мТОР
| МТОР | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Идентификаторы | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | MTOR , FRAP, FRAP1, FRAP2, RAFT1, RAPT1, SKS, механистическая мишень рапамицина, механистическая мишень киназы рапамицина | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | ОМИМ : 601231 ; МГИ : 1928394 ; Гомологен : 3637 ; Генные карты : MTOR ; ОМА : МТОР – ортологи | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||




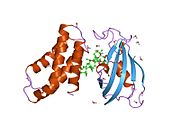
Мишень для млекопитающих рапамицина ( mTOR ), [5] Также называемый механистической мишенью рапамицина и иногда называемый FK506-связывающим белком. 12-рапамицин-ассоциированный белок 1 (FRAP1) представляет собой киназу , которая у людей кодируется MTOR геном . [6] [7] [8] mTOR является членом родственных фосфатидилинозитол-3-киназным киназам семейства протеинкиназ, . [9]
mTOR связывается с другими белками и служит основным компонентом двух отдельных белковых комплексов : комплекса mTOR 1 и комплекса mTOR 2 , которые регулируют различные клеточные процессы. [10] В частности, будучи основным компонентом обоих комплексов, mTOR действует как серин/треониновая протеинкиназа , которая регулирует рост клеток, пролиферацию клеток , подвижность клеток , выживаемость клеток, синтез белка , аутофагию и транскрипцию . [10] [11] Являясь основным компонентом mTORC2, mTOR также действует как тирозиновая протеинкиназа , которая способствует активации рецепторов инсулина и рецепторов инсулиноподобного фактора роста 1 . [12] mTORC2 также участвует в контроле и поддержании актинового цитоскелета . [10] [13]
Открытие
[ редактировать ]Рапа-Нуи (остров Пасхи - Чили)
[ редактировать ]Исследование TOR началось в 1960-х годах во время экспедиции на остров Пасхи (известный жителям острова как Рапа-Нуи ) с целью выявления натуральных продуктов из растений и почвы с возможным терапевтическим потенциалом. В 1972 году Сурен Сегал идентифицировал небольшую молекулу почвенной бактерии Streptomyces hygroscopeus , которую он очистил и первоначально сообщил, что она обладает мощной противогрибковой активностью. Он назвал его рапамицином , отметив его первоначальный источник и активность. [14] [15] Ранние испытания показали, что рапамицин также обладает мощной иммуносупрессивной и цитостатической противораковой активностью. Первоначально рапамицин не вызывал значительного интереса со стороны фармацевтической промышленности до 1980-х годов, когда Уайет-Айерст поддержал усилия Сегала по дальнейшему исследованию влияния рапамицина на иммунную систему. В конечном итоге это привело к одобрению FDA в качестве иммунодепрессанта после трансплантации почки. Однако до одобрения FDA механизм действия рапамицина оставался совершенно неизвестным.
Последующая история
[ редактировать ]Открытие TOR и mTOR произошло в результате независимых исследований природного продукта рапамицина, проведенных Джозефом Хейтманом , Рао Моввой и Майклом Н. Холлом в 1991 году; [16] Дэвид М. Сабатини , Хедие Эрджюмент-Бромадж, Мэри Луи, Пол Темпст и Соломон Х. Снайдер в 1994 году; [7] и Кэндис Дж. Сэйберс, Мэри М. Мартин, Грегори Дж. Брунн, Джози М. Уильямс, Фрэнсис Дж. Дюмон, Грегори Видеррехт и Роберт Т. Абрахам в 1995 году. [8] В 1991 году, работая с дрожжами, Холл и его коллеги идентифицировали гены TOR1 и TOR2. [16] В 1993 году Роберт Кафферки, Джордж Ливи и коллеги, а также Жаннет Кунц, Майкл Н. Холл и коллеги независимо клонировали гены, которые опосредуют токсичность рапамицина в грибах, известные как гены TOR/DRR. [17] [18]
