ИРАК1
| ИРАК1 | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | IRAK1 , IRAK, pelle, киназа 1, связанная с рецептором интерлейкина 1 | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | Опустить : 300283 ; МГИ : 107420 ; Гомологен : 37496 ; GeneCards : IRAK1 ; ОМА : IRAK1 – ортологи | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||




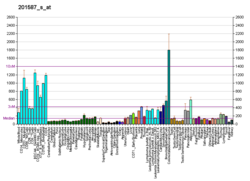
Киназа 1, ассоциированная с рецептором интерлейкина-1 ( IRAK-1), представляет собой фермент человека, кодируемый IRAK1 геном . [ 5 ] [ 6 ] IRAK-1 играет важную роль в регуляции экспрессии воспалительных генов иммунными клетками, такими как моноциты и макрофаги , которые, в свою очередь, помогают иммунной системе уничтожать бактерии, вирусы и другие патогены . IRAK-1 является частью семейства IRAK, состоящего из IRAK-1, IRAK-2, IRAK-3 и IRAK-4, и активируется воспалительными молекулами, высвобождаемыми сигнальными путями во время патогенной атаки. [ 7 ] IRAK-1 классифицируется как киназный фермент, который регулирует механизмы как врожденной , так и адаптивной иммунной системы. [ 8 ]
Структура
[ редактировать ]IRAK-1 содержит N-концевой домен смерти ( DD ), домен ProST, центрально расположенный киназный домен и C-концевой домен . DD на IRAK-1 действует как платформа взаимодействия для других DD-содержащих белков, в первую очередь для адаптерного белка, фактора миелоидной дифференцировки 88, MyD88 .
Домен proST содержит аминокислотные остатки серина, пролина и треонина и используется для облегчения взаимодействия IRAK-1 с другими членами или белками семейства IRAK. Например, аутофосфорилирование может происходить несколько раз в домене ProST, что позволяет IRAK-1 диссоциировать от MyD88, связанного с DD, сохраняя при этом взаимодействие с нижестоящими белками, такими как TNF фактор 6, связанный с рецептором ( TRAF-6 ), для инициации дальнейший путь передачи сигналов. [ 7 ]
Более того, IRAK-1 содержит инвариантный лизин внутри центрально расположенного киназного домена. Инвариантный лизин действует как сайт связывания АТФ и медиатор каталитической функции и активности киназы. [ 7 ] [ 9 ]
IRAK-1 также содержит остаток тирозина (Tyr 262 ), который конформационно изменяет активный сайт IRAK-1 путем ингибирования гидрофильного кармана позади сайта связывания и тем самым позволяет IRAK-1 оставаться в активном состоянии. Например, связывание АТФ с сайтом связывания IRAK-1 может легко происходить в присутствии Tyr. 266 , потому что Тир 266 будет занимать гидрофильный карман, где конкурентные ингибиторы АТФ могут связываться и нарушать каталитическую функцию. [ 7 ]
Активация
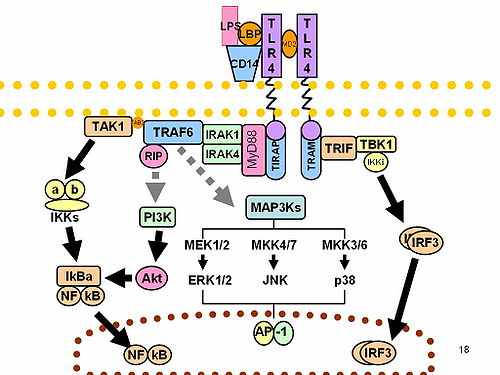
[ редактировать ]В присутствии чужеродных патогенов сигнальные пути, индуцированные IRAK-1, могут активироваться Toll-подобными рецепторами ( TLR ) или рецепторами семейства интерлейкина-1 ( IL-1R в ответ ). TLR распознают патоген-ассоциированные молекулярные паттерны ( PAMP ), экспрессируемые на бактериях, а IL-1R распознают и связывают провоспалительные цитокины семейства IL-1. И TLR, и IL-1R опосредуют сигнальный каскад, который включает связывание MyD88 с рецептором, олигомеризацию MyD88, рекрутирование IRAK-1 через DD, мультимеризацию IRAK-1 и, в конечном итоге, активацию киназы и дальнейшую последующую передачу сигналов. [ 10 ] [ 7 ]
IRAK-1 также можно активировать при взаимодействии с другими членами семьи IRAK. IRAK-1 и IRAK-4 могут активировать друг друга, используя DD в качестве платформы для MyD88. IRAK-1 IRAK-4 сначала фосфорилирует IRAK-1, который катализирует каскад автофосфорилирования , происходящий в три этапа. IRAK-1 сначала фосфорилируется по Thr209, вызывая конформационные изменения . Затем IRAK-1 фосфорилируется по Thr387, делая IRAK-1 полностью активным. Наконец, аутофосфорилирование нескольких остатков в области proST стимулирует высвобождение IRAK-1 из рецепторного комплекса. [ 7 ]
