CCR5
| CCR5 | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Идентификаторы | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | CCR5 , CC-CKR-5, CCCKR5, CCR-5, CD195, CKR-5, CKR5, CMKBR5, IDDM22, хемокиновый рецептор 5 с мотивом CC (ген/псевдоген), рецептор 5 хемокина с мотивом CC | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | Опустить : 601373 ; МГИ : 107182 ; Гомологен : 37325 ; Генные карты : CCR5 ; OMA : CCR5 — ортологи | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||




Рецептор хемокинов CC 5-го типа , также известный как CCR5 или CD195 , представляет собой белок на поверхности лейкоцитов , который участвует в работе иммунной системы, поскольку действует как рецептор для хемокинов . [5]
У человека ген CCR5 , кодирующий белок CCR5, расположен на коротком (p) плече в положении 21 хромосомы 3 . Некоторые популяции унаследовали мутацию Дельта 32 , приводящую к генетической делеции части гена CCR5. Гомозиготные носители этой мутации устойчивы к заражению макрофаготропными (М-тропными) штаммами ВИЧ-1 . [6] [7] [8] [9] [10] [11]
Функция
[ редактировать ]Белок CCR5 принадлежит к бета-хемокиновых рецепторов семейству интегральных мембранных белков . [12] [13] Это рецептор, связанный с G-белком. [12] который действует как хемокиновый рецептор в группе хемокинов CC .
CCR5 Родственные лиганды включают CCL3 , CCL4 (также известные как MIP 1 α и 1 β соответственно) и CCL3L1 . [14] [15] Кроме того, CCR5 взаимодействует с CCL5 ( белком хемотаксического цитокина, также известным как RANTES). [14] [16] [17]
CCR5 преимущественно экспрессируется на Т-клетках , макрофагах , дендритных клетках , эозинофилах , микроглии и субпопуляции клеток рака молочной железы или простаты. [18] [19] Экспрессия CCR5 избирательно индуцируется во время процесса раковой трансформации и не экспрессируется в нормальных эпителиальных клетках молочной железы или предстательной железы. Примерно 50% случаев рака молочной железы человека экспрессируют CCR5, в первую очередь при тройном негативном раке молочной железы . [18] Ингибиторы CCR5 блокировали миграцию и метастазирование клеток рака молочной железы и простаты, экспрессирующих CCR5, что позволяет предположить, что CCR5 может функционировать как новая терапевтическая мишень. [18] [19] [20] Недавние исследования показывают, что CCR5 экспрессируется в подмножестве раковых клеток с характеристиками раковых стволовых клеток, которые, как известно, вызывают резистентность к терапии, и что ингибиторы CCR5 увеличивают количество клеток, убитых современной химиотерапией. [21] Вполне вероятно, что CCR5 играет роль в воспалительных реакциях на инфекцию, хотя его точная роль в нормальной иммунной функции неясна. Области этого белка также имеют решающее значение для связывания хемокиновых лигандов, функционального ответа рецептора и активности корецептора ВИЧ. [22]
Модуляция активности CCR5 способствует непатогенному течению инфекции вирусом иммунодефицита обезьян (SIV) у нескольких африканских видов приматов, не являющихся человеком, которые являются длительными естественными хозяевами SIV и избегают иммунодефицита при заражении. [23] Эти регуляторные механизмы включают: генетические делеции, которые аннулируют экспрессию CCR5 на клеточной поверхности, [24] подавление CCR5 на поверхности CD4+ Т-клеток, в частности на клетках памяти, [25] и отсроченное начало экспрессии CCR5 на CD4+ Т-клетках во время развития. [26] [27]

ВИЧ
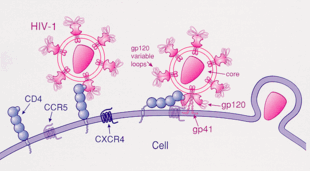
[ редактировать ]ВИЧ-1 чаще всего использует хемокиновые рецепторы CCR5 и/или CXCR4 в качестве корецепторов для проникновения в целевые иммунологические клетки. [28] Эти рецепторы расположены на поверхности иммунных клеток хозяина, обеспечивая путь проникновения вируса ВИЧ-1 для заражения клетки. [29] Структура гликопротеина оболочки ВИЧ-1 необходима для обеспечения проникновения вируса ВИЧ-1 в клетку-мишень. [29] Структура оболочечного гликопротеина состоит из двух белковых субъединиц, отщепленных от белка-предшественника Gp160, кодируемого геном env ВИЧ-1 : внешней субъединицы Gp120 и трансмембранной субъединицы Gp41. [29] Эта оболочечная гликопротеиновая структура представляет собой шипообразную структуру, расположенную на поверхности вириона, и состоит из тримера гетеродимеров Gp120-Gp41. [29] Белок оболочки Gp120 является имитатором хемокина. [28] Хотя ему не хватает уникальной структуры хемокина, он все же способен связываться с хемокиновыми рецепторами CCR5 и CXCR4. [28] Во время инфекции ВИЧ-1 субъединица гликопротеина оболочки Gp120 связывается с гликопротеином CD4 и корецептором ВИЧ-1, экспрессируемым на клетке-мишени, образуя гетеротримерный комплекс. [28] Образование этого комплекса стимулирует высвобождение фузогенного пептида, вызывающего слияние вирусной мембраны с мембраной клетки-мишени. [28] Поскольку связывание только с CD4 может иногда приводить к выделению gp120, gp120 должен затем связаться с корецептором CCR5, чтобы продолжить слияние. Тирозинсульфатированный аминоконец этого корецептора является «важной детерминантой» связывания с гликопротеином gp120. [30] Корецептор также распознает область V1-V2 gp120 и мостиковый лист (антипараллельный четырехцепочечный β-лист, который соединяет внутренний и внешний домены gp120). Стебель V1-V2 может влиять на «использование корецептора посредством своего пептидного состава, а также степени N-связанного гликозилирования». Однако, в отличие от V1-V2, петля V3 сильно вариабельна и, таким образом, является наиболее важным фактором, определяющим специфичность корецептора. [30] Нормальные лиганды этого рецептора, RANTES , MIP-1β и MIP-1α , способны подавлять инфекцию ВИЧ-1 in vitro . У людей, инфицированных ВИЧ, вирусы, использующие CCR5, являются преобладающим видом, выделяемым на ранних стадиях вирусной инфекции. [31] предполагая, что эти вирусы могут иметь избирательное преимущество во время передачи или острой фазы заболевания. Более того, по крайней мере половина всех инфицированных людей на протяжении всего периода заражения содержит только вирусы, использующие CCR5.
