворенной яйцеклетки .
Клеточная структура

Каждая клетка заключена в клеточную мембрану , которая отделяет ее цитоплазму от внеклеточного пространства . [41] Клеточная мембрана состоит из липидного бислоя , включающего холестерин , который находится между фосфолипидами и сохраняет их текучесть при различных температурах. Клеточные мембраны являются полупроницаемыми , пропуская небольшие молекулы, такие как кислород, углекислый газ и вода, одновременно ограничивая движение более крупных молекул и заряженных частиц, таких как ионы . [42] Клеточные мембраны также содержат мембранные белки , в том числе интегральные мембранные белки , которые проходят через мембрану и служат мембранными переносчиками , а также периферические белки , которые свободно прикрепляются к внешней стороне клеточной мембраны, действуя как ферменты, формирующие клетку. [43] Клеточные мембраны участвуют в различных клеточных процессах, таких как клеточная адгезия , хранение электрической энергии и передача сигналов клетками , и служат поверхностью прикрепления для некоторых внеклеточных структур, таких как клеточная стенка , гликокаликс и цитоскелет .
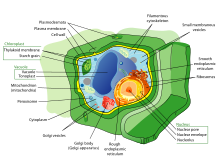
В цитоплазме клетки находится множество биомолекул, таких как белки и нуклеиновые кислоты . [44] Помимо биомолекул, эукариотические клетки имеют специализированные структуры, называемые органеллами , которые имеют собственные липидные бислои или являются пространственными единицами. [45] Эти органеллы включают ядро клетки , которое содержит большую часть клеточной ДНК, или митохондрии , которые генерируют аденозинтрифосфат (АТФ) для питания клеточных процессов. Другие органеллы, такие как эндоплазматическая сеть и аппарат Гольджи, играют роль в синтезе и упаковке белков соответственно. Биомолекулы, такие как белки, могут быть поглощены лизосомами , другой специализированной органеллой. Растительные клетки имеют дополнительные органеллы, которые отличают их от клеток животных, такие как клеточная стенка, которая обеспечивает поддержку растительной клетки, хлоропласты , которые собирают энергию солнечного света для производства сахара, и вакуоли , которые обеспечивают хранение и структурную поддержку, а также участвуют в размножении и расщеплении. семян растений. [45] Эукариотические клетки также имеют цитоскелет, состоящий из микротрубочек , промежуточных нитей и микрофиламентов , которые обеспечивают поддержку клетки и участвуют в движении клетки и ее органелл. [45] С точки зрения структурного состава микротрубочки состоят из тубулина (например, α-тубулина и β-тубулина ), тогда как промежуточные филаменты состоят из волокнистых белков. [45] Микрофиламенты состоят из молекул актина , которые взаимодействуют с другими цепями белков. [45]
Метаболизм
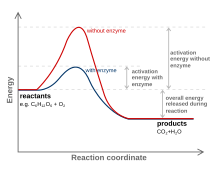
Всем клеткам требуется энергия для поддержания клеточных процессов. Метаболизм – это совокупность химических реакций в организме. Тремя основными целями метаболизма являются: преобразование пищи в энергию для запуска клеточных процессов; преобразование продуктов питания/топлива в мономерные строительные блоки; и устранение метаболических отходов . Эти катализируемые ферментами реакции позволяют организмам расти и размножаться, сохранять свои структуры и реагировать на окружающую среду. Метаболические реакции можно разделить на катаболические — расщепление соединений (например, расщепление глюкозы до пирувата в результате клеточного дыхания ); или анаболический — накопление ( синтез ) соединений (таких как белки, углеводы, липиды и нуклеиновые кислоты). Обычно катаболизм высвобождает энергию, а анаболизм ее потребляет. Химические реакции метаболизма организованы в метаболические пути , в которых одно химическое вещество посредством ряда стадий превращается в другое химическое вещество, причем каждый этап обеспечивается определенным ферментом. Ферменты имеют решающее значение для метаболизма, поскольку они позволяют организмам запускать желаемые реакции, требующие энергии, которые не происходят сами по себе. связывая их со спонтанными реакциями , которые высвобождают энергию. Ферменты действуют как катализаторы — они позволяют реакции протекать быстрее, не потребляя ее, — за счет уменьшения количества энергии активации, необходимой для превращения реагентов в продукты . Ферменты также позволяют регулировать скорость метаболической реакции, например, в ответ на изменения в среде клетки или на сигналы других клеток.
Клеточное дыхание

Клеточное дыхание — это набор метаболических реакций и процессов, которые происходят в клетках для преобразования химической энергии питательных веществ в аденозинтрифосфат (АТФ) и последующего высвобождения отходов. [46] Реакции, участвующие в дыхании, являются катаболическими реакциями , которые расщепляют большие молекулы на более мелкие, высвобождая энергию. Дыхание — один из ключевых способов высвобождения клеткой химической энергии для поддержания клеточной активности. Общая реакция происходит в ряд биохимических стадий, некоторые из которых являются окислительно-восстановительными реакциями. Хотя клеточное дыхание технически является реакцией горения , оно явно не похоже на ту, которая происходит в клетке, из-за медленного, контролируемого высвобождения энергии в результате серии реакций.
Сахар в форме глюкозы является основным питательным веществом, используемым клетками животных и растений при дыхании. Клеточное дыхание с участием кислорода называется аэробным дыханием, которое имеет четыре стадии: гликолиз , цикл лимонной кислоты (или цикл Кребса), цепь переноса электронов и окислительное фосфорилирование . [47] Гликолиз — это метаболический процесс, происходящий в цитоплазме, в ходе которого глюкоза превращается в два пирувата , при этом одновременно образуются две чистые молекулы АТФ. [47] Затем каждый пируват окисляется до ацетил-КоА с помощью комплекса пируватдегидрогеназы , который также генерирует НАДН и диоксид углерода. Ацетил-КоА участвует в цикле лимонной кислоты, который происходит внутри митохондриального матрикса. В конце цикла общий выход из 1 молекулы глюкозы (или 2 пируватов) составляет 6 НАДН, 2 ФАДН 2 и 2 молекулы АТФ. Наконец, следующая стадия — окислительное фосфорилирование, которое у эукариот происходит в кристах митохондрий . Окислительное фосфорилирование включает в себя цепь переноса электронов, которая представляет собой серию из четырех белковых комплексов , которые переносят электроны от одного комплекса к другому, тем самым высвобождая энергию из НАДН и ФАДН 2 , которая связана с перекачкой протонов (ионов водорода) через внутреннюю митохондриальную мембрану. ( хемиосмос ), порождающий движущую силу протонов . [47] Энергия движущей силы протонов заставляет фермент АТФ-синтазу синтезировать больше АТФ путем фосфорилирования АДФ . Перенос электронов заканчивается тем, что молекулярный кислород становится конечным акцептором электронов .
Если бы кислорода не было, пируват не метаболизировался бы в результате клеточного дыхания, а подвергался бы процессу ферментации . Пируват не транспортируется в митохондрии, а остается в цитоплазме, где превращается в продукты жизнедеятельности , которые могут быть удалены из клетки. Это служит цели окисления переносчиков электронов, чтобы они могли снова выполнять гликолиз, и удаления избытка пирувата. Ферментация окисляет НАДН до НАД. + поэтому его можно повторно использовать в гликолизе. В отсутствие кислорода ферментация предотвращает накопление НАДН в цитоплазме и обеспечивает НАД. + для гликолиза. Этот продукт жизнедеятельности варьируется в зависимости от организма. В скелетных мышцах продуктом жизнедеятельности является молочная кислота . Этот тип брожения называется молочнокислым брожением . При напряженных физических нагрузках, когда потребности в энергии превышают запасы энергии, дыхательная цепь не может переработать все атомы водорода, к которым присоединен НАДН. Во время анаэробного гликолиза НАД + регенерирует, когда пары водорода соединяются с пируватом с образованием лактата. Образование лактата катализируется лактатдегидрогеназой в обратимой реакции. Лактат также можно использовать в качестве непрямого предшественника гликогена печени. Во время восстановления, когда кислород становится доступным, НАД + присоединяется к водороду из лактата с образованием АТФ. У дрожжей отходами являются этанол и углекислый газ. Этот тип ферментации известен как алкогольная или этаноловая ферментация . АТФ, образующийся в этом процессе, производится путем фосфорилирования на уровне субстрата , для которого не требуется кислород.
Фотосинтез

Фотосинтез — это процесс, используемый растениями и другими организмами для преобразования энергии света в химическую энергию , которая позже может быть высвобождена для подпитки метаболической деятельности организма посредством клеточного дыхания. Эта химическая энергия хранится в молекулах углеводов, таких как сахара, которые синтезируются из углекислого газа и воды. [48] [49] [50] В большинстве случаев кислород выделяется в виде отходов. Большинство растений, водорослей и цианобактерий осуществляют фотосинтез, который в значительной степени отвечает за производство и поддержание содержания кислорода в атмосфере Земли, а также поставляет большую часть энергии, необходимой для жизни на Земле. [51]
Фотосинтез состоит из четырех стадий: поглощение света , транспорт электронов, синтез АТФ и фиксация углерода . [47] Поглощение света — это начальный этап фотосинтеза, при котором энергия света поглощается пигментами хлорофилла, прикрепленными к белкам в мембранах тилакоидов . Поглощенная энергия света используется для удаления электронов от донора (воды) к первичному акцептору электронов, хинону , обозначенному как Q. На втором этапе электроны перемещаются от хинонного первичного акцептора электронов через ряд переносчиков электронов, пока не достигнут конечный акцептор электронов, который обычно представляет собой окисленную форму НАДФ. + , который восстанавливается до НАДФН, процесс, который происходит в белковом комплексе, называемом фотосистемой I (PSI). Транспорт электронов связан с перемещением протонов (или водорода) из стромы к тилакоидной мембране, что формирует градиент pH через мембрану, поскольку водород становится более концентрированным в просвете, чем в строме. Это аналогично протондвижущей силе, генерируемой на внутренней мембране митохондрий при аэробном дыхании. [47]
На третьем этапе фотосинтеза движение протонов вниз по градиенту концентрации из просвета тилакоида в строму через АТФ-синтазу сопровождается синтезом АТФ той же АТФ-синтазой. [47] НАДФН и АТФ, генерируемые в результате светозависимых реакций на второй и третьей стадиях соответственно, обеспечивают энергию и электроны для стимулирования синтеза глюкозы путем фиксации атмосферного углекислого газа в существующие органические соединения углерода, такие как рибулозобисфосфат (RuBP) в последовательность светонезависимых (или темновых) реакций, называемая циклом Кальвина . [52]
Передача сигналов ячейки
Передача сигналов (или связь) клеток — это способность клеток получать, обрабатывать и передавать сигналы с окружающей средой и с самими собой. [53] [54] Сигналы могут быть нехимическими, такими как свет, электрические импульсы и тепло, или химическими сигналами (или лигандами ), которые взаимодействуют с рецепторами , которые могут быть встроены в клеточную мембрану другой клетки или расположены глубоко внутри клетки. [55] [54] Обычно выделяют четыре типа химических сигналов: аутокринные , паракринные , юкстакринные и гормональные . [55] При аутокринной передаче сигналов лиганд воздействует на ту же клетку, которая его высвобождает. Например, опухолевые клетки могут бесконтрольно размножаться, поскольку они выделяют сигналы, которые инициируют их собственное самоделение. При паракринной передаче сигналов лиганд диффундирует в близлежащие клетки и воздействует на них. Например, клетки головного мозга, называемые нейронами, выделяют лиганды, называемые нейротрансмиттерами , которые диффундируют через синаптическую щель и связываются с рецептором на соседней клетке, такой как другой нейрон или мышечная клетка . При юкстакринной передаче сигналов существует прямой контакт между передающими сигнал и отвечающими клетками. Наконец, гормоны — это лиганды, которые проходят через кровеносную систему животных или сосудистую систему растений, чтобы достичь клеток-мишеней. Как только лиганд связывается с рецептором, он может влиять на поведение другой клетки, в зависимости от типа рецептора. Например, нейротрансмиттеры, связывающиеся с инотропными рецепторами, могут изменять возбудимость клетки-мишени. Другие типы рецепторов включают рецепторы протеинкиназы (например, рецептор гормона инсулина ) и рецепторы, связанные с G-белком . Активация рецепторов, связанных с G-белком, может инициировать каскады вторичных мессенджеров . Процесс, посредством которого химический или физический сигнал передается через клетку в виде серии молекулярных событий, называется трансдукцией сигнала .
