CD28
| CD28 | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Идентификаторы | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | CD28 , Tp44, молекула CD28 | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | ОМИМ : 186760 ; МГИ : 88327 ; Гомологен : 4473 ; Генные карты : CD28 ; ОМА : CD28 – ортологи | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||




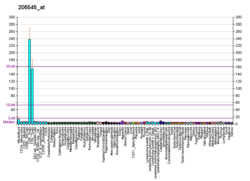

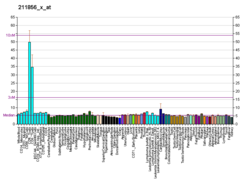
CD28 (кластер дифференциации 28) является одним из белков, экспрессируемых на Т-клетках , которые обеспечивают костимулирующие сигналы, необходимые для активации и выживания Т-клеток. Стимуляция Т-клеток через CD28 в дополнение к рецептору Т-клеток ( TCR ) может дать мощный сигнал для выработки различных интерлейкинов ( , IL-6 в частности ).
CD28 является рецептором белков CD80 (B7.1) и CD86 (B7.2). При активации лигандами Toll-подобных рецепторов экспрессия CD80 усиливается в антигенпрезентирующих клетках (APC). Экспрессия CD86 на антигенпрезентирующих клетках является конститутивной (экспрессия не зависит от факторов окружающей среды).
CD28 является единственным рецептором B7, конститутивно экспрессируемым на наивных Т-клетках . Ассоциация TCR наивной Т-клетки с комплексом MHC : антиген без взаимодействия CD28:B7 приводит к образованию Т-клетки, которая является анергической .
Кроме того, CD28 также был идентифицирован на стромальных клетках костного мозга, плазматических клетках, нейтрофилах и эозинофилах, но функциональное значение CD28 для этих клеток до конца не изучено. [5] [6] [7] [8] Обычно сообщается, что CD28 экспрессируется на 50% CD8+ Т-клеток и более чем на 80% CD4+ Т-клетках у человека, но в ходе активации некоторые Т-клетки теряют эту молекулу. Некоторые Т-клетки, подвергшиеся воздействию антигена, теряют CD28 и впоследствии могут быть повторно активированы без участия CD28. Эти CD28 − Т-клетки обычно характеризуются как антигенспецифические и терминально дифференцированные, и их часто называют Т-клетками памяти (ТМ) . Кроме того, уровень положительного CD28 снижается с возрастом. [9]
Будучи гомодимером двух цепей с доменами Ig, CD28 связывает молекулы B7 на APC и может способствовать пролиферации и дифференцировке Т-клеток, стимулировать выработку фактора роста и индуцировать экспрессию антиапоптотических белков. [10] По данным ряда исследований, после рождения все клетки человека экспрессируют CD28. Однако у взрослых 20-30% CD8+ Т-клеток теряют экспрессию CD28, тогда как у пожилых людей (+80 лет) до 50-60% CD8+ клеток теряют способность экспрессировать CD28. [11] Но эти утверждения лишь предполагают, что потеря экспрессии CD28 отмечает функциональную дифференцировку цитотоксических клеток памяти в рамках клональной экспансии. [12]
В общем, CD28 является основной костимулирующей молекулой для активации Т-клеток, но эффективная костимуляция необходима только для некоторой активации Т-клеток. В этом случае в отсутствие костимулирующих сигналов взаимодействие дендритных и Т-клеток приводит к анергии Т-клеток. Важность костимулирующего пути подчеркивается тем фактом, что антагонисты костимулирующих молекул нарушают иммунные реакции как in vitro , так и in vivo . [13] Но, как упоминалось ранее, в ходе активации, например, ТМ теряют эту молекулу и приобретают CD28-независимое существование. [14]
Сигнализация
[ редактировать ]CD28 обладает внутриклеточным доменом с несколькими остатками, которые имеют решающее значение для его эффективной передачи сигналов. Мотив YMNM, начинающийся с тирозина 170, в частности, имеет решающее значение для рекрутирования белков, содержащих SH2 -домен, особенно PI3K . [15] Грб2 [16] и Гадс . Остаток Y170 важен для индукции Bcl-xL посредством mTOR и усиления транскрипции IL-2 посредством PKCθ , но не влияет на пролиферацию и приводит к небольшому снижению продукции IL-2. Остаток N172 (как часть YMNM) важен для связывания Grb2 и Gads и, по-видимому, способен индуцировать мРНК стабильность NF-κB IL-2, но не транслокацию . Индукция NF-κB, по-видимому, в гораздо большей степени зависит от связывания Gads как с YMNM, так и с двумя богатыми пролином мотивами внутри молекулы. Однако мутация последней аминокислоты мотива, M173, которая не способна связывать PI3K, но способна связывать Grb2 и Gads, дает мало NF-κB или IL-2, что позволяет предположить, что эти Grb2 и Gads не способны компенсировать потеря ПИ3К. Транскрипция IL-2, по-видимому, имеет две стадии; Y170-зависимая, PI3K-зависимая начальная фаза, которая обеспечивает транскрипцию, и PI3K-независимая вторая фаза, которая зависит от образования иммунного синапса , что приводит к повышению стабильности мРНК IL-2. Оба необходимы для полноценного производства Ил-2.