CCR5 является основным корецептором, используемым gp120 последовательно с CD4. В результате этого связывания gp41, другой белковый продукт gp160, высвобождается из метастабильной конформации и встраивается в мембрану клетки-хозяина. Хотя это не было подтверждено, связывание gp120-CCR5 включает два важных этапа: 1) Тирозинсульфатированный аминоконец этого корецептора является «важной детерминантой» связывания с gp120 (как указывалось ранее) 2) После этапа 1 ., должно существовать взаимное действие (синергия, взаимодействие) между gp120 и трансмембранными доменами CCR5. [30]
CCR5 необходим для распространения штамма R5 вируса ВИЧ-1. [32] Знание механизма, с помощью которого этот штамм ВИЧ-1 опосредует инфекцию, побудило исследования по разработке терапевтических мер по блокированию функции CCR5. [33] Ряд новых экспериментальных препаратов против ВИЧ, называемых антагонистами рецептора CCR5 , был разработан для вмешательства в связывание между белком оболочки Gp120 и корецептором ВИЧ CCR5. [32] Эти экспериментальные препараты включают PRO140 ( CytoDyn ), Викривирок (исследования фазы III были отменены в июле 2010 года) ( Schering Plow ), Аплавирок (GW-873140) ( GlaxoSmithKline ) и Маравирок (UK-427857) ( Pfizer ). Маравирок был одобрен FDA к использованию в августе 2007 года. [32] На данный момент это единственный ингибитор, одобренный FDA для клинического использования, став таким образом первым ингибитором CCR5. [30] Проблема этого подхода заключается в том, что, хотя CCR5 является основным корецептором, с помощью которого ВИЧ инфицирует клетки, он не единственный такой корецептор. Вполне возможно, что под давлением отбора ВИЧ эволюционирует и начнет использовать другой корецептор. Однако изучение устойчивости вируса к AD101, молекулярному антагонисту CCR5, показало, что резистентные вирусы не переключались на другой корецептор (CXCR4), а продолжали использовать CCR5: они либо связывались с альтернативными доменами CCR5, либо с рецептором на более высокая близость. Однако, поскольку существует еще один корецептор, вполне вероятно, что отсутствие гена CCR5 не делает человека невосприимчивым к вирусу; человеку просто будет сложнее заразиться им. Кроме того, вирус все еще имеет доступ к CD4. В отличие от CCR5, который не требуется (о чем свидетельствуют те, кто ведет здоровый образ жизни даже при отсутствии этого гена в результате мутации дельта32), CD4 имеет решающее значение для системы иммунной защиты организма. [34] Даже без наличия какого-либо корецептора (даже CCR5), вирус все равно может проникать в клетки, если gp41 претерпит изменение (включая его цитоплазматический хвост), которое приведет к независимости CD4 без необходимости CCR5 и/или CXCR4. как дверной проем. [35]
Рак
[ редактировать ]Экспрессия CCR5 индуцируется в эпителиальных клетках молочной железы и предстательной железы при трансформации. [18] [19] Индукция экспрессии CCR5 способствует клеточной инвазии, миграции и метастазированию. [5] [18] [21] Индукция метастазирования включает возвращение к месту метастазирования. Было показано, что ингибиторы CCR5, включая маравирок и леронлимаб, блокируют метастазирование в легкие клеточных линий рака молочной железы человека. [18] [36] В доклинических исследованиях на иммунокомпетентных мышах ингибиторы CCR5 блокировали метастазы в кости и мозг. [19] Ингибиторы CCR5 также уменьшают инфильтрацию опухолеассоциированных макрофагов. [37] Клиническое исследование фазы 1 ингибитора CCR5 у пациентов с метастатическим раком толстой кишки, получавших тяжелое предварительное лечение, продемонстрировало объективный клинический ответ и снижение метастатической опухолевой нагрузки. [38]
Мозг
[ редактировать ]Повышенные уровни CCR5 являются частью воспалительной реакции на инсульт и смерть. Блокирование CCR5 с помощью Маравирока (препарата, одобренного для лечения ВИЧ) может ускорить выздоровление после инсульта. [39] [40]
В развивающемся мозге хемокиновые рецепторы, такие как CCR5, влияют на миграцию и соединение нейронов. После инсульта они, по-видимому, уменьшают количество мест соединения на нейронах вблизи места повреждения. [39]
CCR5-Δ32
[ редактировать ]CCR5-Δ32 (или CCR5-D32 или CCR5 дельта 32) представляет собой аллель CCR5. [41] [42]
CCR5 Δ32 представляет собой делецию из 32 пар оснований, которая вводит преждевременный стоп-кодон в локус рецептора CCR5, что приводит к нефункциональному рецептору. [43] [44] CCR5 необходим для проникновения М-тропного вируса ВИЧ-1. [45] Лица, гомозиготные (обозначенные Δ32/Δ32) по CCR5 Δ32, не экспрессируют функциональные рецепторы CCR5 на поверхности своих клеток и устойчивы к инфекции ВИЧ-1 , несмотря на многократное воздействие высокого риска. [45] Лица, гетерозиготные (+/Δ32) по мутантному аллелю, имеют снижение функциональных рецепторов CCR5 на поверхности клеток более чем на 50% из-за димеризации между мутантными рецепторами и рецепторами дикого типа, которая препятствует транспортировке CCR5 на поверхность клетки. [46] Гетерозиготы-носители устойчивы к инфекции ВИЧ-1 по сравнению с дикими типами, а при заражении гетерозиготы демонстрируют сниженную вирусную нагрузку и прогрессирование СПИДа на 2-3 года медленнее по сравнению с дикими типами. [43] [45] [47] Также было показано, что гетерозиготность по этому мутантному аллелю улучшает вирусологический ответ на антиретровирусное лечение. [48] CCR5 Δ32 имеет частоту (гетерозиготных) аллелей 9% в Европе и частоту гомозигот 1%. [49]
Недавние исследования показывают, что CCR5 Δ32 улучшает когнитивные функции и память. В 2016 году исследователи показали, что удаление гена CCR5 у мышей значительно улучшило их память. [50] CCR5 является мощным подавителем пластичности нейронов, обучения и памяти; Чрезмерная активация CCR5 вирусными белками может способствовать когнитивному дефициту, связанному с ВИЧ. [51]
Эволюционная история и возраст аллеля
[ редактировать ]Аллель CCR5 Δ32 отличается недавним происхождением, неожиданно высокой частотой и отчетливым географическим распространением. [52] которые вместе предполагают, что (а) он возник в результате единственной мутации и (б) исторически подвергался положительному отбору.
В двух исследованиях использовался анализ сцепления для оценки возраста делеции CCR5 Δ32, предполагая, что количество рекомбинаций и мутаций, наблюдаемых в геномных областях, окружающих делецию CCR5 Δ32, будет пропорционально возрасту делеции. [42] [53] Используя выборку из 4000 человек из 38 этнических групп, Stephens et al. подсчитали, что делеция CCR5-Δ32 произошла 700 лет назад (275–1875, 95% доверительный интервал). Другая группа, Libert et al. (1998) использовали микросателлитные мутации, чтобы оценить возраст мутации CCR5 Δ32 в 2100 лет (700–4800, 95% доверительный интервал). На основании наблюдаемых событий рекомбинации они оценили возраст мутации в 2250 лет (900–4700, 95% доверительный интервал). [53] Третья гипотеза основана на градиенте частоты аллелей с севера на юг в Европе, который показывает, что самая высокая частота аллелей наблюдалась в странах Северной Европы , а самая низкая частота аллелей - в южной Европе. Поскольку исторически викинги оккупировали эти страны, вполне возможно, что аллель распространилась по Европе из-за расселения викингов в VIII-X веках. [54] Позднее викинги были заменены варягами в России, что, возможно, способствовало наблюдаемому сдвигу частоты аллелей с востока на запад. [52] [54]
ВИЧ-1 первоначально был передан от шимпанзе ( Pan troglodytes ) человеку в начале 1900-х годов в юго-восточном Камеруне, Африка, [55] в результате контакта с инфицированной кровью и биологическими жидкостями во время разделки мяса диких животных. [56] Однако ВИЧ-1 фактически отсутствовал в Европе до 1980-х годов. [57] Таким образом, учитывая средний возраст аллеля CCR5-Δ32, составляющий примерно 1000 лет, можно установить, что ВИЧ-1 не оказывал давления отбора на человеческую популяцию достаточно долго, чтобы достичь нынешних частот. [52] Следовательно, другие патогены были предложены в качестве агентов положительной селекции для CCR5 Δ32, включая бубонную чуму ( Yersinia pestis ) и оспа ( Variola major ).Другие данные свидетельствуют о том, что частота аллелей подвергалась отрицательному давлению отбора из-за патогенов, которые получили более широкое распространение во время римской экспансии. [58] Идея о том, что отрицательный отбор сыграл роль в низкой частоте аллеля, также подтверждается экспериментами с использованием нокаутных мышей и гриппа А, которые продемонстрировали, что присутствие рецептора CCR5 важно для эффективного ответа на патоген. [59] [60]
Доказательства единственной мутации
[ редактировать ]Некоторые данные свидетельствуют о том, что аллель CCR5 Δ32 эволюционировал только один раз. [52] Во-первых, CCR5 Δ32 имеет относительно высокую частоту в нескольких различных европейских популяциях, но сравнительно отсутствует в популяциях азиатов, Ближнего Востока и американских индейцев. [42] предполагая, что единственная мутация произошла после расхождения европейцев от их африканского предка. [42] [43] [61] Во-вторых, анализ генетического сцепления показывает, что мутация возникает на однородном генетическом фоне, подразумевая, что мутация унаследована от общего предка. [53] Это было продемонстрировано, показав, что аллель CCR5 Δ32 находится в сильном неравновесии по сцеплению с высокополиморфными микросателлитами. Более 95% хромосом CCR5 Δ32 также несут аллель IRI3.1-0, а 88% — аллель IRI3.2. Напротив, микросателлитные маркеры IRI3.1-0 и IRI3.2-0 были обнаружены только в 2 или 1,5% хромосом, несущих аллель CCR5 дикого типа. [53] Эти доказательства неравновесия по сцеплению подтверждают гипотезу о том, что большинство, если не все, аллели CCR5 Δ32 возникли в результате одного мутационного события. Наконец, аллель CCR5 Δ32 имеет уникальное географическое распространение, указывающее на единственное северное происхождение с последующей миграцией. Исследование, измеряющее частоты аллелей в 18 европейских популяциях, выявило градиент с севера на юг: самые высокие частоты аллелей наблюдаются в финских и мордовских популяциях (16%), а самые низкие — на Сардинии (4%). [53]
Положительный выбор
[ редактировать ]В отсутствие отбора одной мутации потребовалось бы примерно 127 500 лет, чтобы частота в популяции выросла до 10%. [42] По оценкам, основанным на скорости генетической рекомбинации и мутаций, возраст аллеля составляет от 1000 до 2000 лет. Это несоответствие является признаком положительного отбора.