Функция
[ редактировать ]IRAK-1 кодирует киназу 1, ассоциированную с рецептором интерлейкина-1, которая представляет собой серин-треониновую протеинкиназу , которая при стимуляции связывается с рецептором интерлейкина-1 (IL1R) . IRAK-1 необходим для продукции провоспалительных цитокинов ниже сигнальных путей TLR и IL-1R. Более того, IRAK-1 отвечает за индуцированную IL1 активацию транскрипционного фактора NF-каппа B. При связывании со своим рецептором IRAK-1 активируется, как описано в разделе «Активация» , а затем диссоциирует от своего рецепторного комплекса. IRAK-1 диссоциирует от рецептора вместе с TRAF6 - лигазой убиквитина E3, которая занимает промежуточное положение между различными типами рецепторов для экзогенных или эндогенных медиаторов и активации транскрипционных ответов через пути NF-каппа B и MAPK . [ 11 ] IRAK-1 и TRAF-6 затем связываются с TAK-1-связывающим белком-1 (TAB-1), после чего следует связывание с киназой, активируемой трансформирующим фактором роста β (TAK-1), и TAB-2, образуя новый комплекс. Затем этот комплекс транслоцируется в цитоплазму , где он связывается с убиквитинлигазами, такими как убиквитин-конъюгирующий фермент- 13 UBC-13 и убиквитин-конъюгирующий фермент E2 вариант-1 (UEV-1a), что приводит к убиквитинированию и деградации TRAF-6. Затем активируется ТАК-1 и происходит фосфорилирование комплекса ингибитора киназы κB (IKK), состоящего из IKKα, IKKβ и IKKγ. MAPK также активируются в процессе. Наконец, NF-κB активируется для регулирования транскрипции провоспалительных генов. [ 7 ] Альтернативно, активация IRAK-1 пути NF-κB может регулироваться путем убиквитинирования Lys. 134 и Лис 180 . [ 12 ] [ 7 ]
Альтернативно сплайсированные варианты транскрипта, кодирующие разные изоформы, были обнаружены для гена IRAK1. [ 13 ] В настоящее время существует три варианта дифференциального сплайсинга IRAK1 — IRAK1, IRAK1b и IRAK1c. Было обнаружено, что IRAK1 подвергается сумойлированию , способствуя его перемещению в ядро, а не в цитоплазму при патогенной атаке. Примечательно, что IRAK1c остается стабильным при сумойлировании, не подвергается модификации при тех же обстоятельствах и локализуется только в цитоплазме. [ 14 ]
Киназная активность IRAK-1 не является единственным белком, участвующим в провоспалительных иммунных реакциях, однако она служит белком-адаптером, который эффективно связывает MyD88, IRAK-4, белки, взаимодействующие с toll ( TOLLIP ). [ 15 ] вместе образуют комплекс, который индуцирует IL-1R-опосредованную активацию NF-κB. [ 15 ] [ 7 ]
Регулирование
[ редактировать ]Активность IRAK-1 регулируется во время его активации и функционирования. Аутофосфорилирование играет роль в активации IRAK-1 (см. «Активация опосредованную протеасомами »), а также опосредует деградацию, , что приводит к потере белка IRAK1. [ 16 ] Альтернативно, IRAK-1 может регулироваться на уровне транскрипции . Сплайсинговый вариант IRAK-1b лишен киназной активности и устойчив к деградации, опосредованной протеасомами. Более того, сплайсированный вариант IRAK-1c имеет укороченную и, таким образом, мутированную последовательность на С-конце своего киназного домена и действует как негативный регулятор сигнальных путей TLR и IL-1R. [ 7 ] [ 16 ]
Взаимодействия
[ редактировать ]Было показано, что IRAK1 взаимодействует со следующими белками:
Клиническое значение
[ редактировать ]Передача сигналов IRAK-1 участвует в ревматоидном артрите . [ 35 ] [ 36 ] Более того, IRAK-1 играет значительную роль в развитии рака.