CD28 также содержит два богатых пролином мотива, которые способны связывать SH3 -содержащие белки. Itk и Tec способны связываться с N-концом этих двух мотивов, который сразу следует за Y170 YMNM; Lck связывает С-конец. И Itk, и Lck способны фосфорилировать остатки тирозина, что затем позволяет связывать SH2-содержащие белки с CD28. Связывание Tec с CD28 усиливает продукцию IL-2, что зависит от связывания его доменов SH3 и PH с CD28 и PIP3 соответственно. С-концевой мотив, богатый пролином, в CD28 важен для доставки Lck и липидных рафтов в иммунный синапс через филамин-А. Мутация двух пролинов в С-концевом мотиве приводит к снижению пролиферации и продукции IL-2, но нормальной индукции Bcl-xL. Фосфорилирование тирозина в мотиве PYAP (Y191 в зрелом CD28 человека) образует сайт связывания с высоким сродством для домена SH2 киназы src Lck , который, в свою очередь, связывается с серинкиназой PKC-θ . [17]
Структура
[ редактировать ]Первая структура CD28 была получена в 2005 году группой биологов Т-клеток Оксфордского университета . [18]
Структура белка CD28 содержит 220 аминокислот, кодируемых геном, состоящим из четырех экзонов. Это гликозилированный гомодимер с дисульфидной связью массой 44 кДа, экспрессирующийся на поверхности клетки. Структура содержит парные домены суперсемейства иммуноглобулинов V-set (IgSF). Эти домены связаны с отдельными трансмембранными доменами и цитоплазматическими доменами, которые содержат важные сигнальные мотивы. [19] Как и CTLA4 , CD28 имеют очень схожие петли, аналогичные CDR3 . [20] В комплексе CD28-CD80 две молекулы CD80 сходятся так, что их проксимальные домены мембраны сталкиваются стерически, несмотря на наличие обоих сайтов связывания лигандов для CD28. [18]
члены семьи CD28
[ редактировать ]CD28 принадлежит к группе членов подсемейства костимулирующих молекул, которые характеризуются внеклеточным вариабельным иммуноглобулиноподобным доменом. Члены этого подсемейства также включают гомологичные рецепторы ICOS , CTLA4 , PD1 , PD1H и BTLA . [21] Тем не менее, только CD28 конститутивно экспрессируется на Т-клетках мыши, тогда как ICOS и CTLA4 индуцируются стимуляцией рецептора Т-клеток и в ответ на цитокины, такие как IL-2 . CD28 и CTLA4 очень гомологичны и конкурируют за один и тот же лиганд – CD80 и CD86 . [22] CTLA4 связывает CD80 и CD86 всегда сильнее, чем CD28, что позволяет CTLA4 конкурировать с CD28 за лиганд и подавлять ответы эффекторных Т-клеток. [23] Но было показано, что CD28 и CTLA4 оказывают противоположное влияние на стимуляцию Т-клеток. CD28 действует как активатор, а CTLA4 действует как ингибитор. [24] [25] ICOS и CD28 также являются близкородственными генами, но по функциям они не могут заменять друг друга. Противоположные роли CD28 и ICOS по сравнению с CTLA4 приводят к тому, что эти рецепторы действуют как реостат для иммунного ответа посредством конкурентных про- и противовоспалительных эффектов. [26]
Как мишень для наркотиков
[ редактировать ]Препарат TGN1412 , который был произведен немецкой биотехнологической компанией TeGenero и неожиданно вызвал в ходе испытаний полиорганную недостаточность , является суперагонистом CD28. К сожалению, часто игнорируется тот факт, что те же рецепторы существуют и на других клетках, помимо лимфоцитов . Также было обнаружено, что CD28 стимулирует эозинофильные гранулоциты , где его связывание с анти-CD28 приводит к высвобождению IL-2 , IL4 , IL-13 и IFN-γ . [27] [28]
Известно, что CD28 и CTL4 могут быть критическими регуляторами аутоиммунных заболеваний на мышиной модели. [29] [30] Но данных от пациентов о роли CD28 в заболеваниях человека меньше.