Предполагается, что ВИЧ-1 проник в человеческое население Африки в начале 1900-х годов. [55] но о симптоматических инфекциях не сообщалось до 1980-х годов. Таким образом, эпидемия ВИЧ-1 слишком молода, чтобы быть источником положительного отбора, который увеличил частоту CCR5 Δ32 с нуля до 10% за 2000 лет.
Защита от бубонной чумы . Стивенс и др. (1998) предполагают, что бубонная чума ( Yersinia pestis ) оказала положительное селективное давление на CCR5 Δ32. [42] Эта гипотеза была основана на времени и серьезности пандемии «черной смерти», унесшей жизни 30% европейского населения всех возрастов в период с 1346 по 1352 год. [62] После Черной смерти были менее серьезные, периодические эпидемии. В отдельных городах наблюдался высокий уровень смертности, но общая смертность в Европе составляла всего несколько процентов. [62] [63] [64] В 1655–1656 годах вторая пандемия, получившая название «Великая чума», унесла жизни 15–20% населения Лондона. [65] [66] [ сомнительно – обсудить ] Важно отметить, что эпидемии чумы были периодическими. Бубонная чума — зоонозное заболевание, поражающее преимущественно грызунов, распространяющееся блохами и лишь изредка заражающее человека. [67] Передача бубонной чумы от человека к человеку не происходит, хотя она может возникать при легочной чуме, поражающей легкие. [68] Только когда плотность грызунов низкая, инфицированные блохи вынуждены питаться альтернативными хозяевами, такими как люди, и в этих обстоятельствах может возникнуть эпидемия среди людей. [67] Основываясь на популяционно-генетических моделях, Гальвани и Слаткин (2003) утверждают, что прерывистый характер эпидемий чумы не создал достаточно сильной силы отбора, чтобы довести частоту аллеля CCR5 Δ32 до 10% в Европе. [41] Чтобы проверить эту гипотезу, Гальвани и Слаткин (2003) смоделировали историческое давление отбора, вызванное чумой и оспой. [41]
Защита от оспы . Чума была смоделирована согласно историческим источникам. [69] [70] а повозрастная смертность от оспы была получена из возрастного распределения оспенных захоронений в Йорке (Англия) между 1770 и 1812 годами. [63] Оспой преимущественно заражаются молодые представители населения дорепродуктивного возраста, поскольку они являются единственными людьми, которые не были иммунизированы или умерли от перенесенной инфекции. Поскольку оспа преимущественно убивает представителей популяции, не достигших репродуктивного возраста, она оказывает более сильное селективное давление, чем чума. [41] В отличие от чумы, оспа не имеет резервуара среди животных и передается только от человека к человеку. [71] [72] Авторы подсчитали, что если бы чума отбирала CCR5 Δ32, частота этого аллеля все равно была бы менее 1%, в то время как оспа оказала селективную силу, достаточную для достижения 10%.
Гипотеза о том, что оспа осуществляет положительную селекцию по CCR5 Δ32, также биологически правдоподобна, поскольку поксвирусы, как и ВИЧ, проникают в лейкоциты с помощью хемокиновых рецепторов. [73] Напротив, Yersinia pestis — это бактерия с совершенно другой биологией.
Хотя европейцы — единственная группа, имеющая субпопуляции с высокой частотой CCR5 Δ32, они не единственная популяция, подвергшаяся селекции в результате оспы, которая была распространена по всему миру до того, как в 1980 году она была объявлена искорененной. Самые ранние безошибочные описания оспа появилась в V веке нашей эры в Китае, в VII веке нашей эры в Индии и Средиземноморье и в X веке нашей эры в Юго-Западной Азии. [72] Напротив, мутация CCR5 Δ 32 обнаружена только в популяциях Европы, Западной Азии и Северной Африки. [74] Аномально высокая частота CCR5 Δ32 в этих популяциях, по-видимому, требует как уникального происхождения в Северной Европе, так и последующего отбора с помощью оспы.
Потенциальные затраты
[ редактировать ]CCR5 Δ32 может быть полезен для хозяина при некоторых инфекциях (например, ВИЧ-1, возможно, оспа), но вреден при других (например, клещевой энцефалит , вирус Западного Нила ). Полезна или вредна функция CCR5 в контексте конкретной инфекции, зависит от сложного взаимодействия между иммунной системой и патогеном. [75]
В целом исследования показывают, что мутация CCR5 Δ32 может играть пагубную роль в постинфекционных воспалительных процессах, которые могут повредить ткани и вызвать дальнейшую патологию. [76] Наилучшие доказательства этой предполагаемой антагонистической плейотропии обнаружены при флавивирусных инфекциях. В целом многие вирусные инфекции протекают бессимптомно или вызывают лишь легкие симптомы у подавляющего большинства населения. Однако у некоторых невезучих людей наблюдается особенно деструктивное клиническое течение, которое иначе необъяснимо, но, по-видимому, генетически опосредовано. Было обнаружено, что пациенты, гомозиготные по CCR5 Δ32, подвергаются более высокому риску нейроинвазивной формы клещевого энцефалита (вызванного флавивирусом ). [77] Кроме того, функциональный CCR5 может потребоваться для предотвращения симптоматического заболевания после заражения вирусом Западного Нила, другим флавивирусом; CCR5 Δ32 был связан с ранним развитием симптомов и более выраженными клиническими проявлениями после заражения вирусом Западного Нила. [78]
Это открытие у людей подтвердило ранее наблюдавшийся эксперимент на животной модели гомозиготности CCR5 Δ32. После заражения вирусом Западного Нила у мышей CCR5 Δ32 заметно увеличивались титры вируса в центральной нервной системе и наблюдалась повышенная смертность. [79] по сравнению с мышами дикого типа, что позволяет предположить, что экспрессия CCR5 необходима для создания сильной защиты хозяина от вируса Западного Нила.
