Толл-подобный рецептор 4
| ТЛР4 | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Идентификаторы | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | TLR4 , ARMD10, CD284, TLR-4, TOLL, толл-подобный рецептор 4 | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | Опустить : 603030 ; МГИ : 96824 ; Гомологен : 41317 ; Генные карты : TLR4 ; ОМА : TLR4 — ортологи | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||




Toll-подобный рецептор 4 (TLR4), также обозначаемый как CD284 ( кластер дифференцировки 284), является ключевым активатором врожденного иммунного ответа и играет центральную роль в борьбе с бактериальными инфекциями. TLR4 представляет собой трансмембранный белок массой около 95 кДа , кодируемый TLR4 геном .
TLR4 принадлежит к семейству толл-подобных рецепторов , которое является представителем рецепторов распознавания образов (PRR), названных так из-за их способности распознавать эволюционно консервативные компоненты микроорганизмов (бактерий, вирусов, грибов и паразитов), называемые молекулярными паттернами, связанными с патогенами (PAMP). ). Распознавание PAMP PRR вызывает быструю активацию врожденного иммунитета, необходимого для борьбы с инфекционными заболеваниями. [ 5 ]
TLR4 экспрессируется в иммунных клетках преимущественно миелоидного происхождения, включая моноциты, макрофаги и дендритные клетки (ДК). [ 5 ] Он также экспрессируется на более низком уровне на некоторых неиммунных клетках, включая эпителий, эндотелий, плацентарные клетки и бета-клетки островков Лангерганса. Большинство миелоидных клеток экспрессируют также большое количество CD14 , закрепленного на плазматической мембране , что облегчает активацию TLR4 с помощью LPS и контролирует последующую интернализацию LPS-активированного TLR4, важного для передачи сигналов и деградации рецептора. [ 6 ] [ 7 ]
Основными лигандами TLR4 являются липополисахариды (ЛПС), основные компоненты внешней мембраны грамотрицательных бактерий и некоторых грамположительных бактерий . TLR4 также может быть активирован эндогенными соединениями, называемыми молекулярными паттернами, связанными с повреждением ( DAMP ), включая белок 1 группового бокса с высокой подвижностью ( HMGB1 ), белки S100 или гистоны . Эти соединения высвобождаются при повреждении тканей, а также при отмирании или некрозе клеток. [ 8 ] [ 9 ] [ 10 ] [ 11 ] [ 12 ]
Функция
[ редактировать ]Первой функцией, описанной для TLR4, было распознавание экзогенных молекул патогенов (PAMP), в частности молекул ЛПС грамотрицательных бактерий. [ 13 ] Как рецептор распознавания образов , TLR4 играет фундаментальную роль в распознавании патогенов и активации врожденного иммунитета , который является первой линией защиты от вторжения микроорганизмов. Во время инфекции TLR4 реагирует на ЛПС, присутствующий в тканях и кровотоке, и запускает провоспалительные реакции, способствующие уничтожению вторгшихся бактерий. [ 13 ]
TLR4 также участвует в распознавании эндогенных молекул DAMP, что приводит к иным результатам передачи сигналов, чем PAMP, как количественно, так и качественно. [ 14 ] [ 12 ] DAMP могут активировать TLR4 в неинфекционных условиях, вызывая восстановление тканей и активацию преимущественно провоспалительных реакций. [ 8 ] [ 9 ] [ 10 ] [ 11 ] [ 12 ] Обычно воспаление играет защитную роль. Это сложный и скоординированный процесс, за которым следуют пути разрешения, которые восстанавливают целостность и функцию тканей. Однако в некоторых случаях чрезмерная и/или плохо регулируемая воспалительная реакция на DAMP может быть губительной для организма, ускоряя развитие или прогрессирование патологий, таких как ряд раковых и нейродегенеративных заболеваний (как обсуждается ниже).
TLR4 связывает ЛПС с помощью LPS-связывающего белка (LBP) и CD14, а также незаменимого вклада белка MD-2, стабильно связанного с внеклеточным фрагментом рецептора. [ 15 ] Передача сигналов TLR4 реагирует на сигналы путем формирования комплекса с использованием внеклеточного домена повторов, богатого лейцином (LRR) и внутриклеточного домена рецептора toll/интерлейкина-1 (TIR). Стимуляция ЛПС индуцирует ряд взаимодействий с несколькими вспомогательными белками, которые образуют комплекс TLR4 на поверхности клетки. Распознавание ЛПС инициируется связыванием ЛПС с белком LBP . Этот комплекс LPS-LBP переносит LPS на CD14 , который представляет собой гликозилфосфатидилинозитол-заякоренный мембранный белок, который связывает комплекс LPS-LBP и облегчает перенос LPS на белок MD-2 , который связан с внеклеточным доменом TLR4. Связывание ЛПС способствует димеризации комплекса TLR4/MD-2. Конформационные изменения TLR4 вызывают рекрутирование внутриклеточных адаптерных белков, содержащих домен TIR, который необходим для активации нижестоящего сигнального пути.
В связывании молекулы ЛПС с комплексом TLR4/MD-2 участвуют ацильные цепи и фосфатные группы липида А, консервативной части ЛПС и основного индуктора провоспалительных ответов на ЛПС. [ 16 ] [ 17 ]
На активацию TLR4 и реакцию на ЛПС большое влияние оказывают полисахаридный домен и молекулярная структура фрагмента липида А молекул ЛПС. Гексацилированные и дифосфорилированные ЛПС, такие как ЛПС Escherichia coli (O111:B4), являются одними из наиболее мощных агонистов TLR4, тогда как недостаточно ацилированные и дефосфорилированные виды ЛПС обладают более слабой провоспалительной активностью, особенно в клетках человека. [ 18 ] Структурные детерминанты этого явления обнаружены в комплексе TLR4/MD-2, а также в белке CD14. [ 16 ] [ 19 ] Полисахаридная часть, ковалентно связанная с липидом А, также играет незаменимую роль в активации TLR4 посредством CD14/TLR4/MD-2. [ 20 ] Однако, помимо домена липида А, важную роль в связывании и активации молекул ЛПС играет полисахаридный фрагмент, поскольку было показано, что один только фрагмент липида А значительно менее активен, чем полная молекула ЛПС. [ 21 ]
Сигнализация
[ редактировать ]В отличие от всех других TLR, стимуляция TLR4 запускает два сигнальных пути, называемых MyD88 -зависимым и TRIF -зависимым, после адаптерных белков, участвующих в их индукции. [ 22 ] MyD88-зависимая передача сигналов запускается TLR4, локализованным на плазматической мембране, тогда как TRIF-зависимая сигнализация запускается TLR4, интернализованным в эндосомах.
Эти сигнальные пути приводят к выработке двух наборов цитокинов. MyD88-зависимый путь индуцирует выработку провоспалительных цитокинов, тогда как TRIF-зависимый путь индуцирует выработку интерферонов I типа и хемокинов. [ 22 ] [ 23 ] Молекулярная структура лигандов TLR4 (в частности, ЛПС), а также их комплексообразование с белками или липидами сильно влияют на действие этих сигнальных путей, связанных с TLR4, что приводит к различным балансам цитокинов. [ 24 ] [ 25 ] [ 26 ] [ 27 ]
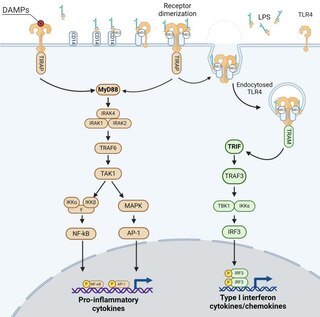
MyD88 – зависимый путь
[ редактировать ]MyD88-зависимый путь регулируется двумя адаптер-ассоциированными белками: геном 88 первичного ответа миелоидной дифференцировки ( MyD88 ) и адаптерным белком, содержащим домен TIR ( TIRAP ). Он также включает активацию киназ, ассоциированных с рецептором IL-1 ( IRAK ), и молекул-адаптеров фактора 6, ассоциированного с рецептором TNF ( TRAF6 ). TRAF6 индуцирует активацию TAK1 (активируемой трансформирующим фактором роста-β киназы 1), что приводит к активации каскадов MAPK (митоген-активируемой протеинкиназы) и киназ IκB ( IKK ), называемых IKKα и IKKβ. [ 28 ] Сигнальный путь IKKs приводит к индукции транскрипционного фактора NF-κB , тогда как активация каскадов MAPK приводит к активации другого транскрипционного фактора AP-1 . [ 28 ] [ 29 ] Эти два фактора транскрипции вызывают экспрессию генов, кодирующих провоспалительные медиаторы, такие как фактор некроза опухоли α (TNF-α), интерлейкин (IL)-6 и интерфероны типа III (IFNλ1/2). [ 30 ] [ 31 ] [ 32 ]
TRIF – зависимый путь
[ редактировать ]TRIF-зависимый путь включает интернализацию TLR4 в эндосомах и рекрутирование адаптерных белков, содержащих TIR-домен адаптера, индуцирующего интерферон-β ( TRIF ) и TRIF-родственную адаптерную молекулу (TRAM). Сигналы TRAM-TRIF активируют убиквитинлигазу TRAF3 с последующей активацией неканонических киназ IKK: TANK-связывающей киназы 1 (TBK1) и IKKε. TBK1 фосфорилирует консенсусный мотив pLxIS TRIF, который необходим для рекрутирования регуляторного фактора интерферона (IRF) 3. IRF3 также фосфорилируется TBK1, а затем диссоциирует от TRIF, димеризуется и перемещается в ядро. [ 33 ] Наконец, IRF3 индуцирует экспрессию генов, кодирующих IFN типа I, таких как интерферон бета (IFN-β), хемокин CCL5/RANTES и гены, регулируемые интерфероном, например, кодирующие хемокин CXCL10/IP-10. [ 30 ] [ 31 ] [ 32 ] [ 34 ] Известно, что TRIF-зависимый сигнальный путь TLR4 играет центральную роль в стимуляции клеток врожденного иммунитета, таких как макрофаги, созревании ДК, а также индукции и рекрутирования адаптивных иммунных ответов Th1. [ 35 ]
Активация иммунных клеток
[ редактировать ]Активация TLR4 с помощью ЛПС обеспечивает быструю стимуляцию широкого спектра клеток врожденного иммунитета, таких как макрофаги и ДК. Это приводит к секреции провоспалительных интерферонов I типа, цитокинов, хемокинов. Уровни продукции этих цитокинов/хемокинов варьируются в зависимости от степени активации сигнальных путей MyD88 и TRIF молекулами-агонистами TLR4. Активация TLR4 также вызывает стимуляцию презентации антигена и активацию костимулирующих молекул (таких как CD40 , CD80 и CD86 ) на клетках врожденного иммунитета, которые необходимы для презентации антигена Т-лимфоцитам. [ 36 ] [ 37 ] Это объясняет, почему известно, что активация TLR4 с помощью LPS также стимулирует генерацию эффективных адаптивных иммунных ответов и индуцирует их рекрутирование, поляризацию и поддержание через панель продуцируемых цитокинов и хемокинов. [ 37 ] [ 22 ]
Сигнальные пути TRIF и MyD88 оказывают разное, но взаимодополняющее влияние на активацию иммунных клеток. Было показано, что стимуляция макрофагов строго зависит от активации пути TRIF, тогда как активация и созревание DC зависят как от путей MyD88, так и от TRIF. [ 38 ] [ 39 ] [ 40 ] [ 41 ] Повышенная экспрессия костимулирующих молекул и молекул MHC является признаком созревания DC, необходимого для презентации антигена этими клетками. [ 42 ] Однако были обнаружены существенные различия в сигнальных путях, приводящих к этому явлению. В макрофагах активация костимулирующих молекул строго зависит от TRIF-зависимого пути, тогда как в ДК участвуют как MyD88-, так и TRIF-зависимый путь. [ 43 ] [ 44 ] [ 22 ] [ 45 ] Повышенное присутствие на клеточной поверхности костимулирующих молекул, а также MHC II является признаком созревания DC, необходимого для презентации антигена этими клетками. [ 46 ]
Также было обнаружено, что активация сигнальных путей MyD88 и TRIF индуцирует поляризацию Th1 ответов Т-клеток посредством созревания DC и панели продуцируемых цитокинов. [ 47 ] [ 48 ] [ 49 ] Однако низкая активация пути MYD88 важна для эффективной дифференцировки цитотоксических Т-клеток, способствуя слиянию эндосом, несущих MHC I, с фагосомами, обеспечивающими перекрестную презентацию антигенов. [ 47 ] Напротив, мощная активация пути MYD88 индуцирует чрезмерную выработку провоспалительных цитокинов, что приводит к опасным для жизни патологическим последствиям, таким как цитокиновый шторм.