Клеточный цикл

Клеточный цикл — это серия событий, происходящих в клетке , которые заставляют ее делиться на две дочерние клетки. Эти события включают дублирование его ДНК и некоторых его органелл , а также последующее разделение его цитоплазмы на две дочерние клетки в процессе, называемом клеточным делением . [56] В эукариотах (т.е. клетках животных, растений, грибов и простейших ) существует два различных типа клеточного деления: митоз и мейоз . [57] Митоз — это часть клеточного цикла, в ходе которого реплицированные хромосомы разделяются на два новых ядра. Деление клеток приводит к образованию генетически идентичных клеток, в которых сохраняется общее число хромосом. Обычно митозу (делению ядра) предшествует S-стадия интерфазы (во время которой ДНК реплицируется) и часто за ней следуют телофаза и цитокинез ; который делит цитоплазму , органеллы и клеточную мембрану одной клетки на две новые клетки, содержащие примерно равные доли этих клеточных компонентов. Различные стадии митоза в совокупности определяют митотическую фазу клеточного цикла животных — деление материнской клетки на две генетически идентичные дочерние клетки. [58] Клеточный цикл — это жизненно важный процесс, посредством которого одноклеточная оплодотворенная яйцеклетка развивается в зрелый организм, а также процесс волос, кожи, клеток крови и некоторых внутренних органов обновления . После деления клеток каждая из дочерних клеток начинает интерфазу нового цикла. В отличие от митоза, мейоз приводит к образованию четырех гаплоидных дочерних клеток в результате одного раунда репликации ДНК с последующими двумя делениями. [59] Гомологичные хромосомы разделяются в первом делении ( мейоз I ), а сестринские хроматиды — во втором делении ( мейоз II ). Оба этих цикла деления клеток используются в процессе полового размножения в определенный момент их жизненного цикла. Считается, что оба они присутствуют у последнего общего предка эукариот.
Прокариоты (т.е. археи и бактерии) также могут подвергаться делению клеток (или бинарному делению ). В отличие от процессов митоза и мейоза у эукариот бинарное деление у прокариот происходит без образования веретенообразного аппарата на клетке. Перед бинарным делением ДНК в бактерии плотно свернута. После того, как он развернулся и размножился, он притягивается к отдельным полюсам бактерии, увеличивая размер, готовясь к расщеплению. Рост новой клеточной стенки начинает отделять бактерию (вызванный полимеризацией FtsZ и образованием «Z-кольца»). [60] Новая клеточная стенка ( перегородка ) полностью развивается, что приводит к полному расщеплению бактерии. Новые дочерние клетки имеют плотно свернутые палочки ДНК, рибосомы и плазмиды .
Половое размножение и мейоз
Мейоз является центральной особенностью полового размножения у эукариот, и наиболее фундаментальной функцией мейоза , по-видимому, является сохранение целостности генома , который передается потомству родителями. [61] [62] Два аспекта полового размножения , мейотическая рекомбинация и ауткроссинг , вероятно, поддерживаются соответственно адаптивными преимуществами рекомбинационного восстановления повреждений геномной ДНК и генетической комплементации , которая маскирует проявление вредных рецессивных мутаций . [63]
Положительный эффект генетической комплементации, возникающий в результате ауткроссинга (перекрестного оплодотворения), также называется гибридной силой или гетерозисом. Чарльз Дарвин в своей книге 1878 года «Эффекты скрещивания и самооплодотворения в растительном мире». [64] в начале главы XII отмечалось: «Первый и самый важный из выводов, которые можно сделать из наблюдений, приведенных в этом томе, заключается в том, что перекрестное оплодотворение обычно полезно, а самоопыление часто вредно, по крайней мере, для растений, на которых Я экспериментировал». Генетическая изменчивость , часто возникающая как побочный продукт полового размножения, может обеспечить долгосрочные преимущества тем половым линиям, которые участвуют в ауткроссинге . [63]
Генетика
Наследование
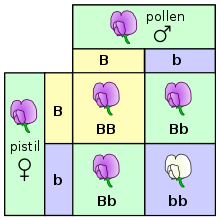
Генетика – это научное исследование наследственности. [65] [66] [67] В частности, менделевское наследование — это процесс, посредством которого гены и признаки передаются от родителей к потомству. [27] У него есть несколько принципов. Во-первых, генетические характеристики, аллели , дискретны и имеют альтернативные формы (например, пурпурный против белого или высокий против карлика), каждая из которых унаследована от одного из двух родителей. Основан на законе доминирования и единообразия , который гласит, что некоторые аллели являются доминантными , а другие — рецессивными ; организм хотя бы с одним доминантным аллелем будет демонстрировать фенотип этого доминантного аллеля. Во время формирования гамет аллели каждого гена разделяются, так что каждая гамета несет только одну аллель для каждого гена. Гетерозиготные особи производят гаметы с одинаковой частотой двух аллелей. Наконец, закон независимого ассортимента гласит, что гены разных признаков могут разделяться независимо во время образования гамет, т. е. гены не связаны между собой. Исключением из этого правила могут быть черты, сцепленные с полом . Тестовые скрещивания можно проводить для экспериментального определения основного генотипа организма с доминантным фенотипом. [68] Квадрат Пеннета можно использовать для прогнозирования результатов тестового скрещивания. Хромосомная теория наследования , утверждающая, что гены находятся в хромосомах, была подтверждена Томаса Морганса экспериментами с плодовыми мушками , которые установили половую связь между цветом глаз и полом у этих насекомых. [69]
Гены и ДНК
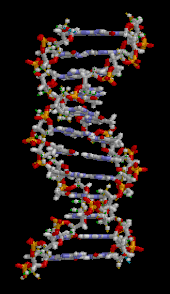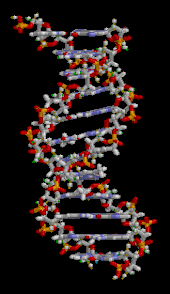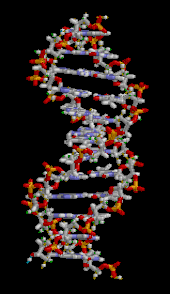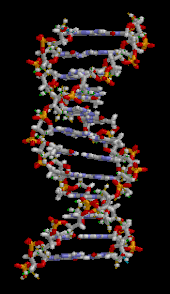
Ген — это единица наследственности , которая соответствует участку дезоксирибонуклеиновой кислоты (ДНК), несущему генетическую информацию, контролирующую форму или функцию организма. ДНК состоит из двух полинуклеотидных цепей, которые скручиваются друг вокруг друга, образуя двойную спираль . [70] Он встречается в виде линейных хромосом у эукариот и кольцевых хромосом у прокариот . Набор хромосом в клетке известен как ее геном . У эукариотов ДНК находится преимущественно в ядре клетки . [71] У прокариотов ДНК находится внутри нуклеоида . [72] Генетическая информация хранится в генах, а полная совокупность в организме называется его генотипом . [73] Репликация ДНК — это полуконсервативный процесс, при котором каждая цепь служит матрицей для новой цепи ДНК. [70] Мутации – это наследственные изменения в ДНК. [70] Они могут возникать спонтанно в результате ошибок репликации, которые не были исправлены при корректуре, или могут быть вызваны окружающей среды, мутагенами такими как химические вещества (например, азотистая кислота , бензопирен ) или радиация (например, рентгеновские лучи , гамма-лучи , ультрафиолетовое излучение). , частицы, испускаемые нестабильными изотопами). [70] Мутации могут приводить к фенотипическим эффектам, таким как потеря функции, усиление функции и условные мутации. [70] Некоторые мутации полезны, поскольку являются источником генетических вариаций для эволюции. [70] Другие вредны, если они приводят к потере функции генов, необходимых для выживания. [70]
Экспрессия генов
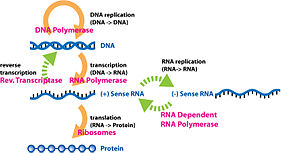
Экспрессия генов — это молекулярный процесс, посредством которого генотип, закодированный в ДНК, порождает наблюдаемый фенотип в белках тела организма. Этот процесс резюмируется центральной догмой молекулярной биологии , сформулированной Фрэнсисом Криком в 1958 году. [74] [75] [76] Согласно Центральной догме, генетическая информация передается от ДНК к РНК и белку. Существует два процесса экспрессии генов: транскрипция (ДНК в РНК) и трансляция (РНК в белок). [77]
Генная регуляция
Регуляция экспрессии генов факторами окружающей среды и на разных стадиях развития может происходить на каждом этапе процесса, таком как транскрипция , сплайсинг РНК , трансляция и посттрансляционная модификация белка. [78] На экспрессию генов может влиять положительная или отрицательная регуляция, в зависимости от того, какой из двух типов регуляторных белков, называемых факторами транскрипции, связывается с последовательностью ДНК рядом с промотором или на нем. [78] Группа генов, имеющих один и тот же промотор, называется опероном и встречается главным образом у прокариот и некоторых низших эукариот (например, Caenorhabditis elegans ). [78] [79] При позитивной регуляции экспрессии генов активатор представляет собой фактор транскрипции, который стимулирует транскрипцию, когда он связывается с последовательностью рядом с промотором или на нем. Негативная регуляция происходит, когда другой фактор транскрипции, называемый репрессором, связывается с последовательностью ДНК, называемой оператором , которая является частью оперона, чтобы предотвратить транскрипцию. Репрессоры могут ингибироваться соединениями, называемыми индукторами (например, аллолактозой ), тем самым позволяя происходить транскрипции. [78] Специфические гены, которые могут быть активированы индукторами, называются индуцибельными генами , в отличие от конститутивных генов , которые практически постоянно активны. [78] В отличие от обоих структурные гены кодируют белки, которые не участвуют в регуляции генов. [78] Помимо регуляторных событий, связанных с промотором, экспрессия генов также может регулироваться эпигенетическими изменениями хроматина , который представляет собой комплекс ДНК и белка, обнаруженный в эукариотических клетках. [78]
Гены, развитие и эволюция
Развитие — это процесс, при котором многоклеточный организм (растение или животное) проходит ряд изменений, начиная с одной клетки и принимая различные формы, характерные для его жизненного цикла. [80] В основе развития лежат четыре ключевых процесса: детерминация , дифференциация , морфогенез и рост. Детерминация определяет судьбу клетки, которая в процессе развития становится более ограничительной. Дифференцировка — это процесс, посредством которого специализированные клетки возникают из менее специализированных клеток, таких как стволовые клетки . [81] [82] Стволовые клетки — это недифференцированные или частично дифференцированные клетки , которые могут дифференцироваться в различные типы клеток и размножаться бесконечно, производя больше одних и тех же стволовых клеток. [83] Клеточная дифференциация резко меняет размер клетки, ее форму, мембранный потенциал , метаболическую активность и чувствительность к сигналам, что во многом обусловлено тщательно контролируемыми модификациями экспрессии генов и эпигенетики . За некоторыми исключениями, клеточная дифференциация почти никогда не предполагает изменения самой последовательности ДНК . [84] Таким образом, разные клетки могут иметь совершенно разные физические характеристики, несмотря на одинаковый геном . Морфогенез, или развитие формы тела, является результатом пространственных различий в экспрессии генов. [80] Небольшая часть генов в геноме организма, называемая набором генетических инструментов развития, контролирует развитие этого организма. Этот набор генов высококонсервативен среди типов , а это означает, что они древние и очень похожи у широко разделенных групп животных. Различия в использовании набора генов влияют на строение тела, а также на количество, идентичность и структуру частей тела. Среди наиболее важных генов инструментария являются Hox -гены . Hox-гены определяют, где повторяющиеся части, такие как многочисленные позвонки змей, будут расти в развивающемся эмбрионе или личинке. [85]
Эволюция
Эволюционные процессы