Ссылки
[ редактировать ]- ^ Jump up to: а б с GRCh38: Версия Ensembl 89: ENSG00000184216 – Ensembl , май 2017 г.
- ^ Jump up to: а б с GRCm38: выпуск Ensembl 89: ENSMUSG00000031392 – Ensembl , май 2017 г.
- ^ «Ссылка на Human PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Ссылка на Mouse PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ Jump up to: а б Музио М., Ни Дж., Фэн П., Диксит В.М. (ноябрь 1997 г.). «Член семейства IRAK (Pelle) IRAK-2 и MyD88 как проксимальные медиаторы передачи сигналов IL-1». Наука . 278 (5343): 1612–1615. Бибкод : 1997Sci...278.1612M . дои : 10.1126/science.278.5343.1612 . ПМИД 9374458 .
- ^ Цао З, Хензель В.Дж., Гао X (февраль 1996 г.). «IRAK: киназа, связанная с рецептором интерлейкина-1». Наука . 271 (5252): 1128–1131. Бибкод : 1996Sci...271.1128C . дои : 10.1126/science.271.5252.1128 . ПМИД 8599092 . S2CID 42977425 .
- ^ Jump up to: а б с д и ж г час я дж Джайн А., Качановска С., Давила Э. (2014). «Передача сигналов киназы, связанной с рецептором IL-1, и ее роль в воспалении, прогрессировании рака и резистентности к терапии» . Границы в иммунологии . 5 : 553. дои : 10.3389/fimmu.2014.00553 . ПМЦ 4233944 . ПМИД 25452754 .
- ^ Дэн Й., Хан Б.Х., Цао Б.П. (2013). «Системная красная волчанка». Принципы и практика медицинской генетики Эмери и Римоэна : 1–22. дои : 10.1016/B978-0-12-383834-6.00081-1 . ISBN 9780123838346 .
- ^ Ван З., Веше Х., Стивенс Т., Уокер Н., Йе У.К. (май 2009 г.). «Ингибиторы ИРАК-4 при воспалении» . Актуальные темы медицинской химии . 9 (8): 724–737. дои : 10.2174/156802609789044407 . ПМК 3182414 . ПМИД 19689377 .
- ^ Ван Л., Цяо К., Феррао Р., Шен С., Хэтчер Дж.М., Бурлаге С.Дж. и др. (декабрь 2017 г.). «Кристаллическая структура IRAK1 человека» . Труды Национальной академии наук Соединенных Штатов Америки . 114 (51): 13507–13512. Бибкод : 2017PNAS..11413507W . дои : 10.1073/pnas.1714386114 . ПМЦ 5754798 . ПМИД 29208712 .
- ^ Дайничи Т., Мацумото Р., Мостафа А., Кабашима К. (2019). «Иммунный контроль с помощью TRAF6-опосредованных путей эпителиальных клеток в EIME (эпителиальной иммунной микросреде)» . Границы в иммунологии . 10 : 1107. дои : 10.3389/fimmu.2019.01107 . ПМК 6532024 . ПМИД 31156649 .
- ^ Конг Ф., Лю З., Джайн В.Г., Шима К., Сузуки Т., Маглия Л.Дж. и др. (ноябрь 2017 г.). «Ингибирование убиквитинирования IRAK1 определяет чувствительность к глюкокортикоидам при TLR9-индуцированном воспалении в макрофагах» . Журнал иммунологии . 199 (10): 3654–3667. doi : 10.4049/jimmunol.1700443 . ПМЦ 5672817 . ПМИД 29038250 .