Влияние активации TLR4 на врожденную и адаптивную иммунную систему объясняет, почему агонисты TLR4, такие как производные ЛПС, были разработаны в качестве адъювантов для вакцин. Среди них — монофосфорилированный липид А (MPL) компании GSK, детоксицированный липид А, полученный из ЛПС сальмонеллы, который является первым и единственным природным иммуностимулятором, одобренным в качестве адъюванта в пяти человеческих вакцинах. [ 50 ] [ 51 ] [ 52 ]
Эволюционная история и полиморфизм
[ редактировать ]TLR4 возник, когда TLR2 и TLR4 разошлись около 500 миллионов лет назад, в начале эволюции позвоночных. [ 53 ] Выравнивание последовательностей экзонов TLR4 человека и человекообразных обезьян показало, что в человеческом TLR4 не произошло значительной эволюции с момента нашего расхождения с нашим последним общим предком с шимпанзе; Экзоны TLR4 человека и шимпанзе различаются только тремя заменами, тогда как люди и павианы сходны на 93,5% во внеклеточном домене. [ 54 ] Примечательно, что люди обладают большим количеством ранних стоп-кодонов в TLR4, чем человекообразные обезьяны; в исследовании 158 человек по всему миру 0,6% имели нонсенс-мутацию. [ 55 ] [ 56 ] Это говорит о том, что на TLR4 человека оказывается более слабое эволюционное давление, чем на наших родственников-приматов. Распределение полиморфизмов TLR4 человека соответствует миграции из Африки, и вполне вероятно, что полиморфизмы возникли в Африке до миграции на другие континенты. [ 56 ] [ 57 ]
У людей были идентифицированы различные однонуклеотидные полиморфизмы (SNP) TLR4. Для некоторых из них сообщалось о связи с повышенной восприимчивостью к грамотрицательным бактериальным инфекциям или более быстрому прогрессированию и более тяжелому течению сепсиса у пациентов в критическом состоянии. Однако они очень редки, и их частота варьирует в зависимости от этнического происхождения. Двумя преобладающими SNP являются Asp299Gly и Thr399Ile, частота которых составляет <10% у европеоидной популяции и еще ниже у азиатской популяции. [ 58 ] Эти два SNP представляют собой миссенс-мутации, связанные, таким образом, с потерей функции, что может объяснить их негативное влияние на инфекционный контроль. Исследования действительно показали, что SNP TLR4 D299G ограничивает ответ на LPS, ставя под угрозу рекрутирование MyD88 и TRIF в TLR4 и, следовательно, секрецию цитокинов, но не влияя на экспрессию TLR4. [ 59 ] [ 60 ] Структурный анализ человеческого TLR4 с SNP D299G позволяет предположить, что это изменение аминокислоты влияет на взаимодействие Ван-дер-Ваальса и водородные связи в богатых лейцином повторах, модулируя свойства его поверхности, которые могут влиять на связывание лиганда LPS с TLR4. [ 61 ]
Клиническое значение
[ редактировать ]Сообщается, что TLR4 играет роль друга и врага при различных заболеваниях человека, таких как бактериальные инфекции и рак. Эта двойная роль TLR4 зависит от интенсивности, продолжительности и места (поверхностная или эндосома) его активации, его полиморфизма и баланса активации сигнальных путей (MyD88 против TRIF).
Инфекционные заболевания
[ редактировать ]TLR4 играет центральную роль в контроле бактериальных инфекций посредством распознавания молекул ЛПС грамотрицательных и некоторых грамположительных бактерий. [ 62 ] Во время инфекций TLR4 на клетках врожденного иммунитета активируются молекулами ЛПС, присутствующими в тканях и кровотоке. Это активирует врожденный иммунитет, первую линию защиты от вторжения микроорганизмов, и запускает провоспалительные реакции, которые способствуют уничтожению вторгшихся бактерий. [ 13 ] Обычно воспаление играет защитную роль. Это сложный и скоординированный процесс, за которым следуют пути разрешения, которые восстанавливают целостность и функцию тканей. Однако в некоторых случаях чрезмерное и неконтролируемое воспаление, вызванное TLR4 во время инфекции, может привести к сепсису и септическому шоку . [ 33 ] Инфекции грамотрицательными бактериями, такими как Escherichia coli и Pseudomonas aeruginosa, являются преобладающими причинами тяжелого сепсиса у людей. Некоторые исследования связали полиморфизм TLR4 (SNP Asp299Gly и Thr399Ile) с повышенной восприимчивостью к сепсису из-за грамотрицательной инфекции, но другие исследования не удалось это подтвердить. [ 63 ]
Рак
[ редактировать ]Роль TLR4 в контроле прогрессирования рака и в терапии рака хорошо документирована.
