Телофаза
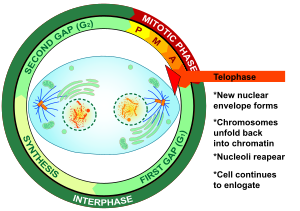
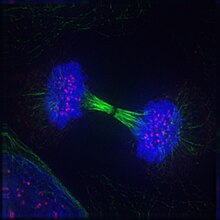
Телофаза (от древнегреческого τέλος ( télos ) «конец, результат, завершение» и φάσις (phásis) «появление») — заключительная стадия как мейоза , так и митоза в эукариотической клетке . Во время телофазы эффекты профазы и прометафазы ( распад ядрышка и ядерной мембраны) меняются местами. Когда хромосомы достигают полюсов клетки, ядерная оболочка заново собирается вокруг каждого набора хроматид , вновь появляются ядрышки , и хромосомы начинают деконденсироваться обратно в расширенный хроматин , который присутствует во время интерфазы . Митотическое веретено разбирается, а оставшиеся микротрубочки веретена деполимеризуются. Телофаза составляет примерно 2% продолжительности клеточного цикла .
Цитокинез обычно начинается до поздней телофазы. [1] и, по завершении, два дочерних ядра разделяются между парой отдельных дочерних клеток.
Телофаза в первую очередь обусловлена дефосфорилированием субстратов митотической циклин-зависимой киназы (Cdk). [2]
Дефосфорилирование субстратов Cdk
[ редактировать ]Фосфорилирование . белков-мишеней M-Cdks (митотических циклин-зависимых киназ) управляет сборкой веретена, конденсацией хромосом и разрушением ядерной оболочки на ранних стадиях митоза Дефосфорилирование этих же субстратов приводит к разборке веретена, деконденсации хромосом и реформированию дочерних ядер в телофазе. Установление степени дефосфорилирования, допускающей телофазные события, требует как инактивации Cdks, так и активации фосфатаз . [ нужна ссылка ]
Инактивация Cdk является, прежде всего, результатом разрушения связанного с ним циклина . Циклины подвергаются протеолитическому расщеплению с помощью комплекса, способствующего анафазе (APC), также известного как циклосома. [3] убиквитин-лигаза. Активный, связанный с CDC20 APC (APC/C CDC20 ) нацелен на деградацию митотических циклинов, начиная с анафазы . [4] Экспериментальное добавление неразлагаемого М-циклина к клеткам индуцирует остановку клеточного цикла в постанафазном/претелофазном состоянии с конденсированными хромосомами, сегрегированными к полюсам клетки, интактным митотическим веретеном и отсутствием реформирования ядерной оболочки. Это было показано на яйцах лягушек ( Xenopus ) , плодовых мушках ( Drosophilla melanogaster ), почкующихся ( Saccharomyces cerevisiae ) и делящихся ( Schizosaccharomyces pombe ) дрожжах, а также на нескольких линиях клеток человека. [5]
Потребность в активации фосфатазы можно увидеть у почкующихся дрожжей, которые не имеют избыточных фосфатаз для выхода из митоза и полагаются на фосфатазу cdc14 . Блокирование активации cdc14 в этих клетках приводит к такому же фенотипическому аресту, как и блокирование деградации M-циклина. [4] [2]
Исторически считалось, что анафаза и телофаза — это события, которые происходят пассивно после удовлетворения контрольной точки сборки веретена (SAC), которая определяет переход метафаза -анафаза. [6] Однако существование дифференциальных фаз активности cdc14 между анафазой и телофазой указывает на дополнительные, неизученные контрольные точки позднего митоза . Cdc14 активируется путем его высвобождения в ядро, секвестрации в ядрышке и последующего экспорта в цитоплазму. Путь раннего анафазного высвобождения Cdc-14, который стабилизирует веретено, также высвобождает cdc14 из ядрышка, но ограничивает его ядром. Полное высвобождение и поддержание активации cdc14 достигается с помощью отдельного пути сети митотического выхода (MEN) в достаточной степени (чтобы запустить разборку веретена и сборку ядерной оболочки) только после поздней анафазы. [7] [8]
Дефосфорилирование, опосредованное Cdc14, активирует нижестоящие регуляторные процессы, уникальные для телофазы. Например, дефосфорилирование CDH1 позволяет APC/C связывать CDH1. БТР/С CDH1 нацелен на CDC20 для протеолиза, что приводит к переключению клеток с APC/C CDC20 в АПК/С CDH1 активность. [5] Убиквитинирование митотических циклинов продолжается наряду с убиквитинированием APC/C. CDH1 -специфические мишени, такие как компонент митотического веретена дрожжей, Ase1, [2] и cdc5, деградация которого необходима для возврата клеток в фазу G1 . [7]
Дополнительные механизмы, управляющие телофазой
[ редактировать ]цельной клетки Сдвиг профиля фосфопротеинов является лишь самым широким из многих регуляторных механизмов, способствующих началу отдельных событий телофазы.
