Циклин
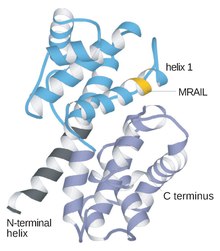
Циклины представляют собой белки, которые контролируют продвижение клетки по клеточному циклу путем активации циклин-зависимых киназ (CDK). [1]
Этимология
[ редактировать ]Циклины были первоначально обнаружены Р. Тимоти Хантом в 1982 году при изучении клеточного цикла морских ежей. [2] [3]
В интервью журналу The Life Scientific (вышедшем в эфир 13 декабря 2011 г.), которое вел Джим Аль-Халили , Р. Тимоти Хант объяснил, что название «циклин» первоначально было названо в честь его хобби, связанного с ездой на велосипеде. И только после присвоения названия его важность в клеточном цикле стала очевидной. Поскольку это было уместно, название прижилось. [4] Р. Тимоти Хант : «Кстати, название циклин, которое я придумал, на самом деле было шуткой, потому что в то время мне очень нравилось кататься на велосипеде, но они приходили и уходили в камере…» [4]
Функция
[ редактировать ]
Первоначально циклины были названы так потому, что их концентрация циклически меняется в течение клеточного цикла. (Обратите внимание, что циклины теперь классифицируются в соответствии с их консервативной структурой циклинового ящика, и не все эти циклины изменяются в уровне в течение клеточного цикла. [5] ) Колебания циклинов, а именно колебания экспрессии генов циклинов и их разрушение с помощью протеасомного пути, опосредованного убиквитином, вызывают колебания активности Cdk, управляющие клеточным циклом. Циклин образует комплекс с Cdk, который начинает активироваться, но для полной активации также требуется фосфорилирование. Cdk Образование комплекса приводит к активации активного сайта . Сами циклины не обладают ферментативной активностью, но имеют сайты связывания для некоторых субстратов и направляют Cdks в определенные субклеточные места. [5]
Циклины, связываясь с зависимыми киназами , такими как белок p34 / cdc2 / cdk1 , образуют фактор, способствующий созреванию . MPFs активируют другие белки посредством фосфорилирования . Эти фосфорилированные белки, в свою очередь, ответственны за определенные события во время деления клеток, такие как образование микротрубочек и ремоделирование хроматина . Циклины можно разделить на четыре класса в зависимости от их поведения в клеточном цикле соматических клеток позвоночных и дрожжевых клеток: циклины G1, циклины G1/S, S-циклины и M-циклины. Это деление полезно, когда речь идет о большинстве клеточных циклов, но оно не является универсальным, поскольку некоторые циклины имеют разные функции или время в разных типах клеток.
Уровень циклинов G1/S повышается в конце G1 и снижается в начале S-фазы. Комплекс Cdk-G1/S-циклин начинает индуцировать начальные процессы репликации ДНК, прежде всего, блокируя системы, предотвращающие S-фазную активность Cdk в G1. Циклины также способствуют другим действиям, способствующим развитию клеточного цикла, таким как дупликация центросом у позвоночных или тело полюса веретена у дрожжей. Увеличение присутствия G1/S-циклинов сопровождается увеличением количества S-циклинов.
Циклины G1 ведут себя не так, как другие циклины, поскольку концентрации увеличиваются постепенно (без колебаний) на протяжении клеточного цикла в зависимости от роста клеток и внешних сигналов, регулирующих рост. Присутствие G-циклинов координирует рост клеток с вступлением в новый клеточный цикл.
S-циклины связываются с Cdk, и комплекс напрямую индуцирует репликацию ДНК. Уровни S-циклинов остаются высокими не только на протяжении S-фазы, но и во время G2 и раннего митоза, а также способствуют ранним событиям митоза.