Медицинские применения
[ редактировать ]Генетический подход, включающий внутритела, блокирующие экспрессию CCR5, был предложен в качестве лечения ВИЧ-1- инфицированных лиц. [80] Когда Т-клетки модифицировались так, что они больше не экспрессируют CCR5, смешивались с немодифицированными Т-клетками, экспрессирующими CCR5, а затем подвергались заражению ВИЧ-1, модифицированные Т-клетки, которые не экспрессируют CCR5, в конечном итоге захватывали культуру, как ВИЧ-1. убивает немодифицированные Т-клетки. Этот же метод можно использовать in vivo для создания пула устойчивых к вирусу клеток у инфицированных людей. [80]
Эта гипотеза была проверена на пациенте со СПИДом, у которого также развился миелолейкоз , и которого лечили химиотерапией для подавления рака. Затем для восстановления иммунной системы использовался трансплантат костного мозга, содержащий стволовые клетки от подходящего донора. Однако трансплантация была выполнена от донора с 2 копиями гена мутации CCR5-Δ32. Через 600 дней пациент был здоров и имел неопределяемый уровень ВИЧ в крови, а также в исследованных тканях головного мозга и прямой кишки. [7] [81] Перед трансплантацией также были обнаружены низкие уровни ВИЧ X4 , который не использует рецептор CCR5. Однако после трансплантации этот тип ВИЧ также не был обнаружен. [7] Однако этот результат согласуется с наблюдением, что клетки, экспрессирующие вариант белка CCR5-Δ32, лишены на своей поверхности рецепторов CCR5 и CXCR4 , тем самым придавая устойчивость к широкому спектру вариантов ВИЧ, включая ВИЧX4. [82] Спустя более шести лет у пациента сохранилась устойчивость к ВИЧ, и было объявлено, что он излечился от ВИЧ-инфекции. [8]
Набор ВИЧ-положительных пациентов в клиническое исследование начался в 2009 году, в ходе которого клетки пациентов были генетически модифицированы с помощью нуклеазы цинковых пальцев , чтобы нести признак CCR5-Δ32, а затем повторно вводились в организм в качестве потенциального лечения ВИЧ. [83] [84] Результаты, представленные в 2014 году, были многообещающими. [11]
Вдохновленный первым человеком, когда-либо излечившимся от ВИЧ, «Берлинским пациентом» , компания StemCyte начала сотрудничество с банками пуповинной крови по всему миру для систематического скрининга образцов пуповинной крови на наличие мутации CCR5, начиная с 2011 года. [85] [86] [87]
В ноябре 2018 года Цзянькуй Хэ объявил, что он отредактировал два человеческих эмбриона, пытаясь отключить ген CCR5, который кодирует рецептор, который ВИЧ использует для проникновения в клетки. Он сказал, что девочки-близнецы, Лулу и Нана , родились несколькими неделями ранее и что девочки все еще несут функциональные копии CCR5 наряду с отключенными CCR5 ( мозаицизм ), следовательно, они все еще уязвимы к ВИЧ. Работа была широко осуждена как неэтичная, опасная и преждевременная. [88] [89]
См. также
[ редактировать ]- Открытие и разработка антагонистов рецептора CCR5
- Ингибитор входа
- ВИЧ-тропизм
- Стивен Крон
- иммунитет к ВИЧ
Ссылки
[ редактировать ]- ^ Jump up to: а б с GRCh38: Версия Ensembl 89: ENSG00000160791 – Ensembl , май 2017 г.
- ^ Jump up to: а б с GRCm38: выпуск Ensembl 89: ENSMUSG00000079227 – Ensembl , май 2017 г.
- ^ «Ссылка на Human PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Ссылка на Mouse PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ Jump up to: а б Цзяо X, Наваб О, Патель Т, Коссенков АВ, Халама Н, Джагер Д и др. (октябрь 2019 г.). «Последние достижения в борьбе с раком CCR5 и его роль в иммуноонкологии» . Исследования рака . 79 (19): 4801–4807. дои : 10.1158/0008-5472.CAN-19-1167 . ПМК 6810651 . ПМИД 31292161 .
- ^ де Сильва Э., депутат Штумпфа (декабрь 2004 г.). «ВИЧ и аллель устойчивости CCR5-Delta32» . Письма FEMS по микробиологии . 241 (1): 1–12. дои : 10.1016/j.femsle.2004.09.040 . ПМИД 15556703 .
- ^ Jump up to: а б с Хюттер Г., Новак Д., Мосснер М., Ганепола С., Мюссиг А., Аллерс К. и др. (февраль 2009 г.). «Долгосрочный контроль ВИЧ с помощью трансплантации стволовых клеток CCR5 Delta32/Delta32» . Медицинский журнал Новой Англии . 360 (7): 692–698. doi : 10.1056/NEJMoa0802905 . ПМИД 19213682 .
- ^ Jump up to: а б Аллерс К., Хюттер Г., Хофманн Дж., Лодденкемпер С., Ригер К., Тиль Э. и др. (март 2011 г.). «Доказательства лечения ВИЧ-инфекции путем трансплантации стволовых клеток CCR5Δ32/Δ32» . Кровь . 117 (10): 2791–2799. doi : 10.1182/blood-2010-09-309591 . ПМИД 21148083 .
- ^ Чжэнь А, Кухня S (декабрь 2013 г.). «Генная терапия ВИЧ-инфекции на основе стволовых клеток» . Вирусы . 6 (1): 1–12. дои : 10.3390/v6010001 . ПМЦ 3917429 . ПМИД 24368413 .
- ^ Кей М.А., Уокер Б.Д. (март 2014 г.). «Инженерия клеточной устойчивости к ВИЧ». Медицинский журнал Новой Англии . 370 (10): 968–969. дои : 10.1056/NEJMe1400593 . ПМИД 24597871 .
- ^ Jump up to: а б Тебас П., Стейн Д., Тан В.В., Фрэнк И., Ван С.К., Ли Дж. и др. (март 2014 г.). «Редактирование гена CCR5 в аутологичных CD4 Т-клетках людей, инфицированных ВИЧ» . Медицинский журнал Новой Англии . 370 (10): 901–910. дои : 10.1056/NEJMoa1300662 . ПМК 4084652 . ПМИД 24597865 .
- ^ Jump up to: а б «CCR5 - хемокиновый (CC-мотив) рецептор 5 (ген/псевдоген)» . Домашний справочник по генетике . Архивировано из оригинала 24 сентября 2009 года . Проверено 30 августа 2009 г.
- ^ Самсон М., Лаббе О., Моллеро С., Вассар Г., Пармантье М. (март 1996 г.). «Молекулярное клонирование и функциональная экспрессия нового гена рецептора CC-хемокина человека». Биохимия . 35 (11): 3362–3367. дои : 10.1021/bi952950g . ПМИД 8639485 .
- ^ Jump up to: а б Стрюиф С., Ментен П., Ленартс Дж.П., Пут В., Д'Хезе А., Де Клерк Э. и др. (июль 2001 г.). «Различные способности связывания природных изоформ LD78beta макрофагального воспалительного белка-1-альфа с хемокиновыми рецепторами CC 1, 3 и 5 влияют на их анти-ВИЧ-1-активность и хемотаксическую активность в отношении нейтрофилов и эозинофилов» . Европейский журнал иммунологии . 31 (7): 2170–2178. doi : 10.1002/1521-4141(200107)31:7<2170::AID-IMMU2170>3.0.CO;2-D . ПМИД 11449371 .
- ^ Миякава Т., Обару К., Маэда К., Харада С., Мицуя Х. (февраль 2002 г.). «Идентификация аминокислотных остатков, имеющих решающее значение для LD78beta, варианта воспалительного белка-1-альфа макрофагов человека, связывания с CCR5 и ингибирования репликации вируса иммунодефицита человека R5 типа 1» . Журнал биологической химии . 277 (7): 4649–4655. дои : 10.1074/jbc.M109198200 . ПМИД 11734558 .
- ^ Слимани Х., Шарно Н., Фрог Э., Саффар Л., Васси Р., Вита С. и др. (октябрь 2003 г.). «Взаимодействие RANTES с синдеканом-1 и синдеканом-4, экспрессируемыми первичными макрофагами человека » Биохимические и биофизические акты (ББА) – Биомембраны . 1617 (1–2): 80–88. дои : 10.1016/j.bbamem.2003.09.006 . ПМИД 14637022 .
- ^ Праудфут А.Е., Фритчли С., Борлат Ф., Шоу Дж.П., Вильбуа Ф., Цвален С. и др. (апрель 2001 г.). «Мотив BBXB RANTES является основным сайтом связывания гепарина и контролирует селективность рецептора» . Журнал биологической химии . 276 (14): 10620–10626. дои : 10.1074/jbc.M010867200 . ПМИД 11116158 .
- ^ Jump up to: а б с д и ж Веласко-Веласкес М., Цзяо Х., Де Ла Фуэнте М., Пестель Т.Г., Эртель А., Лисанти М.П. и др. (август 2012 г.). «Антагонист CCR5 блокирует метастазирование базальных клеток рака молочной железы» . Исследования рака . 72 (15): 3839–3850. дои : 10.1158/0008-5472.CAN-11-3917 . ПМИД 22637726 .
- ^ Jump up to: а б с д Сиколи Д., Цзяо Х., Джу Х., Веласко-Веласкес М., Эртель А., Аддия С. и др. (декабрь 2014 г.). «Антагонисты рецептора CCR5 блокируют метастазирование в кость линий метастатических клеток рака предстательной железы, трансформированных онкогеном v-Src» . Исследования рака . 74 (23): 7103–7114. дои : 10.1158/0008-5472.CAN-14-0612 . ПМЦ 4294544 . ПМИД 25452256 .