- Опосредованное анафазой расстояние хромосом от метафазной пластинки может запускать пространственные сигналы начала телофазы. [6]
- Важным регулятором и эффектором телофазы является cdc48 (гомологичным дрожжевому cdc48 является человеческий p97 как структурно, так и функционально), белок, который механически использует свою АТФазную активность для изменения конформации целевого белка. Cdc48 необходим для разборки веретена, сборки ядерной оболочки и деконденсации хромосом. Cdc48 модифицирует белки, структурно участвующие в этих процессах, а также некоторые убиквитинированные белки, которые таким образом направляются на протеасому. . [2] [9] [10]
Разборка митотического веретена
[ редактировать ]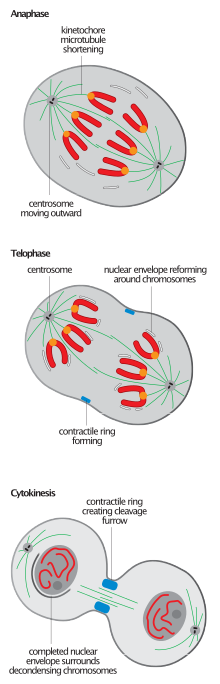
Разрыв митотического веретена, характерный для завершения митоза у всех эукариот, является событием, наиболее часто используемым для определения перехода от анафазы-B к телофазе. [2] [6] хотя начало повторной сборки ядра обычно предшествует разборке шпинделя. [11]
Разборка веретена представляет собой необратимый процесс, который должен привести не к окончательной деградации, а к реорганизации составляющих микротрубочек; микротрубочки отделяются от кинетохор и тел полюсов веретена и возвращаются в свои интерфазные состояния. [ нужна ссылка ]
Деполимеризация веретена во время телофазы происходит с плюсового конца и, таким образом, представляет собой реверс сборки веретена. [12] Последующая сборка массива микротрубочек, в отличие от сборки поляризованного веретена, является интерполярной. Это особенно очевидно в клетках животных, которые должны немедленно, после разборки митотического веретена, установить антипараллельный пучок микротрубочек, известный как центральное веретено, чтобы регулировать цитокинез. [2] АТФаза p97 необходима для создания относительно стабильных и длинных интерфазных массивов микротрубочек после разборки высокодинамичных и относительно коротких митотических. [9]
Хотя сборка веретена хорошо изучена и охарактеризована как процесс, в котором предварительные структуры формируются с помощью SAC, молекулярная основа разборки веретена не изучена в сопоставимых деталях. Поздний митотический каскад дефосфорилирования субстратов M-Cdk с помощью MEN, как широко полагают, ответственен за разборку веретена. Состояния фосфорилирования факторов, стабилизирующих и дестабилизирующих микротрубочки, а также нуклеаторов микротрубочек являются ключевыми регуляторами их активности. [9] Например, NuMA представляет собой сшивающий минус-конец белок и субстрат Cdk, диссоциация которого от микротрубочек осуществляется за счет его дефосфорилирования во время телофазы. [2]
Общая модель разборки веретена у дрожжей состоит в том, что на три функционально перекрывающихся подпроцесса: расцепление веретена, дестабилизацию и деполимеризацию в первую очередь влияют APC/C. CDH1 , киназы, специфичные для стабилизатора микротрубочек, и деполимеразы микротрубочек, направленные на плюс-конец, соответственно. Известно, что эти эффекторы высоко консервативны у дрожжей и высших эукариот. БТР/С CDH1 нацелен на сшивание белков, связанных с микротрубочками (NuMA, Ase1, Cin1 и других). AuroraB (дрожжевой IpI1) фосфорилирует связанный с веретеном стабилизирующий белок EB1 (дрожжевой Bim1), который затем диссоциирует от микротрубочек, и дестабилизатор She1, который затем связывается с микротрубочками. Кинезин8 (дрожжевой Kip3), АТФ-зависимая деполимераза, ускоряет деполимеризацию микротрубочек на плюс-конце. Было показано, что одновременное нарушение этих механизмов, но не какого-либо одного, приводит к резкой гиперстабильности веретена во время телофазы, что указывает на функциональное перекрытие, несмотря на разнообразие механизмов. [13]
