Вирус клещевого энцефалита
| Вирус клещевого энцефалита | |
|---|---|
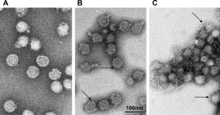
| |
| ВКЭ при различных уровнях pH | |
| Классификация вирусов | |
| (без рейтинга): | Вирус |
| Область : | Рибовирия |
| Королевство: | Орторнавиры |
| Тип: | Китриновирикота |
| Сорт: | Фласувирицеты |
| Заказ: | Желтовирусы |
| Семья: | Флавивирусиды |
| Род: | Флавивирус |
| Разновидность: | Вирус клещевого энцефалита
|
| Штаммы | |
| |
Вирус клещевого энцефалита ( ВКЭ ) представляет собой РНК-вирус с положительной цепью, связанный с клещевым энцефалитом, из рода Flavivirus .
Классификация
[ редактировать ]Таксономия
[ редактировать ]ВКЭ является представителем рода Flavivirus . К другим близким родственникам, членам серокомплекса ВКЭ, относятся вирус Омской геморрагической лихорадки , вирус Кясанурской лесной болезни , вирус Алхурма , вирус Лупинг-ил и вирус Лангат . [ 1 ]
Подтипы
[ редактировать ]ВКЭ имеет три подтипа:
- Западноевропейский подтип (ранее вирус центральноевропейского энцефалита, CEEV; основной переносчик клещей: Ixodes ricinus );
- Сибирский подтип (ранее Западно-Сибирский вирус; основной переносчик клещей: Ixodes persulcatus );
- Дальневосточный подтип (ранее вирус русского весенне-летнего энцефалита, RSSEV; основной переносчик клещей: Ixodes persulcatus ). [ 2 ]
Эталонным штаммом является штамм Sofjin. [ 3 ]
Вирусология
[ редактировать ]Структура
[ редактировать ]ВКЭ представляет собой с положительным смыслом одноцепочечный РНК-вирус , содержащийся в сферическом оболочке капсида размером 40–60 нм. [ 1 ] Геном ВКЭ имеет размер примерно 11 КБ, содержит 5'-кэп , одну открытую рамку считывания с 3'- и 5'- UTR и не содержит полиаденилирования . [ 1 ] Как и другие флавивирусы, [ 4 ] Геном ВКЭ кодирует десять вирусных белков, три структурных и семь неструктурных . [ 5 ] [ 1 ]
Структурными белками являются C ( капсид ), PrM (предмембранный), который расщепляется с образованием конечного мембранного белка (M) и белка оболочки (E). Семь неструктурных белков: NS1, NS2A, NS2B, NS3, NS4A, NS4B и NS5. Известна роль некоторых неструктурных белков, NS5 служит РНК-зависимой РНК-полимеразой , NS3 обладает протеазной (в комплексе с NS2B) и геликазной активностью. [ 5 ] [ 1 ] Структурные и неструктурные белки не требуются для того, чтобы геном был инфекционным. [ 1 ] Все вирусные белки экспрессируются в виде одного большого полипротеина порядка C, PrM, E, NS1, NS2A, NS2B, NS3, NS4A, NS4B, NS5. [ 5 ]
Вирусные генетические детерминанты патогенности
[ редактировать ]Белок оболочки участвует в связывании рецепторов и нейровирулентности, при этом повышенное сродство к связыванию гликозаминогликанов снижает нейроинвазивность. [ 6 ] На конформацию белка Е во время секреции вирусных частиц также влияет гликозилирование. [ 7 ] Была продемонстрирована иммуногенность NS1 ВКЭ, демонстрирующая его способность вызывать окислительный стресс и вызывать экспрессию субъединиц иммунопротеасом. Кроме того, было замечено, что он стимулирует выработку цитокинов. [ 8 ] Белок NS5 обладает антагонистической активностью интерферона , поскольку он подавляет экспрессию субъединицы рецептора IFN. Неструктурный белок 5 (NS5) влияет на нейропатогенез путем ослабления роста нейритов. Нетранслируемая область 3 (UTR3) и UTR 5 влияют на циклизацию и репликацию геномной РНК, а также на транспорт вирусной РНК в дендритах, что влияет на нейрогенез и синаптическую связь. [ 6 ]