- ^ «Ген Энтрез: киназа 1, ассоциированная с рецептором интерлейкина-1 IRAK1» .
- ^ Су Дж, Рихтер К, Чжан С, Гу Ц, Ли Л (февраль 2007 г.). «Дифференциальная регуляция вариантов сплайсинга киназы 1, связанной с рецептором интерлейкина-1 (IRAK1)». Молекулярная иммунология . 44 (5): 900–905. дои : 10.1016/j.molimm.2006.03.021 . ПМИД 16690127 .
- ^ Jump up to: а б «Белок, взаимодействующий с tollIP [Homo sapiens (человек)] - Ген - NCBI» . www.ncbi.nlm.nih.gov . Проверено 5 марта 2022 г.
- ^ Jump up to: а б Готтипати С., Рао Н.Л., Фунг-Леунг В.П. (февраль 2008 г.). «IRAK1: критический сигнальный медиатор врожденного иммунитета». Сотовая сигнализация . 20 (2): 269–276. doi : 10.1016/j.cellsig.2007.08.009 . ПМИД 17890055 .
- ^ Jump up to: а б с д Виндхейм М., Стаффорд М., Пегги М., Коэн П. (март 2008 г.). «Интерлейкин-1 (IL-1) индуцирует связанное с Lys63 полиубиквитинирование киназы 1, связанной с рецептором IL-1, чтобы облегчить связывание NEMO и активацию киназы IkappaBalpha» . Молекулярная и клеточная биология . 28 (5): 1783–1791. дои : 10.1128/MCB.02380-06 . ПМК 2258775 . ПМИД 18180283 .
- ^ Jump up to: а б Виг Э, Грин М, Лю Ю, Ю КЮ, Квон Х.Дж., Тиан Дж. и др. (март 2001 г.). «SIMPL представляет собой специфичный для фактора некроза опухоли регулятор активности ядерного фактора каппаВ» . Журнал биологической химии . 276 (11): 7859–7866. дои : 10.1074/jbc.M010399200 . ПМИД 11096118 .
- ^ Jump up to: а б Конзе Д.Б., Ву С.Дж., Томас Дж.А., Ландстрем А., Эшвелл Дж.Д. (май 2008 г.). «Lys63-связанное полиубиквитинирование IRAK-1 необходимо для активации NF-kappaB, опосредованной рецептором интерлейкина-1 и толл-подобным рецептором» . Молекулярная и клеточная биология . 28 (10): 3538–3547. дои : 10.1128/MCB.02098-07 . ПМЦ 2423148 . ПМИД 18347055 .
- ^ Чен BC, Ву WT, Хо FM, Линь WW (июль 2002 г.). «Ингибирование индуцированной интерлейкином-1бета активации NF-каппа B кальций/кальмодулин-зависимой протеинкиназной киназой происходит посредством активации Akt, связанной с фосфорилированием киназы, связанной с рецептором интерлейкина-1, и разобщением MyD88» . Журнал биологической химии . 277 (27): 24169–24179. дои : 10.1074/jbc.M106014200 . ПМИД 11976320 .
- ^ Jump up to: а б Веше Х., Гао Х, Ли Х, Киршнинг С.Дж., Старк Г.Р., Цао З. (июль 1999 г.). «IRAK-M является новым членом семейства киназ, ассоциированных с рецептором интерлейкина-1 (IRAK)» . Журнал биологической химии . 274 (27): 19403–19410. дои : 10.1074/jbc.274.27.19403 . ПМИД 10383454 .
- ^ Jump up to: а б Ли С., Стрелов А., Фонтана Э.Дж., Веше Х. (апрель 2002 г.). «ИРАК-4: новый член семейства IRAK со свойствами IRAK-киназы» . Труды Национальной академии наук Соединенных Штатов Америки . 99 (8): 5567–5572. Бибкод : 2002PNAS...99.5567L . дои : 10.1073/pnas.082100399 . ПМК 122810 . ПМИД 11960013 .