- ^ Веласко-Веласкес М., Холальпа В., Пестель Р.Г. (ноябрь 2014 г.). «Потенциал воздействия на CCL5/CCR5 при раке молочной железы». Мнение экспертов о терапевтических целях . 18 (11): 1265–1275. дои : 10.1517/14728222.2014.949238 . ПМИД 25256399 . S2CID 7976259 .
- ^ Jump up to: а б Цзяо X, Веласко-Веласкес М.А., Ван М., Ли З., Руй Х., Пек А.Р. и др. (апрель 2018 г.). «CCR5 управляет восстановлением повреждений ДНК и увеличением стволовых клеток рака молочной железы» . Исследования рака . 78 (7): 1657–1671. дои : 10.1158/0008-5472.CAN-17-0915 . ПМК 6331183 . ПМИД 29358169 .
- ^ Бармания Ф, Пеппер М.С. (декабрь 2013 г.). «Хемокиновый рецептор CC пятого типа (CCR5): новая мишень для контроля ВИЧ-инфекции» . Прикладная и трансляционная геномика . 2 (а): 3–16. дои : 10.1016/j.atg.2013.05.004 . ПМЦ 5133339 . ПМИД 27942440 .
- ^ Ясинска А.Дж., Пандреа I, Апетрей С (27 января 2022 г.). «CCR5 как корецептор вируса иммунодефицита человека и вирусов иммунодефицита обезьян: прототип романа любви и ненависти» . Границы в иммунологии . 13 : 835994. дои : 10.3389/fimmu.2022.835994 . ISSN 1664-3224 . ПМЦ 8829453 . ПМИД 35154162 .
- ^ Чен З., Квон Д., Джин З., Монард С., Телфер П., Джонс М.С. и др. (декабрь 1998 г.). «Естественное заражение гомозиготного красношапочного мангабея delta24 CCR5 R2b-тропным вирусом иммунодефицита обезьян» . Журнал экспериментальной медицины . 188 (11): 2057–2065. дои : 10.1084/jem.188.11.2057 . ПМК 2212380 . ПМИД 9841919 .
- ^ Пайардини М., Серваси Б., Рейес-Авилес Е., Миччи Л., Ортис А.М., Чахруди А. и др. (июнь 2011 г.). «Низкие уровни инфекции SIV в CD⁴⁺ Т-клетках центральной памяти мангабея связаны с ограниченной экспрессией CCR5» . Природная медицина . 17 (7): 830–836. дои : 10.1038/нм.2395 . ПМЦ 3253129 . ПМИД 21706028 .
- ^ Ма Д., Ясинска А.Дж., Фейертаг Ф., Виджевардана В., Кристофф Дж., Хе Т. и др. (май 2014 г.). «Факторы, связанные с передачей вируса иммунодефицита симана у естественного африканского примата-хозяина, не являющегося человеком, в дикой природе» . Журнал вирусологии . 88 (10): 5687–5705. дои : 10.1128/JVI.03606-13 . ПМК 4019088 . ПМИД 24623416 .
- ^ Пандреа И., Онанга Р., Сукьер С., Муинга-Ондем А., Бурри О., Боунс М. и др. (июнь 2008 г.). «Недостаток CD4+ CCR5+ Т-клеток может предотвратить передачу вируса иммунодефицита обезьян естественным хозяевам-приматам, не являющимся человеком, при грудном вскармливании» . Журнал вирусологии . 82 (11): 5501–5509. дои : 10.1128/JVI.02555-07 . ПМК 2395173 . ПМИД 18385229 .
- ^ Jump up to: а б с д и Мерфи ПМ (февраль 2001 г.). «Вирусная эксплуатация и подрыв иммунной системы посредством мимикрии хемокинов» . Природная иммунология . 2 (2): 116–122. дои : 10.1038/84214 . ПМИД 11175803 . S2CID 29364407 .
- ^ Jump up to: а б с д Алхатиб Г. (март 2009 г.). «Биология CCR5 и CXCR4» . Современное мнение о ВИЧ и СПИДе . 4 (2): 96–103. doi : 10.1097/COH.0b013e328324bbec . ПМЦ 2718543 . ПМИД 19339947 .
- ^ Jump up to: а б с д «Ингибиторы CCR5 и ВИЧ» .
- ^ Андерсон Дж., Аккина Р. (сентябрь 2007 г.). «Полный нокдаун CCR5 с помощью миРНК, экспрессируемых лентивирусным вектором, и защита трансгенных макрофагов от инфекции ВИЧ-1» . Генная терапия . 14 (17): 1287–1297. дои : 10.1038/sj.gt.3302958 . ПМИД 17597795 .
- ^ Jump up to: а б с Либерман-Блюм СС, Фунг Х.Б., Бандрес Х.К. (июль 2008 г.). «Маравирок: антагонист рецептора CCR5 для лечения инфекции ВИЧ-1». Клиническая терапия . 30 (7): 1228–1250. дои : 10.1016/s0149-2918(08)80048-3 . ПМИД 18691983 .
- ^ Назари Р., Джоши С. (август 2008 г.). «CCR5 как мишень для генной терапии ВИЧ-1». Современная генная терапия . 8 (4): 264–272. дои : 10.2174/156652308785160674 . ПМИД 18691022 .
- ^ Старр Д.Б. (19 ноября 2009 г.). «Есть ли люди, генетически предрасположенные к иммунитету к ВИЧ?» . Технический интерактив . Спросите генетика . Проверено 5 августа 2024 г.
- ^ Тейлор Б.М., Фоулк Дж.С., Флинко Р., Эредиа А., ДеВико А., Рейтц М. (июнь 2008 г.). «Изменение gp41 вируса иммунодефицита человека приводит к снижению зависимости CCR5 и независимости CD4» . Журнал вирусологии . 82 (11): 5460–5471. дои : 10.1128/JVI.01049-07 . ПМК 2395218 . ПМИД 18353949 .
- ^ Цзяо X, Ван М, Чжан З, Ли З, Ни Д, Эштон А.В. и др. (январь 2021 г.). «Леронлимаб, гуманизированное моноклональное антитело к CCR5, блокирует клеточные метастазы рака молочной железы и усиливает гибель клеток, вызванную химиотерапией, повреждающей ДНК» . Исследование рака молочной железы . 23 (1): 11. дои : 10.1186/s13058-021-01391-1 . ПМЦ 7825185 . PMID 33485378 .
- ^ Франкенбергер С., Рабе Д., Байнер Р., Санкарашарма Д., Чада К., Крауш Т. и др. (октябрь 2015 г.). «Супрессоры метастазов регулируют микроокружение опухоли, блокируя рекрутирование прометастатических макрофагов, связанных с опухолью» . Исследования рака . 75 (19): 4063–4073. дои : 10.1158/0008-5472.CAN-14-3394 . ПМЦ 4592465 . ПМИД 26238785 .
- ^ Халама Н., Зёрниг И., Бертел А., Калерт С., Клупп Ф., Суарес-Кармона М. и др. (апрель 2016 г.). «Эксплуатацию опухолевых иммунных клеток при метастазах колоректального рака можно эффективно контролировать с помощью терапии против CCR5 у онкологических больных» . Раковая клетка . 29 (4): 587–601. дои : 10.1016/j.ccell.2016.03.005 . ПМИД 27070705 .
- ^ Jump up to: а б «Препарат от ВИЧ может улучшить восстановление после инсульта» . НаукаМаг . 21 февраля 2019 года . Проверено 22 февраля 2019 г.
- ^ Джой М.Т., Бен Ассаяг Э., Шабашов-Стоун Д., Лираз-Зальцман С., Маццителли Дж., Аренас М. и др. (февраль 2019 г.). «CCR5 является терапевтической мишенью для восстановления после инсульта и черепно-мозговой травмы» . Клетка . 176 (5): 1143–1157.e13. дои : 10.1016/j.cell.2019.01.044 . ПМК 7259116 . ПМИД 30794775 .
- ^ Jump up to: а б с д Гальвани А.П., Слаткин М. (декабрь 2003 г.). «Оценка чумы и оспы как исторического селективного давления на аллель устойчивости к ВИЧ CCR5-Delta 32» . Труды Национальной академии наук Соединенных Штатов Америки . 100 (25): 15276–15279. Бибкод : 2003PNAS..10015276G . дои : 10.1073/pnas.2435085100 . ПМК 299980 . ПМИД 14645720 .