Хорошо известно, что стимуляция TLR4 природными производными и ЛПС вызывает сильную противоопухолевую активность. Эта противоопухолевая активность связана со способностью ЛПС стимулировать врожденный иммунитет через TLR4, что приводит к выработке провоспалительных цитокинов и интерферонов 1 типа, а также к непрямой генерации адаптивных противоопухолевых ответов. [ 64 ] [ 65 ]
Первые сведения об эффективности агонистов TLR4, таких как ЛПС, в иммунотерапии рака были получены в XIX веке, когда было обнаружено, что бактериальные инфекции вызывают регрессию опухолей. [ 66 ] Позже доктор Уильям Коли продемонстрировал терапевтическую эффективность смешанной бактериальной вакцины, так называемого «токсина Коли», против рака человека. [ 67 ] С тех пор был сделан ряд разработок в лечении или профилактике рака с использованием смесей бактерий, сильно активирующих TLR4 за счет содержания ЛПС. Противотуберкулезная вакцина Bacillus Calmette-Guerin (BCG) была одобрена Федеральным управлением по лекарственным средствам (FDA) в 1990 году для местного лечения поверхностного рака мочевого пузыря. БЦЖ способствует созреванию дендритных клеток, и этот эффект зависит от TLR4 (а также TLR2). [ 68 ] Имеются также сообщения о лечении плоскоклеточного рака полости рта , желудка, головы и шеи и шейки матки лиофилизированным стрептококковым препаратом ОК-432 (пицибанил). [ 69 ] Механизм действия OK-432 включает активацию TLR4, поскольку OKA-432 не ингибирует рост опухоли при нокауте TLR4, как это происходит на мышах дикого типа. [ 70 ]
Очищенный ЛПС также продемонстрировал высокую противоопухолевую эффективность в качестве системного терапевтического средства на нескольких моделях опухолей. [ 71 ] [ 72 ] В 90-х годах клинические испытания по оценке внутривенного введения ЛПС пациентам с раком дали положительные результаты, включая несколько случаев стабилизации заболевания и частичного ответа. Однако сообщалось об ограничении токсичности при дозах в диапазоне нг/кг, которые слишком низки для достижения значительного противоопухолевого эффекта. [ 73 ]
Впоследствии были произведены и оценены в клинике детоксицированные агонисты TLR4 (производные ЛПС). Сюда входит MPL, химически модифицированный ЛПС, который был первым агонистом TLR4, одобренным и коммерциализированным GSK в составе 5 вакцин для человека (ВПЧ, зостер, гепатит B, малярия, RSV). MPL исследовался в качестве адъюванта для лечебных противоопухолевых вакцин после одобрения мелацина в Канаде для лечения пациентов со злокачественной меланомой. [ 74 ] Также были разработаны синтетические производные ЛПС на основе структур фрагментов дефосфорилированного липида А, которые подтвердили мощную адъювантную и противоопухолевую активность в качестве терапевтических средств. В частности, внутриопухолевое введение глюкопиранозил-липидного адъюванта (GLA-SE/G100), синтетического детоксицированного аналога липида А, сформулированного в стабильной эмульсии, продемонстрировало противоопухолевый иммунный ответ и регресс опухоли у пациентов с карциномой Меркеля. [ 75 ] и сильная адъювантная активность в исследованиях фазы 2 в сочетании с пембролизумабом у пациентов с фолликулярной лимфомой. [ 76 ] [ 77 ]
Помимо признанной противоопухолевой эффективности активации TLR4 с помощью ЛПС, некоторые исследования показывают, что TLR4 может также способствовать развитию некоторых видов рака (рак простаты, печени, молочной железы и легких) и может способствовать резистентности к химиотерапии паклитакселом при раке молочной железы. [ 78 ] Некоторые клинические исследования также показали потенциальную корреляцию между экспрессией TLR4 в опухолевых клетках и прогрессированием опухоли. Однако в многочисленных клинических исследованиях, проведенных с природными ЛПС или производными ЛПС, о таком эффекте не сообщалось. Напротив, в исследованиях фазы 2 с GLA сообщалось о положительной связи между исходной экспрессией TLR4 в опухолях и увеличением общей скорости ответа. [ 77 ]
Потенциальное влияние TLR4 на прогрессирование некоторых видов рака было связано с избыточной выработкой провоспалительных цитокинов посредством активации сигнального пути TLR4-MyD88/NF-kB. [ 79 ] [ 80 ] [ 81 ] Несколько исследований показали, что это опосредовано неправильным использованием передачи сигналов DAMP опухолевыми клетками. [ 12 ] [ 82 ] [ 14 ]
Многие DAMP высвобождаются умирающими или некротическими опухолевыми клетками и присутствуют во время прогрессирования рака. DAMP, высвобождаемые из опухолевых клеток, могут напрямую активировать экспрессируемый опухолью TLR4, что вызывает химиорезистентность, миграцию, инвазию и метастазирование. Кроме того, хроническое воспаление, индуцированное DAMP в микроокружении опухоли, вызывает увеличение популяций иммуносупрессоров, таких как макрофаги М2, супрессорные клетки миелоидного происхождения (MDSC) и регуляторные Т-клетки (Tregs). [ 12 ] Было обнаружено, что DAMP, такие как белки HMGB1, S100 и белки теплового шока (HSP), сильно активируют воспалительные пути и высвобождают IL-1, IL-6, LT-β, IFN-γ, TNF и трансформирующий фактор роста (TGF). )-β способствует воспалению, иммуносупрессии, ангиогенезу и пролиферации опухолевых клеток. [ 11 ]
В нескольких исследованиях оценивалась потенциальная связь этого полиморфизма TLR4 с риском рака, но данные весьма противоречивы. Однако некоторые метаанализы предполагают связь SNP D299G с раком желудка, вирусно-индуцированным и женским раком (шейки матки, яичников). [ 83 ]
Нейрогенеративные заболевания
[ редактировать ]Все больше данных свидетельствует о роли TLR4 в развитии и прогрессировании нейрогенеративных расстройств, таких как болезнь Альцгеймера, болезнь Паркинсона и болезнь Хантингтона. В головном мозге TLR4 экспрессируется нейронами, а также ненейрональными глиальными клетками, включая микроглию, астроциты и олигодендроциты. TLR4 экспрессируется преимущественно микроглией и в меньшей степени астроцитами, олигодендроцитами и нейронами. [ 5 ] Микроглия является представителем системы мононуклеарных фагоцитов в головном мозге, и активация TLR4 регулирует некоторые их функции, такие как фагоцитарная активность. [ 84 ] [ 13 ]
Было высказано предположение, что активация микроглиального TLR4 защищает от нейродегенеративных заболеваний или замедляет их развитие, в частности, за счет усиления клиренса нейротоксических белков, таких как Aβ и его агрегатов, благодаря повышенной фагоцитарной и аутофагической активности. [ 85 ]
Однако считается, что хроническая активация TLR4 связана с глиальной гибелью нейронов из-за чрезмерной секреции провоспалительных цитотоксинов, приводящей к нейровоспалению, ключевому фактору в развитии многих нейродегенеративных заболеваний. [ 86 ] [ 87 ] В головном мозге TLR4 может активироваться различными эндогенными DAMP в дополнение к белкам, связанным с патологией, таким как агрегаты амилоидных пептидов (Aβ) или α-синуклеина. [ 88 ] Все эти структуры связывают TLR4 и активируют нижестоящие сигнальные пути в глии, индуцируя секрецию активных форм кислорода (АФК) и провоспалительных цитокинов, таких как IL-1β и TNF-α, что может привести к повреждению и гибели нейронов. [ 86 ] [ 89 ] [ 90 ] Гибель нейронов сопровождается выбросом DAMP во внеклеточное пространство, которое затем может дополнительно активировать TLR4, усугубляя нейровоспаление. [ 91 ] У пациентов с болезнью Альцгеймера (БА) уровни циркулирующих DAMP, таких как HMGB1 и растворимый RAGE, значительно повышены, что коррелирует с уровнями бета-амилоида. [ 92 ] У пациентов с АД уровень S100B в сыворотке также тесно связан с тяжестью заболевания. [ 93 ] Роль оси HMGB1-TLR4 очень важна в патогенезе болезни Паркинсона (БП). Уровни белков HMGB1 и TLR4 в сыворотке были значительно повышены у пациентов с БП и коррелировали со стадиями БП. [ 94 ]
Нацеливание на TLR4 с помощью агонистов или антагонистов или модуляция его нижестоящих сигнальных путей может иметь терапевтический потенциал при лечении нейродегенеративных заболеваний. [ 95 ] Специфические антагонисты TLR4 могут подавлять нейровоспаление за счет снижения перепроизводства медиаторов воспаления и цитотоксинов глией. Однако антагонисты TLR4 могут оказывать неблагоприятное воздействие на ЦНС, ингибируя фагоцитоз глии, снижая клиренс белка и препятствуя миелинизации. [ 96 ] Некоторые исследования показали, что селективные агонисты TLR4 могут быть полезны, усиливая фагоцитарную активность микроглии, что приводит к усилению очистки поврежденных тканей и аномальных белковых агрегатов, связанных с несколькими различными заболеваниями ЦНС. Было обнаружено, что повторные инъекции MPL в апирогенных дозах значительно улучшают состояние мышей с патологией, связанной с AD. [ 97 ] MPL привел к значительному снижению нагрузки Aβ в головном мозге, а также к усилению когнитивных функций. MPL индуцировал мощный фагоцитарный ответ микроглии, вызывая при этом умеренную воспалительную реакцию. Однако побочные эффекты могут быть вызваны агонистами TLR 4, индуцирующими секрецию медиаторов воспаления. Таким образом, исследования показали, что агонисты TLR4, которые избирательно активируют сигнальный путь TRIF, могут быть очень полезны при лечении нейродегенеративных заболеваний за счет увеличения фагоцитарной активности глиальных клеток без значительного увеличения глиальных цитокинов и цитотоксинов. [ 96 ]
Лекарства, нацеленные на TLR4
[ редактировать ]Было показано, что TLR4 важен для долгосрочных побочных эффектов опиоидных анальгетиков . Были протестированы различные лиганды мю-опиоидных рецепторов , и было обнаружено, что они также обладают действием в качестве агонистов или антагонистов TLR4, при этом опиоидные агонисты, такие как (+)-морфин, являются агонистами TLR4, тогда как опиоидные антагонисты, такие как налоксон, являются антагонистами TLR4. Активация TLR4 приводит к высвобождению модуляторов воспаления, включая TNF-α и интерлейкин-1 , и считается, что постоянное высвобождение этих модуляторов на низком уровне со временем снижает эффективность лечения опиоидными препаратами и участвует как в развитии толерантности, так и в развитии толерантности. к опиоидным анальгетикам, [ 98 ] [ 99 ] а также в возникновении побочных эффектов, таких как гипералгезия и аллодиния , которые могут стать проблемой после длительного использования опиоидных препаратов. [ 100 ] [ 101 ] Было показано, что препараты, блокирующие действие TNF-α или IL-1β, усиливают анальгетический эффект опиоидов и уменьшают развитие толерантности и других побочных эффектов. [ 102 ] [ 103 ] и это также было продемонстрировано на препаратах, которые блокируют сам TLR4.
Было обнаружено, что реакция TLR4 на опиоидные препараты не зависит от энантиомеров , поэтому «неестественные» энантиомеры опиоидных препаратов, таких как морфин и налоксон , которым не хватает сродства к опиоидным рецепторам, по-прежнему производят ту же активность в отношении TLR4, что и их «нормальные» энантиомеры. энантиомеры. [ 104 ] [ 105 ] Это означает, что неприродные энантиомеры опиоидных антагонистов, такие как (+)-налоксон, можно использовать для блокирования активности TLR4 опиоидных анальгетиков, оставляя при этом анальгетическую активность, опосредованную мю-опиоидным рецептором, неизменной. [ 106 ] [ 105 ] [ 107 ] Это также может быть механизмом благоприятного воздействия сверхнизких доз налтрексона на опиоидную аналгезию. [ 108 ]
Морфин вызывает воспаление путем связывания с белком антигена лимфоцитов 96 , который, в свою очередь, заставляет белок связываться с Toll-подобным рецептором 4 (TLR4). [ 109 ] Морфин-индуцированная активация TLR4 ослабляет боли подавление опиоидами и усиливает развитие толерантности к опиоидам и зависимости , злоупотребления наркотиками и других негативных побочных эффектов, таких как угнетение дыхания и гипералгезия. на основе опиоидов Кандидаты в лекарства, нацеленные на TLR4, могут улучшить методы лечения боли . [ 110 ]
Агонисты
[ редактировать ]Помимо ЛПС и его производных, постулируется существование до 30 природных агонистов TLR4 с разнообразной химической структурой. Однако, помимо DAMP, другие не являются прямыми активаторами TLR4 и, следовательно, могут действовать как шапероны для TLR4 или как промоторы интернализации LPS. [ 8 ] [ 111 ] [ 112 ]
Антагонисты
[ редактировать ]По состоянию на 2020 год не было конкретных антагонистов TLR4, одобренных в качестве лекарств. [ 113 ]
- Амитриптилин [ 114 ]
- Циклобензаприн [ 114 ]
- Эриторан [ 115 ]
- Кетотифен [ 114 ]
- Имипрамин [ 114 ]
- Миансерин [ 114 ]
- Из Ибудилы [ 116 ]
- Пиноцембрин [ 117 ]
- Ресаторвид [ 118 ]
- М62812
- Налоксон [ 119 ]
- (+)-Налоксон («неестественный» изомер, не обладающий сродством к опиоидным рецепторам, поэтому избирательно ингибирует TLR4) [ 105 ]
- Налтрексон [ 119 ]
- (+)-Налтрексон [ 119 ]
- ЛПС-РС [ 119 ]
- Пропентофиллин [ нужна ссылка ]
- Пентоксифиллин [ 120 ] (и подавлять экспрессию TLR4 [ 121 ] )
- Тапентадол (смешанный агонист/антагонист)
- TLR4-IN-C34 [ 122 ]
- Пальмитоилэтаноламид [ 123 ]
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с GRCh38: Версия Ensembl 89: ENSG00000136869 – Ensembl , май 2017 г.