Жизненный цикл
[ редактировать ]Передача инфекции
[ редактировать ]Заражение переносчиком начинается , когда клещ ест кровь инфицированного хозяина . Это может произойти на любом этапе жизненного цикла клеща, однако считается, что горизонтальная передача между инфицированными нимфами и неинфицированными личинками , питающимися одним и тем же хозяином, является ключевым фактором поддержания циркуляции ВКЭ. [ 9 ] [ 1 ] ВКЭ в крови хозяина заражает клеща через среднюю кишку , откуда он может попасть в слюнные железы и передаться следующему хозяину. У невзрослых клещей ВКЭ передается транстадиально путем заражения клеток, которые не разрушаются во время линьки , поэтому клещ остается заразным на протяжении всей своей жизни. [ 9 ] Зараженные взрослые клещи могут откладывать инфицированные яйца, передавая вирус трансовариально . [ 10 ]
Репликация
[ редактировать ]
У человека инфекция начинается с кожи (за исключением случаев пищевого происхождения, около 1% инфекций) в месте укуса инфицированного клеща, где преимущественно поражаются клетки Лангерганса и макрофаги кожи. [ 5 ] Белки оболочки (E) ВКЭ распознают гепарансульфат (и, вероятно, другие рецепторы) на поверхности клетки-хозяина и эндоцитозируются по клатрин -опосредованному пути . Подкисление поздней эндосомы вызывает конформационные изменения в белках Е, приводящие к слиянию с последующим снятием оболочки и высвобождением генома одноцепочечной РНК в цитоплазму. [ 11 ] [ 1 ]
Вирусный полипротеин транслируется и встраивается в мембрану ЭР , где он обрабатывается на цитозольной стороне пептидазами хозяина и в просвете под действием вирусных ферментов. Вирусные белки C, NS3 и NS5 расщепляются в цитозоль (хотя NS3 может образовывать комплекс с NS2B или NS4A для осуществления протеолитической или хеликазной активности), тогда как оставшиеся неструктурные белки изменяют структуру мембраны ЭР. Эта измененная мембрана позволяет собирать репликационные комплексы, в которых вирусный геном реплицируется вирусной РНК-зависимой РНК-полимеразой NS5. [ 11 ] [ 5 ]
Вновь реплицированные геномы вирусной РНК затем упаковываются белками C, находясь на цитозольной стороне мембраны ЭР, образуя незрелый нуклеокапсид , и получают белки E и PrM, организованные в виде гетеродимера, во время отпочкования в просвет ЭР. Незрелый вирион имеет остроконечную форму по сравнению со зрелой частицей. Частица проходит через аппарат Гольджи и транс -сеть Гольджи в условиях все более кислой среды, в результате чего вирион созревает с отщеплением сегмента Pr от белка М и образованием компетентных к слиянию гомодимеров белка Е. Однако расщепленный сегмент Pr остается связанным с белковым комплексом до выхода. [ 1 ] [ 11 ]
Вирус высвобождается из клетки-хозяина при слиянии транспортного пузырька с мембраной клетки-хозяина, расщепленные сегменты Pr теперь диссоциируют, в результате чего образуется полностью зрелый инфекционный вирус. [ 1 ] [ 11 ] Однако иногда также выделяются частично зрелые и незрелые вирусы; незрелые вирусы неинфекционны, поскольку белки Е не способны к слиянию, частично зрелые вирусы все еще способны заражать. [ 11 ]
Патогенез и иммунный ответ
[ редактировать ]