Сборка ядерной оболочки
[ редактировать ]Основными компонентами ядерной оболочки являются двойная мембрана, комплексы ядерных пор и ядерная пластинка, внутренняя по отношению к внутренней ядерной мембране. Эти компоненты демонтируются во время профазы и прометафазы и реконструируются во время телофазы, когда ядерная оболочка реформируется на поверхности разделенных сестринских хроматид. [14] [15] Ядерная мембрана фрагментируется и частично поглощается эндоплазматическим ретикулумом во время прометафазы, а нацеливание везикул ЭР, содержащих белок внутренней ядерной мембраны, на хроматин происходит во время телофазы, что представляет собой обращение этого процесса. Мембранообразующие везикулы агрегируют непосредственно на поверхности хроматина, где латерально сливаются в сплошную мембрану. [2]
Ran-GTP необходим для ранней сборки ядерной оболочки на поверхности хромосом: он высвобождает компоненты оболочки, секвестрированные импортином β во время раннего митоза. Ran-GTP локализуется вблизи хромосом во время митоза, но не запускает диссоциацию белков ядерной оболочки от импортина β до тех пор, пока мишени M-Cdk не будут дефосфорилированы в телофазе. [2] Эти компоненты оболочки включают несколько компонентов ядерных пор, наиболее изученным из которых является каркасный белок ядерных пор ELYS , который может распознавать участки ДНК, богатые парами оснований A:T (in vitro), и, следовательно, может напрямую связываться с ДНК. [16] Однако эксперименты с экстрактами яиц Xenopus пришли к выводу, что ELYS не может связываться с голой ДНК и напрямую связывает только димеры гистонов и нуклеосомы. [17] После связывания с хроматином ELYS рекрутирует другие компоненты каркаса ядерных пор и трансмембранные белки ядерных пор. Комплекс ядерных пор организованно собирается и интегрируется в ядерную оболочку с последовательным добавлением Nup107-160, POM121 и FG Nups. [18]
Спорным является вопрос о том, включает ли механизм повторной сборки ядерной мембраны начальную сборку ядерных пор и последующее рекрутирование мембранных везикул вокруг пор, или же ядерная оболочка формируется в основном из расширенных цистерн ЭР, предшествующих сборке ядерных пор:
- В клетках, где ядерная мембрана фрагментируется на везикулы, не относящиеся к ER, во время митоза, Ran-GTP-зависимый путь может направлять эти дискретные популяции везикул в хроматин, где они сливаются, чтобы реформировать ядерную оболочку. [19] [16]
- В клетках, где ядерная мембрана поглощается эндоплазматической сетью во время митоза, повторная сборка включает латеральное расширение вокруг хроматина со стабилизацией расширяющейся мембраны над поверхностью хроматина. [20] Исследования, утверждающие, что этот механизм является предпосылкой образования ядерных пор, обнаружили, что связанные с голым хроматином комплексы Nup107–160 присутствуют в виде отдельных единиц, а не в виде собранных препор. [21] [16]