- ^ Перейти обратно: а б с GRCm38: выпуск Ensembl 89: ENSMUSG00000039005 – Ensembl , май 2017 г.
- ^ «Ссылка на Human PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Ссылка на Mouse PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ Перейти обратно: а б с Вауре С, Лю Ю (2014). «Сравнительный обзор экспрессии и функциональности толл-подобного рецептора 4 у разных видов животных» . Границы в иммунологии . 5 : 316. дои : 10.3389/fimmu.2014.00316 . ПМК 4090903 . ПМИД 25071777 .
- ^ Манке К., Бехер Э., Риккарди-Кастаньоли П., Люгер Т.А., Шварц Т., Граббе С. (1997). «CD14 экспрессируется подмножествами мышиных дендритных клеток и активируется липополисахаридом». В Риккарди-Кастаньоли П (ред.). Дендритные клетки в фундаментальной и клинической иммунологии . Достижения экспериментальной медицины и биологии. Том. 417. Бостон, Массачусетс: Springer US. стр. 145–159. дои : 10.1007/978-1-4757-9966-8_25 . ISBN 978-1-4757-9968-2 . ПМИД 9286353 .
- ^ Сабро И., Джонс Э.К., Ашер Л.Р., Уайт М.К., Дауэр С.К. (май 2002 г.). «Toll-подобный рецептор (TLR)2 и TLR4 в гранулоцитах периферической крови человека: решающая роль моноцитов в лейкоцитарных липополисахаридных реакциях». Журнал иммунологии . 168 (9): 4701–4710. дои : 10.4049/jimmunol.168.9.4701 . ПМИД 11971020 .
- ^ Перейти обратно: а б с Ян Х., Ван Х., Джу З., Рагаб А.А., Лундбек П., Лонг В. и др. (январь 2015 г.). «MD-2 необходим для дисульфидной HMGB1-зависимой передачи сигналов TLR4» . Журнал экспериментальной медицины . 212 (1): 5–14. дои : 10.1084/jem.20141318 . ПМЦ 4291531 . ПМИД 25559892 .
- ^ Перейти обратно: а б Цзян Д., Лян Дж., Фань Дж., Ю С., Чен С., Луо Ю. и др. (ноябрь 2005 г.). «Регуляция повреждения и восстановления легких с помощью Toll-подобных рецепторов и гиалуронана». Природная медицина . 11 (11): 1173–1179. дои : 10.1038/nm1315 . ПМИД 16244651 . S2CID 11765495 .
- ^ Перейти обратно: а б Фан Х, Анг Б, Сюй Х, Хуан Х, Ву Ю, Сунь Ю и др. (март 2014 г.). «TLR4 необходим для активации дендритных клеток и усиления противоопухолевого ответа Т-клеток за счет DAMP, высвобождаемых из раковых клеток, подвергшихся химическому стрессу» . Клеточная и молекулярная иммунология . 11 (2): 150–159. дои : 10.1038/cmi.2013.59 . ПМЦ 4003380 . ПМИД 24362470 .
- ^ Перейти обратно: а б с Эрнандес С., Хюбенер П., Швабе РФ (ноябрь 2016 г.). «Молекулярные закономерности, связанные с повреждением при раке: палка о двух концах» . Онкоген . 35 (46): 5931–5941. дои : 10.1038/onc.2016.104 . ПМК 5119456 . ПМИД 27086930 .
- ^ Перейти обратно: а б с д и Чан Джи, Ли Дж.В., Ким Ю.С., Ли С.Э., Хан Х.Д., Хонг К.Дж. и др. (декабрь 2020 г.). «Взаимодействие между белками опухолевого происхождения и Toll-подобными рецепторами» . Экспериментальная и молекулярная медицина . 52 (12): 1926–1935. дои : 10.1038/s12276-020-00540-4 . ПМК 8080774 . ПМИД 33299138 .
- ^ Перейти обратно: а б с д Молтени М., Джемма С., Россетти С. (2016). «Роль Toll-подобного рецептора 4 в инфекционном и неинфекционном воспалении» . Медиаторы воспаления . 2016 : 6978936. doi : 10.1155/2016/6978936 . ПМЦ 4887650 . ПМИД 27293318 .
- ^ Перейти обратно: а б Ро Дж.С., Зон Д.Х. (август 2018 г.). «Молекулярные закономерности, связанные с повреждением при воспалительных заболеваниях» . Иммунная сеть . 18 (4): е27. дои : 10.4110/in.2018.18.e27 . ПМК 6117512 . ПМИД 30181915 .
- ^ Цукамото Х., Такеучи С., Кубота К., Кобаяши Ю., Козакай С., Укай И. и др. (июнь 2018 г.). «Липополисахарид (ЛПС)-связывающий белок стимулирует CD14-зависимую интернализацию Toll-подобного рецептора 4 и индуцированную ЛПС активацию оси TBK1-IKKϵ-IRF3» . Журнал биологической химии . 293 (26): 10186–10201. дои : 10.1074/jbc.M117.796631 . ПМК 6028956 . ПМИД 29760187 .
- ^ Перейти обратно: а б Пак Б.С., Сон Д.Х., Ким Х.М., Чхве Б.С., Ли Х., Ли Джо (апрель 2009 г.). «Структурные основы распознавания липополисахаридов комплексом TLR4-MD-2». Природа . 458 (7242): 1191–1195. Бибкод : 2009Natur.458.1191P . дои : 10.1038/nature07830 . ПМИД 19252480 . S2CID 4396446 .
- ^ Пак Б.С., Ли Джо (декабрь 2013 г.). «Распознавание липополисахаридного рисунка комплексами TLR4» . Экспериментальная и молекулярная медицина . 45 (12): е66. дои : 10.1038/эмм.2013.97 . ПМЦ 3880462 . ПМИД 24310172 .
- ^ Стеймле А., Аутенрит И.Б., Фрик Дж.С. (август 2016 г.). «Структура и функции: модификации липида А у комменсалов и патогенов» . Международный журнал медицинской микробиологии . 306 (5): 290–301. дои : 10.1016/j.ijmm.2016.03.001 . ПМИД 27009633 .
- ^ Келли С.Л., Лукк Т., Наир С.К., Таппинг Р.И. (февраль 2013 г.). «Кристаллическая структура человеческого растворимого CD14 представляет собой изогнутый соленоид с гидрофобным аминоконцевым карманом» . Журнал иммунологии . 190 (3): 1304–1311. doi : 10.4049/jimmunol.1202446 . ПМЦ 3552104 . ПМИД 23264655 .
- ^ Мурои М., Танамото К. (ноябрь 2002 г.). «Полисахаридная часть играет незаменимую роль в индуцированной липополисахаридом сальмонеллы активации NF-каппаВ через человеческий толл-подобный рецептор 4» . Инфекция и иммунитет . 70 (11): 6043–6047. дои : 10.1128/IAI.70.11.6043-6047.2002 . ПМК 130318 . ПМИД 12379680 .
- ^ Кавайон Дж. М., Фиттинг С., Карофф М., Хаффнер-Кавайон Н. (март 1989 г.). «Диссоциация клеточно-ассоциированного интерлейкина-1 (IL-1) и высвобождение IL-1, индуцированное липополисахаридом и липидом А» . Инфекция и иммунитет . 57 (3): 791–797. дои : 10.1128/iai.57.3.791-797.1989 . ПМЦ 313178 . ПМИД 2537258 .
- ^ Перейти обратно: а б с д Шен Х., Тесар Б.М., Уокер В.Е., Гольдштейн Д.Р. (август 2008 г.). «Двойная передача сигналов MyD88 и TRIF имеет решающее значение для максимального TLR4-индуцированного созревания дендритных клеток» . Журнал иммунологии . 181 (3): 1849–1858. doi : 10.4049/jimmunol.181.3.1849 . ПМК 2507878 . ПМИД 18641322 .
- ^ Накаяма М., Ники Ю., Кавасаки Т., Такеда Ю., Икегами Х., Тояма Ю. и др. (октябрь 2013 г.). «Ось IL-32-PAR2 представляет собой врожденный иммунный сенсор, обеспечивающий альтернативную передачу сигналов оси LPS-TRIF» . Научные отчеты 3 (1): 2960. Бибкод : 2013NatSR...3E2960N . дои : 10.1038/srep02960 . ПМЦ 3797434 . ПМИД 24129891 .
- ^ Придмор AC, Джарвис Г.А., Джон CM, Джек Д.Л., Дауэр С.К., Рид RC (июль 2003 г.). «Активация toll-подобного рецептора 2 (TLR2) и TLR4/MD2 Neisseria не зависит от сиалирования капсулы и липоолигосахаридов (LOS), но широко варьируется между LOS из разных штаммов» . Инфекция и иммунитет . 71 (7): 3901–3908. дои : 10.1128/IAI.71.7.3901-3908.2003 . ПМК 161978 . ПМИД 12819075 .
- ^ Стивенсон Х.Н., Джон К.М., Наз Н., Гундогду О., Доррелл Н., Рен Б.В. и др. (июль 2013 г.). «Сиалирование липоолигосахаридов Campylobacter jejuni, фосфорилирование и модификации амидно-эфирных связей позволяют точно настроить активацию Toll-подобного рецептора 4 человека» . Журнал биологической химии . 288 (27): 19661–19672. дои : 10.1074/jbc.M113.468298 . ПМЦ 3707672 . ПМИД 23629657 .