За исключением случаев пищевого заражения, заражение начинается с кожи в месте укуса клеща. клетки кожи (или клетки Лангерганса (ДК)). дендритные Предпочтительно нацелены на [ 5 ] Первоначально вирус реплицируется локально, и иммунный ответ запускается, когда вирусные компоненты распознаются цитозольными рецепторами распознавания образов (PRR), такими как Toll-подобные рецепторы (TLR). [ 12 ] Распознавание вызывает высвобождение цитокинов, включая интерфероны (IFN) α, β и γ , а также хемокины, привлекающие мигрирующие иммунные клетки к месту укуса. [ 5 ] На этом этапе инфекцию можно остановить и вылечить до появления заметных симптомов. Примечательно, что слюна клеща усиливает инфекцию, модулируя иммунный ответ хозяина и подавляя сигналы апоптоза. [ 12 ] Если инфекция продолжается, мигрирующие ДК и макрофаги заражаются и перемещаются в местный дренирующий лимфатический узел, где активируются -ядерные лейкоциты , моноциты и система комплемента . полиморфно [ 12 ]
Дренирующий лимфатический узел может также служить местом амплификации вируса, откуда ВКЭ получает системный доступ. Эта виремическая стадия соответствует первой симптоматической фазе прототипного двухфазного паттерна клещевого энцефалита. [ 1 ] ВКЭ отдает предпочтение нейрональной ткани и является нейроинвазивным. [ 13 ] Начальная стадия виремии обеспечивает доступ к ряду предпочтительных тканей. Однако точный механизм проникновения ВКЭ в центральную нервную систему (ЦНС) неясен. [ 13 ] [ 12 ] [ 1 ] Существует несколько предлагаемых механизмов нарушения гематоэнцефалического барьера (ГЭБ) ВКЭ: 1) Механизм «троянского коня», при котором ВКЭ получает доступ к ЦНС при инфицировании иммунных клеток, проходящих через ГЭБ; [ 12 ] [ 5 ] [ 13 ] 2) Нарушение и повышение проницаемости ГЭБ иммунными цитокинами; [ 13 ] 3) Путем заражения обонятельных нейронов; [ 5 ] 4) Путем ретроградного транспорта по периферическим нервам в ЦНС; [ 5 ] 5) Инфицирование клеток, входящих в состав ГЭБ. [ 5 ] [ 12 ]
Инфекция ЦНС вызывает вторую фазу классической двухфазной инфекции, связанной с европейским подтипом. Заболевание ЦНС является иммунопатологическим; высвобождение воспалительных цитокинов в сочетании с действием цитотоксических CD8+ Т-клеток и, возможно, NK-клеток приводит к воспалению и апоптозу инфицированных клеток, что отвечает за многие симптомы ЦНС. [ 12 ] [ 13 ]
Гуморальный ответ
[ редактировать ]В ответ на инфекцию вырабатываются специфические к ВКЭ антитела IgM и IgG. [ 1 ] Антитела IgM появляются и достигают пика первыми, а также достигают более высоких уровней и обычно исчезают примерно через 1,5 месяца после заражения, хотя существуют значительные различия от пациента к пациенту. Уровни IgG достигают пика примерно через 6 недель после появления симптомов со стороны ЦНС, затем слегка снижаются, но не исчезают, что, вероятно, обеспечивает пациенту пожизненный иммунитет. [ 1 ] [ 5 ]
Эволюция
[ редактировать ]Предок современных штаммов, по-видимому, разделился на несколько клад примерно 2750 лет назад. [ 14 ] Сибирский и подтипы разошлись дальневосточный около 2250 лет назад. Второй анализ предполагает более раннюю дату эволюции (3300 лет назад) с быстрым увеличением числа штаммов, начавшимся примерно 300 лет назад. [ 15 ] Различные штаммы вируса передавались в Японию как минимум трижды в период 260–430 лет назад. [ 16 ] [ 17 ] Штаммы, циркулирующие в Латвии , по-видимому, произошли как из России, так и из Западной Европы. [ 18 ] в то время как те, что находятся в Эстонии, судя по всему, произошли из России . [ 19 ] Литовские штаммы, по-видимому , родственны штаммам из Западной Европы. [ 20 ] Филогенетический анализ показывает, что европейский и сибирский подтипы ВКЭ тесно связаны, а дальневосточный подтип ближе к вирусу Louping Ill. [ 1 ] Однако по антигенному родству, основанному на белках E, NS3 и NS5, все три подтипа очень схожи, и вирус Louping III является ближайшим родственником за пределами коллективной группы ВКЭ. [ 21 ]