- ^ Jump up to: а б с д и ж Стивенс Дж.К., Райх Д.Э., Гольдштейн Д.Б., Шин Х.Д., Смит М.В., Кэррингтон М. и др. (июнь 1998 г.). «Датирование происхождения аллеля устойчивости к СПИДу CCR5-Delta32 путем слияния гаплотипов» . Американский журнал генетики человека . 62 (6): 1507–1515. дои : 10.1086/301867 . ПМЦ 1377146 . ПМИД 9585595 .
- ^ Jump up to: а б с Дин М., Кэррингтон М., Винклер С., Хаттли Г.А., Смит М.В., Алликметс Р. и др. (сентябрь 1996 г.). «Генетическое ограничение инфекции ВИЧ-1 и прогрессирования до СПИДа за счет делеции аллели структурного гена CKR5. Исследование роста и развития гемофилии, многоцентровое когортное исследование СПИДа, многоцентровое когортное исследование гемофилии, когортное исследование города Сан-Франциско, исследование ALIVE» . Наука . 273 (5283): 1856–1862. Бибкод : 1996Sci...273.1856D . дои : 10.1126/science.273.5283.1856 . ПМИД 8791590 . S2CID 21160580 .
- ^ Самсон М., Либерт Ф., Доранц Б.Дж., Ракер Дж., Лиснард С., Фарбер С.М. и др. (август 1996 г.). «Устойчивость к инфекции ВИЧ-1 у лиц европеоидной расы, несущих мутантные аллели гена хемокинового рецептора CCR-5». Природа . 382 (6593): 722–725. Бибкод : 1996Natur.382..722S . дои : 10.1038/382722a0 . ПМИД 8751444 . S2CID 4370818 .
- ^ Jump up to: а б с Лю Р., Пакстон В.А., Чой С., Черадини Д., Мартин С.Р., Хорук Р. и др. (август 1996 г.). «Гомозиготный дефект корецептора ВИЧ-1 объясняет устойчивость некоторых людей, неоднократно контактировавших с инфекцией ВИЧ-1» . Клетка . 86 (3): 367–377. дои : 10.1016/s0092-8674(00)80110-5 . ПМИД 8756719 . S2CID 16965624 .
- ^ Бенкиран М., Джин Д.Ю., Чун Р.Ф., Коуп Р.А., Жанг К.Т. (декабрь 1997 г.). «Механизм трансдоминантного ингибирования CCR5-опосредованной инфекции ВИЧ-1 с помощью ccr5delta32» . Журнал биологической химии . 272 (49): 30603–30606. дои : 10.1074/jbc.272.49.30603 . ПМИД 9388191 .
- ^ Майкл Н.Л., Луи Л.Г., Рорбо А.Л., Шульц К.А., Дайхофф Д.Э., Ван К.Э. и др. (октябрь 1997 г.). «Роль полиморфизмов CCR5 и CCR2 в передаче ВИЧ-1 и прогрессировании заболевания». Природная медицина . 3 (10): 1160–1162. дои : 10.1038/нм1097-1160 . ПМИД 9334732 . S2CID 23196048 .
- ^ Лоришесс Ж.Дж., Персо А., Теодору И., Рузиу С., Дельфраси Ж.Ф., Мейер Л. (май 2007 г.). «Улучшение вирусологического ответа на высокоактивную антиретровирусную терапию у ВИЧ-1-инфицированных пациентов, несущих делецию CCR5 Delta32» . ВИЧ-медицина . 8 (4): 213–219. дои : 10.1111/j.1468-1293.2007.00455.x . ПМИД 17461848 . S2CID 24900064 .
- ^ Мерфи К., Уивер С. (2016). Иммунобиология Джейнвей, 9-е издание . Гирляндная наука. п. 582. ИСБН 978-0-8153-4550-3 .
- ^ «Мозг китайских близнецов CRISPR мог быть случайно улучшен » Обзор технологий . Проверено 22 февраля 2019 г.
- ^ Чжоу М., Гринхилл С., Хуан С., Сильва Т.К., Сано Ю., Ву С. и др. (декабрь 2016 г.). «CCR5 подавляет пластичность коры, а также обучение и память гиппокампа» . электронная жизнь . 5 . doi : 10.7554/eLife.20985 . ПМЦ 5213777 . ПМИД 27996938 .
- ^ Jump up to: а б с д Гальвани А.П., Новембре Дж. (февраль 2005 г.). «Эволюционная история мутации CCR5-Delta32, устойчивой к ВИЧ» . Микробы и инфекции . 7 (2): 302–309. дои : 10.1016/j.micinf.2004.12.006 . ПМИД 15715976 .
- ^ Jump up to: а б с д и Либер Ф., Кошо П., Бекман Г., Самсон М., Аксенова М., Цао А. и др. (март 1998 г.). «Мутация deltaccr5, обеспечивающая защиту от ВИЧ-1 среди населения европеоидной расы, имеет единственное и недавнее происхождение в Северо-Восточной Европе» . Молекулярная генетика человека . 7 (3): 399–406. дои : 10.1093/hmg/7.3.399 . ПМИД 9466996 .
- ^ Jump up to: а б Люкотт Дж. (сентябрь 2001 г.). «Распространение делеции 32 пар оснований гена CCR5 в Западной Европе. Гипотеза о возможном распространении мутации викингами в исторические времена». Иммунология человека . 62 (9): 933–936. дои : 10.1016/S0198-8859(01)00292-0 . ПМИД 11543895 .
- ^ Jump up to: а б Кил Б.Ф., Ван Хеуверсвин Ф., Ли Ю., Бэйлз Э., Такехиса Дж., Сантьяго М.Л. и др. (июль 2006 г.). «Резервуары пандемического и непандемического ВИЧ-1 у шимпанзе» . Наука . 313 (5786): 523–526. Бибкод : 2006Sci...313..523K . дои : 10.1126/science.1126531 . ПМК 2442710 . ПМИД 16728595 .
- ^ Хан Б.Х., Шоу GM, Де Кок К.М., Sharp PM (январь 2000 г.). «СПИД как зооноз: последствия для науки и общественного здравоохранения». Наука . 287 (5453): 607–614. Бибкод : 2000Sci...287..607H . дои : 10.1126/science.287.5453.607 . ПМИД 10649986 .
- ^ Матич С. «ВИЧ/СПИД в Европе: переход от смертного приговора к лечению хронических заболеваний» (PDF) . Всемирная организация здравоохранения.
- ^ Фор Э., Ройер-Каренци М. (декабрь 2008 г.). «Сформировано ли европейское пространственное распределение устойчивого к ВИЧ-1 аллеля CCR5-Delta32 разрушением патоценоза из-за исторической римской экспансии?» (PDF) . Инфекция, генетика и эволюция . 8 (6): 864–874. дои : 10.1016/j.meegid.2008.08.007 . ПМИД 18790087 . S2CID 32881202 .
- ^ Доусон Т.К., Бек М.А., Кузиел В.А., Хендерсон Ф., Маэда Н. (июнь 2000 г.). «Контрастные эффекты дефицита CCR5 и CCR2 при воспалительной реакции легких на вирус гриппа А» . Американский журнал патологии . 156 (6): 1951–1959. дои : 10.1016/S0002-9440(10)65068-7 . ПМК 1850091 . ПМИД 10854218 .
- ^ Варгас А.Е., Сехим Г., Корреа Дж.Ф., Гомеш П.А., Маседо Г.С., де Медейрос Р.М. и др. (июль 2009 г.). «Плюсы и минусы отсутствующего хемокинового рецептора — комментарии Эрика Фора и Мануэлы к статье «Сформировано ли европейское пространственное распределение устойчивого к ВИЧ-1 аллеля CCR5-D32 в результате разрушения патоценоза из-за исторической римской экспансии?» Ройер-Каренци (2008)». Инфекция, генетика и эволюция . 9 (4): 387–389. дои : 10.1016/j.meegid.2009.01.001 . ПМИД 19472441 .
- ^ О'Брайен С.Дж., Дин М. (сентябрь 1997 г.). «В поисках генов устойчивости к СПИДу». Научный американец . 277 (3): 44–51. Бибкод : 1997SciAm.277c..44O . doi : 10.1038/scientificamerican0997-44 . ПМИД 9274039 .