Эволюция — центральная организующая концепция в биологии. Это изменение наследственных характеристик популяций в течение последующих поколений . [86] [87] При искусственном отборе животных разводили по определенным признакам. [88] Учитывая, что черты наследуются, популяции содержат разнообразную смесь признаков, а воспроизводство способно увеличить любую популяцию, Дарвин утверждал, что в естественном мире именно природа играла роль людей в отборе определенных признаков. [88] Дарвин пришел к выводу, что особи, обладающие наследственными признаками, лучше адаптированными к окружающей среде, с большей вероятностью выживут и произведут больше потомства, чем другие особи. [88] Далее он пришел к выводу, что это приведет к накоплению благоприятных черт в последующих поколениях, тем самым увеличивая соответствие между организмами и окружающей их средой. [89] [90] [91] [88] [92]
Видообразование
Вид — это группа организмов, которые спариваются друг с другом, а видообразование — это процесс, посредством которого одна линия разделяется на две линии в результате эволюции независимо друг от друга. [93] Для того чтобы произошло видообразование, необходима репродуктивная изоляция . [93] Репродуктивная изоляция может быть результатом несовместимости между генами, как описано моделью Бейтсона-Добжанского-Мюллера . Репродуктивная изоляция также имеет тенденцию увеличиваться по мере генетической дивергенции . Видообразование может произойти, когда существуют физические барьеры, разделяющие предковый вид, - процесс, известный как аллопатрическое видообразование . [93]
Филогения

Филогения — это эволюционная история определенной группы организмов или их генов. [94] Это можно представить с помощью филогенетического дерева — диаграммы, показывающей линии происхождения организмов или их генов. Каждая линия, нарисованная на оси времени дерева, представляет собой родословную потомков определенного вида или популяции. Когда линия делится на две части, она представляется в виде развилки или разделения на филогенетическом дереве. [94] Филогенетические деревья являются основой для сравнения и группировки различных видов. [94] Различные виды, которые имеют общие черты, унаследованные от общего предка, описываются как имеющие гомологичные признаки (или синапоморфию ). [95] [96] [94] Филогения составляет основу биологической классификации. [94] Эта система классификации основана на рангах: высший ранг соответствует домену, за которым следуют царство , тип , класс , отряд , семейство , род и вид . [94] Все организмы можно отнести к одному из трех доменов : археи (первоначально архебактерии), бактерии (первоначально эубактерии) или эукарии (включает царства грибов, растений и животных). [97]
История жизни
История жизни на Земле прослеживает эволюцию организмов от самого раннего возникновения жизни до наших дней. Земля образовалась около 4,5 миллиардов лет назад, и вся жизнь на Земле, как живая, так и вымершая, произошла от последнего универсального общего предка , жившего около 3,5 миллиардов лет назад . [98] [99] Геологи разработали геологическую шкалу времени , которая делит историю Земли на основные подразделения, начиная с четырех эонов ( гадейский , архейский , протерозойский и фанерозойский ), первые три из которых вместе известны как докембрийский период , который длился примерно 4 миллиарда лет. годы. [100] Каждый эон можно разделить на эры: фанерозойский эон начался 539 миллионов лет назад. [101] подразделяется на палеозойскую , мезозойскую и кайнозойскую эры. [100] Эти три эры вместе составляют одиннадцать периодов ( кембрийский , ордовикский , силурийский , девонский , каменноугольный , пермский , триасовый , юрский , меловой , третичный и четвертичный ). [100]
Сходство всех известных современных видов указывает на то, что в процессе эволюции они отделились от своего общего предка. [102] Биологи рассматривают повсеместное распространение генетического кода как свидетельство универсального общего происхождения всех бактерий , архей и эукариот . [103] [10] [104] [105] Микробные маты из сосуществовавших бактерий и архей были доминирующей формой жизни в раннем архее , и считается, что многие основные этапы ранней эволюции произошли в этой среде. [106] Самые ранние свидетельства существования эукариот датируются 1,85 миллиарда лет назад. [107] [108] и хотя они, возможно, присутствовали раньше, их диверсификация ускорилась, когда они начали использовать кислород в своем метаболизме . Позже, около 1,7 миллиарда лет назад, многоклеточные организмы стали появляться , дифференцированные клетки которых выполняли специализированные функции. [109]
Водорослеобразные многоклеточные наземные растения возникли примерно 1 миллиард лет назад. [110] хотя данные свидетельствуют о том, что микроорганизмы сформировали самые ранние наземные экосистемы , по крайней мере, 2,7 миллиарда лет назад. [111] Считается, что микроорганизмы проложили путь к появлению наземных растений в ордовикский период. Наземные растения были настолько успешными, что, как полагают, они способствовали вымиранию в позднем девоне . [112]
Эдиакарская биота появляется в эдиакарский период. [113] в то время как позвоночные , наряду с большинством других современных типов, возникли около 525 миллионов лет назад во время кембрийского взрыва . [114] В пермский период на суше господствовали синапсиды , в том числе предки млекопитающих , [115] но большая часть этой группы вымерла в результате пермско-триасового вымирания 252 миллиона лет назад. [116] Во время восстановления после этой катастрофы архозавры стали самыми многочисленными наземными позвоночными; [117] одна группа архозавров, динозавры, доминировала в юрском и меловом периодах. [118] После того , как мел-палеогеновое вымирание 66 миллионов лет назад привело к гибели нептичьих динозавров, [119] млекопитающие быстро увеличивались в размерах и разнообразии . [120] Такие массовые вымирания , возможно, ускорили эволюцию, предоставив возможность диверсификации новым группам организмов. [121]
Разнообразие
Бактерии и археи

Бактерии представляют собой тип клеток которые составляют обширную область прокариотических микроорганизмов , . несколько микрометров Обычно бактерии имеют длину и имеют различную форму : от сфер до палочек и спиралей . Бактерии были одними из первых форм жизни, появившихся на Земле, и присутствуют в большинстве ее сред обитания . Бактерии обитают в почве, воде, кислых горячих источниках , радиоактивных отходах . [122] и глубокая биосфера земной коры . Бактерии также живут в симбиотических и паразитических отношениях с растениями и животными. Большинство бактерий не охарактеризовано, и только около 27 процентов типов бактерий имеют виды, которые можно выращивать в лаборатории. [123]

Археи представляют собой другой домен прокариотических клеток и первоначально были классифицированы как бактерии, получив название архебактерии (в царстве архебактерий ), термин, который вышел из употребления. [124] Клетки архей обладают уникальными свойствами, отличающими их от двух других доменов : бактерий и эукариот . Археи далее делятся на несколько признанных типов . Археи и бактерии в целом схожи по размеру и форме, хотя некоторые археи имеют очень разные формы, например, плоские и квадратные клетки Haloquadratum walsbyi . [125] Несмотря на это морфологическое сходство с бактериями, археи обладают генами и несколькими метаболическими путями , которые более тесно связаны с таковыми у эукариот, особенно в отношении ферментов, участвующих в транскрипции и трансляции . Другие аспекты биохимии архей уникальны, например, их зависимость от эфирных липидов в клеточных мембранах . [126] включая археолы . Археи используют больше источников энергии, чем эукариоты: они варьируются от органических соединений , таких как сахара, до аммиака , ионов металлов или даже газообразного водорода . Солеустойчивые археи ( Haloarchaea ) используют солнечный свет в качестве источника энергии, а другие виды архей фиксируют углерод , но, в отличие от растений и цианобактерий , ни один из известных видов архей не делает и то, и другое. Археи размножаются бесполым путем бинарным делением , фрагментацией или почкованием ; В отличие от бактерий, ни один из известных видов архей не образует эндоспор .
Первыми наблюдаемыми архей были экстремофилы , живущие в экстремальных условиях, таких как горячие источники и соленые озера, без других организмов. Усовершенствованные инструменты молекулярного обнаружения привели к обнаружению архей практически во всех средах обитания , включая почву, океаны и болота . Археи особенно многочисленны в океанах, а археи в планктоне могут быть одной из самых многочисленных групп организмов на планете.
Археи — важная часть жизни на Земле . Они являются частью микробиоты всех организмов. В микробиоме человека они важны в кишечнике , ротовой полости и на коже. [127] Их морфологическое, метаболическое и географическое разнообразие позволяет им выполнять множество экологических ролей: фиксацию углерода; круговорот азота; оборот органических соединений; микробных симбиотических и синтрофных сообществ. и поддержание , например, [128]
Эукариоты

Предполагается, что эукариоты отделились от архей, после чего последовал их эндосимбиоз с бактериями (или симбиогенез ), давший начало митохондриям и хлоропластам, которые сейчас являются частью современных эукариотических клеток. [129] Основные линии эукариот диверсифицировались в докембрии около 1,5 миллиардов лет назад и могут быть разделены на восемь основных клад : альвеоляты , экскаваты , страменопили , растения, ризарии , амебозои , грибы и животные. [129] Пять из этих клад вместе известны как протисты , которые в основном представляют собой микроскопические эукариотические организмы, не являющиеся растениями, грибами или животными. [129] Хотя вполне вероятно, что протисты имеют общего предка ( последнего общего предка эукариот ), [130] Сами по себе протисты не составляют отдельную кладу, поскольку некоторые протисты могут быть более тесно связаны с растениями, грибами или животными, чем с другими протистами. Подобно таким группам, как водоросли , беспозвоночные или простейшие , группа протистов не является формальной таксономической группой, но используется для удобства. [129] [131] Большинство протистов одноклеточные; их называют микробными эукариотами. [129]
Растения — это в основном многоклеточные организмы , преимущественно фотосинтезирующие эукариоты царства Plantae , за исключением грибов и некоторых водорослей . Растительные клетки произошли в результате эндосимбиоза цианобактерии в ранний эукариот около одного миллиарда лет назад, который дал начало хлоропластам. [132] Первые несколько клад, возникшие в результате первичного эндосимбиоза, были водными, и большинство водных фотосинтезирующих эукариотических организмов все вместе описываются как водоросли, что является удобным термином, поскольку не все водоросли тесно связаны между собой. [132] Водоросли состоят из нескольких отдельных клад, таких как глаукофиты , которые представляют собой микроскопические пресноводные водоросли, которые, возможно, по форме напоминали раннего одноклеточного предка Plantae. [132] В отличие от глаукофитов, другие группы водорослей, такие как красные и зеленые водоросли, являются многоклеточными. Зеленые водоросли состоят из трех основных клад: хлорофитов , колеохетофитов и каменных водорослей . [132]
Грибы – это эукариоты, которые переваривают пищу вне своего тела. [133] секретируют пищеварительные ферменты, которые расщепляют крупные молекулы пищи перед их поглощением через клеточные мембраны. Многие грибы также являются сапробами , питаясь мертвыми органическими веществами, что делает их важными разлагателями в экологических системах. [133]
Животные – многоклеточные эукариоты. За некоторыми исключениями, животные потребляют органический материал , дышат кислородом , способны двигаться , могут размножаться половым путем и растут из полой сферы клеток , бластулы , во время эмбрионального развития . более 1,5 миллиона живых животных видов Описано , но , из которых около 1 миллиона являются насекомыми , по оценкам, всего насчитывается более 7 миллионов видов животных. Они имеют сложные взаимодействия друг с другом и окружающей средой, образуя сложные пищевые сети . [134]