- ^ Александр-Флойд Дж., Басс А.Р., Харбертс Э.М., Грубау Д., Буксбаум Дж.Д., Бродский И.Е. и др. (август 2022 г.). Боймлер А.Дж. (ред.). «Варианты липида А по-разному активируют TLR4 человека и неканоническую инфламмасому и требуют основного олигосахарида для активации воспалительной сомы» . Инфекция и иммунитет . 90 (8): e0020822. дои : 10.1128/iai.00208-22 . ПМЦ 9387229 . ПМИД 35862709 .
- ^ Бономм Д., Сантеккья И., Вернель-Пойяк Ф., Карофф М., Жермон П., Мюррей Г. и др. (август 2020 г.). «Лептоспиральный ЛПС избегает интернализации мышиного TLR4 и TRIF-ассоциированных антимикробных реакций через О-антиген и связанные с ним липопротеины» . ПЛОС Патогены . 16 (8): e1008639. дои : 10.1371/journal.ppat.1008639 . ПМЦ 7447051 . ПМИД 32790743 .
- ^ Перейти обратно: а б Полссон-Макдермотт Э.М., О'Нил Л.А. (октябрь 2004 г.). «Передача сигнала липополисахаридным рецептором, Toll-подобным рецептором-4» . Иммунология . 113 (2): 153–162. дои : 10.1111/j.1365-2567.2004.01976.x . ПМК 1782563 . ПМИД 15379975 .
- ^ Лу Ю.К., Йе У.К., Охаши П.С. (май 2008 г.). «Путь передачи сигнала LPS/TLR4». Цитокин . 42 (2): 145–151. дои : 10.1016/j.cyto.2008.01.006 . ПМИД 18304834 .
- ^ Перейти обратно: а б Мейснер Ф., Шелтема Р.А., Молленкопф Х.Дж., Манн М. (апрель 2013 г.). «Прямая протеомная количественная оценка секретома активированных иммунных клеток». Наука . 340 (6131): 475–478. Бибкод : 2013Sci...340..475M . дои : 10.1126/science.1232578 . ПМИД 23620052 . S2CID 40513139 .
- ^ Перейти обратно: а б Каваи Т., Такеучи О., Фудзита Т., Иноуэ Дж., Мюльрадт П.Ф., Сато С. и др. (ноябрь 2001 г.). «Липополисахарид стимулирует MyD88-независимый путь и приводит к активации IFN-регуляторного фактора 3 и экспрессии подмножества генов, индуцируемых липополисахаридами». Журнал иммунологии . 167 (10): 5887–5894. дои : 10.4049/jimmunol.167.10.5887 . ПМИД 11698465 .
- ^ Перейти обратно: а б Шанте Х., Гиссе АС, Пилетт С, Сибилле Ю (октябрь 2007 г.). «ЛПС индуцирует выработку IL-10 альвеолярными макрофагами человека посредством MAP-киназ- и Sp1-зависимых механизмов» . Респираторные исследования . 8 (1): 71. дои : 10.1186/1465-9921-8-71 . ПМК 2080632 . ПМИД 17916230 .
- ^ Перейти обратно: а б Цесельска А., Матыйек М., Квятковска К. (февраль 2021 г.). «Торговля TLR4 и CD14 и его влияние на провоспалительную передачу сигналов, индуцированную ЛПС» . Клеточные и молекулярные науки о жизни . 78 (4): 1233–1261. дои : 10.1007/s00018-020-03656-y . ПМЦ 7904555 . ПМИД 33057840 .
- ^ О'Нил Л.А., Голенбок Д., Боуи А.Г. (июнь 2013 г.). «История Toll-подобных рецепторов - новое определение врожденного иммунитета». Обзоры природы. Иммунология . 13 (6): 453–460. дои : 10.1038/nri3446 . hdl : 2262/72552 . ПМИД 23681101 . S2CID 205491986 .
- ^ Ватанабэ С., Кумазава Ю., Иноуэ Дж. (2013). «Липосомальный липополисахарид инициирует TRIF-зависимый сигнальный путь независимо от CD14» . ПЛОС ОДИН . 8 (4): e60078. Бибкод : 2013PLoSO...860078W . дои : 10.1371/journal.pone.0060078 . ПМЦ 3615118 . ПМИД 23565187 .
- ^ Лиен Э., Минс ТК, Хейне Х., Ёсимура А., Кусумото С., Фукасе К. и др. (февраль 2000 г.). «Toll-подобный рецептор 4 обеспечивает лиганд-специфическое распознавание бактериального липополисахарида» . Журнал клинических исследований . 105 (4): 497–504. дои : 10.1172/JCI8541 . ПМК 289161 . ПМИД 10683379 .
- ^ Перейти обратно: а б Шетаб Бушехри М.А., Лампрехт А. (ноябрь 2018 г.). «Иммунотерапия рака на основе TLR4: обзор достижений и недостатков». Молекулярная фармацевтика . 15 (11): 4777–4800. doi : 10.1021/acs.molpharmaceut.8b00691 . ПМИД 30226786 . S2CID 52297047 .
- ^ Кайшо Т., Такеучи О., Каваи Т., Хосино К., Акира С. (май 2001 г.). «Индуцированное эндотоксином созревание дендритных клеток с дефицитом MyD88». Журнал иммунологии . 166 (9): 5688–5694. дои : 10.4049/jimmunol.166.9.5688 . ПМИД 11313410 .
- ^ Хоэбе К., Янссен Э.М., Ким СО, Алексопулу Л., Флавелл Р.А., Хан Дж. и др. (декабрь 2003 г.). «Повышающая регуляция костимулирующих молекул, индуцированная липополисахаридом и двухцепочечной РНК, происходит по триф-зависимым и триф-независимым путям». Природная иммунология . 4 (12): 1223–1229. дои : 10.1038/ni1010 . ПМИД 14625548 . S2CID 8505015 .
- ^ Шен Х., Тесар Б.М., Уокер В.Е., Гольдштейн Д.Р. (август 2008 г.). «Двойная передача сигналов MyD88 и TRIF имеет решающее значение для максимального TLR4-индуцированного созревания дендритных клеток» . Журнал иммунологии . 181 (3): 1849–1858. doi : 10.4049/jimmunol.181.3.1849 . ПМК 2507878 . ПМИД 18641322 .
- ^ Тромбетта Э.С., Эберсолд М., Гарретт В., Пайпарт М., Меллман I (февраль 2003 г.). «Активация лизосомальной функции во время созревания дендритных клеток». Наука . 299 (5611): 1400–1403. дои : 10.1126/science.1080106 . ПМИД 12610307 . S2CID 46594244 .
- ^ Терли С.Дж., Инаба К., Гарретт В.С., Эберсолд М., Унтернарер Дж., Штайнман Р.М. и др. (апрель 2000 г.). «Транспорт комплексов пептид-MHC класса II в развивающихся дендритных клетках». Наука . 288 (5465): 522–527. дои : 10.1126/science.288.5465.522 . ПМИД 10775112 .
- ^ Кайшо Т., Такеучи О., Каваи Т., Хосино К., Акира С. (май 2001 г.). «Индуцированное эндотоксином созревание дендритных клеток с дефицитом MyD88». Журнал иммунологии . 166 (9): 5688–5694. дои : 10.4049/jimmunol.166.9.5688 . ПМИД 11313410 .
- ^ Хоэбе К., Янссен Э.М., Ким СО, Алексопулу Л., Флавелл Р.А., Хан Дж. и др. (декабрь 2003 г.). «Повышающая регуляция костимулирующих молекул, индуцированная липополисахаридом и двухцепочечной РНК, происходит по триф-зависимым и триф-независимым путям». Природная иммунология . 4 (12): 1223–1229. дои : 10.1038/ni1010 . ПМИД 14625548 . S2CID 8505015 .
- ^ Тромбетта Э.С., Эберсолд М., Гарретт В., Пайпарт М., Меллман I (февраль 2003 г.). «Активация лизосомальной функции во время созревания дендритных клеток». Наука . 299 (5611): 1400–1403. дои : 10.1126/science.1080106 . ПМИД 12610307 . S2CID 46594244 .
- ^ Терли С.Дж., Инаба К., Гарретт В.С., Эберсолд М., Унтернарер Дж., Штайнман Р.М. и др. (апрель 2000 г.). «Транспорт комплексов пептид-MHC класса II в развивающихся дендритных клетках». Наука . 288 (5465): 522–527. дои : 10.1126/science.288.5465.522 . ПМИД 10775112 .
- ^ Перейти обратно: а б Наир-Гупта П., Баккарини А., Тунг Н., Сейффер Ф., Флори О., Хуанг Ю. и др. (июль 2014 г.). «Сигналы TLR индуцируют фагосомальную доставку MHC-I из эндосомального отсека рециркуляции, чтобы обеспечить перекрестную презентацию» . Клетка . 158 (3): 506–521. дои : 10.1016/j.cell.2014.04.054 . ПМК 4212008 . ПМИД 25083866 .
- ^ Хан Дж.Э., Вуй С.Р., Ким К.С., Чо Ю.Дж., Чо В.Дж., Ли Н.Г. (22 января 2014 г.). Шин EC (ред.). «Характеристика структуры и иммуностимулирующей активности вакцинного адъюванта де-О-ацилированного липоолигосахарида» . ПЛОС ОДИН . 9 (1): e85838. Бибкод : 2014PLoSO...985838H . дои : 10.1371/journal.pone.0085838 . ПМК 3899070 . ПМИД 24465739 .
- ^ Шариф О., Большаков В.Н., Рейнс С., Ньюхэм П., Перкинс Н.Д. (январь 2007 г.). «Транскрипционное профилирование LPS-индуцированного ответа NF-kappaB в макрофагах» . БМК Иммунология . 8 (1): 1. дои : 10.1186/1471-2172-8-1 . ПМЦ 1781469 . ПМИД 17222336 .