Другими потенциальными лекарствами, находящимися в доклинической разработке, являются агонистические аптамеры CD28 с иммуностимулирующими свойствами на модели опухоли мыши. [31] моноклональное анти-CD28 Fab'-антитело FR104 , [32] или октапептид AB103, который предотвращает гомодимеризацию CD28. [33]
Взаимодействия
[ редактировать ]Было показано, что CD28 взаимодействует с:
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Перейти обратно: а б с GRCh38: Версия Ensembl 89: ENSG00000178562 – Ensembl , май 2017 г.
- ^ Перейти обратно: а б с GRCm38: выпуск Ensembl 89: ENSMUSG00000026012 – Ensembl , май 2017 г.
- ^ «Ссылка на Human PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Ссылка на Mouse PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ Грей Паркин, Кирстин; Стефан, Роберт П.; Апиладо, Рон-Гран; Лилль-Элганиан, Дебора А.; Ли, Кельвин П.; Саха, Бхаскар; Витте, Памела Л. (1 сентября 2002 г.). «Экспрессия CD28 стромальными клетками костного мозга и ее участие в B-лимфопоэзе» . Журнал иммунологии . 169 (5): 2292–2302. дои : 10.4049/jimmunol.169.5.2292 . ISSN 0022-1767 . ПМИД 12193694 . S2CID 22737782 .
- ^ Розански, Шерил Х.; Аренс, Рамон; Карлсон, Луиза М.; Наир, Джаякумар; Бойсе, Лоуренс Х.; Чанан-Хан, Ашер А.; Шенбергер, Стивен П.; Ли, Кельвин П. (20 июня 2011 г.). «Устойчивый ответ антител зависит от функции CD28 в плазматических клетках, находящихся в костном мозге» . Журнал экспериментальной медицины . 208 (7): 1435–1446. дои : 10.1084/jem.20110040 . ISSN 1540-9538 . ПМК 3135367 . ПМИД 21690252 .
- ^ Венупрасад, К.; Параб, Прадип; Прасад, видеорегистратор; Шарма, Сатьян; Банерджи, Пинаки П.; Дешпанде, Маниша; Митра, Дипендра К.; Пал, Субрата; Бхадра, Ранджан; Митра, Дебашис; Саха, Бхаскар (май 2001 г.). <1536::aid-immu1536>3.0.co;2-8 «Иммунобиология экспрессии CD28 на нейтрофилах человека. I. CD28 регулирует миграцию нейтрофилов путем модуляции экспрессии CXCR-1» . Европейский журнал иммунологии . 31 (5): 1536–1543. doi : 10.1002/1521-4141(200105)31:5<1536::aid-immu1536>3.0.co;2-8 . ISSN 0014-2980 . ПМИД 11465111 . S2CID 22349635 .
- ^ Уэрли, Г.; Деко, В.; Луазо, С.; Лойенс, М.; Чихара, Дж.; Оно, Н.; Капрон, М. (сентябрь 2004 г.). «CD28 и секреторный иммуноглобулин А-зависимая активация эозинофилов: ингибирование высвобождения медиатора противоаллергическим препаратом суплатаст тозилат» . Клиническая и экспериментальная аллергия . 34 (9): 1379–1387. дои : 10.1111/j.1365-2222.2004.02036.x . ISSN 0954-7894 . ПМИД 15347370 . S2CID 21120027 .