- ^ Фицджеральд К.А., Палссон-Макдермотт Э.М., Боуи А.Г., Джеффрис К.А., Мэнселл А.С., Брейди Г. и др. (сентябрь 2001 г.). «Mal (похожий на MyD88-адаптер) необходим для передачи сигнала Toll-подобного рецептора-4». Природа . 413 (6851): 78–83. Бибкод : 2001Natur.413...78F . дои : 10.1038/35092578 . ПМИД 11544529 . S2CID 4333764 .
- ^ Jump up to: а б с «Ген IRAK1 | Белок IRAK1 | Антитело IRAK1» . База данных генов человека GeneCards .
- ^ Цянь Ю, Коммане М, Ниномия-Цудзи Дж, Мацумото К, Ли Х (ноябрь 2001 г.). «IRAK-опосредованная транслокация TRAF6 и TAB2 при индуцированной интерлейкином-1 активации NFkappa B» . Журнал биологической химии . 276 (45): 41661–41667. дои : 10.1074/jbc.M102262200 . ПМИД 11518704 .
- ^ Цао З, Сюн Дж, Такеучи М, Курама Т, Гёддел Д.В. (октябрь 1996 г.). «TRAF6 является преобразователем сигнала интерлейкина-1». Природа . 383 (6599): 443–446. Бибкод : 1996Natur.383..443C . дои : 10.1038/383443a0 . ПМИД 8837778 . S2CID 4269027 .
- ^ Такацуна Х., Като Х., Года Дж., Акияма Т., Мория А., Окамото Ю. и др. (апрель 2003 г.). «Идентификация TIFA как адаптерного белка, который связывает фактор 6, связанный с рецептором фактора некроза опухоли (TRAF6), с киназой-1, ассоциированной с рецептором интерлейкина-1 (IL-1) (IRAK-1) в передаче сигналов рецептора IL-1» . Журнал биологической химии . 278 (14): 12144–12150. дои : 10.1074/jbc.M300720200 . ПМИД 12566447 .
- ^ Линг Л., Гёддел Д.В. (август 2000 г.). «T6BP, белок, взаимодействующий с TRAF6, участвующий в передаче сигналов IL-1» . Труды Национальной академии наук Соединенных Штатов Америки . 97 (17): 9567–9572. Бибкод : 2000PNAS...97.9567L . дои : 10.1073/pnas.170279097 . ПМК 16905 . ПМИД 10920205 .
- ^ Такаесу Г., Ниномия-Цудзи Дж., Кисида С., Ли Х, Старк Г.Р., Мацумото К. (апрель 2001 г.). «Киназа, связанная с рецептором интерлейкина-1 (IL-1), приводит к активации TAK1, индуцируя транслокацию TAB2 в сигнальном пути IL-1» . Молекулярная и клеточная биология . 21 (7): 2475–2484. дои : 10.1128/MCB.21.7.2475-2484.2001 . ПМК 86880 . ПМИД 11259596 .
- ^ Чен Ф., Ду Ю, Чжан З, Чен Г, Чжан М, Шу Х.Б. и др. (апрель 2008 г.). «Синтенин отрицательно регулирует TRAF6-опосредованную передачу сигналов IL-1R/TLR4». Сотовая сигнализация . 20 (4): 666–674. doi : 10.1016/j.cellsig.2007.12.002 . ПМИД 18234474 . S2CID 28812149 .
- ^ Ньютон К., Мацумото М.Л., Вертц И.Е., Киркпатрик Д.С., Лилль Дж.Р., Тан Дж. и др. (август 2008 г.). «Редактирование цепи убиквитина, выявленное с помощью антител, специфичных к полиубиквитиновой связи» . Клетка . 134 (4): 668–678. дои : 10.1016/j.cell.2008.07.039 . ПМИД 18724939 .
- ^ Сяо Х., Цянь В., Сташке К., Цянь Ю., Цуй Г., Дэн Л. и др. (май 2008 г.). «Пеллино 3b отрицательно регулирует индуцированную интерлейкином-1 TAK1-зависимую активацию NF kappaB» . Журнал биологической химии . 283 (21): 14654–14664. дои : 10.1074/jbc.M706931200 . ПМК 2386918 . ПМИД 18326498 .