- ^ Паавонен Дж., Дженкинс Д., Бош Ф.К., Науд П., Салмерон Дж., Уилер К.М. и др. (июнь 2007 г.). «Эффективность профилактической адъювантной бивалентной вирусоподобной вакцины L1 против заражения вирусом папилломы человека типов 16 и 18 у молодых женщин: промежуточный анализ двойного слепого рандомизированного контролируемого исследования III фазы». Ланцет . 369 (9580): 2161–2170. дои : 10.1016/S0140-6736(07)60946-5 . ПМИД 17602732 . S2CID 26318328 .
- ^ Кунди М. (апрель 2007 г.). «Новая вакцина против гепатита В, созданная с использованием улучшенной адъювантной системы». Экспертная оценка вакцин . 6 (2): 133–140. дои : 10.1586/14760584.6.2.133 . ПМИД 17408363 . S2CID 35472093 .
- ^ Гарсон Н., Ди Паскуале А (январь 2017 г.). «От открытия до лицензирования: история адъювантной системы» . Человеческие вакцины и иммунотерапия . 13 (1): 19–33. дои : 10.1080/21645515.2016.1225635 . ПМК 5287309 . ПМИД 27636098 .
- ^ Бейтлер Б., Рели М. (2002). «Эволюция TIR, дорожных сборов и TLRS: функциональные выводы из вычислительной биологии». Члены семейства Toll-подобных рецепторов и их лиганды . Актуальные темы микробиологии и иммунологии. Том. 270. стр. 1–21. дои : 10.1007/978-3-642-59430-4_1 . ISBN 978-3-642-63975-3 . ПМИД 12467241 .
- ^ Смирнова И., Полторак А., Чан Е.К., Макбрайд С., Бейтлер Б. (2000). «Филогенетическая изменчивость и полиморфизм локуса toll-подобного рецептора 4 (TLR4)» . Геномная биология . 1 (1): ИССЛЕДОВАНИЕ002. doi : 10.1186/gb-2000-1-1-research002 . ПМК 31919 . ПМИД 11104518 .
- ^ Куах Х., Уилсон Д., Лаваль Г., Патин Е., Манри Дж., Гиберт Дж. и др. (декабрь 2013 г.). «Различное селективное давление формирует эволюцию Toll-подобных рецепторов в популяциях человека и африканских человекообразных обезьян» . Молекулярная генетика человека . 22 (23): 4829–4840. дои : 10.1093/hmg/ddt335 . ПМК 3820138 . ПМИД 23851028 .
- ^ Перейти обратно: а б Баррейро Л.Б., Бен-Али М., Куах Х., Лаваль Г., Патин Э., Пикрелл Дж.К. и др. (июль 2009 г.). «Эволюционная динамика Toll-подобных рецепторов человека и их различный вклад в защиту хозяина» . ПЛОС Генетика . 5 (7): e1000562. дои : 10.1371/journal.pgen.1000562 . ПМК 2702086 . ПМИД 19609346 .
- ^ Плантинга Т.С., Иоана М., Алонсо С., Исагирре Н., Эрвелла М., Йостен Л.А. и др. (2012). «Эволюционная история полиморфизмов TLR4 в Европе» . Журнал врожденного иммунитета . 4 (2): 168–175. дои : 10.1159/000329492 . ПМК 6741577 . ПМИД 21968286 .
- ^ Норин М., Шах М.А., Молл С.М., Чоудхари С., Хусейн Т., Ахмед И. и др. (март 2012 г.). «Полиморфизмы TLR4 и восприимчивость к заболеваниям». Исследование воспаления . 61 (3): 177–188. дои : 10.1007/s00011-011-0427-1 . ПМИД 22277994 . S2CID 9500302 .
- ^ Лонг Х., О'Коннор Б.П., Земанс Р.Л., Чжоу X, Ян IV, Шварц Д.А. (2 апреля 2014 г.). «Полиморфизм Asp299Gly Toll-подобного рецептора 4, но не Thr399Ile, влияет на передачу сигналов и функцию TLR4» . ПЛОС ОДИН . 9 (4): е93550. Бибкод : 2014PLoSO...993550L . дои : 10.1371/journal.pone.0093550 . ПМЦ 3973565 . ПМИД 24695807 .
- ^ Фигероа Л., Сюн Ю., Сонг С., Пиао В., Фогель С.Н., Медведев А.Е. (май 2012 г.). «Полиморфизм Asp299Gly изменяет передачу сигналов TLR4, препятствуя рекрутированию MyD88 и TRIF» . Журнал иммунологии . 188 (9): 4506–4515. doi : 10.4049/jimmunol.1200202 . ПМК 3531971 . ПМИД 22474023 .
- ^ Ото У, Ямакава Н, Акаси-Такамура С, Мияке К, Симидзу Т (ноябрь 2012 г.). «Структурный анализ полиморфизмов D299G и T399I человеческого Toll-подобного рецептора 4» . Журнал биологической химии . 287 (48): 40611–40617. дои : 10.1074/jbc.M112.404608 . ПМЦ 3504774 . ПМИД 23055527 .
- ^ Акира С., Такеда К. (июль 2004 г.). «Передача сигналов Toll-подобного рецептора». Обзоры природы. Иммунология . 4 (7): 499–511. дои : 10.1038/nri1391 . ПМИД 15229469 .
- ^ Нетеа М.Г., Вейменга К., О'Нил Л.А. (май 2012 г.). «Генетическая изменчивость Toll-подобных рецепторов и восприимчивость к болезням». Природная иммунология . 13 (6): 535–542. дои : 10.1038/ni.2284 . ПМИД 22610250 . S2CID 24438756 .
- ^ Четтаб К., Фицсиммонс С., Новиков А., Денис М., Фелип С., Мате Д. и др. (2023). «Системно вводимый детоксицированный агонист TLR4 демонстрирует мощную противоопухолевую активность и приемлемый профиль переносимости в доклинических моделях» . Границы в иммунологии . 14 : 1066402. дои : 10.3389/fimmu.2023.1066402 . ПМК 10200957 . ПМИД 37223101 .
- ^ Ричерт И., Бершар П., Аббес Л., Новиков А., Четтаб К., Вандермотен А. и др. (сентябрь 2023 г.). «Агонист TLR4 вызывает регрессию остеосаркомы, индуцируя противоопухолевый иммунный ответ и перепрограммируя макрофаги M2 в макрофаги M1» . Раки . 15 (18): 4635. doi : 10.3390/cancers15184635 . ПМЦ 10526955 . ПМИД 37760603 .
- ^ Маруяма К., Селмани З., Исии Х., Ямагути К. (март 2011 г.). «Врожденный иммунитет и терапия рака». Международная иммунофармакология . 11 (3): 350–357. дои : 10.1016/j.intimp.2010.09.012 . ПМИД 20955832 .
- ^ Старнес СО (май 1992 г.). «Токсины Коли в перспективе». Природа . 357 (6373): 11–12. Бибкод : 1992Natur.357...11S . дои : 10.1038/357011a0 . ПМИД 1574121 . S2CID 4265230 .
- ^ Цудзи С., Мацумото М., Такеучи О., Акира С., Адзума И., Хаяши А. и др. (декабрь 2000 г.). Кауфманн С.Х. (ред.). «Созревание дендритных клеток человека скелетом клеточной стенки Mycobacterium bovis bacillus Calmette-Guérin: участие толл-подобных рецепторов» . Инфекция и иммунитет . 68 (12): 6883–6890. дои : 10.1128/IAI.68.12.6883-6890.2000 . ПМК 97794 . ПМИД 11083809 .
- ^ Риома Ю, Мория Ю, Окамото М, Канайя И, Сайто М, Сато М (01 сентября 2004 г.). «Биологический эффект ОК-432 (пицибанила) и возможное применение в терапии дендритных клеток» . Противораковые исследования . 24 (5С): 3295–3301. ПМИД 15515424 .
- ^ Окамото М., Осикава Т., Тано Т., Охе Г., Фуруичи С., Нисикава Х. и др. (февраль 2003 г.). «Участие передачи сигналов Toll-подобного рецептора 4 в продукцию гамма-интерферона и противоопухолевый эффект стрептококкового агента ОК-432». Журнал Национального института рака . 95 (4): 316–326. дои : 10.1093/jnci/95.4.316 . ПМИД 12591988 .
- ^ Шир М.Б., Перро М. (апрель 1944 г.). «Химическое лечение опухолей. IX. Реакция мышей с первичными подкожными опухолями на инъекцию вызывающего кровотечение бактериального полисахарида1». JNCI: Журнал Национального института рака . 4 (5): 461–476. дои : 10.1093/jnci/4.5.461 .
- ^ Берендт М.Дж., Норт Р.Дж., Кирштейн Д.П. (декабрь 1978 г.). «Иммунологические основы эндотоксин-индуцированной регрессии опухоли. Необходимость наличия предсуществующего состояния сопутствующего противоопухолевого иммунитета» . Журнал экспериментальной медицины . 148 (6): 1560–1569. дои : 10.1084/jem.148.6.1560 . ПМК 2185097 . ПМИД 309922 .
- ^ Энгельхардт Р., Макенсен А., Галанос С. (май 1991 г.). «Фаза I исследования внутривенного введения эндотоксина (Salmonella abortus equi) у онкологических больных» . Исследования рака . 51 (10): 2524–2530. ПМИД 2021932 .
- ^ «Мелацин - обзор | Темы ScienceDirect» . www.sciencedirect.com . Проверено 20 февраля 2024 г.
- ^ Бхатия С., Миллер Н.Дж., Лу Х., Лонгино Н.В., Ибрани Д., Шинохара М.М. и др. (февраль 2019 г.). «Внутриопухолевый G100, агонист TLR4, индуцирует противоопухолевые иммунные ответы и регрессию опухоли у пациентов с клеточной карциномой Меркеля» . Клинические исследования рака . 25 (4): 1185–1195. дои : 10.1158/1078-0432.CCR-18-0469 . ПМК 6368904 . ПМИД 30093453 .
- ^ Халвани А.С., Панизо С., Исуфи И., Эррера А.Ф., Окада С.И., Кулл Э.Х. и др. (апрель 2022 г.). «Исследование фазы 1/2 внутриопухолевого G100 (агониста TLR4) с пембролизумабом или без него при фолликулярной лимфоме». Лейкемия и лимфома . 63 (4): 821–833. дои : 10.1080/10428194.2021.2010057 . ПМИД 34865586 . S2CID 244943266 .