- ^ Диас, Дэвид; Чара, Луи; Чеваррия, Юлиус; Убеда, Мэри; Муньос, Летисия; Барсенилья, Хью; Санчес, Майкл Анхель; Морено, Зайда; Монсеррат, Джордж; Альбильос, Августин; Прието, Альфред (2011). «Потеря поверхностных антигенов является консервативной особенностью апоптотических лимфоцитов нескольких видов млекопитающих» . Клеточная иммунология . 271 (1): 163–172. doi : 10.1016/j.cellimm.2011.06.018 . ISSN 0008-8749 . ПМИД 21745657 .
- ^ Эсенстен, Джонатан Х.; Хелу, Инес А.; Чопра, Гаурав; Вайс, Артур; Блюстоун, Джеффри А. (май 2016 г.). «Костимуляция CD28: от механизма к терапии» . Иммунитет . 44 (5): 973–988. doi : 10.1016/j.immuni.2016.04.020 . ПМЦ 4932896 . ПМИД 27192564 .
- ^ ФАНЬОНИ, ФФ; ВЕСКОВИНИ, Р.; МАЦЗОЛА, М.; БОЛОНЬЯ, Г.; НИГРО, Э.; ЛАВАГЕТТО, Г.; ФРАНЧЕШИ, К.; ПАССЕРИ, М.; САНСОНИ, П. (август 1996 г.). «Экспансия цитотоксических CD8 + CD28 - Т-клеток у здоровых стареющих людей, включая долгожителей» . Иммунология . 88 (4): 501–507. дои : 10.1046/j.1365-2567.1996.d01-689.x . ISSN 0019-2805 . ПМЦ 1456634 . ПМИД 8881749 .
- ^ Чемберлен, Уинстон Д.; Фальта, Майкл Т.; Котцин, Брайан Л. (март 2000 г.). «Функциональные подмножества клонально расширенных CD8+ Т-клеток памяти у пожилых людей» . Клиническая иммунология . 94 (3): 160–172. дои : 10.1006/clim.1999.4832 . ПМИД 10692235 .
- ^ Часовня, Хелен (2018). Основы клинической иммунологии: 6-е издание . Мэнсел Хейни, Сирадж А. Мисбах, Нил Сноуден, Войтех Тон. Прага. ISBN 978-80-7553-396-8 . OCLC 1031053171 .
{{cite book}}: CS1 maint: отсутствует местоположение издателя ( ссылка ) - ^ Моу, Д.; Эспиноза, Дж.; Ло, диджей; Кирк, AD (ноябрь 2014 г.). «CD28-негативные Т-клетки: является ли их потеря нашей выгодой?: CD28-негативные Т-клетки» . Американский журнал трансплантологии . 14 (11): 2460–2466. дои : 10.1111/ajt.12937 . ПМЦ 4886707 . ПМИД 25323029 .
- ^ Прасад К.В., Кай Ю.К., Рааб М., Дакворт Б., Кэнтли Л., Шолсон С.Е., Радд CE (март 1994 г.). «Т-клеточный антиген CD28 взаимодействует с липидкиназой фосфатидилинозитол-3-киназой посредством цитоплазматического мотива Tyr(P)-Met-Xaa-Met» . Труды Национальной академии наук Соединенных Штатов Америки . 91 (7): 2834–8. Бибкод : 1994PNAS...91.2834P . дои : 10.1073/pnas.91.7.2834 . ПМК 43465 . ПМИД 8146197 .
- ^ Шнайдер Х., Кай Ю.К., Прасад К.В., Шолсон С.Е., Радд CE (апрель 1995 г.). «Т-клеточный антиген CD28 связывается с комплексом GRB-2/SOS, регуляторами p21ras». Европейский журнал иммунологии . 25 (4): 1044–50. дои : 10.1002/eji.1830250428 . ПМИД 7737275 . S2CID 23540587 .
- ^ Конг К.Ф., Йокосука Т., Канониго-Балансио А.Дж., Исаков Н., Сайто Т., Альтман А. (октябрь 2011 г.). «Мотив в домене V3 киназы PKC-θ определяет его локализацию в иммунологическом синапсе и функционирует в Т-клетках посредством ассоциации с CD28» . Природная иммунология . 12 (11): 1105–12. дои : 10.1038/ni.2120 . ПМК 3197934 . ПМИД 21964608 .
- ^ Перейти обратно: а б Эванс Э.Дж., Эснуф Р.М., Мансо-Санчо Р., Гилберт Р.Дж., Джеймс Дж.Р., Ю.С. и др. (март 2005 г.). «Кристаллическая структура растворимого комплекса CD28-Fab». Природная иммунология . 6 (3): 271–9. дои : 10.1038/ni1170 . ПМИД 15696168 . S2CID 23630078 .