- ^ Бернс К., Клатворти Дж., Мартин Л., Мартинон Ф., Пламптон С., Маскера Б. и др. (июнь 2000 г.). «Толлип, новый компонент пути IL-1RI, связывает IRAK с рецептором IL-1». Природная клеточная биология . 2 (6): 346–351. дои : 10.1038/35014038 . ПМИД 10854325 . S2CID 32036101 .
- ^ Хэ Х, Цзин З, Ченг Г (2014). «МикроРНК: новые регуляторы сигнальных путей Toll-подобных рецепторов» . БиоМед Исследования Интернэшнл . 2014 : 945169. doi : 10.1155/2014/945169 . ПМЦ 3977468 . ПМИД 24772440 .
- ^ «Ген IRAK1» . Домашний справочник по генетике . Национальная медицинская библиотека США. Архивировано из оригинала 20 февраля 2017 года.
- ^ Сингер Дж.В., Флейшман А., Аль-Файюми С., Маскаренхас Ж.О., Ю К., Агарвал А. (сентябрь 2018 г.). «Ингибирование киназы 1, связанной с рецептором интерлейкина-1 (IRAK1), как терапевтическая стратегия» . Онкотаргет . 9 (70): 33416–33439. дои : 10.18632/oncotarget.26058 . ПМК 6161786 . ПМИД 30279971 .
Дальнейшее чтение
[ редактировать ]- Аурон ЧП (1999). «Рецептор интерлейкина 1: взаимодействие лигандов и передача сигнала». Обзоры цитокинов и факторов роста . 9 (3–4): 221–237. дои : 10.1016/S1359-6101(98)00018-5 . ПМИД 9918122 .
- Цао З, Сюн Дж, Такеучи М, Курама Т, Гёддел Д.В. (октябрь 1996 г.). «TRAF6 является преобразователем сигнала интерлейкина-1». Природа . 383 (6599): 443–446. Бибкод : 1996Natur.383..443C . дои : 10.1038/383443a0 . ПМИД 8837778 . S2CID 4269027 .
- Бреннер В., Ньякатура Г., Розенталь А., Платцер М. (август 1997 г.). «Геномная организация двух новых генов человека Xq28: компактное расположение гамма-IDH и дельта-TRAP консервативно у крыс и мышей». Геномика . 44 (1): 8–14. дои : 10.1006/geno.1997.4822 . ПМИД 9286695 .
- Хуан Дж, Гао X, Ли С, Цао Z (ноябрь 1997 г.). «Для привлечения IRAK в комплекс рецепторов интерлейкина 1 требуется дополнительный белок рецептора интерлейкина 1» . Труды Национальной академии наук Соединенных Штатов Америки . 94 (24): 12829–12832. Бибкод : 1997PNAS...9412829H . дои : 10.1073/pnas.94.24.12829 . ПМЦ 24223 . ПМИД 9371760 .
- Муцио М, Натоли Г, Саккани С, Левреро М, Мантовани А (июнь 1998 г.). «Сигнальный путь человека: расхождение ядерного фактора kappaB и активации JNK/SAPK выше фактора 6, связанного с рецептором фактора некроза опухоли (TRAF6)» . Журнал экспериментальной медицины . 187 (12): 2097–2101. дои : 10.1084/jem.187.12.2097 . ПМК 2212359 . ПМИД 9625770 .
- Маскера Б., Рэй К., Бернс К., Вольпе Ф. (апрель 1999 г.). «Сверхэкспрессия ферментативно неактивной киназы, связанной с рецептором интерлейкина-1, активирует ядерный фактор-каппаВ» . Биохимический журнал . 339 (2): 227–231. дои : 10.1042/0264-6021:3390227 . ПМК 1220149 . ПМИД 10191251 .
- Веше Х., Гао Х, Ли Х, Киршнинг С.Дж., Старк Г.Р., Цао З. (июль 1999 г.). «IRAK-M является новым членом семейства киназ, ассоциированных с рецептором интерлейкина-1 (IRAK)» . Журнал биологической химии . 274 (27): 19403–19410. дои : 10.1074/jbc.274.27.19403 . ПМИД 10383454 .