Вирусы
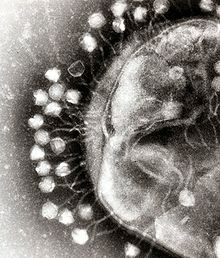
Вирусы — это субмикроскопические инфекционные агенты которые размножаются внутри клеток организмов , . [135] Вирусы заражают все виды форм жизни , от животных и растений до микроорганизмов , включая бактерии и археи . [136] [137] более 6000 видов вирусов . Подробно описано [138] Вирусы встречаются практически во всех экосистемах Земли и являются самым многочисленным типом биологических объектов. [139] [140]
Происхождение вирусов в эволюционной истории жизни неясно: некоторые, возможно, произошли от плазмид — фрагментов ДНК, которые могут перемещаться между клетками, — тогда как другие, возможно, произошли от бактерий. В эволюции вирусы являются важным средством горизонтального переноса генов , который увеличивает генетическое разнообразие аналогично половому размножению . [141] Поскольку вирусы обладают некоторыми, но не всеми характеристиками жизни, их называют «организмами на грани жизни». [142] и как самовоспроизводители . [143]
Экология
Экология – это наука о распространении и изобилии жизни, взаимодействии организмов с окружающей средой . [144]
Экосистемы
Сообщество влажностью живых ( биотических ) организмов в сочетании с неживыми ( абиотическими ) компонентами (например, водой, светом, радиацией, температурой, , атмосферой , кислотностью и почвой) окружающей их среды называется экосистемой . [145] [146] [147] Эти биотические и абиотические компоненты связаны друг с другом через круговорот питательных веществ и потоки энергии. [148] Энергия Солнца поступает в систему посредством фотосинтеза и включается в ткани растений. Питаясь растениями и друг другом, животные перемещают материю и энергию по системе. Они также влияют на количество присутствующей растительной и микробной биомассы . Разлагая мертвое органическое вещество , разлагающие вещества высвобождают углерод обратно в атмосферу и облегчают круговорот питательных веществ , превращая питательные вещества, хранящиеся в мертвой биомассе, обратно в форму, которая может быть легко использована растениями и другими микробами. [149]
Население

Популяция – это группа организмов одного вида , занимающая определенную территорию и размножающаяся из поколения в поколение. [150] [151] [152] [153] [154] Численность населения можно оценить, умножив плотность населения на площадь или объем. Емкость — это окружающей среды максимальный размер популяции вида , который может поддерживаться в этой конкретной среде с учетом имеющихся продуктов питания, среды обитания , воды и других ресурсов . [155] На емкость популяции могут влиять изменения условий окружающей среды, такие как изменения в наличии ресурсов и стоимости их содержания. Что касается человеческих популяций , новые технологии , такие как Зеленая революция, со временем помогли увеличить пропускную способность Земли для людей, что загнало в тупик попытки предсказания надвигающегося сокращения численности населения, самый известный из которых был сделан Томасом Мальтусом в 18 веке. [150]
Сообщества

Сообщество – это группа популяций видов, населяющих одну и ту же географическую территорию в одно и то же время. Биологическое взаимодействие — это воздействие, которое пары организмов, живущих вместе в сообществе, оказывают друг на друга. Они могут быть как одного вида (внутривидовые взаимодействия), так и разных видов (межвидовые взаимодействия). Эти эффекты могут быть краткосрочными, как опыление и хищничество , или долгосрочными; оба часто сильно влияют на эволюцию рассматриваемых видов. Длительное взаимодействие называется симбиозом . Симбиозы варьируются от мутуализма , выгодного обоим партнерам, до конкуренции , вредной для обоих партнеров. [157] Каждый вид участвует в качестве потребителя, ресурса или того и другого во взаимодействиях потребитель-ресурс , которые образуют ядро пищевых цепей или пищевых сетей . [158] В любой пищевой сети существуют разные трофические уровни , причем самый низкий уровень — это первичные продуценты (или автотрофы ), такие как растения и водоросли, которые преобразуют энергию и неорганический материал в органические соединения , которые затем могут быть использованы остальной частью сообщества. [51] [159] [160] На следующем уровне находятся гетеротрофы — виды, которые получают энергию за счет расщепления органических соединений других организмов. [158] Гетеротрофы, потребляющие растения, являются первичными потребителями (или травоядными ), тогда как гетеротрофы, потребляющие травоядных, являются вторичными потребителями (или хищниками ). А те, кто едят вторичных потребителей, являются третичными потребителями и так далее. Всеядные гетеротрофы способны потреблять на нескольких уровнях. Наконец, существуют редуценты , которые питаются отходами жизнедеятельности или мертвыми телами организмов. [158] В среднем общее количество энергии, включаемой в биомассу трофического уровня в единицу времени, составляет около одной десятой энергии трофического уровня, которую он потребляет. Оставшиеся девяносто процентов энергии, не потребляемой следующим трофическим уровнем, составляют отходы и мертвый материал, используемые редуцентами, а также тепло, теряемое в результате метаболизма. [161]
Биосфера
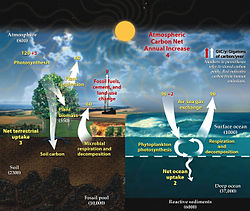
В глобальной экосистеме или биосфере материя существует в виде различных взаимодействующих компартментов, которые могут быть биотическими или абиотическими, а также доступными или недоступными, в зависимости от их формы и местоположения. [163] Например, вещества наземных автотрофов одновременно биотичны и доступны для других организмов, тогда как вещества горных пород и минералов абиотичны и недоступны. Биогеохимический цикл — это путь, по которому определенные элементы материи переворачиваются или перемещаются через биотические ( биосфера ) и абиотические ( литосфера , атмосфера и гидросфера ) отсеки Земли. Существуют биогеохимические циклы азота , углерода и воды .
Сохранение
Биология сохранения — это изучение сохранения биоразнообразия Земли с целью защиты видов , их среды обитания и экосистем от чрезмерных темпов вымирания и разрушения биотических взаимодействий. [164] [165] [166] Он занимается факторами, влияющими на поддержание, утрату и восстановление биоразнообразия, а также наукой о поддержании эволюционных процессов, которые порождают генетическое , популяционное, видовое и экосистемное разнообразие. [167] [168] [169] [170] Беспокойство вызвано оценками, предполагающими, что до 50% всех видов на планете исчезнут в течение следующих 50 лет. [171] что способствовало бедности, голоданию и обнулит ход эволюции на этой планете. [172] [173] Биоразнообразие влияет на функционирование экосистем, которые предоставляют разнообразные услуги , от которых зависят люди. Биологи-природоохранители исследуют и обучают тенденциям утраты биоразнообразия видов , исчезновения и негативному влиянию, которое они оказывают на нашу способность поддерживать благополучие человеческого общества. Организации и граждане реагируют на текущий кризис биоразнообразия посредством планов действий по сохранению, которые направляют исследовательские, мониторинговые и образовательные программы, которые затрагивают проблемы на местном и глобальном уровнях. [174] [167] [168] [169]
См. также
- Биология в художественной литературе
- Глоссарий биологии
- Список биологических сайтов
- Список биологов
- Список биологических журналов
- Список тем по биологии
- Список наук о жизни
- Список тем омики по биологии
- Национальная ассоциация учителей биологии
- Очерк биологии
- Периодическая таблица наук о жизни в четырех вопросах Тинбергена
- Научный туризм
- Терминология биологии
Ссылки
- ^ Jump up to: а б с д и ж г час Урри, Лиза; Каин, Майкл; Вассерман, Стивен; Минорский, Петр; Рис, Джейн (2017). «Эволюция, темы биологии и научные исследования». Кэмпбелл Биология (11-е изд.). Нью-Йорк: Пирсон. стр. 2–26. ISBN 978-0134093413 .
- ^ Jump up to: а б с д и Хиллис, Дэвид М.; Хеллер, Х. Крейг; Хакер, Салли Д.; Ласковски, Марта Дж.; Садава, Дэвид Э. (2020). «Изучение жизни». Жизнь: наука биологии (12-е изд.). У. Х. Фриман. ISBN 978-1319017644 .
- ^ Jump up to: а б с д и Фриман, Скотт; Куиллин, Ким; Эллисон, Лизабет; Блэк, Майкл; Подгорски, Грег; Тейлор, Эмили; Кармайкл, Джефф (2017). «Биология и тройка жизни». Биологические науки (6-е изд.). Хобокен, Нью-Джерси: Пирсон. стр. 1–18. ISBN 978-0321976499 .
- ^ Моделл, Гарольд; Клифф, Уильям; Майкл, Джоэл; Макфарланд, Дженни; Вендерот, Мэри Пэт; Райт, Энн (декабрь 2015 г.). «Взгляд физиолога на гомеостаз» . Достижения в области физиологического образования . 39 (4): 259–266. дои : 10.1152/advan.00107.2015 . ПМЦ 4669363 . ПМИД 26628646 .
- ^ Дэвис, ПК; Рипер, Э; Тушинский, Ю.А. (январь 2013 г.). «Самоорганизация и снижение энтропии в живой клетке» . Биосистемы . 111 (1): 1–10. Бибкод : 2013BiSys.111....1D . doi : 10.1016/j.biosystems.2012.10.005 . ПМЦ 3712629 . ПМИД 23159919 .
- ^ На основе определения: «Глоссарий терминов проекта Aquarena Wetlands Project» . Техасский государственный университет в Сан-Маркосе. Архивировано из оригинала 8 июня 2004 г.
- ^ Крейг, Нэнси (2014). Молекулярная биология, Принципы функционирования генома . ОУП Оксфорд. ISBN 978-0-19-965857-2 .
- ^ Москони, Франческо; Жюлу, Томас; Депра, Николя; Синха, Дипак Кумар; Аллеманд, Жан-Франсуа; Винсент Крокетт; Бенсимон, Дэвид (2008). «Некоторые нелинейные задачи биологии». Нелинейность . 21 (8): Т131. Бибкод : 2008Nonli..21..131M . дои : 10.1088/0951-7715/21/8/T03 . S2CID 119808230 .
- ^ Хауэлл, Элизабет (8 декабря 2014 г.). «Как жизнь стала сложной и могла ли она произойти за пределами Земли?» . Журнал астробиологии . Архивировано из оригинала 17 августа 2018 года . Проверено 14 февраля 2018 г.
{{cite web}}: CS1 maint: неподходящий URL ( ссылка ) - ^ Jump up to: а б Пирс, Бен К.Д.; Таппер, Эндрю С.; Пудриц, Ральф Э .; и др. (1 марта 2018 г.). «Ограничение временного интервала возникновения жизни на Земле». Астробиология . 18 (3): 343–364. arXiv : 1808.09460 . Бибкод : 2018AsBio..18..343P . дои : 10.1089/ast.2017.1674 . ПМИД 29570409 . S2CID 4419671 .
- ^ Jump up to: а б Линдберг, Дэвид К. (2007). «Наука до греков». Истоки западной науки: европейская научная традиция в философском, религиозном и институциональном контексте (2-е изд.). Чикаго, Иллинойс: Издательство Чикагского университета. стр. 1–20. ISBN 978-0-226-48205-7 .
- ^ Jump up to: а б Грант, Эдвард (2007). «Древний Египет до Платона». История натуральной философии: от древнего мира до девятнадцатого века . Нью-Йорк: Издательство Кембриджского университета. стр. 1–26 . ISBN 978-052-1-68957-1 .
- ^ Магнер, Лоис Н. (2002). История наук о жизни, переработанная и расширенная . ЦРК Пресс. ISBN 978-0-203-91100-6 . Архивировано из оригинала 24 марта 2015 г.
- ^ Серафини, Энтони (2013). Эпическая история биологии . Спрингер. ISBN 978-1-4899-6327-7 . Архивировано из оригинала 15 апреля 2021 года . Проверено 14 июля 2015 г.
- ^ Одно или несколько предыдущих предложений включают текст из публикации, которая сейчас находится в свободном доступе : Чисхолм, Хью , изд. (1911). « Теофраст ». Британская энциклопедия (11-е изд.). Издательство Кембриджского университета.
- ^ Фахд, Туфик (1996). «Ботаника и сельское хозяйство». В Морлоне, Режис; Рашед, Рошди (ред.). Энциклопедия истории арабской науки . Том. 3. Рутледж . п. 815. ИСБН 978-0-415-12410-2 .
- ^ Магнер, Лоис Н. (2002). История наук о жизни, переработанная и расширенная . ЦРК Пресс. стр. 133–44. ISBN 978-0-203-91100-6 . Архивировано из оригинала 24 марта 2015 г.
- ^ Сапп, Январь (2003). «7». Бытие: эволюция биологии . Нью-Йорк: Издательство Оксфордского университета. ISBN 978-0-19-515618-8 .
- ^ Коулман, Уильям (1977). Биология в девятнадцатом веке: проблемы формы, функции и трансформации . Нью-Йорк: Издательство Кембриджского университета. ISBN 978-0-521-29293-1 .