- ^ Каррено Б.М., Коллинз М. (апрель 2002 г.). «Семейство лигандов B7 и его рецепторы: новые пути костимуляции и ингибирования иммунных ответов». Ежегодный обзор иммунологии . 20 (1): 29–53. doi : 10.1146/annurev.immunol.20.091101.091806 . ПМИД 11861596 .
- ^ Чжан X, Шварц Дж.К., Алмо С.К., Натенсон С.Г. (март 2003 г.). «Кристаллическая структура рецептор-связывающего домена человеческого B7-2: понимание организации и передачи сигналов» . Труды Национальной академии наук Соединенных Штатов Америки . 100 (5): 2586–91. Бибкод : 2003PNAS..100.2586Z . дои : 10.1073/pnas.252771499 . ПМЦ 151384 . ПМИД 12606712 .
- ^ Чен Л., Flies DB (апрель 2013 г.). «Молекулярные механизмы костимуляции и коингибирования Т-клеток» . Обзоры природы. Иммунология . 13 (4): 227–42. дои : 10.1038/nri3405 . ПМЦ 3786574 . ПМИД 23470321 .
- ^ Линсли П.С., Кларк Э.А., Ледбеттер Дж.А. (июль 1990 г.). «Т-клеточный антиген CD28 опосредует адгезию с В-клетками путем взаимодействия с антигеном активации B7/BB-1» . Труды Национальной академии наук Соединенных Штатов Америки . 87 (13): 5031–5. Бибкод : 1990PNAS...87.5031L . дои : 10.1073/pnas.87.13.5031 . ПМК 54255 . ПМИД 2164219 .
- ^ Энгельхардт Дж. Дж., Салливан Т. Дж., Эллисон Дж. П. (июль 2006 г.). «Сверхэкспрессия CTLA-4 ингибирует ответы Т-клеток посредством CD28-B7-зависимого механизма» . Журнал иммунологии . 177 (2): 1052–61. дои : 10.4049/jimmunol.177.2.1052 . ПМИД 16818761 . S2CID 7990944 .
- ^ Краммель М.Ф., Эллисон Дж.П. (август 1995 г.). «CD28 и CTLA-4 оказывают противоположные эффекты на реакцию Т-клеток на стимуляцию» . Журнал экспериментальной медицины . 182 (2): 459–65. дои : 10.1084/jem.182.2.459 . ПМК 2192127 . ПМИД 7543139 .
- ^ Валунас Т.Л., Леншоу Д.Д., Баккер С.И., Линсли П.С., Фриман Г.Дж., Грин Дж.М. и др. (август 1994 г.). «CTLA-4 может действовать как негативный регулятор активации Т-клеток». Иммунитет . 1 (5): 405–13. дои : 10.1016/1074-7613(94)90071-х . ПМИД 7882171 .
- ^ Линтерман М.А., Ригби Р.Дж., Вонг Р., Сильва Д., Уизерс Д., Андерсон Г. и др. (февраль 2009 г.). «Рокин дифференцирует специализированные функции дублированных генов костимулирующих рецепторов Т-клеток CD28 и ICOS» . Иммунитет . 30 (2): 228–41. doi : 10.1016/j.immuni.2008.12.015 . ПМИД 19217324 .
- ^ Уэрли Г., Роджер Н., Луазо С., Домбрович Д., Капрон А., Капрон М. (август 1999 г.). «Экспрессия CD28 и CD86 эозинофилами человека и роль в секреции цитокинов типа 1 (интерлейкин 2 и гамма-интерферон): ингибирование комплексами иммуноглобулина А» . Журнал экспериментальной медицины . 190 (4): 487–95. дои : 10.1084/jem.190.4.487 . ПМК 2195599 . ПМИД 10449520 .
- ^ Уэрли Г., Лейси П., Юнес А.Б., Роджер Н., Луазо С., Мокбель Р., Капрон М. (октябрь 2002 г.). «Человеческие эозинофилы экспрессируют и высвобождают IL-13 после CD28-зависимой активации» . Журнал биологии лейкоцитов . 72 (4): 769–79. дои : 10.1189/jlb.72.4.769 . ПМИД 12377947 . S2CID 10820672 .