- ^ Перейти обратно: а б Флауэрс С, Панизо С, Исуфи I, Эррера А.Ф., Окада С, Кулл Э.Х. и др. (08.12.2017). «Внутриопухолевый G100 индуцирует системный иммунитет и регрессию абскопальной опухоли у пациентов с фолликулярной лимфомой: результаты исследования фазы 1/2, изучающего G100 отдельно и в сочетании с пембролизумабом» . Кровь . 130 : 2771. doi : 10.1182/blood.V130.Suppl_1.2771.2771 (неактивен 20 февраля 2024 г.). ISSN 0006-4971 .
{{cite journal}}: CS1 maint: DOI неактивен по состоянию на февраль 2024 г. ( ссылка ) - ^ Раджпут С., Волк-Дрейпер Л.Д., Ран С. (август 2013 г.). «TLR4 является новым фактором, определяющим реакцию на паклитаксел при раке молочной железы» . Молекулярная терапия рака . 12 (8): 1676–1687. дои : 10.1158/1535-7163.MCT-12-1019 . ПМЦ 3742631 . ПМИД 23720768 .
- ^ Чжан Р., Чжао Дж., Сюй Дж., Цзяо Д.С., Ван Дж., Гун ZQ и др. (октябрь 2017 г.). «Андрографолид подавляет пролиферацию клеток SW620 рака толстой кишки человека посредством сигнального пути TLR4/NF-κB/MMP-9» . Письма об онкологии . 14 (4): 4305–4310. дои : 10.3892/ol.2017.6669 . ПМК 5604146 . ПМИД 28943944 .
- ^ Ван Ч., Ван П.Дж., Се Ю.К., Ло С., Ли Ю.К., Чен Ю.К. и др. (февраль 2018 г.). «Резистин способствует прогрессированию рака молочной железы посредством TLR4-опосредованной индукции мезенхимальных фенотипов и свойств стволовости». Онкоген . 37 (5): 589–600. дои : 10.1038/onc.2017.357 . ПМИД 28991224 . S2CID 24926622 .
- ^ Келли М.Г., Альверо А.Б., Чен Р., Силаси Д.А., Абрахамс В.М., Чан С. и др. (апрель 2006 г.). «Передача сигналов TLR-4 способствует росту опухоли и хеморезистентности к паклитакселу при раке яичников». Исследования рака . 66 (7): 3859–3868. дои : 10.1158/0008-5472.CAN-05-3948 . ПМИД 16585214 .
- ^ Хадемальхоссейни М, депутат Арабабади (май 2019 г.). «Toll-подобный рецептор 4 и рак молочной железы: обновленный систематический обзор». Рак молочной железы . 26 (3): 265–271. дои : 10.1007/s12282-018-00935-2 . ПМИД 30543015 . S2CID 56143069 .
- ^ Чжу Л, Юань Х, Цзян Т, Ван Р, Ма Х, Чжан С (20 декабря 2013 г.). «Связь полиморфизмов TLR2 и TLR4 с риском развития рака: метаанализ» . ПЛОС ОДИН . 8 (12): е82858. Бибкод : 2013PLoSO...882858Z . дои : 10.1371/journal.pone.0082858 . ПМЦ 3869723 . ПМИД 24376595 .
- ^ Уордилл Х.Р., Ван Себилле Ю.З., Мандер К.А., Гибсон Р.Дж., Логан Р.М., Боуэн Дж.М. и др. (февраль 2015 г.). «Передача сигналов Toll-подобного рецептора 4: общий биологический механизм токсичности, связанной с режимом лечения: новая гипотеза нейропатии и желудочно-кишечной токсичности». Обзоры лечения рака . 41 (2): 122–128. дои : 10.1016/j.ctrv.2014.11.005 . ПМИД 25512119 .
- ^ Тахара К., Ким Х.Д., Джин Джей-Джей, Максвелл Дж.А., Ли Л., Фукучи К. (ноябрь 2006 г.). «Роль передачи сигналов толл-подобных рецепторов в поглощении и выведении бета-беты» . Мозг . 129 (Часть 11): 3006–3019. дои : 10.1093/brain/awl249 . ПМЦ 2445613 . ПМИД 16984903 .
- ^ Перейти обратно: а б Бьюкенен М.М., Хатчинсон М., Уоткинс Л.Р., Инь Х. (июль 2010 г.). «Толл-подобный рецептор 4 при патологиях ЦНС» . Журнал нейрохимии . 114 (1): 13–27. дои : 10.1111/j.1471-4159.2010.06736.x . ПМК 2909662 . ПМИД 20402965 .
- ^ Цинь Ю, Лю Ю, Хао В, Декер Ю, Томич И, Менгер, доктор медицинских наук и др. (октябрь 2016 г.). «Стимуляция TLR4 ослабляет симптомы и патологии, связанные с болезнью Альцгеймера, у тау-трансгенных мышей». Журнал иммунологии . 197 (8): 3281–3292. doi : 10.4049/jimmunol.1600873 . ПМИД 27605009 .
- ^ Гамбузза М., Ликата Н., Палелла Е., Чели Д., Фоти Куццола В., Итальяно Д. и др. (октябрь 2011 г.). «Нацеливание на Toll-подобные рецепторы: новые методы лечения рассеянного склероза». Журнал нейроиммунологии . 239 (1–2): 1–12. дои : 10.1016/j.jneuroim.2011.08.010 . ПМИД 21889214 . S2CID 3277551 .
- ^ Ранникко Э.Х., Вебер С.С., Кале П.Дж. (сентябрь 2015 г.). «Экзогенный α-синуклеин индуцирует воспалительные реакции, зависимые от толл-подобного рецептора 4, в астроцитах» . BMC Нейронаука . 16:57 . дои : 10.1186/s12868-015-0192-0 . ПМК 4562100 . ПМИД 26346361 .
- ^ Уолтер С., Летембре М., Лю Ю., Хейне Х., Пенке Б., Хао В. и др. (2007). «Роль толл-подобного рецептора 4 в нейровоспалении при болезни Альцгеймера». Клеточная физиология и биохимия . 20 (6): 947–956. дои : 10.1159/000110455 . ПМИД 17982277 . S2CID 6752610 .
- ^ Земельная РГ (февраль 2015 г.). «Роль молекулярных структур, связанных с повреждением, в заболеваниях человека: Часть I - Стимулирование воспаления и иммунитета» . Медицинский журнал Университета Султана Кабуса . 15 (1): e9–e21. ПМК 4318613 . ПМИД 25685392 .
- ^ Фестофф Б.В., Саджа Р.К., ван Дреден П., Кукулло Л. (август 2016 г.). «HMGB1 и тромбин опосредуют дисфункцию гематоэнцефалического барьера, выступая в качестве биомаркеров нейровоспаления и прогрессирования нейродегенерации при болезни Альцгеймера» . Журнал нейровоспаления . 13 (1): 194. дои : 10.1186/s12974-016-0670-z . ПМЦ 4995775 . ПМИД 27553758 .
- ^ Чавес М.Л., Камоццато А.Л., Феррейра Э.Д., Пьязенски И., Кочханн Р., Далл'Игна О. и др. (январь 2010 г.). «Уровни белков S100B и NSE в сыворотке крови у пациентов с болезнью Альцгеймера» . Журнал нейровоспаления . 7 :6. дои : 10.1186/1742-2094-7-6 . ПМЦ 2832635 . ПМИД 20105309 .
- ^ Ян Ю, Хань С., Го Л, Гуань Ц (апрель 2018 г.). «Высокая экспрессия оси HMGB1-TLR4 и ее нижестоящих сигнальных факторов у пациентов с болезнью Паркинсона и взаимосвязь патологической стадии» . Мозг и поведение . 8 (4): e00948. дои : 10.1002/brb3.948 . ПМЦ 5893335 . ПМИД 29670828 .
- ^ Ву Л, Сянь Х, Сюй Г, Тан З, Донг Ф, Чжан М и др. (21 августа 2022 г.). «Толл-подобный рецептор 4: многообещающая терапевтическая мишень для лечения болезни Альцгеймера» . Медиаторы воспаления . 2022 : 7924199. дои : 10.1155/2022/7924199 . ПМЦ 9420645 . ПМИД 36046763 .
- ^ Перейти обратно: а б Лейтнер Г.Р., Венцель Т.Дж., Маршалл Н., Гейтс Э.Дж., Клегерис А. (октябрь 2019 г.). «Нацеливание на толл-подобный рецептор 4 для модуляции нейровоспаления при заболеваниях центральной нервной системы». Мнение экспертов о терапевтических целях . 23 (10): 865–882. дои : 10.1080/14728222.2019.1676416 . ПМИД 31580163 . S2CID 203652175 .
- ^ Мишо Ж.П., Халле М., Лампрон А., Терио П., Префонтен П., Филали М. и др. (январь 2013 г.). «Стимуляция Toll-подобного рецептора 4 детоксицированным лигандом монофосфориллипидом А улучшает патологию, связанную с болезнью Альцгеймера» . Труды Национальной академии наук Соединенных Штатов Америки . 110 (5): 1941–1946. Бибкод : 2013ПНАС..110.1941М . дои : 10.1073/pnas.1215165110 . ПМЦ 3562771 . ПМИД 23322736 .
- ^ Шавит Ю., Вольф Г., Гошен И., Лившиц Д., Йирмия Р. (май 2005 г.). «Интерлейкин-1 противодействует морфиновой анальгезии и лежит в основе толерантности к морфину». Боль . 115 (1–2): 50–59. дои : 10.1016/j.pain.2005.02.003 . ПМИД 15836969 . S2CID 7286123 .
- ^ Мохан С., Дэвис Р.Л., ДеСильва Ю., Стивенс К.В. (октябрь 2010 г.). «Двойная регуляция мю-опиоидных рецепторов в клетках нейробластомы SK-N-SH с помощью морфина и интерлейкина-1β: доказательства перекрестных помех между опиоидами и иммунитетом» . Журнал нейроиммунологии . 227 (1–2): 26–34. дои : 10.1016/j.jneuroim.2010.06.007 . ПМЦ 2942958 . ПМИД 20615556 .