- ^ Майр, Эрнст. Рост биологической мысли , глава 4
- ^ Майр, Эрнст. Рост биологической мысли , глава 7
- ^ * Дарвин, Фрэнсис , изд. (1909). Основы «Происхождения видов», очерк, написанный в 1842 году (PDF) . Кембридж: напечатано в University Press. п. 53. LCCN 61057537 . OCLC 1184581 . Архивировано (PDF) из оригинала 4 марта 2016 года . Проверено 27 ноября 2014 г.
- ^ Гулд, Стивен Джей . Структура эволюционной теории . Издательство Belknap Press Гарвардского университета: Кембридж, 2002. ISBN 0-674-00613-5 . п. 187.
- ^ Майр, Эрнст. «Рост биологической мысли» , глава 10: «Дарвиновские доказательства эволюции и общего происхождения»; и глава 11: «Причины эволюции: естественный отбор».
- ^ Ларсон, Эдвард Дж. (2006). «Ч. 3» . Эволюция: замечательная история научной теории . Издательская группа Random House. ISBN 978-1-58836-538-5 . Архивировано из оригинала 24 марта 2015 г.
- ^ Хениг (2000). На. цит . стр. 134–138.
- ^ Jump up to: а б Мико, Илона (2008). «Принципы наследственности Грегора Менделя составляют краеугольный камень современной генетики. Так что же они собой представляют?» . Природное образование . 1 (1): 134. Архивировано из оригинала 19 июля 2019 г. Проверено 13 мая 2021 г.
- ^ Футуйма, Дуглас Дж.; Киркпатрик, Марк (2017). «Эволюционная биология». Эволюция (4-е изд.). Сандерленд, Массачусетс: Sinauer Associates. стр. 3–26.
- ^ Благородный, Иван (14 апреля 2003 г.). «Геном человека наконец-то завершен» . Новости Би-би-си . Архивировано из оригинала 14 июня 2006 г. Проверено 22 июля 2006 г.
- ^ Jump up to: а б Урри, Лиза; Каин, Майкл; Вассерман, Стивен; Минорский, Петр; Рис, Джейн (2017). «Химический контекст жизни». Кэмпбелл Биология (11-е изд.). Нью-Йорк: Пирсон. стр. 28–43. ISBN 978-0134093413 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с Фриман, Скотт; Куиллин, Ким; Эллисон, Лизабет; Блэк, Майкл; Подгорски, Грег; Тейлор, Эмили; Кармайкл, Джефф (2017). «Вода и углерод: химическая основа жизни». Биологические науки (6-е изд.). Хобокен, Нью-Джерси: Пирсон. стр. 55–77. ISBN 978-0321976499 .
- ^ Jump up to: а б Урри, Лиза; Каин, Майкл; Вассерман, Стивен; Минорский, Петр; Рис, Джейн (2017). «Углерод и молекулярное разнообразие жизни». Кэмпбелл Биология (11-е изд.). Нью-Йорк: Пирсон. стр. 56–65. ISBN 978-0134093413 .
- ^ Хиллис, Дэвид М.; Садава, Давид; Хилл, Ричард В.; Прайс, Мэри В. (2014). «Углеродное и молекулярное разнообразие жизни». Принципы жизни (2-е изд.). Сандерленд, Массачусетс: Sinauer Associates. стр. 56–65. ISBN 978-1464175121 .
- ^ Jump up to: а б с Фриман, Скотт; Куиллин, Ким; Эллисон, Лизабет; Блэк, Майкл; Подгорски, Грег; Тейлор, Эмили; Кармайкл, Джефф (2017). «Структура и функции белка». Биологические науки (6-е изд.). Хобокен, Нью-Джерси: Пирсон. стр. 78–92. ISBN 978-0321976499 .
- ^ Jump up to: а б с Урри, Лиза; Каин, Майкл; Вассерман, Стивен; Минорский, Петр; Рис, Джейн (2017). «Структура и функции крупных биологических молекул». Кэмпбелл Биология (11-е изд.). Нью-Йорк: Пирсон. стр. 66–92. ISBN 978-0134093413 .
- ^ Фриман, Скотт; Куиллин, Ким; Эллисон, Лизабет; Блэк, Майкл; Подгорски, Грег; Тейлор, Эмили; Кармайкл, Джефф (2017). «Введение в углеводы». Биологические науки (6-е изд.). Хобокен, Нью-Джерси: Пирсон. стр. 107–118. ISBN 978-0321976499 .
- ^ Фриман, Скотт; Куиллин, Ким; Эллисон, Лизабет; Блэк, Майкл; Подгорски, Грег; Тейлор, Эмили; Кармайкл, Джефф (2017). «Липиды, мембраны и первые клетки». Биологические науки (6-е изд.). Хобокен, Нью-Джерси: Пирсон. стр. 119–141. ISBN 978-0321976499 .
- ^ Фриман, Скотт; Куиллин, Ким; Эллисон, Лизабет; Блэк, Майкл; Подгорски, Грег; Тейлор, Эмили; Кармайкл, Джефф (2017). «Нуклеиновые кислоты и мир РНК». Биологические науки (6-е изд.). Хобокен, Нью-Джерси: Пирсон. стр. 93–106. ISBN 978-0321976499 .
- ^ Маццарелло, П. (май 1999 г.). «Объединяющая концепция: история клеточной теории». Природная клеточная биология . 1 (1): Е13–15. дои : 10.1038/8964 . ПМИД 10559875 . S2CID 7338204 .
- ^ Кэмпбелл, Нил А.; Уильямсон, Брэд; Хейден, Робин Дж. (2006). Биология: исследование жизни . Бостон: Пирсон Прентис Холл. ISBN 978-0132508827 . Архивировано из оригинала 2 ноября 2014 г. Проверено 13 мая 2021 г.
- ^ Урри, Лиза; Каин, Майкл; Вассерман, Стивен; Минорский, Петр; Рис, Джейн (2017). «Структура и функция мембраны». Кэмпбелл Биология (11-е изд.). Нью-Йорк: Пирсон. стр. 126–142. ISBN 978-0134093413 .
- ^ Альбертс, Б.; Джонсон, А.; Льюис, Дж.; и др. (2002). Молекулярная биология клетки (4-е изд.). Нью-Йорк: Garland Science. ISBN 978-0-8153-3218-3 . Архивировано из оригинала 20 декабря 2017 г.
- ^ Том Херрманн; Сандип Шарма (2 марта 2019 г.). «Физиология, Мембрана» . СтатПерлз . PMID 30855799 . Архивировано из оригинала 17 февраля 2022 года . Проверено 14 мая 2021 г.
- ^ Альбертс, Брюс; Джонсон, Александр; Льюис, Джулиан; Рафф, Мартин; Робертс, Кейт; Уолтер, Питер (2002). «Движения клеток и формирование тела позвоночных» . Молекулярная биология клетки (4-е изд.). Архивировано из оригинала 22 января 2020 г. Проверено 13 мая 2021 г. В тексте Альбертса обсуждается, как «клеточные строительные блоки» движутся, формируя развивающиеся эмбрионы . также принято описывать Малые молекулы, такие как аминокислоты, как « молекулярные строительные блоки Архивировано 22 января 2020 г. в Wayback Machine . ».
- ^ Jump up to: а б с д и Хиллис, Дэвид М.; Садава, Давид; Хилл, Ричард В.; Прайс, Мэри В. (2014). «Клетки: рабочие единицы жизни». Принципы жизни (2-е изд.). Сандерленд, Массачусетс: Sinauer Associates. стр. 60–81. ISBN 978-1464175121 .
- ^ Бэйли, Регина. «Клеточное дыхание» . Архивировано из оригинала 5 мая 2012 г.
- ^ Jump up to: а б с д и ж Лодиш, Харви; Берк, Арнольд; Кайзер, Крис А.; Кригер, Монти; Скотт, Мэтью П.; Бретшер, Энтони; Плоэ, Хидде; Мацудайра, Пол (2008). «Клеточная энергетика». Молекулярно-клеточная биология (6-е изд.). Нью-Йорк: WH Freeman and Company. стр. 479–532. ISBN 978-0716776017 .
- ^ «фотосинтез» . Интернет-словарь этимологии . Архивировано из оригинала 7 марта 2013 г. Проверено 23 мая 2013 г.
- ^ φῶς . Лидделл, Генри Джордж ; Скотт, Роберт ; Греко-английский лексикон в проекте «Персей»
- ^ σύνθεσις . Лидделл, Генри Джордж ; Скотт, Роберт ; Греко-английский лексикон в проекте «Персей»
- ^ Jump up to: а б Брайант, округ Колумбия; Фригаард, Нью-Йорк (ноябрь 2006 г.). «Освещение фотосинтеза и фототрофии прокариот». Тенденции в микробиологии . 14 (11): 488–496. дои : 10.1016/j.tim.2006.09.001 . ПМИД 16997562 .
- ^ Рис, Дж.; Урри, Л.; Каин, М. (2011). Биология (международное изд.). Река Аппер-Сэддл, Нью-Джерси: Pearson Education . стр. 235, 244 . ISBN 978-0-321-73975-9 .
Это первоначальное включение углерода в органические соединения известно как фиксация углерода.
- ^ Нейтцель, Джеймс; Расбанд, Мэтью. «Сотовая связь» . Природное образование . Архивировано из оригинала 29 сентября 2010 года . Проверено 29 мая 2021 г.
- ^ Jump up to: а б «Сотовая сигнализация» . Природное образование . Архивировано из оригинала 31 октября 2010 года . Проверено 29 мая 2021 г.
- ^ Jump up to: а б Хиллис, Дэвид М.; Садава, Давид; Хилл, Ричард В.; Прайс, Мэри В. (2014). «Клеточные мембраны и передача сигналов». Принципы жизни (2-е изд.). Сандерленд, Массачусетс: Sinauer Associates. стр. 82–104. ISBN 978-1464175121 .
- ^ Мартин, Э.А.; Хайн, Р. (2020). Биологический словарь (6-е изд.). Оксфорд: Издательство Оксфордского университета. ISBN 978-0199204625 . OCLC 176818780 .
- ^ Гриффитс, Эй Джей (2012). Введение в генетический анализ (10-е изд.). Нью-Йорк: WH Freeman. ISBN 978-1429229432 . OCLC 698085201 .
- ^ «10.2 Клеточный цикл – Биология 2e | OpenStax» . openstax.org . 28 марта 2018 г. Архивировано из оригинала 29 ноября 2020 г. Проверено 24 ноября 2020 г.
- ^ Фриман, Скотт; Куиллин, Ким; Эллисон, Лизабет; Блэк, Майкл; Подгорски, Грег; Тейлор, Эмили; Кармайкл, Джефф (2017). "Мейоз". Биологические науки (6-е изд.). Хобокен, Нью-Джерси: Пирсон. стр. 271–289. ISBN 978-0321976499 .
- ^ Казираги, А.; Суйго, Л.; Валоти, Э.; Страньеро, В. (февраль 2020 г.). «Нацеливание на деление бактериальных клеток: связывающий сайт-центрированный подход к наиболее многообещающим ингибиторам незаменимого белка FtsZ» . Антибиотики . 9 (2): 69. doi : 10.3390/antibiotics9020069 . ПМК 7167804 . ПМИД 32046082 .
- ^ Брандейс М. Идеи нового века о древнем сексе: отделение мейоза от спаривания может решить загадку вековой давности. Biol Rev Camb Philos Soc. Май 2018 г.;93(2):801-810. дои: 10.1111/brv.12367. Epub, 14 сентября 2017 г. PMID: 28913952.
- ^ Хёрандль Э. Апомиксис и парадокс пола у растений. Энн Бот. 18 марта 2024 г.: mcae044. doi: 10.1093/aob/mcae044. Epub перед печатью. PMID: 38497809
- ^ Jump up to: а б Бернштейн Х., Байерли Х.К., Хопф Ф.А., Мишод Р.Э. Генетические повреждения, мутации и эволюция пола. Наука. 1985, 20 сентября; 229 (4719): 1277-81. doi: 10.1126/science.3898363. PMID: 3898363
- ^ Дарвин, CR 1878. Эффекты перекрестного и самооплодотворения в растительном царстве. Лондон: Джон Мюррей. darwin-online.org.uk
- ^ Гриффитс, Энтони Дж.; Весслер, Сьюзен Р.; Кэрролл, Шон Б.; Добли, Джон (2015). «Генетическая революция». Введение в генетический анализ (11-е изд.). Сандерленд, Массачусетс: WH Freeman & Company. стр. 1–30. ISBN 978-1464109485 .