Оболочка сглаживается и расширяется, охватывая весь набор хроматид. в ядерные поры Вероятно, это происходит из-за импорта ламина , который может удерживаться внутри сплошной мембраны. Ядерные оболочки экстрактов яиц Xenopus не смогли сгладиться, когда ядерный импорт ламина был ингибирован, оставаясь морщинистыми и тесно связанными с конденсированными хромосомами. [22] Однако в случае латерального расширения ER импорт ядра инициируется до завершения повторной сборки ядерной оболочки, что приводит к временному внутриядерному белковому градиенту между дистальной и медиальной сторонами формирующегося ядра. [18]
Субъединицы ламина, разобранные в профазе, инактивируются и секвестрируются во время митоза. Повторная сборка ламины запускается дефосфорилированием ламина (и дополнительно метилэтерификацией остатков COOH на ламине -B ). Ламин-B может воздействовать на хроматин уже в середине анафазы. Во время телофазы, когда импорт ядра восстанавливается, ламин-А входит в реформинговое ядро, но продолжает медленно собираться в периферическую пластинку в течение нескольких часов на протяжении всей фазы G1. [16]
Экстракты яиц Xenopus и линии раковых клеток человека были основными моделями, используемыми для изучения повторной сборки ядерной оболочки. [18]
Дрожжам не хватает ламинов; их ядерная оболочка остается неповрежденной на протяжении всего митоза, а деление ядра происходит во время цитокинеза. [23] [11]
Хромосомная деконденсация
[ редактировать ]Деконденсация хромосом (также известная как релаксация или разуплотнение) в расширенный хроматин необходима для возобновления в клетке интерфазных процессов и происходит параллельно со сборкой ядерной оболочки во время телофазы у многих эукариот. [2] MEN-опосредованное дефосфорилирование Cdk необходимо для деконденсации хромосом. [2] [5]
У позвоночных деконденсация хромосом начинается только после импорта ядра восстановления . Если транспорт ламина через ядерные поры предотвращен, хромосомы остаются конденсированными после цитокинеза, и клетки не могут повторно войти в следующую S-фазу. [16] У млекопитающих лицензирование ДНК для S-фазы (ассоциация хроматина с множеством белковых факторов, необходимых для его репликации) также происходит одновременно с созреванием ядерной оболочки во время поздней телофазы. [24] [25] Это может быть объяснено восстановлением механизма ядерного импорта интерфазных ядерных и цитоплазматических локализаций белков во время телофазы и является доказательством этого.
См. также
[ редактировать ]- Цитоскелет - сеть нитевидных белков, образующая внутренний каркас клеток.
Ссылки
[ редактировать ]- ^ Рис, Джейн; Урри, Лиза; Каин, Майкл; Вассерман, Стивен; Минорский, Петр; Джексон, Роберт (2011). Кэмпбелл Биология (10-е изд.). Пирсон. ISBN 978-0-321-77565-8 .
- ↑ Перейти обратно: Перейти обратно: а б с д и ж г час я дж к Морган Д. (2007). Клеточный цикл . Лондон, Великобритания: New Science Press Ltd., стр. 154–155. ISBN 978-0-9539181-2-6 .
- ^ Хуанг Ю.Л., Хуан Дж., Питерс Дж.М., Маклафлин М.Э., Тай С.И., Пеллман Д. (февраль 1997 г.). «APC-опосредованный протеолиз Ase1 и морфогенез митотического веретена». Наука . 275 (5304): 1311–4. дои : 10.1126/science.275.5304.1311 . ПМИД 9036857 . S2CID 12265554 .