Рапамицин подавляет активность грибов на фазе G1 клеточного цикла. У млекопитающих он подавляет иммунную систему, блокируя фазовый переход G1 в S в Т-лимфоцитах . [19] Таким образом, он используется в качестве иммунодепрессанта после трансплантации органов. [20] Интерес к рапамицину возобновился после открытия в 1987 году структурно родственного иммуносупрессивного природного продукта FK506 (позже названного такролимусом). В 1989–90 годах было установлено, что FK506 и рапамицин ингибируют рецептора Т-клеток (TCR) и рецептора IL-2. сигнальные пути , соответственно. [21] [22] Два натуральных продукта были использованы для открытия белков, связывающих FK506 и рапамицин , включая FKBP12 , а также для доказательства того, что FKBP12-FK506 и FKBP12-рапамицин могут действовать посредством механизмов усиления функции, нацеленных на различные клеточные функции. Эти исследования включали ключевые исследования Фрэнсиса Дюмона и Нолана Сигала из компании Merck, которые показали, что FK506 и рапамицин действуют как взаимные антагонисты. [23] [24] Эти исследования предполагали, что FKBP12 является возможной мишенью рапамицина, но предположили, что комплекс может взаимодействовать с другим элементом механистического каскада. [25] [26]
В 1991 году кальциневрин был идентифицирован как мишень FKBP12-FK506. [27] Характер FKBP12-рапамицина оставался загадкой до тех пор, пока генетические и молекулярные исследования на дрожжах не установили FKBP12 как мишень рапамицина и не выявили TOR1 и TOR2 как мишени FKBP12-рапамицина в 1991 и 1993 годах. [16] [28] за которыми последовали исследования в 1994 году, когда несколько групп, работая независимо, обнаружили киназу mTOR как свою прямую мишень в тканях млекопитающих. [6] [7] [20] Анализ последовательности mTOR показал, что он является прямым ортологом белков, кодируемых дрожжевыми мишенями генов рапамицина 1 и 2 (TOR1 и TOR2 ), которые Джозеф Хейтман, Рао Мовва и Майкл Н. Холл идентифицировали в августе 1991 г. и мае 1993 г. Независимо от этого Джордж Ливи и его коллеги позже сообщили об тех же генах, которые они назвали доминантной устойчивостью к рапамицину 1 и 2 (DRR1 и DRR2) , в исследованиях, опубликованных в октябре 1993 года.
Белок, который теперь называется mTOR, первоначально был назван FRAP Стюартом Л. Шрайбером и RAFT1 Дэвидом М. Сабатини; [6] [7] FRAP1 использовался в качестве официального символа гена у людей. Из-за разных названий mTOR, который впервые использовал Роберт Т. Абрахам, [6] все чаще принимается сообществом ученых, работающих над путем mTOR, для обозначения белка и в знак уважения к первоначальному открытию белка TOR в дрожжах, который Джо Хейтман, Рао Мовва и Майк назвали TOR, «мишень рапамицина». Зал. TOR был первоначально обнаружен в Biozentrum и Sandoz Pharmaceuticals в 1991 году в Базеле, Швейцария, и название TOR отдает дань уважения этому открытию, поскольку TOR по-немецки означает «дверь» или «ворота», а город Базель когда-то был окружен стеной, перемежающейся ворота в город, в том числе знаменитый Шпалентор . [29] «mTOR» первоначально означало «мишень рапамицина для млекопитающих», но позже значение «m» было изменено на «механистический». [30] Аналогичным образом, с последующими открытиями рыба-зебра TOR была названа zTOR, TOR Arabidopsis thaliana получила название AtTOR, а TOR дрозофилы получила название dTOR. (HGNC) официально изменил название гена FRAP1 В 2009 году Комитет по номенклатуре генов HUGO на mTOR, что означает «механистическая мишень рапамицина». [31]
Открытие TOR и последующая идентификация mTOR открыли двери для молекулярных и физиологических исследований того, что сейчас называется путем mTOR, и оказали каталитический эффект на развитие области химической биологии, где небольшие молекулы используются в качестве зондов. биология.