- ^ Гриффитс, Энтони Дж. Ф.; Миллер, Джеффри Х.; Сузуки, Дэвид Т.; Левонтин, Ричард С.; Гелбарт, Уильям М., ред. (2000). «Генетика и организм: Введение» . Введение в генетический анализ (7-е изд.). Нью-Йорк: WH Freeman. ISBN 978-0-7167-3520-5 .
- ^ Хартл, Д.; Джонс, Э. (2005). Генетика: анализ генов и геномов (6-е изд.). Джонс и Бартлетт. ISBN 978-0-7637-1511-3 .
- ^ Мико, Илона (2008). «Испытательные кресты» . Природное образование . 1 (1): 136. Архивировано из оригинала 21 мая 2021 г. Проверено 28 мая 2021 г.
- ^ Мико, Илона (2008). «Томас Хант Морган и половая связь» . Природное образование . 1 (1): 143. Архивировано из оригинала 20 мая 2021 г. Проверено 28 мая 2021 г.
- ^ Jump up to: а б с д и ж г Хиллис, Дэвид М.; Садава, Давид; Хилл, Ричард В.; Прайс, Мэри В. (2014). «ДНК и ее роль в наследственности». Принципы жизни (2-е изд.). Сандерленд, Массачусетс: Sinauer Associates. стр. 172–193. ISBN 978-1464175121 .
- ^ Рассел, Питер (2001). iГенетика . Нью-Йорк: Бенджамин Каммингс. ISBN 0-8053-4553-1 .
- ^ Танбихлер, М; Ван, Южная Каролина; Шапиро, Л. (октябрь 2005 г.). «Бактериальный нуклеоид: высокоорганизованная и динамичная структура» . Журнал клеточной биохимии . 96 (3): 506–21. дои : 10.1002/jcb.20519 . ПМИД 15988757 . S2CID 25355087 .
- ^ «Определение генотипа - Определения Медицинского словаря» . Medterms.com. 19 марта 2012 г. Архивировано из оригинала 21 сентября 2013 г. Проверено 2 октября 2013 г.
- ^ Крик, Фрэнсис Х. (1958). «О синтезе белка». Симпозиумы Общества экспериментальной биологии . 12 : 138–63. ПМИД 13580867 .
- ^ Крик, Фрэнсис Х. (август 1970 г.). «Центральная догма молекулярной биологии». Природа . 227 (5258): 561–3. Бибкод : 1970Natur.227..561C . дои : 10.1038/227561a0 . ПМИД 4913914 . S2CID 4164029 .
- ^ «Центральная догма перевернута». Природа . 226 (5252): 1198–9. Июнь 1970 г. Бибкод : 1970Natur.226.1198. . дои : 10.1038/2261198a0 . ПМИД 5422595 . S2CID 4184060 .
- ^ Лин, Ихан; Еловиц, Майкл Б. (2016). «Центральная догма становится цифровой» . Молекулярная клетка . 61 (6): 791–792. doi : 10.1016/j.molcel.2016.03.005 . ПМИД 26990983 .
- ^ Jump up to: а б с д и ж г Хиллис, Дэвид М.; Садава, Давид; Хилл, Ричард В.; Прайс, Мэри В. (2014). «Регуляция экспрессии генов». Принципы жизни (2-е изд.). Сандерленд, Массачусетс: Sinauer Associates. стр. 215–233. ISBN 978-1464175121 .
- ^ Кин, Джек Д.; Тененбаум, Скотт А. (2002). «Эукариотические мРНП могут представлять собой посттранскрипционные опероны» . Молекулярная клетка . 9 (6): 1161–1167. дои : 10.1016/s1097-2765(02)00559-2 . ПМИД 12086614 .
- ^ Jump up to: а б Хиллис, Дэвид М.; Садава, Давид; Хилл, Ричард В.; Прайс, Мэри В. (2014). «Гены, развитие и эволюция». Принципы жизни (2-е изд.). Сандерленд, Массачусетс: Sinauer Associates. стр. 273–298. ISBN 978-1464175121 .
- ^ Slack, JMW (2013) Основная биология развития. Уайли-Блэквелл, Оксфорд.
- ^ Слэк, JMW (2007). «Метаплазия и трансдифференцировка: от чистой биологии к клинике». Nature Reviews Молекулярно-клеточная биология . 8 (5): 369–378. дои : 10.1038/nrm2146 . ПМИД 17377526 . S2CID 3353748 .
- ^ Атала, Энтони; Ланца, Роберт (2012). Справочник по стволовым клеткам . Академическая пресса. п. 452. ИСБН 978-0-12-385943-3 . Архивировано из оригинала 12 апреля 2021 г. Проверено 28 мая 2021 г.
- ^ Янес, Оскар; Кларк, Джули; Вонг, Диана М.; Патти, Гэри Дж.; Санчес-Руис, Антонио; Бентон, Х. Пол; Траугер, Суния А.; Деспон, Кэролайн; Дин, Шэн; Сюздак, Гэри (июнь 2010 г.). «Метаболическое окисление регулирует дифференцировку эмбриональных стволовых клеток» . Химическая биология природы . 6 (6): 411–417. дои : 10.1038/nchembio.364 . ПМЦ 2873061 . ПМИД 20436487 .
- ^ Кэрролл, Шон Б. «Происхождение формы» . Естественная история . Архивировано из оригинала 9 октября 2018 года . Проверено 9 октября 2016 г.
Биологи могли бы с уверенностью сказать, что формы меняются и что естественный отбор является важной силой, способствующей изменениям. Однако они ничего не могли сказать о том, как осуществляется это изменение. Как изменяются тела или части тела или как возникают новые структуры, оставалось полной загадкой.
- ^ Холл, Брайан К.; Халлгримссон, Бенедикт (2007). Эволюция Стрикбергера . Издательство Джонс и Бартлетт. стр. 4–6. ISBN 978-1-4496-4722-3 . Архивировано из оригинала 26 марта 2023 г. Проверено 27 мая 2021 г.
- ^ «Ресурсы эволюции» . Вашингтон, округ Колумбия: Национальные академии наук, техники и медицины . 2016. Архивировано из оригинала 3 июня 2016 г.
- ^ Jump up to: а б с д Урри, Лиза; Каин, Майкл; Вассерман, Стивен; Минорский, Петр; Рис, Джейн (2017). «Происхождение с модификациями: дарвиновский взгляд на жизнь». Кэмпбелл Биология (11-е изд.). Нью-Йорк: Пирсон. стр. 466–483. ISBN 978-0134093413 .
- ^ Левонтин, Ричард К. (ноябрь 1970 г.). «Единицы выбора» (PDF) . Ежегодный обзор экологии и систематики . 1 :1–18. doi : 10.1146/annurev.es.01.110170.000245 . JSTOR 2096764 . S2CID 84684420 . Архивировано (PDF) из оригинала 6 февраля 2015 г.
- ^ Дарвин, Чарльз (1859). О происхождении видов , Джон Мюррей.
- ^ Футуйма, Дуглас Дж.; Киркпатрик, Марк (2017). «Эволюционная биология». Эволюция (4-е изд.). Сандерленд, Массачусетс: Sinauer Associates. стр. 3–26.
- ^ Хиллис, Дэвид М.; Садава, Давид; Хилл, Ричард В.; Прайс, Мэри В. (2014). «Процессы эволюции». Принципы жизни (2-е изд.). Сандерленд, Массачусетс: Sinauer Associates. стр. 299–324. ISBN 978-1464175121 .
- ^ Jump up to: а б с Хиллис, Дэвид М.; Садава, Давид; Хилл, Ричард В.; Прайс, Мэри В. (2014). «Видообразование». Принципы жизни (2-е изд.). Сандерленд, Массачусетс: Sinauer Associates. стр. 343–356. ISBN 978-1464175121 .
- ^ Jump up to: а б с д и ж Хиллис, Дэвид М.; Садава, Давид; Хилл, Ричард В.; Прайс, Мэри В. (2014). «Реконструкция и использование филогений». Принципы жизни (2-е изд.). Сандерленд, Массачусетс: Sinauer Associates. стр. 325–342. ISBN 978-1464175121 .
- ^ Китчинг, Ян Дж.; Фори, Питер Л.; Уильямс, Дэвид М. (2001). «Кладистика» . У Левина, Саймон А. (ред.). Энциклопедия биоразнообразия (2-е изд.). Эльзевир. стр. 33–45. дои : 10.1016/B978-0-12-384719-5.00022-8 . ISBN 9780123847201 . Архивировано из оригинала 29 августа 2021 года . Проверено 29 августа 2021 г. )
- ^ Футуйма, Дуглас Дж.; Киркпатрик, Марк (2017). «Филогения: Единство и разнообразие жизни». Эволюция (4-е изд.). Сандерленд, Массачусетс: Sinauer Associates. стр. 401–429.
- ^ Вёзе, ЧР; Кандлер, О; Уилис, ML (июнь 1990 г.). «На пути к естественной системе организмов: предложение по доменам архей, бактерий и эукариев» . Труды Национальной академии наук Соединенных Штатов Америки . 87 (12): 4576–79. Бибкод : 1990PNAS...87.4576W . дои : 10.1073/pnas.87.12.4576 . ПМК 54159 . ПМИД 2112744 .
- ^ Монтевиль, М; Моссио, М; Пошевиль, А; Лонго, Дж. (октябрь 2016 г.). «Теоретические основы биологии: вариации» . Прогресс биофизики и молекулярной биологии . От века генома к веку организма: новые теоретические подходы. 122 (1): 36–50. doi : 10.1016/j.pbiomolbio.2016.08.005 . ПМИД 27530930 . S2CID 3671068 . Архивировано из оригинала 20 марта 2018 г.
- ^ Де Дюв, Кристиан (2002). Развитие жизни: молекулы, разум и смысл . Нью-Йорк: Издательство Оксфордского университета. п. 44 . ISBN 978-0-19-515605-8 .
- ^ Jump up to: а б с Хиллис, Дэвид М.; Садава, Давид; Хилл, Ричард В.; Прайс, Мэри В. (2014). «История жизни на Земле». Принципы жизни (2-е изд.). Сандерленд, Массачусетс: Sinauer Associates. стр. 357–376. ISBN 978-1464175121 .
- ^ «Стратиграфическая схема 2022» (PDF) . Международная стратиграфическая комиссия. Февраль 2022 г. Архивировано (PDF) из оригинала 2 апреля 2022 г. Проверено 25 апреля 2022 г.
- ^ Футуйма 2005 г.
- ^ Футуйма, диджей (2005). Эволюция . Синауэр Ассошиэйтс. ISBN 978-0-87893-187-3 . OCLC 57311264 .
- ^ Розинг, Миник Т. (29 января 1999 г.). " 13 Микрочастицы обедненного углеродом углерода в осадочных породах морского дна возрастом > 3700 млн лет назад из Западной Гренландии». Science . 283 (5402): 674–676. Bibcode : 1999Sci...283..674R . doi : 10.1126/science.283.5402. 674 . ПМИД 9924024 .
- ^ Отомо, Йоко; Какегава, Такеши; Исида, Акизуми; и др. (январь 2014 г.). «Свидетельства наличия биогенного графита в метаосадочных породах раннего архея Исуа». Природа Геонауки . 7 (1): 25–28. Бибкод : 2014NatGe...7...25O . дои : 10.1038/ngeo2025 .
- ^ Нисбет, Юан Г.; Фаулер, CMR (7 декабря 1999 г.). «Архейская метаболическая эволюция микробных матов» . Труды Королевского общества Б. 266 (1436): 2375–2382. дои : 10.1098/rspb.1999.0934 . ПМК 1690475 .