- ↑ Перейти обратно: Перейти обратно: а б Альбертс Б., Джонсон А., Льюис Дж., Морган Д., Рафф М., Робертс К., Уолтер П. (2015). Молекулярная биология клетки (6-е изд.). Нью-Йорк, штат Нью-Йорк: Garland Science, Taylor and Francisco Group. стр. 995–996. ISBN 978-0-8153-4432-2 .
- ↑ Перейти обратно: Перейти обратно: а б с Инзе Д. (2007). Контроль клеточного цикла и развитие растений . Оксфорд, Великобритания: Blackwell Publishing Ltd., стр. 99–103 . ISBN 978-1-4051-5043-9 .
- ↑ Перейти обратно: Перейти обратно: а б с Афонсу О, Матос I, Майато Х (2014). «Пространственное управление анафазно-телофазным переходом» . Клеточный цикл . 13 (19): 2985–6. дои : 10.4161/15384101.2014.959853 . ПМЦ 4614036 . ПМИД 25486554 .
- ↑ Перейти обратно: Перейти обратно: а б Монже-Касас Ф, Керальт Э (2017). Сеть митотического выхода . Нью-Йорк, штат Нью-Йорк: Humana Press. стр. 3–8. ISBN 9781493965007 .
- ^ Йеллман К.М., Редер Г.С. (2015). «Раннее высвобождение Cdc14 в анафазе, FEAR, ограничено ядром и необязательно для эффективного выхода из митоза» . ПЛОС ОДИН . 10 (6): e0128604. Бибкод : 2015PLoSO..1028604Y . дои : 10.1371/journal.pone.0128604 . ПМЦ 4474866 . ПМИД 26090959 .
- ↑ Перейти обратно: Перейти обратно: а б с Цао К., Накадзима Р., Мейер Х.Х., Чжэн Ю (октябрь 2003 г.). «ААА-АТФаза Cdc48/p97 регулирует разборку веретена в конце митоза» . Клетка . 115 (3): 355–67. дои : 10.1016/S0092-8674(03)00815-8 . ПМИД 14636562 .
- ^ Хетцер М., Мейер Х.Х., Вальтер Т.К., Бильбао-Кортес Д., Уоррен Г., Маттадж И.В. (декабрь 2001 г.). «Различные комплексы AAA-АТФаза p97 функционируют на отдельных этапах сборки ядра». Природная клеточная биология . 3 (12): 1086–91. дои : 10.1038/ncb1201-1086 . ПМИД 11781570 . S2CID 19261807 .
- ↑ Перейти обратно: Перейти обратно: а б Аист-младший (1 января 2002 г.). «Митоз и моторные белки у нитчатых аскомицетов, Nectria haematococca и некоторых родственных грибов». Международный обзор цитологии . 212 : 239–63. дои : 10.1016/S0074-7696(01)12007-3 . ISBN 9780123646163 . ПМИД 11804038 .
- ^ Вудрафф Дж. Б. (2011). Механизмы разборки и позиционирования митотического веретена у Saccharomyces cerevisiae (Диссертация). Калифорнийский университет в Беркли.
- ^ Вудрафф Дж. Б., Друбин Д. Г., Барнс Дж. (ноябрь 2010 г.). «Разборка митотического веретена происходит посредством отдельных подпроцессов, управляемых комплексом, способствующим анафазе, киназой Aurora B и кинезином-8» . Журнал клеточной биологии . 191 (4): 795–808. дои : 10.1083/jcb.201006028 . ПМК 2983061 . ПМИД 21079246 .
- ^ Яэль А., Чой Дж., ДеСэ Дж., Журуковски В., Висем Р., Рай К. (2013). Биология . Университет Райса, Хьюстон, Техас 77005: Колледж OpenStax. стр. 281–283. ISBN 978-1-938168-09-3 .
{{cite book}}: CS1 maint: местоположение ( ссылка ) - ^ Лодиш, Харви; Берк, Арнольд; Зипурски, С. Лоуренс; Мацудайра, Пол; Балтимор, Дэвид; Дарнелл, Джеймс (2000). Молекулярно-клеточная биология. 4-е издание . У. Х. Фриман. стр. Раздел 13.4.