- ^ Jump up to: а б МакЭведи С. (февраль 1988 г.). «Бубонная чума». Научный американец . 258 (2): 118–123. Бибкод : 1988SciAm.258b.118M . doi : 10.1038/scientificamerican0288-118 . ПМИД 3055286 .
- ^ Jump up to: а б Галера С (1998). Демография городов раннего Нового времени: Йорк в шестнадцатом и семнадцатом веках . Лондон: Макмиллан.
- ^ Хэтчер Дж (1977). Чума, население и английская экономика 1348-1530 гг . Лондон: Макмиллан.
- ^ «Великая чума в Лондоне, 1665 год» . Заражение, исторические взгляды на болезни и эпидемии . Гарвардский университет . Проверено 2 марта 2015 г.
- ^ «ДНК в лондонской могиле может помочь раскрыть тайны Великой чумы» . 8 сентября 2016 г. Архивировано из оригинала 9 сентября 2016 г. Проверено 18 сентября 2016 г.
- ^ Jump up to: а б Килинг М.Дж., Гиллиган, Калифорния (ноябрь 2000 г.). «Бубонная чума: метапопуляционная модель зооноза» . Слушания. Биологические науки . 267 (1458): 2219–2230. дои : 10.1098/rspb.2000.1272 . ПМК 1690796 . ПМИД 11413636 .
- ^ Эпплби AB (1980). «Исчезновение чумы: продолжающаяся загадка». Обзор экономической истории . 33 (2): 161–173. дои : 10.2307/2595837 . JSTOR 2595837 . ПМИД 11614424 .
- ^ Рассел Дж.С. (1948). Британское средневековое население . Альбукерке: Издательство Университета Нью-Мексико.
- ^ Твигг Дж. (1984). Черная смерть: биологическая переоценка . Лондон: Бэтсфод.
- ^ Чапин CV (1913). «Изменения типов инфекционных заболеваний, как показывает история оспы в Соединенных Штатах, 1815–1912 годы» . J Заразить Дис . 13 (2): 171–196. дои : 10.1093/infdis/13.2.171 .
- ^ Jump up to: а б Феннер Ф (1998). «Историческая виньетка: жизнь с поксвирусами и издателями». Достижения в области исследования вирусов . 51 : 1–33. дои : 10.1016/S0065-3527(08)60782-4 . ISBN 978-0-12-039851-5 . ПМИД 9891584 .
- ^ Лалани А.С., Мастерс Дж., Цзэн В., Барретт Дж., Панну Р., Эверетт Х. и др. (декабрь 1999 г.). «Использование хемокиновых рецепторов поксвирусами». Наука . 286 (5446): 1968–1971. дои : 10.1126/science.286.5446.1968 . ПМИД 10583963 .
- ^ Новембре Дж., Гальвани А.П., Слаткин М. (ноябрь 2005 г.). «Географическое распространение аллеля устойчивости к ВИЧ CCR5 Delta32» . ПЛОС Биология . 3 (11): е339. дои : 10.1371/journal.pbio.0030339 . ПМК 1255740 . ПМИД 16216086 .
- ^ ОБрайен С.Дж. (март 2024 г.). «Наследие волшебного гена - : от CCR5-∆32 открытия до клинической пользы за одно поколение». Труды Национальной академии наук Соединенных Штатов Америки . 121 (12): e2321907121. дои : 10.1073/pnas.2321907121 . PMC 10962972. PMID 38457490 .
- ^ Кляйн Р.С. (январь 2008 г.). «Движущаяся цель: множественная роль CCR5 при инфекционных заболеваниях» . Журнал инфекционных болезней . 197 (2): 183–186. дои : 10.1086/524692 . ПМИД 18179384 .
- ^ Киндберг Э., Микиин А., Акс С., Акерлинд Б., Вене С., Линдквист Л. и др. (январь 2008 г.). «Делекция гена хемокинового рецептора 5 (CCR5) связана с клещевым энцефалитом» . Журнал инфекционных болезней . 197 (2): 266–269. дои : 10.1086/524709 . ПМИД 18179389 .
- ^ Лим Дж.К., Луи С.И., Глейзер С., Джин С., Джонсон Б., Джонсон Х. и др. (январь 2008 г.). «Генетический дефицит хемокинового рецептора CCR5 является сильным фактором риска симптоматической инфекции вируса Западного Нила: метаанализ 4 когорт эпидемии в США» . Журнал инфекционных болезней . 197 (2): 262–265. дои : 10.1086/524691 . ПМИД 18179388 .
- ^ Гласс В.Г., Лим Дж.К., Холера Р., Плетнев А.Г., Гао Дж.Л., Мерфи П.М. (октябрь 2005 г.). «Хемокиновый рецептор CCR5 способствует доставке лейкоцитов в мозг и выживанию при инфекции вируса Западного Нила» . Журнал экспериментальной медицины . 202 (8): 1087–1098. дои : 10.1084/jem.20042530 . ПМК 2213214 . ПМИД 16230476 .
- ^ Jump up to: а б Стейнбергер П., Андрис-Видхопф Дж., Бюлер Б., Торбетт Б.Е., Барбас К.Ф. (январь 2000 г.). «Функциональная делеция рецептора CCR5 путем внутриклеточной иммунизации приводит к образованию клеток, устойчивых к CCR5-зависимой инфекции ВИЧ-1 и слиянию клеток» . Труды Национальной академии наук Соединенных Штатов Америки . 97 (2): 805–810. Бибкод : 2000PNAS...97..805S . дои : 10.1073/pnas.97.2.805 . ПМЦ 15412 . ПМИД 10639161 .
- ^ Шуфс М. (7 ноября 2008 г.). «Врач, мутация и потенциальное лекарство от СПИДа» . Уолл Стрит Джорнал . Проверено 15 декабря 2010 г.
- ^ Агравал Л., Лу Х, Цинвэнь Дж., ВанХорн-Али З., Николеску И.В., МакДермотт Д.Х. и др. (март 2004 г.). «Роль белка CCR5Delta32 в устойчивости к вирусу иммунодефицита человека типа 1 R5, R5X4 и X4 в первичных клетках CD4+» . Журнал вирусологии . 78 (5): 2277–2287. doi : 10.1128/JVI.78.5.2277-2287.2004 . ПМК 369216 . ПМИД 14963124 .
- ^ «Аутологичные Т-клетки, генетически модифицированные по гену CCR5 нуклеазой цинковых пальцев SB-728 для ВИЧ (цинковые пальцы)» . Национальные институты здравоохранения США. 9 декабря 2009 года . Проверено 30 декабря 2009 г.
- ^ Уэйд Н. (28 декабря 2009 г.). «Цинковые пальцы могут стать ключом к возрождению генной терапии» . Нью-Йорк Таймс . Проверено 30 декабря 2009 г.
- ^ Корбин З. (июль 2012 г.). «Начался план по поиску лекарства от ВИЧ» . Ланцет . 380 (9838): 203–204. дои : 10.1016/s0140-6736(12)61199-4 . ПМИД 22826833 . S2CID 34773481 .
- ^ Гонсалес Г., Парк С., Чен Д., Армитидж С., Шпалл Э., Берингер Р. (сентябрь 2011 г.). «Идентификация и частота использования единиц пуповинной крови, устойчивых к ВИЧ CCR5Δ32/Δ32, в больницах Хьюстона» . ВИЧ-медицина . 12 (8): 481–486. дои : 10.1111/j.1468-1293.2010.00911.x . ПМК 4021858 . ПМИД 21375684 .
- ^ Петц Л.Д., Бернетт Дж.К., Ли Х., Ли С., Тонай Р., Бакалинская М. и др. (2015). «Прогресс в лечении ВИЧ-инфекции с помощью трансплантации гемопоэтических клеток» . Стволовые клетки и клонирование: достижения и применение . 8 : 109–116. дои : 10.2147/SCCAA.S56050 . ПМЦ 4524463 . ПМИД 26251620 .
- ^ Бегли С. (28 ноября 2018 г.). «На фоне возмущения китайский ученый защищает создание детей с отредактированными генами» . СТАТ .
- ^ Коллинз Ф.С. (28 ноября 2018 г.). «Заявление китайского исследователя о заявлении о первых детях с отредактированным геном» . Национальные институты здравоохранения (NIH) . Министерство здравоохранения и социальных служб США.