- ^ Саломон Б., Леншоу DJ, Ри Л., Ашуриан Н., Сингх Б., Шарп А., Блюстоун Дж.А. (апрель 2000 г.). «Костимуляция B7/CD28 необходима для гомеостаза CD4+CD25+ иммунорегуляторных Т-клеток, которые контролируют аутоиммунный диабет» . Иммунитет . 12 (4): 431–40. дои : 10.1016/s1074-7613(00)80195-8 . ПМИД 10795741 .
- ^ Тивол Э.А., Борриелло Ф., Швейцер А.Н., Линч В.П., Блюстоун Дж.А., Шарп А.Х. (ноябрь 1995 г.). «Потеря CTLA-4 приводит к массивной лимфопролиферации и фатальному разрушению тканей многих органов, что раскрывает критическую негативную регуляторную роль CTLA-4» . Иммунитет . 3 (5): 541–547. дои : 10.1016/1074-7613(95)90125-6 . ISSN 1074-7613 . ПМИД 7584144 . S2CID 46680106 .
- ^ Пастор Ф., Солдевилья М.М., Вильянуэва Х., Колониас Д., Иногес С., де Серио А.Л. и др. (июнь 2013 г.). «Аптамеры CD28 как мощные модуляторы иммунного ответа» . Молекулярная терапия: нуклеиновые кислоты . 2 (6): е98. дои : 10.1038/mtna.2013.26 . ПМК 3696906 . ПМИД 23756353 .
- ^ Пуарье Н., Мэри К., Дилек Н., Эрвуэ Дж., Мино Д., Бланшо Г., Ванхове Б. (октябрь 2012 г.). «Доклиническая эффективность и иммунологическая безопасность FR104, моновалентного Fab'-антитела-антагониста против CD28» . Американский журнал трансплантологии . 12 (10): 2630–40. дои : 10.1111/j.1600-6143.2012.04164.x . ПМИД 22759318 . S2CID 715661 .
- ^ Мирзоева С., Паунеску Т., Ванцер М.Б., Ширван А., Кемпфер Р., Волощак Г.Е., Смолл В. (23 июля 2014 г.). «Однократное введение p2TA (AB103), пептида-антагониста CD28, предотвращает воспалительные и тромботические реакции и защищает от желудочно-кишечных повреждений у мышей, подвергшихся облучению всего тела» . ПЛОС ОДИН . 9 (7): e101161. Бибкод : 2014PLoSO...9j1161M . дои : 10.1371/journal.pone.0101161 . ПМК 4108308 . ПМИД 25054224 .
- ^ Эллис Дж. Х., Эшман С., Берден М. Н., Килпатрик К. Е., Морс М. А., Хэмблин П. А. (июнь 2000 г.). «GRID: новый адапторный белок, родственный Grb-2, который взаимодействует с активированным костимулирующим рецептором Т-клеток CD28» . Журнал иммунологии . 164 (11): 5805–14. дои : 10.4049/jimmunol.164.11.5805 . ПМИД 10820259 . S2CID 25739159 .
- ^ Оккенхауг К., Роттапель Р. (август 1998 г.). «Grb2 образует индуцибельный белковый комплекс с CD28 посредством взаимодействия 3-го домена Src-пролина» . Журнал биологической химии . 273 (33): 21194–202. дои : 10.1074/jbc.273.33.21194 . ПМИД 9694876 . S2CID 39280280 .
- ^ Нунес Х.А., Труне А., Олив Д., Кантрелл Д.А. (январь 1996 г.). «Передача сигнала костимулирующим рецептором CD28 на Т-клетках. Регуляция B7-1 и B7-2 адаптерных молекул тирозинкиназы» . Журнал биологической химии . 271 (3): 1591–8. дои : 10.1074/jbc.271.3.1591 . ПМИД 8576157 . S2CID 37740924 .
- ^ Пажес Ф., Рагено М., Класен С., Баттифора М., Куэз Д., Свит Р. и др. (апрель 1996 г.). «Две отдельные внутрицитоплазматические области молекулы адгезии Т-клеток CD28 участвуют в ассоциации фосфатидилинозитол-3-киназы» . Журнал биологической химии . 271 (16): 9403–9. дои : 10.1074/jbc.271.16.9403 . ПМИД 8621607 . S2CID 12566111 .