- Ян Р.Б., Марк М.Р., Герни А.Л., Годовски П.Дж. (июль 1999 г.). «Сигнальные события, индуцированные липополисахарид-активируемым толл-подобным рецептором 2» . Журнал иммунологии . 163 (2): 639–643. дои : 10.4049/jimmunol.163.2.639 . ПМИД 10395652 .
- Томас Дж.А., Аллен Дж.Л., Цен М., Дубникофф Т., Данао Дж., Ляо XC и др. (июль 1999 г.). «Нарушение передачи сигналов цитокинов у мышей, у которых отсутствует киназа, связанная с рецептором IL-1» . Журнал иммунологии . 163 (2): 978–984. дои : 10.4049/jimmunol.163.2.978 . ПМИД 10395695 .
- Райхвальд К., Тизен Дж., Вие Т., Вайцель Дж., Пустка В.А., Розенталь А. и др. (март 2000 г.). «Сравнительный анализ последовательностей локуса MECP2 у человека и мыши выявляет новые транскрибируемые области». Геном млекопитающих . 11 (3): 182–190. дои : 10.1007/s003350010035 . ПМИД 10723722 . S2CID 15901911 .
- Бернс К., Клатворти Дж., Мартин Л., Мартинон Ф., Пламптон С., Маскера Б. и др. (июнь 2000 г.). «Толлип, новый компонент пути IL-1RI, связывает IRAK с рецептором IL-1». Природная клеточная биология . 2 (6): 346–351. дои : 10.1038/35014038 . ПМИД 10854325 . S2CID 32036101 .
- Бёль Г., Кройцер О.Дж., Бригелиус-Флоэ Р. (июль 2000 г.). «Транслокация киназы-1, связанной с рецептором интерлейкина-1 (IRAK-1), в ядро» . Письма ФЭБС . 477 (1–2): 73–78. дои : 10.1016/S0014-5793(00)01759-2 . ПМИД 10899313 .
- Хартли Дж.Л., Темпл Г.Ф., Браш Массачусетс (ноябрь 2000 г.). «Клонирование ДНК с использованием сайт-специфической рекомбинации in vitro» . Геномные исследования . 10 (11): 1788–1795. дои : 10.1101/гр.143000 . ПМК 310948 . ПМИД 11076863 .
- Виг Э, Грин М, Лю Ю, Ю КЮ, Квон Х.Дж., Тиан Дж. и др. (март 2001 г.). «SIMPL представляет собой специфичный для фактора некроза опухоли регулятор активности ядерного фактора каппаВ» . Журнал биологической химии . 276 (11): 7859–7866. дои : 10.1074/jbc.M010399200 . ПМИД 11096118 .
- Ли X, Комман М, Цзян З, Старк Г.Р. (апрель 2001 г.). «IL-1-индуцированная активация NFkappa B и N-концевой киназы c-Jun (JNK) расходится с киназой, ассоциированной с рецептором IL-1 (IRAK)» . Труды Национальной академии наук Соединенных Штатов Америки . 98 (8): 4461–4465. Бибкод : 2001PNAS...98.4461L . дои : 10.1073/pnas.071054198 . ПМК 31857 . ПМИД 11287640 .
- Дженсен Л.Е., Уайтхед А.С. (август 2001 г.). «IRAK1b, новый альтернативный вариант сплайсинга киназы, ассоциированной с рецептором интерлейкина-1 (IRAK), опосредует передачу сигналов интерлейкина-1 и обладает длительной стабильностью» . Журнал биологической химии . 276 (31): 29037–29044. дои : 10.1074/jbc.M103815200 . ПМИД 11397809 .
- Цянь Ю, Коммане М, Ниномия-Цудзи Дж, Мацумото К, Ли Х (ноябрь 2001 г.). «IRAK-опосредованная транслокация TRAF6 и TAB2 при индуцированной интерлейкином-1 активации NFkappa B» . Журнал биологической химии . 276 (45): 41661–41667. дои : 10.1074/jbc.M102262200 . ПМИД 11518704 .