Функция
[ редактировать ]mTOR интегрирует входные данные от вышестоящих путей , включая инсулин , факторы роста (такие как IGF-1 и IGF-2 ) и аминокислоты . [11] mTOR также определяет уровень питательных веществ, кислорода и энергии в клетках. [32] Путь mTOR является центральным регулятором метаболизма и физиологии млекопитающих, играя важную роль в функционировании тканей, включая печень, мышцы, белую и бурую жировую ткань. [33] и мозг, а его регуляция нарушается при таких заболеваниях человека, как диабет , ожирение , депрессия и некоторые виды рака . [34] [35] Рапамицин ингибирует mTOR, связываясь с его внутриклеточным рецептором FKBP 12. [36] [37] Комплекс FKBP12- рапамицин напрямую связывается с доменом FKBP12-рапамицин-связывающий (FRB) mTOR, ингибируя его активность. [37]
В растениях
[ редактировать ]Растения экспрессируют механистическую мишень рапамицина (mTOR) и имеют киназный комплекс TOR. В растениях присутствует только комплекс TORC1, в отличие от мишени рапамицина у млекопитающих, которая также содержит комплекс TORC2. [38] Виды растений имеют белки TOR в доменах протеинкиназы и FKBP-рапамицинсвязывающего домена (FRB), которые имеют аминокислотную последовательность, аналогичную mTOR у млекопитающих. [39]
Роль mTOR в растениях
Известно, что киназный комплекс TOR играет роль в метаболизме растений. Комплекс TORC1 включается, когда растения живут в подходящих для выживания условиях окружающей среды. После активации растительные клетки подвергаются особым анаболическим реакциям. К ним относятся развитие растений, трансляция мРНК и рост клеток внутри растения. Однако активация комплекса TORC1 останавливает возникновение катаболических процессов, таких как аутофагия. [38] Было обнаружено, что передача сигналов TOR-киназы в растениях способствует старению, цветению, росту корней и листьев, эмбриогенезу и активации меристемы над корневым чехликом растения. [40] Также обнаружено, что mTOR активно участвует в развитии эмбриональной ткани растений. [39]
Комплексы
[ редактировать ]
mTOR является каталитической субъединицей двух структурно различных комплексов: mTORC1 и mTORC2. [41] Два комплекса локализуются в разных субклеточных компартментах, тем самым влияя на их активацию и функцию. [42] После активации Rheb mTORC1 локализуется в комплексе Ragulator-Rag на поверхности лизосомы, где затем становится активным в присутствии достаточного количества аминокислот. [43] [44]
mTORC1
[ редактировать ]Комплекс mTOR 1 (mTORC1) состоит из mTOR, регуляторно-ассоциированного белка mTOR ( Raptor ), летального для млекопитающих с белком 8 SEC13 ( mLST8 ), и неосновных компонентов PRAS40 и DEPTOR . [45] [46] Этот комплекс действует как сенсор питательных веществ/энергии/окислительно-восстановительного потенциала и контролирует синтез белка. [11] [45] Активность mTORC1 регулируется рапамицином , инсулином, факторами роста, фосфатидной кислотой , некоторыми аминокислотами и их производными (например, L -лейцином и β-гидрокси-β-метилмасляной кислотой ), механическими стимулами и окислительным стрессом . [45] [47] [48]
mTORC2
[ редактировать ]Комплекс mTOR 2 (mTORC2) состоит из MTOR, нечувствительного к рапамицину компаньона MTOR ( RICTOR ), MLST8 и белка 1, взаимодействующего со стресс-активируемой протеинкиназой млекопитающих ( mSIN1 ). [49] [50] Было показано, что mTORC2 функционирует как важный регулятор актинового цитоскелета посредством стимуляции F- актиновых стрессовых волокон, паксиллина , RhoA , Rac1 , Cdc42 и протеинкиназы Cα ( PKCα ). [50] mTORC2 также фосфорилирует серин/треониновую протеинкиназу Akt/PKB по остатку серина Ser473, тем самым влияя на метаболизм и выживаемость. [51] Фосфорилирование серинового остатка Ser473 Akt с помощью mTORC2 стимулирует фосфорилирование Akt по остатку треонина Thr308 с помощью PDK1 и приводит к полной активации Akt. [52] [53] Кроме того, mTORC2 проявляет тирозиновую протеинкиназную активность и фосфорилирует рецептор инсулиноподобного фактора роста 1 (IGF-1R) и рецептор инсулина (InsR) по остаткам тирозина Tyr1131/1136 и Tyr1146/1151 соответственно, что приводит к полной активации IGF. -ИР и ИнР. [12]