История
[ редактировать ]Хотя первое описание того, что могло быть TBE, появляется в записях 1700-х годов в Скандинавии. [ 13 ] идентификация вируса ВКЭ произошла в Советском Союзе в 1930-х годах. [ 22 ] Расследование началось в связи со вспышкой, как предполагалось, японского энцефалита («Летнего энцефалита») среди советских войск, дислоцированных вдоль границы с Японской империей (нынешней Китайской Народной Республикой ), недалеко от дальневосточного города Хабаровска . Экспедицию возглавил вирусолог Лев А. Зильбер , собравший команду из двадцати молодых ученых в ряде смежных областей, таких как акарология , микробиология, неврология и эпидемиология. [ 23 ] [ 22 ] Экспедиция прибыла в Хабаровск 15 мая 1937 года и разделилась на отряды: Северный под руководством Елизаветы Николаевны Левкович, работавший в Хабаровском крае , и Южный под руководством Александры Дмитриевны Шеболдаевой, работавший в Приморском крае . [ 22 ]
В мае экспедиция определила клещей как вероятный переносчик, собрала клещей I. persucatus путем воздействия на голую кожу энтомологом Александром В. Гуцевичем, а вирусолог Михаил П. Чумаков изолировал вирус от клещей, питающихся намеренно зараженными мышами. Летом пять участников экспедиции заразились ВКЭ, и, хотя смертельных исходов не было, трое из пяти получили тяжелые последствия . [ 22 ]
Экспедиция вернулась в середине августа, а в октябре 1937 года Зильбер и Шеболдова были арестованы по ложному обвинению в распространении японского энцефалита. Эпидемиолог экспедиции Тамара М. Сафонова была арестована в январе следующего года за протест против обвинений против Зильбера и Шеболдовой. В результате арестов была опубликована одна из важных первоначальных работ под авторством экспедиционного акаролога Василия Сергеевича Миронова. Зильбер вышел на свободу в 1939 году и сумел восстановить вместе с Шеболдовой соавторство в этом первоначальном произведении; однако Сафанов и Шеболдова (которую не освободили) провели 18 лет в трудовых лагерях . [ 22 ] [ 23 ]
Ссылки
[ редактировать ]- ^ Jump up to: а б с д и ж г час я дж к л м н тот Мэнсфилд К.Л., Джонсон Н., Фиппс Л.П., Стивенсон-младший, Фукс А.Р., Соломон Т. (август 2009 г.). «Вирус клещевого энцефалита – обзор возникающего зооноза» . Журнал общей вирусологии . 90 (Часть 8): 1781–1794. дои : 10.1099/vir.0.011437-0 . ПМИД 19420159 .
- ^ Гудман Дж.Л., Деннис Д.Т., Соненшайн Д.Э. (2005). «Клещевой энцефалит». Клещевые болезни человека . Вашингтон, округ Колумбия: ASM Press. п. 151. ИСБН 978-1-55581-238-6 .
- ^ Ковалев С.Ю., Мухачева Т.А., Кокорев В.С., Беляева И.В. (апрель 2012 г.). «Вирус клещевого энцефалита: эталонный штамм Софджин и проблема его подлинности». Гены вирусов . 44 (2): 217–224. дои : 10.1007/s11262-011-0690-9 . ПМИД 22095094 . S2CID 12587373 .
- ^ Уайлдер-Смит А., Оой Э.Э., Хорстик О., Уиллс Б. (январь 2019 г.). «Денге». Ланцет . 393 (10169): 350–363. дои : 10.1016/s0140-6736(18)32560-1 . ПМИД 30696575 . S2CID 208789595 .