- ↑ Перейти обратно: Перейти обратно: а б с д и Поллард Т.Д., Эрншоу В.К., Липпинкотт-Шварц Дж., Джонсон Г.Т. (2017). Клеточная биология (3-е изд.). Филадельфия, Пенсильвания: Эльзевир. стр. 770–771. ISBN 978-0-323-34126-4 .
- ^ Зирхут С., Дженнесс С., Кимура Х., Фунабики Х. (июль 2014 г.). «Нуклеосомная регуляция состава хроматина и сборки ядра, выявленная путем истощения гистонов» . Структурная и молекулярная биология природы . 21 (7): 617–25. дои : 10.1038/nsmb.2845 . ПМЦ 4082469 . ПМИД 24952593 .
- ↑ Перейти обратно: Перейти обратно: а б с Гей С., Фойани М (1 января 2015 г.). «Ядерная оболочка и хроматин, замок и ключ целостности генома». Международное обозрение клеточной и молекулярной биологии . 317 : 267–330. дои : 10.1016/bs.ircmb.2015.03.001 . ISBN 9780128022801 . ПМИД 26008788 .
- ^ Кларк П.Р., Чжан С. (2004). «Пространственный и временной контроль сборки ядерной оболочки с помощью Ran GTPase». Симпозиумы Общества экспериментальной биологии (56): 193–204. ПМИД 15565882 .
- ^ Hetzer MW (март 2010 г.). «Ядерная оболочка» . Перспективы Колд-Спринг-Харбор в биологии . 2 (3): а000539. doi : 10.1101/cshperspect.a000539 . ПМК 2829960 . ПМИД 20300205 .
- ^ Лу Л., Ладинский М.С., Кирххаузен Т. (август 2011 г.). «Формирование постмитотической ядерной оболочки из расширенных цистерн ЭР предшествует сборке ядерных пор» . Журнал клеточной биологии . 194 (3): 425–40. дои : 10.1083/jcb.201012063 . ПМК 3153650 . ПМИД 21825076 .
- ^ Визе С., Голдберг М.В., Аллен Т.Д., Уилсон К.Л. (июль 1997 г.). «Сборка ядерной оболочки в экстрактах Xenopus, визуализированная с помощью ЭМ-сканирования, обнаруживает зависимое от транспорта событие« сглаживания оболочки »». Журнал клеточной науки . 110 (13): 1489–502. дои : 10.1242/jcs.110.13.1489 . ПМИД 9224766 .
- ^ Таддеи А., Шобер Х., Гассер С.М. (август 2010 г.). «Почкующееся дрожжевое ядро» . Перспективы Колд-Спринг-Харбор в биологии . 2 (8): а000612. doi : 10.1101/cshperspect.a000612 . ПМЦ 2908769 . ПМИД 20554704 .
- ^ Димитрова Д.С., Прохорова Т.А., Блоу Дж.Дж., Тодоров И.Т., Гилберт Д.М. (январь 2002 г.). «Ядра млекопитающих получают право на репликацию ДНК во время поздней телофазы» . Журнал клеточной науки . 115 (Часть 1): 51–9. дои : 10.1242/jcs.115.1.51 . ПМК 1255924 . ПМИД 11801723 .
- ^ Фукусима К., Ван М., Найто Ю., Учихаши Т., Като Ю., Мукаи С., Ябута Н., Нодзима Х. (март 2017 г.). «GAK фосфорилируется c-Src и перемещается из центросомы в хроматин в конце телофазы» . Клеточный цикл . 16 (5): 415–427. дои : 10.1080/15384101.2016.1241916 . ПМК 5351929 . ПМИД 28135906 .
Внешние ссылки
[ редактировать ] СМИ, связанные с телофазой, на Викискладе?
СМИ, связанные с телофазой, на Викискладе?