Ингибирование рапамицином
[ редактировать ]Рапамицин ( сиролимус ) ингибирует mTORC1, что приводит к подавлению клеточного старения . [54] Это, по-видимому, обеспечивает большую часть полезных эффектов препарата (включая увеличение продолжительности жизни в исследованиях на животных). Подавление резистентности к инсулину сиртуинами объясняет , по крайней мере, часть этого эффекта. [55] Нарушение сиртуина 3 приводит к митохондриальной дисфункции . [56]
Рапамицин оказывает более сложное воздействие на mTORC2, ингибируя его только в определенных типах клеток при длительном воздействии. Нарушение mTORC2 вызывает диабетические симптомы снижения толерантности к глюкозе и нечувствительности к инсулину. [57]
Эксперименты по удалению генов
[ редактировать ]Сигнальный путь mTORC2 менее определен, чем сигнальный путь mTORC1. Функции компонентов комплексов mTORC были изучены с использованием нокдаунов и нокаутов и обнаружено наличие следующих фенотипов:
- NIP7 : Нокдаун снижает активность mTORC2, на что указывает снижение фосфорилирования субстратов mTORC2. [58]
- РИКТОР : Сверхэкспрессия приводит к метастазированию, а нокдаун ингибирует индуцированное фактором роста фосфорилирование PKC. [59] Конститутивная делеция Rictor у мышей приводит к эмбриональной смертности. [60] в то время как тканеспецифическая делеция приводит к множеству фенотипов; Распространенным фенотипом делеции Rictor в печени, белой жировой ткани и бета-клетках поджелудочной железы является системная непереносимость глюкозы и резистентность к инсулину в одной или нескольких тканях. [57] [61] [62] [63] Снижение экспрессии Rictor у мышей снижает продолжительность жизни самцов, но не самок. [64]
- mTOR: Ингибирование mTORC1 и mTORC2 с помощью PP242 [2-(4-амино-1-изопропил-1H-пиразоло[3,4-d]пиримидин-3-ил)-1H-индол-5-ол] приводит к аутофагии или апоптоз ; ингибирование только mTORC2 с помощью PP242 предотвращает фосфорилирование сайта Ser-473 на AKT и арестовывает клетки в фазе G1 клеточного цикла . [65] Генетическое снижение экспрессии mTOR у мышей значительно увеличивает продолжительность жизни. [66]
- PDK1 : Нокаут смертелен; гипоморфный аллель приводит к меньшему объему органа и размеру организма, но нормальной активации АКТ. [67]
- АКТ : Нокаутные мыши испытывают спонтанный апоптоз ( АКТ1 ), тяжелый диабет ( АКТ2 ), маленький мозг ( АКТ3 ) и задержку роста (АКТ1/АКТ2). [68] Мыши, гетерозиготные по AKT1, имеют увеличенную продолжительность жизни. [69]
- TOR1, ортолог mTORC1 S. cerevisiae , является регулятором как углеродного, так и азотистого метаболизма; Штаммы TOR1 KO регулируют реакцию на азот, а также на доступность углерода, что указывает на то, что он является ключевым преобразователем питания у дрожжей. [70] [71]
Клиническое значение
[ редактировать ]Старение
[ редактировать ]
Было обнаружено, что снижение активности TOR увеличивает продолжительность жизни cerevisiae , C. elegans и S. D. melanogaster . [72] [73] [74] [75] Было подтверждено, что ингибитор mTOR рапамицин увеличивает продолжительность жизни мышей. [76] [77] [78] [79] [80]
Предполагается, что некоторые диетические режимы, такие как ограничение калорий и ограничение метионина , вызывают увеличение продолжительности жизни за счет снижения активности mTOR. [72] [73] Некоторые исследования показали, что передача сигналов mTOR может усиливаться с возрастом, по крайней мере, в определенных тканях, таких как жировая ткань, и рапамицин может частично блокировать это увеличение. [81] Альтернативная теория заключается в том, что передача сигналов mTOR является примером антагонистической плейотропии , и хотя высокая передача сигналов mTOR полезна в раннем возрасте, она сохраняется на недопустимо высоком уровне в пожилом возрасте. Ограничение калорий и ограничение метионина могут частично действовать за счет ограничения уровней незаменимых аминокислот , включая лейцин и метионин, которые являются мощными активаторами mTOR. [82] введение лейцина Было показано, что в мозг крыс снижает потребление пищи и массу тела за счет активации пути mTOR в гипоталамусе . [83]