Концентрации М-циклина повышаются, когда клетка начинает вступать в митоз, и достигают пика концентрации в метафазе. Клеточные изменения в клеточном цикле, такие как сборка митотических веретен и выравнивание сестринских хроматид вдоль веретен, индуцируются комплексами M циклин-Cdk. Разрушение М-циклинов во время метафазы и анафазы, после выполнения контрольной точки сборки веретена, вызывает завершение митоза и цитокинеза. [6] Экспрессия циклинов обнаруживается иммуноцитохимически в отдельных клетках в зависимости от содержания клеточной ДНК (фаза клеточного цикла), [7] или в отношении инициации и прекращения репликации ДНК во время S-фазы, можно измерить с помощью проточной цитометрии . [8]
Вирус герпеса саркомы Капоши ( KSHV ) кодирует циклин D-типа (ORF72), который связывает CDK6 и, вероятно, способствует развитию рака, связанного с KSHV. [9]
Доменная структура
[ редактировать ]Циклины обычно сильно отличаются друг от друга по первичной структуре или аминокислотной последовательности. Однако все члены семейства циклинов схожи по 100 аминокислотам, составляющим циклиновый блок. Циклины содержат два домена с одинаковой all-α складкой , первый расположен на N-конце , а второй — на C-конце . Считается, что все циклины содержат сходную третичную структуру из двух компактных доменов 5 α-спиралей. Первым из них является консервативный циклиновый ящик, за пределами которого циклины расходятся. Например, аминоконцевые области S и M циклинов содержат короткие мотивы деструкционного бокса, которые нацелены на эти белки для протеолиза в митозе.
|
|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||

Типы
[ редактировать ]Существует несколько различных циклинов, которые активны в разных частях клеточного цикла и заставляют Cdk фосфорилировать различные субстраты. Существует также несколько «сиротских» циклинов, для которых не выявлен партнер Cdk. Например, циклин F представляет собой циклин-сироту, который необходим для перехода G 2 /M. [12] [13] Исследование C. elegans выявило особую роль митотических циклинов. [14] [15] Примечательно, что недавние исследования показали, что циклин А создает клеточную среду, которая способствует отсоединению микротрубочек от кинетохор в прометафазе, чтобы гарантировать эффективную коррекцию ошибок и точную сегрегацию хромосом. Клетки должны точно разделить свои хромосомы, и это событие основано на двунаправленном прикреплении хромосом к микротрубочкам веретена через специализированные структуры, называемые кинетохорами. На ранних стадиях деления существуют многочисленные ошибки в том, как кинетохоры связываются с микротрубочками веретена. Нестабильные прикрепления способствуют исправлению ошибок, вызывая постоянное отсоединение, перестройку и повторное прикрепление микротрубочек от кинетохор в клетках, когда они пытаются найти правильное прикрепление. Белок циклин А управляет этим процессом, поддерживая его до тех пор, пока ошибки не будут устранены. В нормальных клетках постоянная экспрессия циклина А предотвращает стабилизацию микротрубочек, связанных с кинетохорами, даже в клетках с выровненными хромосомами. По мере снижения уровня циклина А прикрепление микротрубочек становится стабильным, что позволяет хромосомам правильно делиться по мере продолжения клеточного деления. Напротив, в клетках с дефицитом циклина А прикрепление микротрубочек преждевременно стабилизируется. Следовательно, эти клетки могут не исправлять ошибки, что приводит к более высокому уровню неправильного разделения хромосом. [16]
Основные группы
[ редактировать ]Выделяют две основные группы циклинов:
- G 1 /S циклины – необходимы для контроля клеточного цикла при переходе G 1 /S ,
- Циклины G2 переходе G2 / /M – необходимы для контроля клеточного цикла при M ( митоз ). Циклины G 2 /M стабильно накапливаются в течение G 2 и резко разрушаются при выходе клеток из митоза (в конце М-фазы ).