- ^ Jump up to: а б с д и ж г час я дж к л Рузек Д., Авшич Жупанц Т., Борде Дж., Чрдле А., Эйер Л., Карганова Г. и др. (апрель 2019 г.). «Клещевой энцефалит в Европе и России: обзор патогенеза, клиники, терапии и вакцин». Противовирусные исследования . 164 : 23–51. doi : 10.1016/j.antiviral.2019.01.014 . ПМИД 30710567 . S2CID 73414822 .
- ^ Jump up to: а б Ёсии К. (март 2019 г.). «Эпидемиология и патологические механизмы клещевого энцефалита» . Журнал ветеринарной медицины . 81 (3): 343–347. дои : 10.1292/jvms.18-0373 . ПМК 6451894 . PMID 30674746 .
- ^ Ёшии К., Янагихара Н., Исидзука М., Сакаи М., Карива Х. (октябрь 2013 г.). «N-связанный гликан в белке оболочки вируса клещевого энцефалита влияет на секрецию вируса в клетках млекопитающих, но не в клетках клещей» . Журнал общей вирусологии . 94 (Часть 10): 2249–2258. дои : 10.1099/vir.0.055269-0 . hdl : 2115/56524 . ПМИД 23824303 .
- ^ Стародубова Е, Тучинская К, Кузьменко Ю, Латанова А, Тутяева В, Карпов В, Карганова Г (январь 2023 г.). «Активация ранних провоспалительных реакций вирусом NS1 варьируется у штаммов различных подтипов» . Международный журнал молекулярных наук . 24 (2): 1011. doi : 10.3390/ijms24021011 . ПМЦ 9863113 . ПМИД 36674524 .
- ^ Jump up to: а б Наттолл П.А., Джонс Л.Д., Лабуда М., Кауфман В.Р. (январь 1994 г.). «Адаптации арбовирусов к клещам». Журнал медицинской энтомологии . 31 (1): 1–9. дои : 10.7939/r3m03xx77 . ПМИД 8158611 .
- ^ Даниэлова В, Голубова Ю, Пейкох М, Дэниел М (2002). «Возможное значение трансовариальной передачи в циркуляции вируса клещевого энцефалита» . Фолиа Паразитологическая . 49 (4): 323–325. дои : 10.14411/fp.2002.060 . ПМИД 12641208 .
- ^ Jump up to: а б с д и Пулккинен Л.И., Бутчер С.Дж., Анастасина М. (июнь 2018 г.). «Вирус клещевого энцефалита: структурный взгляд» . Вирусы . 10 (7): 350. дои : 10.3390/v10070350 . ПМК 6071267 . ПМИД 29958443 .
- ^ Jump up to: а б с д и ж г Велай А., Пас М., Сесброн М., Гантнер П., Солис М., Сулье Э. и др. (август 2019 г.). «Вирус клещевого энцефалита: молекулярные детерминанты нейропатогенеза нового возбудителя». Критические обзоры по микробиологии . 45 (4): 472–493. дои : 10.1080/1040841X.2019.1629872 . ПМИД 31267816 . S2CID 195787988 .
- ^ Jump up to: а б с д и ж Блом К., Куапио А., Сандберг Дж.Т., Варнайте Р., Михаэльссон Дж., Бьоркстрем Н.К. и др. (2018). «Клеточно-опосредованные иммунные реакции и иммунопатогенез вирусной инфекции клещевого энцефалита человека» . Границы в иммунологии . 9 : 2174. дои : 10.3389/fimmu.2018.02174 . ПМК 6168641 . ПМИД 30319632 .
- ^ Субботина Е.Л., Локтев В.Б. (2012). «[Молекулярная эволюция клещевого энцефалита и вирусов Повассан]». Молекулярная биология . 46 (1): 82–92. дои : 10.1134/S0026893311060148 . ПМИД 22642104 . S2CID 18500235 .