Согласно свободнорадикальной теории старения , [84] активные формы кислорода вызывают повреждение митохондриальных белков и снижают выработку АТФ. Впоследствии, через АТФ-чувствительную AMPK , путь mTOR ингибируется, а синтез белка, потребляющего АТФ, подавляется, поскольку mTORC1 инициирует каскад фосфорилирования, активирующий рибосому . [19] Следовательно, доля поврежденных белков увеличивается. Более того, нарушение mTORC1 напрямую ингибирует митохондриальное дыхание . [85] Этим положительным реакциям на процесс старения противодействуют защитные механизмы: снижение активности mTOR (среди других факторов) усиливает удаление дисфункциональных клеточных компонентов посредством аутофагии . [84]
mTOR является ключевым инициатором секреторного фенотипа, связанного со старением (SASP). [86] Интерлейкин 1 альфа (IL1A) обнаруживается на поверхности стареющих клеток , где он способствует выработке факторов SASP благодаря петле положительной обратной связи с NF-κB. [87] [88] Трансляция мРНК IL1A сильно зависит от активности mTOR. [89] Активность mTOR повышает уровень IL1A, опосредованно MAPKAPK2 . [87] Ингибирование mTOR ZFP36L1 предотвращает деградацию этим белком транскриптов многочисленных компонентов факторов SASP. [90]
Рак
[ редактировать ]Чрезмерная активация передачи сигналов mTOR в значительной степени способствует инициированию и развитию опухолей, и было обнаружено, что активность mTOR нарушается при многих типах рака, включая рак молочной железы, простаты, легких, меланому, мочевого пузыря, головного мозга и почки. [91] Причин конститутивной активации несколько. Среди наиболее распространенных — мутации гена-супрессора опухолей PTEN . Фосфатаза PTEN отрицательно влияет на передачу сигналов mTOR, препятствуя эффекту PI3K , вышестоящего эффектора mTOR. Кроме того, активность mTOR дерегулируется при многих видах рака в результате повышенной активности PI3K или Akt . [92] Аналогичным образом, сверхэкспрессия нижестоящих эффекторов mTOR 4E-BP1 , S6K1 , S6K2 и eIF4E приводит к плохому прогнозу рака. [93] Кроме того, мутации в белках TSC , которые ингибируют активность mTOR, могут привести к состоянию, называемому комплексом туберозного склероза , которое проявляется в виде доброкачественных поражений и увеличивает риск почечно-клеточного рака . [94]
Было показано, что повышение активности mTOR стимулирует прогрессирование клеточного цикла и увеличивает пролиферацию клеток, главным образом, за счет его влияния на синтез белка. Более того, активный mTOR поддерживает рост опухоли также косвенно, подавляя аутофагию . [95] Конститутивно активированная функция mTOR обеспечивает снабжение клеток карциномы кислородом и питательными веществами за счет увеличения трансляции HIF1A и поддержки ангиогенеза . [96] mTOR также способствует другой метаболической адаптации раковых клеток для поддержания их повышенной скорости роста — активации гликолитического метаболизма . Akt2 , субстрат mTOR, в частности mTORC2 , усиливает экспрессию гликолитического фермента PKM2, тем самым способствуя эффекту Варбурга . [97]
Нарушения центральной нервной системы/Функция мозга
[ редактировать ]Аутизм
[ редактировать ]mTOR участвует в отказе механизма «обрезки» возбуждающих синапсов при расстройствах аутистического спектра . [98]
болезнь Альцгеймера