- ^ Комацу Т., Сакурада С., Кацуяма С., Санаи К., Сакурада Т. (2009). Механизм аллодинии, вызываемой интратекально морфин-3-глюкуронидом у мышей . Международное обозрение нейробиологии. Том. 85. стр. 207–19. дои : 10.1016/S0074-7742(09)85016-2 . ISBN 978-0-12-374893-5 . ПМИД 19607972 .
- ^ Льюис С.С., Хатчинсон М.Р., Резвани Н., Лорам Л.С., Чжан Ю., Майер С.Ф. и др. (январь 2010 г.). «Доказательства того, что интратекальный морфин-3-глюкуронид может вызывать усиление боли через толл-подобный рецептор 4/MD-2 и интерлейкин-1бета» . Нейронаука . 165 (2): 569–583. doi : 10.1016/j.neuroscience.2009.10.011 . ПМК 2795035 . ПМИД 19833175 .
- ^ Шен Ч., Цай Р.Ю., Ши М.С., Лин С.Л., Тай Ю.Х., Чиен CC и др. (февраль 2011 г.). «Этанерцепт восстанавливает антиноцицептивный эффект морфина и подавляет нейровоспаление спинного мозга у устойчивых к морфию крыс» . Анестезия и анальгезия . 112 (2): 454–459. дои : 10.1213/ANE.0b013e3182025b15 . ПМИД 21081778 . S2CID 12295407 .
- ^ Хук М.А., Уошберн С.Н., Морено Г., Воллер С.А., Пуга Д., Ли К.Х. и др. (февраль 2011 г.). «Антагонист рецептора IL-1 блокирует вызванное морфином замедление двигательного восстановления после травмы спинного мозга» . Мозг, поведение и иммунитет . 25 (2): 349–359. дои : 10.1016/j.bbi.2010.10.018 . ПМК 3025088 . ПМИД 20974246 .
- ^ Уоткинс Л.Р., Хатчинсон М.Р., Райс К.К., Майер С.Ф. (ноябрь 2009 г.). «Плата» за опиоид-индуцированную активацию глии: повышение клинической эффективности опиоидов за счет воздействия на глию» . Тенденции в фармакологических науках . 30 (11): 581–591. дои : 10.1016/j.tips.2009.08.002 . ПМЦ 2783351 . ПМИД 19762094 .
- ^ Перейти обратно: а б с Хатчинсон М.Р., Чжан Ю., Браун К., Коутс Б.Д., Шридхар М., Шолар П.В. и др. (июль 2008 г.). «Нестереоселективное устранение нейропатической боли налоксоном и налтрексоном: вовлечение toll-подобного рецептора 4 (TLR4)» . Европейский журнал неврологии . 28 (1): 20–29. дои : 10.1111/j.1460-9568.2008.06321.x . ПМК 2588470 . ПМИД 18662331 .
- ^ Хатчинсон М.Р., Коутс Б.Д., Льюис С.С., Чжан Ю., Спрунгер Д.Б., Резвани Н. и др. (ноябрь 2008 г.). «Провоспалительные цитокины противодействуют острой и хронической анальгезии, вызванной опиоидами» . Мозг, поведение и иммунитет . 22 (8): 1178–1189. дои : 10.1016/j.bbi.2008.05.004 . ПМЦ 2783238 . ПМИД 18599265 .
- ^ Хатчинсон М.Р., Льюис С.С., Коутс Б.Д., Резвани Н., Чжан Ю., Визелер Дж.Л. и др. (май 2010 г.). «Возможное участие в активности толл-подобного рецептора 4/миелоидного фактора дифференцировки-2 неактивных изомеров опиоидов вызывает провоспаление позвоночника и связанные с ним поведенческие последствия» . Нейронаука . 167 (3): 880–893. doi : 10.1016/j.neuroscience.2010.02.011 . ПМЦ 2854318 . ПМИД 20178837 .
- ^ Лин С.Л., Цай Р.Ю., Тай Ю.Х., Чернг СН, Ву КТ, Йе СС и др. (февраль 2010 г.). «Сверхмалые дозы налоксона повышают экспрессию интерлейкина-10 и подавляют нейровоспаление в спинном мозге устойчивых к морфию крыс». Поведенческие исследования мозга . 207 (1): 30–36. дои : 10.1016/j.bbr.2009.09.034 . ПМИД 19799935 . S2CID 5128970 .
- ^ «Нейронаука: улучшение эффективности морфия» . Природа . 484 (7395): 419. 26 апреля 2012 г. Бибкод : 2012Natur.484Q.419. . дои : 10.1038/484419а . S2CID 52805136 .
- ^ Драль С. (22 августа 2012 г.). «Малые молекулы нацелены на толл-подобные рецепторы» . Новости химии и техники .
- ^ Манчек-Кебер М., Джерала Р. (февраль 2015 г.). «Постулаты проверки агонистов TLR4». Европейский журнал иммунологии . 45 (2): 356–370. дои : 10.1002/eji.201444462 . ПМИД 25476977 . S2CID 32029412 .
- ^ Ким ХМ, Ким ЮМ (октябрь 2018 г.). «HMGB1: Средство доставки ЛПС при пироптозе, опосредованном каспазой-11» . Иммунитет . 49 (4): 582–584. doi : 10.1016/j.immuni.2018.09.021 . ПМИД 30332623 .
- ^ Ромерио А, Пери Ф (2020). «Увеличение химического разнообразия низкомолекулярных модуляторов TLR4: обзор» . Границы в иммунологии . 11 : 1210. дои : 10.3389/fimmu.2020.01210 . ПМЦ 7381287 . ПМИД 32765484 .
- ^ Перейти обратно: а б с д и Хатчинсон М.Р., Лорам Л.С., Чжан Й., Шридхар М., Резвани Н., Беркельхаммер Д. и др. (июнь 2010 г.). «Доказательства того, что трициклические небольшие молекулы могут обладать активностью толл-подобного рецептора и белка 2 миелоидной дифференцировки» . Нейронаука . 168 (2): 551–563. doi : 10.1016/j.neuroscience.2010.03.067 . ПМЦ 2872682 . ПМИД 20381591 .
- ^ Чен Ф., Цзоу Л., Уильямс Б., Чао В. (ноябрь 2021 г.). «Нацеливание на Toll-подобные рецепторы при сепсисе: от лабораторных исследований к клиническим испытаниям» . Антиоксиданты и окислительно-восстановительная сигнализация . 35 (15): 1324–1339. дои : 10.1089/ars.2021.0005 . ПМЦ 8817700 . ПМИД 33588628 .
- ^ Цзя ZJ, Wu FX, Хуан QH, Лю JM (апрель 2012 г.). «[Toll-подобный рецептор 4: потенциальная терапевтическая мишень при нейропатической боли]». Чжунго И Сюэ Кэ Сюэ Юань Сюэ Бао. Acta Academiae Medicinae Sinicae . 34 (2): 168–173. дои : 10.3881/j.issn.1000-503X.2012.02.013 . ПМИД 22776604 .
- ^ Лан X, Хань X, Ли Q, Ли Q, Гао Ю, Ченг Т и др. (март 2017 г.). «Пиноцембрин защищает геморрагический мозг, прежде всего, ингибируя толл-подобный рецептор 4 и уменьшая микроглию фенотипа М1» . Мозг, поведение и иммунитет . 61 : 326–339. дои : 10.1016/j.bbi.2016.12.012 . ПМЦ 5453178 . ПМИД 28007523 .
- ^ Кайеда А., Такахаши М., Фукуда Х., Окамото Р., Моримото С., Гото М. и др. (декабрь 2019 г.). «Структурный дизайн, синтез и биологическая оценка ингибиторов киназы p38 MAP на основе имидазо[4,5-b]пиридин-2-она: Часть 2». ХимМедХим . 14 (24): 2093–2101. дои : 10.1002/cmdc.201900373 . ПМИД 31697454 . S2CID 207951964 .
- ^ Перейти обратно: а б с д Хатчинсон М.Р., Чжан Ю., Шридхар М., Эванс Дж.Х., Бьюкенен М.М., Чжао Техас и др. (январь 2010 г.). «Доказательства того, что опиоиды могут оказывать действие на толл-подобные рецепторы 4 и MD-2» . Мозг, поведение и иммунитет . 24 (1): 83–95. дои : 10.1016/j.bbi.2009.08.004 . ПМК 2788078 . ПМИД 19679181 .
- ^ Спир Э.М., Даулинг Д.Д., Озог Л.С., Сюй Дж., Ян Дж., Кеннади Дж. и др. (май 2017 г.). «Пентоксифиллин ингибирует опосредованную TLR и воспалением выработку воспалительных цитокинов in vitro в крови человека с большей эффективностью и эффективностью у новорожденных» . Педиатрические исследования . 81 (5): 806–816. дои : 10.1038/пр.2017.6 . ПМИД 28072760 . S2CID 47210724 .
- ^ Шуллер С.С., Висгрилл Л., Херндл Э., Шпиттлер А., Фёрстер-Вальдль Э., Садеги К. и др. (август 2017 г.). «Пентоксифиллин модулирует вызванное ЛПС гипервоспаление в моноцитах недоношенных детей in vitro» . Педиатрические исследования . 82 (2): 215–225. дои : 10.1038/пр.2017.41 . ПМИД 28288151 . S2CID 24897100 .
- ^ Нил М.Д., Цзя Х., Эйер Б., Гуд М., Герриеро С.Дж., Соди С.П. и др. (2013). «Открытие и проверка нового класса низкомолекулярных ингибиторов Toll-подобного рецептора 4 (TLR4)» . ПЛОС ОДИН . 8 (6): e65779. Бибкод : 2013PLoSO...865779N . дои : 10.1371/journal.pone.0065779 . ПМК 3680486 . ПМИД 23776545 .
- ^ Импеллиццери Д., Камполо М., Ди Паола Р., Брускетта Г., де Стефано Д., Эспозито Е. и др. (2015). «Ультрамикронизированный пальмитоилэтаноламид уменьшает воспаление на Th1-опосредованной модели колита» . Европейский журнал воспаления . 13 : 14–31. дои : 10.1177/1721727X15575869 . S2CID 79398556 .
Внешние ссылки
[ редактировать ]- Toll-Like+Receptor+4 в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)
- Обзор всей структурной информации, доступной в PDB для UniProt : O00206 (Toll-подобный рецептор 4) в PDBe-KB .