- ^ Узкатеги Н.Ю., Сиронен Т., Головлева И., Яскеляйнен А.Е., Валимаа Х., Лундквист А. и др. (апрель 2012 г.). «Скорость эволюции и молекулярная эпидемиология вируса клещевого энцефалита в Европе, включая два выделения из одного и того же очага с разницей в 44 года» . Журнал общей вирусологии . 93 (Часть 4): 786–796. дои : 10.1099/vir.0.035766-0 . ПМИД 22205716 .
- ^ Сузуки Y (июнь 2007 г.). «Множественные передачи вируса клещевого энцефалита между Японией и Россией» . Гены и генетические системы . 82 (3): 187–195. дои : 10.1266/ggs.82.187 . ПМИД 17660689 .
- ^ Такашима И., Хаясака Д., Гото А., Карива Х., Мизутани Т. (февраль 2001 г.). «Эпидемиология клещевого энцефалита (КЭ) и филогенетический анализ вирусов КЭ в Японии и на Дальнем Востоке России». Японский журнал инфекционных заболеваний . 54 (1): 1–11. ПМИД 11326122 .
- ^ Вене С., Головлева И., Мавчутко В., Форсгрен М., Калнина В., Плюснин А. (декабрь 2001 г.). «Характеристика вируса клещевого энцефалита из Латвии: данные о совместной циркуляции трех различных подтипов». Журнал медицинской вирусологии . 65 (4): 730–735. дои : 10.1002/jmv.2097 . ПМИД 11745938 . S2CID 22860154 .
- ^ Головлёва И., Вене С., Шёландер К.Б., Василенко В., Плюснин А., Лундквист А. (декабрь 2004 г.). «Характеристика вируса клещевого энцефалита из Эстонии». Журнал медицинской вирусологии . 74 (4): 580–588. дои : 10.1002/jmv.20224 . ПМИД 15484275 . S2CID 28491834 .
- ^ Микьене А, Вене С, Головлева И, Лайсконис А, Линдквист Л, Плюснин А, Лундквист А (декабрь 2001 г.). «Вирус клещевого энцефалита в Литве». Европейский журнал клинической микробиологии и инфекционных заболеваний . 20 (12): 886–888. дои : 10.1007/s10096-001-0637-5 . ПМИД 11837641 . S2CID 38061544 .
- ^ Хайнц FX, Стиасны К (2019). «Глава 2б: Молекулярная и антигенная структура ВКЭ» . Клещевой энцефалит - Книга . дои : 10.33442/978-981-14-0914-1_2b . ISSN 2661-3980 . S2CID 150118109 .
- ^ Jump up to: а б с д и Злобин В.И., Погодина В.В., Каль О. (октябрь 2017 г.). «Краткая история открытия вируса клещевого энцефалита в конце 1930-х годов (по воспоминаниям участников экспедиций, их коллег и родственников)». Клещи и клещевые заболевания . 8 (6): 813–820. дои : 10.1016/j.ttbdis.2017.05.001 . ПМИД 28526419 .
- ^ Jump up to: а б Успенский I (май 2018 г.). «Несколько слов в дополнение к «Краткой истории открытия вируса клещевого энцефалита в конце 1930-х годов» В.И. Злобина, В.В. Погодиной и О. Каль (TTBDIS, 2017, 8, 813-820)». Клещи и клещевые заболевания . 9 (4): 834–835. дои : 10.1016/j.ttbdis.2018.03.007 . ПМИД 29559213 .
Внешние ссылки
[ редактировать ]- Энцефалит+вирусы+клещевой в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)
- Клещевой энцефалит во Всемирной организации здравоохранения
- Книга TBE, 4-е издание, Герхард Доблер, Вильгельм Эрбер, Михаэль Брокер, Хайнц-Йозеф Шмитт, Global Health Press, 25 мая 2021 г. - стр. 386 стр.
| Базы данных органов управления : Национальные |
|---|