[ редактировать ]Передача сигналов mTOR пересекается с патологией болезни Альцгеймера (БА) в нескольких аспектах, что указывает на ее потенциальную роль как фактора, способствующего прогрессированию заболевания. В целом результаты демонстрируют гиперактивность передачи сигналов mTOR в мозге с болезнью Альцгеймера. Например, посмертные исследования мозга человека с болезнью Альцгеймера выявили нарушение регуляции PTEN, Akt, S6K и mTOR. [99] [100] [101] Передача сигналов mTOR, по-видимому, тесно связана с наличием растворимого бета-амилоида (Aβ) и тау-белков, которые агрегируют и образуют два признака заболевания: бляшки Aβ и нейрофибриллярные клубки соответственно. [102] Исследования in vitro показали, что Aβ является активатором пути PI3K/AKT , который, в свою очередь, активирует mTOR. [103] Кроме того, применение Aβ к клеткам N2K увеличивает экспрессию p70S6K, нижестоящей мишени mTOR, которая, как известно, имеет более высокую экспрессию в нейронах, которые в конечном итоге развивают нейрофибриллярные клубки. [104] [105] Клетки яичника китайского хомячка, трансфицированные семейной мутацией AD 7PA2, также демонстрируют повышенную активность mTOR по сравнению с контролем, а гиперактивность блокируется с помощью ингибитора гамма-секретазы. [106] [107] Эти исследования in vitro предполагают, что увеличение концентрации Aβ увеличивает передачу сигналов mTOR; однако считается, что значительно большие цитотоксические концентрации Aβ снижают передачу сигналов mTOR. [108]
В соответствии с данными, полученными in vitro, было показано, что активность mTOR и активированный p70S6K значительно повышаются в коре головного мозга и гиппокампе животных моделей AD по сравнению с контрольной группой. [107] [109] Фармакологическое или генетическое удаление Aβ в животных моделях AD устраняет нарушение нормальной активности mTOR, указывая на прямое участие Aβ в передаче сигналов mTOR. [109] Кроме того, при введении олигомеров Aβ в гиппокамп нормальным мышам наблюдается гиперактивность mTOR. [109] Когнитивные нарушения, характерные для БА, по-видимому, опосредованы фосфорилированием PRAS-40, которое отделяется от mTOR и обеспечивает гиперактивность mTOR при его фосфорилировании; ингибирование фосфорилирования PRAS-40 предотвращает Aβ-индуцированную гиперактивность mTOR. [109] [110] [111] Учитывая эти данные, сигнальный путь mTOR, по-видимому, является одним из механизмов индуцированной Aβ токсичности при БА.
Гиперфосфорилирование тау-белков в нейрофибриллярные клубки является одним из признаков БА. Было показано, что активация p70S6K способствует образованию клубков, а также гиперактивности mTOR за счет увеличения фосфорилирования и снижения дефосфорилирования. [104] [112] [113] [114] Также было высказано предположение, что mTOR способствует развитию тау-патологии за счет увеличения трансляции тау и других белков. [115]
Синаптическая пластичность играет ключевую роль в обучении и памяти — двух процессах, которые серьезно нарушены у пациентов с болезнью Альцгеймера. Было показано, что трансляционный контроль, или поддержание белкового гомеостаза, важен для пластичности нейронов и регулируется mTOR. [107] [116] [117] [118] [119] Как избыточное, так и недостаточное производство белка посредством активности mTOR, по-видимому, способствует нарушению обучения и памяти. Более того, учитывая, что дефицит, возникающий в результате гиперактивности mTOR, можно уменьшить с помощью лечения рапамицином, вполне возможно, что mTOR играет важную роль во влиянии на когнитивные функции посредством синаптической пластичности. [103] [120] Дополнительные доказательства активности mTOR при нейродегенерации получены из недавних результатов, демонстрирующих, что eIF2α-P, вышестоящая мишень пути mTOR, опосредует гибель клеток при прионных заболеваниях посредством устойчивого ингибирования трансляции. [121]
Некоторые данные также указывают на роль mTOR в снижении клиренса Aβ. mTOR является негативным регулятором аутофагии; [122] следовательно, гиперактивность передачи сигналов mTOR должна снижать клиренс Aβ в головном мозге с болезнью Альцгеймера. Нарушения аутофагии могут быть потенциальным источником патогенеза заболеваний, связанных с неправильным сворачиванием белков, включая БА. [123] [124] [125] [126] [127] [128] Исследования с использованием мышиных моделей болезни Хантингтона показывают, что лечение рапамицином облегчает клиренс агрегатов хантингтина. [129] [130] Возможно, то же самое лечение может быть полезно и для очистки отложений Aβ.