Подтипы
[ редактировать ]Конкретные подтипы циклинов вместе с соответствующими им CDK (в скобках):
| Разновидность | Г1 | Г1/С | С | М |
|---|---|---|---|---|
| С. cerevisiae | Cln3 (Cdk1) | Клн 1,2 (Cdk1) | Клб 5,6 (Cdk1) | Клб 1,2,3,4 (Кдк 1) |
| С. помбе | Пук1? ( Кдк2 ) | Puc1, Cig1? (Кдк2) | Сигарета 2, Сигарета 1? (Кдк2) | Кдк13 (Кдк2) |
| Д. меланогастер | циклин D (Cdk4) | циклин E (Cdk2) | циклин Е, А (Cdk2,1) | циклин A, B, B3 (Cdk1) |
| Х. левис | либо неизвестно, либо нет | циклин E (Cdk2) | циклин Е, А (Cdk2,1) | циклин A, B, B3 (Cdk1) |
| Х. сапиенс | циклин D 1,2,3 ( Cdk4 , Cdk6 ) | циклин E ( Cdk2 ) | циклин А ( Cdk2 , Cdk1 ) | циклин B ( Cdk1 ) |
| семья | члены |
|---|---|
| А | CCNA1 , CCNA2 |
| Б | CCNB1 , CCNB2 , CCNB3 |
| С | CCNC |
| Д | CCND1 , CCND2 , CCND3 |
| И | CCNE1 , CCNE2 |
| Ф | ККНФ |
| Г | CCNG1 , CCNG2 |
| ЧАС | ККНХ |
| я | CCNI , CCNI2 |
| Дж | CCNJ , CCNJL |
| К | CCNK |
| л | ККНЛ1 , ККНЛ2 |
| ТО | CCNO |
| П | CCNP |
| Т | ККНТ1 , ККНТ2 |
| И | CCNY , CCNYL1 , CCNYL2 , CCNYL3 |
Другие белки, содержащие этот домен
[ редактировать ]Кроме того, следующий человеческий белок содержит домен циклина:
История
[ редактировать ]Леланд Х. Хартвелл , Р. Тимоти Хант и Пол М. Нерс получили Нобелевскую премию по физиологии и медицине 2001 года за открытие циклина и циклин-зависимой киназы. [17]
Ссылки
[ редактировать ]- ^ Галдериси У, Йори Ф.П., Джордано А. (август 2003 г.). «Регуляция клеточного цикла и нейронная дифференцировка». Онкоген . 22 (33): 5208–19. дои : 10.1038/sj.onc.1206558 . ПМИД 12910258 . S2CID 19528945 .
- ^ Эванс Т., Розенталь Э.Т., Янгблом Дж., Дистел Д., Хант Т. (июнь 1983 г.). «Циклин: белок, определяемый материнской мРНК в яйцах морских ежей, который разрушается при каждом делении дробления». Клетка . 33 (2): 389–96. дои : 10.1016/0092-8674(83)90420-8 . ПМИД 6134587 . S2CID 32305758 .
- ^ «Тим Хант — биографический» . NobelPrize.org .
- ^ Jump up to: а б «Жизнь научная» . Радио Би-би-си 4 . Би-би-си . Проверено 13 декабря 2011 г.
- ^ Jump up to: а б Морган Д. (2006). Клеточный цикл: принципы управления . Оксфорд: Издательство Оксфордского университета. ISBN 978-0-19-920610-0 .
- ^ Клют П., Пайнс Дж. (июнь 1999 г.). «Временной и пространственный контроль разрушения циклина B1 в метафазе». Природная клеточная биология . 1 (2): 82–7. дои : 10.1038/10049 . ПМИД 10559878 . S2CID 21441201 .
- ^ Даржинкевич З., Гонг Дж., Хуан Г., Ардельт Б., Траганос Ф. (сентябрь 1996 г.). «Цитометрия белков циклинов». Цитометрия . 25 (1): 1–13. doi : 10.1002/(SICI)1097-0320(19960901)25:1<1::AID-CYTO1>3.0.CO;2-N . ПМИД 8875049 .