Дальнейшее чтение
[ редактировать ]- Эсенстен Дж. Х., Хелу Ю. А., Чопра Г., Вайс А., Блюстоун Дж. А. (май 2016 г.). «Костимуляция CD28: от механизма к терапии» . Иммунитет . 44 (5): 973–88. doi : 10.1016/j.immuni.2016.04.020 . ПМЦ 4932896 . ПМИД 27192564 .
- Леншоу ди-джей, Валунас Т.Л., Блюстоун Дж.А. (1996). «Система костимуляции Т-клеток CD28/B7». Ежегодный обзор иммунологии . 14 : 233–58. doi : 10.1146/annurev.immunol.14.1.233 . ПМИД 8717514 .
- Гринфилд Э.А., Нгуен К.А., Качру В.К. (1998). «Костимуляция CD28/B7: обзор». Критические обзоры по иммунологии . 18 (5): 389–418. doi : 10.1615/critrevimmunol.v18.i5.10 . ПМИД 9784967 .
- Чанг Т.Т., Качру В.К., Шарп А.Х. (2002). «Роль пути B7-CD28/CTLA-4 при аутоиммунных заболеваниях». Пути передачи сигнала при аутоиммунитете . Современные направления в области аутоиммунитета. Том. 5. С. 113–30. дои : 10.1159/000060550 . ISBN 978-3-8055-7308-5 . ПМИД 11826754 .
{{cite book}}:|journal=игнорируется ( помогите ) - Бур-Джордан Х., Блюстон Дж. А. (январь 2002 г.). «Функция CD28: баланс костимулирующих и регуляторных сигналов». Журнал клинической иммунологии . 22 (1): 1–7. дои : 10.1023/А:1014256417651 . ПМИД 11958588 . S2CID 38060684 .
- Гринуэй А.Л., Холлоуэй Г., Макфи Д.А., Эллис П., Корналл А., Лидман М. (апрель 2003 г.). «Контроль Nef ВИЧ-1 клеточных сигнальных молекул: несколько стратегий содействия репликации вируса». Журнал биологических наук . 28 (3): 323–35. дои : 10.1007/BF02970151 . ПМИД 12734410 . S2CID 33749514 .
- Бенишу С., Бенмера А. (январь 2003 г.). «[Нэф ВИЧ и белки K3/K5 вируса, ассоциированного с саркомой Капоши: «паразиты» пути эндоцитоза]» . Медицина/Науки . 19 (1): 100–6. дои : 10.1051/medsci/2003191100 . ПМИД 12836198 .
- Толструп М., Остергаард Л., Лаурсен А.Л., Педерсен С.Ф., Дах М. (апрель 2004 г.). «ВИЧ/ВИV ускользают от иммунного надзора: сосредоточьтесь на Нефе». Текущие исследования ВИЧ . 2 (2): 141–51. дои : 10.2174/1570162043484924 . ПМИД 15078178 .
- Андерсон Дж.Л., Хоуп Т.Дж. (апрель 2004 г.). «Аксессуарные белки ВИЧ и выживание в клетке-хозяине». Текущие отчеты о ВИЧ/СПИДе . 1 (1): 47–53. дои : 10.1007/s11904-004-0007-x . ПМИД 16091223 . S2CID 34731265 .
- Ли Л, Ли Х.С., Пауза К.Д., Букринский М., Чжао Р.Ю. (2006). «Роль вспомогательных белков ВИЧ-1 в вирусном патогенезе и взаимодействии хозяин-патоген» . Клеточные исследования . 15 (11–12): 923–34. дои : 10.1038/sj.cr.7290370 . ПМИД 16354571 . S2CID 24253878 .
- Печь V, Верхасселт Б (январь 2006 г.). «Моделирование эффектов Nef ВИЧ-1 тимуса». Текущие исследования ВИЧ . 4 (1): 57–64. дои : 10.2174/157016206775197583 . ПМИД 16454711 .
Внешние ссылки
[ редактировать ]- Таблица антигенов мышиного CD
- Таблица антигенов CD человека
- человека Местоположение генома CD28 и страница сведений о гене CD28 в браузере генома UCSC .
- Обзор всей структурной информации, доступной в PDB для UniProt : P10747 (поверхностный гликопротеин CD28, специфичный для Т-клеток) на PDBe-KB .
галерея PDB |
|---|