- ^ Нолл, Эндрю Х .; Жаво, Эммануэль Ж.; Хьюитт, Дэвид; и др. (29 июня 2006 г.). «Эукариотические организмы в протерозойских океанах» . Философские труды Королевского общества Б. 361 (1470): 1023–1038. дои : 10.1098/rstb.2006.1843 . ПМЦ 1578724 . ПМИД 16754612 .
- ^ Федонкин Михаил Александрович (31 марта 2003 г.). «Происхождение Metazoa в свете протерозойской летописи окаменелостей» (PDF) . Палеонтологические исследования . 7 (1): 9–41. дои : 10.2517/prpsj.7.9 . S2CID 55178329 . Архивировано из оригинала (PDF) 26 февраля 2009 г. Проверено 2 сентября 2008 г.
- ^ Боннер, Джон Тайлер (7 января 1998 г.). «Истоки многоклеточности». Интегративная биология . 1 (1): 27–36. doi : 10.1002/(SICI)1520-6602(1998)1:1<27::AID-INBI4>3.0.CO;2-6 .
- ^ Стротер, Пол К.; Баттисон, Лейла; Брейзер, Мартин Д .; и др. (26 мая 2011 г.). «Самые ранние неморские эукариоты Земли». Природа . 473 (7348): 505–509. Бибкод : 2011Natur.473..505S . дои : 10.1038/nature09943 . ПМИД 21490597 . S2CID 4418860 .
- ^ Беральди-Кампези, Уго (23 февраля 2013 г.). «Ранняя жизнь на суше и первые наземные экосистемы» . Экологические процессы . 2 (1): 1–17. Бибкод : 2013ЭкоПр...2....1Б . дои : 10.1186/2192-1709-2-1 .
- ^ Алгео, Томас Дж.; Шеклер, Стивен Э. (29 января 1998 г.). «Наземно-морские телекоммуникации в девоне: связи между эволюцией наземных растений, процессами выветривания и морскими бескислородными явлениями» . Философские труды Королевского общества Б. 353 (1365): 113–130. дои : 10.1098/rstb.1998.0195 . ПМК 1692181 .
- ^ Цзюнь-Юань, Чен; Оливери, Паола; Чиа-Вэй, Ли; и др. (25 апреля 2000 г.). «Разнообразие животных докембрия: предполагаемые фосфатизированные эмбрионы из формации Доушантуо в Китае» . Учеб. Натл. акад. наук. США 97 (9): 4457–4462. Бибкод : 2000PNAS...97.4457C . дои : 10.1073/pnas.97.9.4457 . ПМК 18256 . ПМИД 10781044 .
- ^ ДГ., Шу; ХЛ., Ло; Конвей Моррис, Саймон ; и др. (4 ноября 1999 г.). «Позвоночные животные нижнего кембрия из южного Китая» (PDF) . Природа . 402 (6757): 42–46. Бибкод : 1999Natur.402...42S . дои : 10.1038/46965 . S2CID 4402854 . Архивировано из оригинала (PDF) 26 февраля 2009 г. Проверено 22 января 2015 г.
- ^ Хойт, Дональд Ф. (17 февраля 1997 г.). «Синапсидные рептилии» . ЗОО 138 Зоология позвоночных (Лекция). Помона, Калифорния: Калифорнийский государственный политехнический университет, Помона . Архивировано из оригинала 20 мая 2009 г. Проверено 22 января 2015 г.
- ^ Барри, Патрик Л. (28 января 2002 г.). Филлипс, Тони (ред.). «Великое умирание» . Наука@НАСА . Центр космических полетов Маршалла . Архивировано из оригинала 10 апреля 2010 г. Проверено 22 января 2015 г.
- ^ Таннер, Лоуренс Х.; Лукас, Спенсер Г .; Чепмен, Мэри Г. (март 2004 г.). «Оценка данных и причин вымираний в позднем триасе» (PDF) . Обзоры наук о Земле . 65 (1–2): 103–139. Бибкод : 2004ESRv...65..103T . дои : 10.1016/S0012-8252(03)00082-5 . Архивировано из оригинала (PDF) 25 октября 2007 г. Проверено 22 октября 2007 г.
- ^ Бентон, Майкл Дж. (1997). Палеонтология позвоночных (2-е изд.). Лондон: Чепмен и Холл . ISBN 978-0-412-73800-5 . OCLC 37378512 .
- ^ Фастовский, Дэвид Э.; Шиэн, Питер М. (март 2005 г.). «Вымирание динозавров в Северной Америке» (PDF) . ГСА сегодня . 15 (3): 4–10. doi : 10.1130/1052-5173(2005)015<4:TEOTDI>2.0.CO;2 . Архивировано (PDF) из оригинала 22 марта 2019 г. Проверено 23 января 2015 г.
- ^ Роуч, Джон (20 июня 2007 г.). «Вымирание динозавров стимулировало появление современных млекопитающих» . Национальные географические новости . Вашингтон, округ Колумбия: Национальное географическое общество . Архивировано из оригинала 11 мая 2008 г. Проверено 21 февраля 2020 г.
- Уибл, Джон Р.; Ружье, Гильермо В.; Новачек, Майкл Дж.; и др. (21 июня 2007 г.). «Меловые эвтерийцы и лавразийское происхождение плацентарных млекопитающих вблизи границы K/T». Природа . 447 (7147): 1003–1006. Бибкод : 2007Natur.447.1003W . дои : 10.1038/nature05854 . ПМИД 17581585 . S2CID 4334424 .
- ^ Ван Валкенбург, Блэр (1 мая 1999 г.). «Основные закономерности в истории хищных млекопитающих» . Ежегодный обзор наук о Земле и планетах . 27 : 463–493. Бибкод : 1999AREPS..27..463V . дои : 10.1146/annurev.earth.27.1.463 . Архивировано из оригинала 29 февраля 2020 года . Проверено 15 мая 2021 г.
- ^ Фредриксон, Дж. К.; Захара, Дж. М.; Балквилл, Д.Л. (июль 2004 г.). «Геомикробиология вадозных отложений, сильно загрязненных ядерными отходами, на полигоне в Хэнфорде, штат Вашингтон» . Прикладная и экологическая микробиология . 70 (7): 4230–41. Бибкод : 2004ApEnM..70.4230F . дои : 10.1128/АЕМ.70.7.4230-4241.2004 . ПМК 444790 . ПМИД 15240306 .
- ^ Дудек, Северная Каролина; Вс, CL; Бурштейн, Д. (2017). «Новое микробное разнообразие и функциональный потенциал микробиома полости рта морских млекопитающих» (PDF) . Современная биология . 27 (24): 3752–3762. Бибкод : 2017CBio...27E3752D . дои : 10.1016/j.cub.2017.10.040 . ПМИД 29153320 . S2CID 43864355 . Архивировано (PDF) из оригинала 8 марта 2021 г. Проверено 14 мая 2021 г.
- ^ Пейс, Северная Каролина (май 2006 г.). «Время перемен» . Природа . 441 (7091): 289. Бибкод : 2006Natur.441..289P . дои : 10.1038/441289a . ПМИД 16710401 . S2CID 4431143 .
- ^ Стокениус, В. (октябрь 1981 г.). «Квадратная бактерия Уолсби: тонкая структура ортогонального прокариота» . Журнал бактериологии . 148 (1): 352–60. дои : 10.1128/JB.148.1.352-360.1981 . ПМК 216199 . ПМИД 7287626 .
- ^ «Основная биология архей» . Март 2018 г. Архивировано из оригинала 28 апреля 2021 г. Проверено 14 мая 2021 г.
- ^ Банг, К.; Шмитц, РА (сентябрь 2015 г.). «Археи, связанные с человеческими поверхностями: не следует недооценивать» . Обзоры микробиологии FEMS . 39 (5): 631–48. дои : 10.1093/femsre/fuv010 . ПМИД 25907112 .
- ^ Мойссль-Эйхингер. С.; Паусан, М.; Таффнер, Дж.; Берг, Г.; Банг, К.; Шмитц, Р.А. (январь 2018 г.). «Археи — интерактивные компоненты сложных микробиомов». Тенденции в микробиологии . 26 (1): 70–85. дои : 10.1016/j.tim.2017.07.004 . ПМИД 28826642 .
- ^ Jump up to: а б с д и Хиллис, Дэвид М.; Садава, Давид; Хилл, Ричард В.; Прайс, Мэри В. (2014). «Происхождение и разнообразие эукариот». Принципы жизни (2-е изд.). Сандерленд, Массачусетс: Sinauer Associates. стр. 402–419. ISBN 978-1464175121 .
- ^ О'Мэлли, Морин А.; Леже, Мишель М.; Уайдман, Джереми Г.; Руис-Трилло, Иньяки (18 февраля 2019 г.). «Представления о последнем общем предке эукариот». Экология и эволюция природы . 3 (3). ООО «Спрингер Сайенс энд Бизнес Медиа»: 338–344. Бибкод : 2019NatEE...3..338O . дои : 10.1038/s41559-019-0796-3 . hdl : 10261/201794 . ПМИД 30778187 . S2CID 67790751 .
- ^ Тейлор, FJR 'М. (01.11.2003). «Крах системы двух королевств, подъем протистологии и основание Международного общества эволюционной протистологии (ISEP)» . Международный журнал систематической и эволюционной микробиологии . 53 (6). Общество микробиологов: 1707–1714. дои : 10.1099/ijs.0.02587-0 . ПМИД 14657097 .
- ^ Jump up to: а б с д Хиллис, Дэвид М.; Садава, Давид; Хилл, Ричард В.; Прайс, Мэри В. (2014). «Эволюция растений». Принципы жизни (2-е изд.). Сандерленд, Массачусетс: Sinauer Associates. стр. 420–449. ISBN 978-1464175121 .
- ^ Jump up to: а б Хиллис, Дэвид М.; Садава, Давид; Хилл, Ричард В.; Прайс, Мэри В. (2014). «Эволюция и разнообразие грибов». Принципы жизни (2-е изд.). Сандерленд, Массачусетс: Sinauer Associates. стр. 451–468. ISBN 978-1464175121 .
- ^ Хиллис, Дэвид М.; Садава, Давид; Хилл, Ричард В.; Прайс, Мэри В. (2014). «Происхождение и разнообразие животных». Принципы жизни (2-е изд.). Сандерленд, Массачусетс: Sinauer Associates. стр. 469–519. ISBN 978-1464175121 .
- ^ Ву, К.Дж. (15 апреля 2020 г.). «Во Вселенной больше вирусов, чем звезд. Почему нас заражают лишь некоторые из них? – На Земле существует более квадриллиона квадриллионов отдельных вирусов, но большинство из них не готовы перейти к людям. Сможем ли мы найти те, которые есть?» . Национальное географическое общество . Архивировано из оригинала 28 мая 2020 года . Проверено 18 мая 2020 г.
- ^ Кунин Е.В.; Сенкевич Т.Г.; Доля, В.В. (сентябрь 2006 г.). «Древний мир вирусов и эволюция клеток» . Биология Директ . 1 (1): 29. дои : 10.1186/1745-6150-1-29 . ПМЦ 1594570 . ПМИД 16984643 .
- ^ Циммер, К. (26 февраля 2021 г.). «Тайная жизнь коронавируса. Маслянистый пузырь генов шириной 100 нанометров убил более двух миллионов человек и изменил мир. Ученые не совсем знают, что с этим делать» . Нью-Йорк Таймс . Архивировано из оригинала 28 декабря 2021 г. Проверено 28 февраля 2021 г.
- ^ «Таксономия вирусов: выпуск 2019 г.» . talk.ictvonline.org . Международный комитет по таксономии вирусов. Архивировано из оригинала 20 марта 2020 года . Проверено 25 апреля 2020 г.
- ^ Лоуренс CM; Менон С.; Эйлерс, Би Джей (май 2009 г.). «Структурно-функциональные исследования архейных вирусов» . Журнал биологической химии . 284 (19): 12599–603. дои : 10.1074/jbc.R800078200 . ПМК 2675988 . ПМИД 19158076 .
- ^ Эдвардс, РА; Ровер, Ф. (июнь 2005 г.). «Вирусная метагеномика». Обзоры природы. Микробиология . 3 (6): 504–10. дои : 10.1038/nrmicro1163 . ПМИД 15886693 . S2CID 8059643 .