- ^ Даржинкевич З., Чжао Х., Чжан С., Ли М.Ю., Ли Э.Ю. , Чжан З. (май 2015 г.). «Инициация и прекращение репликации ДНК во время S-фазы в отношении циклинов D1, E и A, p21WAF1, Cdt1 и субъединицы p12 ДНК-полимеразы δ, выявленной в отдельных клетках методом цитометрии» . Онкотаргет . 6 (14): 11735–50. дои : 10.18632/oncotarget.4149 . ПМЦ 4494901 . ПМИД 26059433 .
- ^ Чанг Ю, Мур П.С., Талбот С.Дж., Бошофф Ч.Х., Зарковска Т., Годден-Кент, Патерсон Х., Вайс Р.А., Миттнахт С. (август 1996 г.). «Циклин, кодируемый герпесвирусом KS» . Природа . 382 (6590): 410. Бибкод : 1996Natur.382..410C . дои : 10.1038/382410a0 . ПМИД 8684480 . S2CID 5118433 .
- ^ Браун Н.Р., Нобл М.Э., Эндикотт Дж.А., Гарман Э.Ф., Вакацуки С., Митчелл Э., Расмуссен Б., Хант Т., Джонсон Л.Н. (ноябрь 1995 г.). «Кристаллическая структура циклина А» . Структура . 3 (11): 1235–47. дои : 10.1016/S0969-2126(01)00259-3 . ПМИД 8591034 .
- ^ Дэвис Т.Г., Танна П., Мейер Л., Марко Д., Эйзенбранд Г., Эндикотт Дж.А., Нобл М.Э. (май 2001 г.). «Ингибитор связывания с активным и неактивным CDK2: кристаллическая структура CDK2-циклин А/индирубин-5-сульфонат» . Структура . 9 (5): 389–97. дои : 10.1016/S0969-2126(01)00598-6 . ПМИД 11377199 .
- ^ Фунг Т.К., Пун Р.Ю. (июнь 2005 г.). «Американские горки с митотическими циклинами». Семинары по клеточной биологии и биологии развития . 16 (3): 335–42. дои : 10.1016/j.semcdb.2005.02.014 . ПМИД 15840442 .
- ^ Карп Г (2007). Клеточная и молекулярная биология: концепции и эксперименты . Нью-Йорк: Уайли. стр. 148, 165–170 и 624–664. ISBN 978-0-470-04217-5 .
- ^ ван дер Воет М., Лорсон М.А., Шринивасан Д.Г., Беннетт К.Л., ван ден Хеувел С. (декабрь 2009 г.). «Митотические циклины C. elegans обладают различными, а также перекрывающимися функциями в сегрегации хромосом» . Клеточный цикл . 8 (24): 4091–102. дои : 10.4161/cc.8.24.10171 . ПМК 3614003 . ПМИД 19829076 .
- ^ Рахман М.М., Кипреос ET (январь 2010 г.). «Выявлена конкретная роль митотических циклинов» . Клеточный цикл . 9 (1): 22–3. дои : 10.4161/cc.9.1.10577 . ПМИД 20016257 .
- ^ Бауманн К. (ноябрь 2013 г.). «Клеточный цикл: коррекции циклина А» . Обзоры природы. Молекулярно-клеточная биология . 14 (11): 692. дои : 10.1038/nrm3680 . ПМИД 24064541 . S2CID 34397179 .
- ^ «Нобелевская премия по физиологии и медицине 2001 года» . Нобелевский фонд . Проверено 15 марта 2009 г.
Дальнейшее чтение
[ редактировать ]- Кригер М., Скотт М.П., Мацудайра П.Т., Лодиш Х.Ф., Дарнелл Дж.Э., Зипурски Л., Кайзер С., Берк А. (2004). Молекулярно-клеточная биология (Пятое изд.). Нью-Йорк: WH Freeman and CO. ISBN 0-7167-4366-3 .
Внешние ссылки
[ редактировать ]- ресурсов Eukaryotic Linear Motif Класс мотивов LIG_CYCLIN_1