Дальнейшее чтение
[ редактировать ]- Уилкинсон Д. (сентябрь 1996 г.). «Кофакторы предоставляют ключи входа. ВИЧ-1» . Современная биология . 6 (9): 1051–1053. дои : 10.1016/S0960-9822(02)70661-1 . ПМИД 8805353 . S2CID 18710567 .
- Бродер С.С., Димитров Д.С. (1996). «ВИЧ и рецепторы 7-трансмембранного домена» . Патобиология . 64 (4): 171–179. дои : 10.1159/000164032 . ПМИД 9031325 .
- Чой Х., Мартин К.А., Фарзан М., Содроски Дж., Джерард Н.П., Джерард С. (июнь 1998 г.). «Структурные взаимодействия между хемокиновыми рецепторами, gp120 Env и CD4». Семинары по иммунологии . 10 (3): 249–257. дои : 10.1006/smim.1998.0127 . ПМИД 9653051 .
- Шеппард Х.В., Селум С., Майкл Н.Л., О'Брайен С., Дин М., Кэррингтон М. и др. (март 2002 г.). «Инфекция ВИЧ-1 у лиц с генотипом CCR5-Delta32/Delta32: приобретение синцитий-индуцирующего вируса при сероконверсии». Журнал синдромов приобретенного иммунодефицита . 29 (3): 307–313. дои : 10.1097/00042560-200203010-00013 . ПМИД 11873082 .
- Фридман Б.Д., Лю К.Х., Дель Корно М., Коллман Р.Г. (2003). «Передача сигналов, опосредованная рецептором хемокина gp120 ВИЧ-1, в макрофагах человека». Иммунологические исследования . 27 (2–3): 261–276. дои : 10.1385/IR:27:2-3:261 . ПМИД 12857973 . S2CID 32006625 .
- Эсте Ж.А. (сентябрь 2003 г.). «Проникновение вируса как цель вмешательства по борьбе с ВИЧ». Современная медицинская химия . 10 (17): 1617–1632. дои : 10.2174/0929867033457098 . ПМИД 12871111 .
- Галло С.А., Финнеган С.М., Виард М., Равив Ю., Димитров А., Рават С.С. и др. (июль 2003 г.). «Реакция слияния, опосредованная Env ВИЧ» . Biochimica et Biophysical Acta (BBA) – Биомембраны . 1614 (1): 36–50. дои : 10.1016/S0005-2736(03)00161-5 . ПМИД 12873764 .
- Зайцева М., Педен К., Голдинг Х. (июль 2003 г.). «Корецепторы ВИЧ: роль структуры, посттрансляционных модификаций и интернализации в слиянии вирусных клеток и в качестве мишеней для ингибиторов проникновения» . Biochimica et Biophysical Acta (BBA) – Биомембраны . 1614 (1): 51–61. дои : 10.1016/S0005-2736(03)00162-7 . ПМИД 12873765 .
- Ли С., Лю К.Х., Томкович Б., Йи Ю, Фридман Б.Д., Коллман Р.Г. (ноябрь 2003 г.). «Активация макрофагов через CCR5- и CXCR4-опосредованные сигнальные пути, вызываемые gp120» . Журнал биологии лейкоцитов . 74 (5): 676–682. дои : 10.1189/jlb.0503206 . ПМИД 12960231 . S2CID 11362623 .
- Йи Ю, Ли С., Лю К.Х., Фридман Б.Д., Коллман Р.Г. (2004). «Использование хемокиновых рецепторов и передача сигналов макрофагов gp120 вируса иммунодефицита человека типа 1: значение для нейропатогенеза». Журнал нейровирусологии . 10 (Приложение 1): 91–96. дои : 10.1080/753312758 . ПМИД 14982745 . S2CID 9065929 .
- Зайберт С., Сакмар Т.П. (2004). «Низкомолекулярные антагонисты CCR5 и CXCR4: новый многообещающий класс препаратов против ВИЧ-1». Текущий фармацевтический дизайн . 10 (17): 2041–2062. дои : 10.2174/1381612043384312 . ПМИД 15279544 .
- Катлер CW, Джотвани Р. (апрель 2006 г.). «Экспрессия рецепторов ВИЧ-1, корецепторов и альфа-дефензинов в слизистой оболочке полости рта: таблица устойчивости или восприимчивости к ВИЧ-инфекции?» . Достижения в области стоматологических исследований . 19 (1): 49–51. дои : 10.1177/154407370601900110 . ПМЦ 3750741 . ПМИД 16672549 .
- Ахуебор М.Н., Кэри Дж.А., Суэйн М.Г. (август 2006 г.). «CCR5 при заболеваниях печени, опосредованных Т-клетками: что происходит?» . Журнал иммунологии . 177 (4): 2039–2045. doi : 10.4049/jimmunol.177.4.2039 . ПМИД 16887960 .
- Липп М., Мюллер Г. (2003). «Формирование адаптивного иммунитета: влияние CCR7 и CXCR5 на трафик лимфоцитов». Переговоры Немецкого общества патологии . 87 :90-101. ПМИД 16888899 .
- Балистрери Ч.Р., Карузо С., Гримальди М.П., Листи Ф., Васто С., Орландо В. и др. (апрель 2007 г.). «Рецептор CCR5: биологические и генетические последствия возрастных заболеваний». Анналы Нью-Йоркской академии наук . 1100 (1): 162–172. Бибкод : 2007NYASA1100..162B . дои : 10.1196/анналы.1395.014 . ПМИД 17460174 . S2CID 8437349 .
- Мэдсен Х.О., Поулсен К., Даль О., Кларк Б.Ф., Хьёрт Дж.П. (март 1990 г.). «Ретропсевдогены составляют основную часть семейства генов человеческого фактора элонгации 1 альфа» . Исследования нуклеиновых кислот . 18 (6): 1513–1516. дои : 10.1093/нар/18.6.1513 . ПМК 330519 . ПМИД 2183196 .
- Уэцуки Т., Найто А., Нагата С., Казиро Ю. (апрель 1989 г.). «Выделение и характеристика хромосомного гена человека фактора элонгации полипептидной цепи-1 альфа» . Журнал биологической химии . 264 (10): 5791–5798. дои : 10.1016/S0021-9258(18)83619-5 . ПМИД 2564392 .
- Уайтхарт С.В., Шенбагамурти П., Чен Л., Коттер Р.Дж., Харт Г.В. (август 1989 г.). «Мышиный фактор элонгации 1 альфа (EF-1 альфа) посттрансляционно модифицируется новыми амидно-связанными фрагментами этаноламин-фосфоглицерина. Добавление этаноламин-фосфоглицерина к специфическим остаткам глутаминовой кислоты на EF-1 альфа» . Журнал биологической химии . 264 (24): 14334–14341. дои : 10.1016/S0021-9258(18)71682-7 . ПМИД 2569467 .
- Энн Д.К., Ву М.М., Хуан Т., Карлсон Д.М., Ву Р. (март 1988 г.). «Экспрессия генов, регулируемая ретинолом, в трахеобронхиальных эпителиальных клетках человека. Повышенная экспрессия фактора элонгации EF-1 альфа» . Журнал биологической химии . 263 (8): 3546–3549. дои : 10.1016/S0021-9258(18)68958-6 . ПМИД 3346208 .
- Брэндс Дж. Х., Массен Дж. А., ван Хемерт Ф. Дж., Амонс Р., Мёллер В. (февраль 1986 г.). «Первичная структура альфа-субъединицы человеческого фактора элонгации 1. Структурные аспекты сайтов связывания гуанин-нуклеотидов» . Европейский журнал биохимии . 155 (1): 167–171. дои : 10.1111/j.1432-1033.1986.tb09472.x . ПМИД 3512269 .
Внешние ссылки
[ редактировать ]- Видео и текст из документального фильма PBS об открытии CCR5
- «Хемокиновые рецепторы: CCR5» . База данных IUPHAR по рецепторам и ионным каналам . Международный союз фундаментальной и клинической фармакологии. Архивировано из оригинала 18 января 2021 года . Проверено 21 июля 2006 г.
- ВИЧcoPred Сервер для прогнозирования использования корецепторов ВИЧ (CCR5). PLoS ONE 8(4): e61437
- человека Местоположение генома CCR5 и CCR5 страница сведений о гене в браузере генома UCSC .
- Обзор всей структурной информации, доступной в PDB для UniProt : P51681 (CC-хемокиновый рецептор типа 5) на PDBe-KB .