- ^ Канчая, К.; Фурноус, Г.; Чибани-Ченнуфи, С. (август 2003 г.). «Фаг как агент латерального переноса генов». Современное мнение в микробиологии . 6 (4): 417–24. дои : 10.1016/S1369-5274(03)00086-9 . ПМИД 12941415 .
- ^ Рыбицкий, Е.П. (1990). «Классификация организмов на пороге жизни, или Проблемы вирусной систематики». Южноафриканский научный журнал . 86 : 182–86.
- ^ Кунин Е.В.; Старокадомский, П. (октябрь 2016 г.). «Живы ли вирусы? Парадигма репликатора проливает решающий свет на старый, но ошибочный вопрос» . Исследования по истории и философии биологических и биомедицинских наук . 59 : 125–134. дои : 10.1016/j.shpsc.2016.02.016 . ПМК 5406846 . ПМИД 26965225 .
- ^ Бегон, М; Таунсенд, Чехия; Харпер, Дж.Л. (2006). Экология: От особей к экосистемам (4-е изд.). Блэквелл. ISBN 978-1-4051-1117-1 .
- ^ Места обитания мира . Нью-Йорк: Маршалл Кавендиш. 2004. с. 238. ИСБН 978-0-7614-7523-1 . Архивировано из оригинала 15 апреля 2021 г. Проверено 24 августа 2020 г.
- ^ Тэнсли (1934); Моллес (1999), с. 482; Чапин и др. (2002), с. 380; По данным Шульце и др. (2005); п. 400; По данным Гуревича и др. (2006), с. 522; Смит и Смит 2012, с. Г-5
- ^ Хиллис, Дэвид М.; Садава, Давид; Хилл, Ричард В.; Прайс, Мэри В. (2014). «Распределение экологических систем Земли». Принципы жизни (2-е изд.). Сандерленд, Массачусетс: Sinauer Associates. стр. 845–863. ISBN 978-1464175121 .
- ^ Одум, Юджин П. (1971). Основы экологии (3-е изд.). Нью-Йорк: Сондерс. ISBN 978-0-534-42066-6 .
- ^ Чапин III, Ф. Стюарт; Мэтсон, Памела А.; Муни, Гарольд А. (2002). «Концепция экосистемы». Принципы экологии наземных экосистем . Нью-Йорк: Спрингер. п. 10. ISBN 978-0-387-95443-1 .
- ^ Jump up to: а б Хиллис, Дэвид М.; Садава, Давид; Хилл, Ричард В.; Прайс, Мэри В. (2014). «Популяции». Принципы жизни (2-е изд.). Сандерленд, Массачусетс: Sinauer Associates. стр. 864–897. ISBN 978-1464175121 .
- ^ Урри, Лиза; Каин, Майкл; Вассерман, Стивен; Минорский, Петр; Рис, Джейн (2017). «Популяционная экология». Кэмпбелл Биология (11-е изд.). Нью-Йорк: Пирсон. стр. 1188–1211. ISBN 978-0134093413 .
- ^ «Население» . Биология онлайн. Архивировано из оригинала 13 апреля 2019 года . Проверено 5 декабря 2012 г.
- ^ «Определение популяции (биология)» . Оксфордские словари . Издательство Оксфордского университета. Архивировано из оригинала 4 марта 2016 года . Проверено 5 декабря 2012 г.
сообщество животных, растений или людей, среди членов которого происходит скрещивание
- ^ Хартл, Дэниел (2007). Принципы популяционной генетики . Синауэр Ассошиэйтс . п. 45. ИСБН 978-0-87893-308-2 .
- ^ Чепмен, Эрик Дж.; Байрон, Кэрри Дж. (01 января 2018 г.). «Гибкое применение несущей способности в экологии» . Глобальная экология и охрана природы . 13 : e00365. Бибкод : 2018GEcoC..1300365C . дои : 10.1016/j.gecco.2017.e00365 .
- ^ Одум, Е.П.; Барретт, GW (2005). Основы экологии (5-е изд.). Брукс/Коул, часть Cengage Learning. ISBN 978-0-534-42066-6 . Архивировано из оригинала 20 августа 2011 г.
- ^ Вуттон, Джей Ти; Эммерсон, М. (2005). «Измерение силы взаимодействия в природе». Ежегодный обзор экологии, эволюции и систематики . 36 : 419–44. doi : 10.1146/annurev.ecolsys.36.091704.175535 . JSTOR 30033811 .
- ^ Jump up to: а б с Хиллис, Дэвид М.; Садава, Давид; Хилл, Ричард В.; Прайс, Мэри В. (2014). «Экологические и эволюционные последствия внутри и между видами». Принципы жизни (2-е изд.). Сандерленд, Массачусетс: Sinauer Associates. стр. 882–897. ISBN 978-1464175121 .
- ^ Смит, Ал. (1997). Оксфордский словарь биохимии и молекулярной биологии . Оксфорд [Оксфордшир]: Издательство Оксфордского университета. п. 508. ИСБН 978-0-19-854768-6 .
Фотосинтез – синтез организмами органических химических соединений, особенно. углеводы из углекислого газа с использованием энергии, полученной от света, а не от окисления химических соединений.
- ^ Эдвардс, Катрина. «Микробиология отстойника и подстилающего его молодого, холодного, гидрологически активного склона хребта». Океанографический институт Вудс-Хоул .
- ^ Хиллис, Дэвид М.; Садава, Давид; Хилл, Ричард В.; Прайс, Мэри В. (2014). «Экологические сообщества». Принципы жизни (2-е изд.). Сандерленд, Массачусетс: Sinauer Associates. стр. 898–915. ISBN 978-1464175121 .
- ^ Рибик, Холли (16 июня 2011 г.). «Углеродный цикл» . Земная обсерватория . НАСА. Архивировано из оригинала 5 марта 2016 года . Проверено 5 апреля 2018 г.
- ^ Хиллис, Дэвид М.; Садава, Давид; Хилл, Ричард В.; Прайс, Мэри В. (2014). «Распределение экологических систем Земли». Принципы жизни (2-е изд.). Сандерленд, Массачусетс: Sinauer Associates. стр. 916–934. ISBN 978-1464175121 .
- ^ Сахни, С.; Бентон, MJ (2008). «Восстановление после самого глубокого массового вымирания всех времен» . Труды Королевского общества B: Биологические науки . 275 (1636): 759–65. дои : 10.1098/rspb.2007.1370 . ПМЦ 2596898 . ПМИД 18198148 .
- ^ Суле, Майкл Э.; Уилкокс, Брюс А. (1980). Биология сохранения: эволюционно-экологическая перспектива . Сандерленд, Массачусетс: Sinauer Associates. ISBN 978-0-87893-800-1 .
- ^ Соуле, Майкл Э. (1986). «Что такое биология сохранения?» (PDF) . Бионаука . 35 (11). Американский институт биологических наук: 727–34. дои : 10.2307/1310054 . JSTOR 1310054 . Архивировано из оригинала (PDF) 12 апреля 2019 г. Проверено 15 мая 2021 г.
- ^ Jump up to: а б Хантер, Малкольм Л. (1996). Основы биологии сохранения . Оксфорд: Блэквелл Наука. ISBN 978-0-86542-371-8 .
- ^ Jump up to: а б Меффе, Гэри К.; Марта Дж. Грум (2006). Принципы биологии сохранения (3-е изд.). Сандерленд, Массачусетс: Sinauer Associates. ISBN 978-0-87893-518-5 .
- ^ Jump up to: а б Ван Дайк, Фред (2008). Биология сохранения: основы, концепции, приложения (2-е изд.). Нью-Йорк: Springer-Verlag . дои : 10.1007/978-1-4020-6891-1 . hdl : 11059/14777 . ISBN 978-1402068904 . ОСЛК 232001738 . Архивировано из оригинала 27 июля 2020 г. Проверено 15 мая 2021 г.
- ^ Сахни, С.; Бентон, MJ; Ферри, Пенсильвания (2010). «Связь между глобальным таксономическим разнообразием, экологическим разнообразием и распространением позвоночных на суше» . Письма по биологии . 6 (4): 544–7. дои : 10.1098/rsbl.2009.1024 . ПМК 2936204 . ПМИД 20106856 .
- ^ Ко, Лиан Пин; Данн, Роберт Р.; Содхи, Навджот С.; Колвелл, Роберт К.; Проктор, Хизер С.; Смит, Винсент С. (2004). «Сосуществование видов и кризис биоразнообразия». Наука . 305 (5690): 1632–4. Бибкод : 2004Sci...305.1632K . дои : 10.1126/science.1101101 . ПМИД 15361627 . S2CID 30713492 .
- ^ Оценка экосистемы тысячелетия (2005). Экосистемы и благополучие человека: синтез биоразнообразия. Институт мировых ресурсов, Вашингтон, округ Колумбия [1]. Архивировано 14 октября 2019 г. в Wayback Machine.
- ^ Джексон, JBC (2008). «Экологическое вымирание и эволюция в дивном новом океане» . Труды Национальной академии наук . 105 (Приложение 1): 11458–65. Бибкод : 2008PNAS..10511458J . дои : 10.1073/pnas.0802812105 . ПМК 2556419 . ПМИД 18695220 .
- ^ Соул, Майкл Э. (1986). Природоохранная биология: наука о дефиците и разнообразии . Синауэр Ассошиэйтс. п. 584. ИСБН 978-0-87893-795-0 .
Дальнейшее чтение
- Альбертс, Б.; Джонсон, А.; Льюис, Дж.; Рафф, М.; Робертс, К.; Уолтер, П. (2002). Молекулярная биология клетки (4-е изд.). Гирлянда. ISBN 978-0-8153-3218-3 . OCLC 145080076 .
- Бегон, М.; Таунсенд, Чехия; Харпер, Дж.Л. (2005). Экология: от людей к экосистемам (4-е изд.). Блэквелл Паблишинг Лимитед. ISBN 978-1-4051-1117-1 . OCLC 57639896 .
- Кэмпбелл, Нил (2004). Биология (7-е изд.). Издательская компания Бенджамина-Каммингса. ISBN 978-0-8053-7146-8 . OCLC 71890442 .
- Колинво, Поль (1979). Почему большие свирепые животные редки: взгляд эколога (переиздание). Издательство Принстонского университета. ISBN 978-0-691-02364-9 . ОСЛК 10081738 .
- Майр, Эрнст (1982). Рост биологической мысли: разнообразие, эволюция и наследование . Издательство Гарвардского университета. ISBN 978-0-674-36446-2 . Архивировано из оригинала 3 октября 2015 г. Проверено 27 июня 2015 г.
- Хоугланд, Махлон (2001). Как устроена жизнь . Jones and Bartlett Publishers Inc. ISBN 978-0-7637-1688-2 . ОСЛК 223090105 .
- Янови, Джон (2004). Как стать биологом (2-е изд.). Книги о бизонах. ISBN 978-0-8032-7620-8 . OCLC 55138571 .
- Джонсон, Джордж Б. (2005). Биология, визуализация жизни . Холт, Райнхарт и Уинстон. ISBN 978-0-03-016723-2 . ОСЛК 36306648 .
- Тобин, Аллан; Душек, Дженни (2005). Спрашивая о жизни (3-е изд.). Бельмонт, Калифорния: Уодсворт. ISBN 978-0-534-40653-0 .
Внешние ссылки
- Биология в Керли
- Филокод ОГУ
- Биология онлайн – словарь Wiki
- Серия видеолекций MIT по биологии
- OneZoom Древо жизни
Ссылки на журналы
- PLOS Biology Рецензируемый журнал с открытым доступом, издаваемый Публичной научной библиотекой.
- Current Biology : универсальный журнал, публикующий оригинальные исследования из всех областей биологии.
- Biology Letters : влиятельный журнал Королевского общества , публикующий рецензируемые статьи по биологии, представляющие общий интерес.
- Наука : всемирно известный научный журнал AAAS – см. разделы о науках о жизни.
- Международный журнал биологических наук : биологический журнал, публикующий важные рецензируемые научные статьи.
- Перспективы биологии и медицины : междисциплинарный научный журнал, публикующий эссе широкой значимости.

Элементы природы |
|---|