Лимфопролиферативные заболевания
[ редактировать ]Гиперактивные пути mTOR были выявлены при некоторых лимфопролиферативных заболеваниях, таких как аутоиммунный лимфопролиферативный синдром (АЛПС), [131] мультицентрическая болезнь Кастлемана , [132] и посттрансплантационное лимфопролиферативное заболевание (ПТЛД). [133]
Синтез белка и рост клеток
[ редактировать ]Активация mTORC1 необходима для синтеза белка миофибриллярных мышц и гипертрофии скелетных мышц у людей в ответ как на физические упражнения , так и на прием определенных аминокислот или производных аминокислот. [134] [135] Стойкая инактивация передачи сигналов mTORC1 в скелетных мышцах способствует потере мышечной массы и силы во время мышечного истощения в пожилом возрасте, раковой кахексии и атрофии мышц из -за отсутствия физической активности . [134] [135] [136] Активация mTORC2, по-видимому, опосредует рост нейритов в дифференцированных клетках нейро2а мыши . [137] Периодическая активация mTOR в префронтальных нейронах с помощью β-гидрокси-β-метилбутирата ингибирует возрастное снижение когнитивных функций, связанное с обрезкой дендритов у животных, что также наблюдается у людей. [138]


Лизосомальное повреждение ингибирует mTOR и индуцирует аутофагию
[ редактировать ]Активный mTORC1 расположен на лизосомах . mTOR заблокирован [140] когда лизосомальная мембрана повреждается различными экзогенными или эндогенными агентами, такими как проникающие через мембрану бактерии , проникающие через мембрану химические вещества, образующие осмотически активные продукты (этот тип повреждения можно смоделировать с использованием проникающих через мембрану предшественников дипептидов, которые полимеризуются в лизосомах), агрегатов амилоидных белков (см. выше раздел о болезни Альцгеймера ) и цитоплазматические органические или неорганические включения, включая кристаллы уратов и кристаллический кремнезем . [140] Процесс инактивации mTOR после лизосомы/эндомембраны опосредуется белковым комплексом, называемым GALTOR. [140] В сердце ГАЛТОРА [140] представляет собой галектин-8 , член суперсемейства β-галактозидсвязывающих цитозольных лектинов, называемых галектинами , который распознает повреждение лизосомальной мембраны путем связывания с экспонированными гликанами на просветной стороне ограничивающей эндомембраны. После повреждения мембраны галектин-8, который обычно связывается с mTOR в гомеостатических условиях, больше не взаимодействует с mTOR, а теперь вместо этого связывается с SLC38A9 , RRAGA / RRAGB и LAMTOR1 , ингибируя Ragulator (комплекс LAMTOR1-5). обмена гуаниновых нуклеотидов функцию - [140]
TOR является негативным регулятором аутофагии в целом, его лучше всего изучать во время реакции на голодание. [141] [142] [143] [144] [145] что является метаболической реакцией. Однако во время лизосомального повреждения ингибирование mTOR активирует реакцию аутофагии в функции контроля качества, что приводит к процессу, называемому лизофагией. [146] который удаляет поврежденные лизосомы. На этой стадии другой галектин , галектин-3 , взаимодействует с TRIM16, направляя селективную аутофагию поврежденных лизосом. [147] [148] TRIM16 собирает ULK1 и основные компоненты (Beclin 1 и ATG16L1 ) других комплексов (Beclin 1 - VPS34 - ATG14 и ATG16L1 - ATG5 - ATG12 ), инициируя аутофагию . [148] многие из них находятся под отрицательным контролем непосредственно mTOR, например, комплекса ULK1-ATG13, [143] [144] [145] или косвенно, например, компоненты PI3K класса III (Beclin 1, ATG14 и VPS34), поскольку они зависят от активации фосфорилирования с помощью ULK1, когда оно не ингибируется mTOR. Эти компоненты, управляющие аутофагией, физически и функционально связаны друг с другом, интегрируя все процессы, необходимые для формирования аутофагосом: (i) комплекс ULK1- ATG13 - FIP200/RB1CC1 связывается с механизмом конъюгации LC3B / GABARAP посредством прямых взаимодействий между FIP200/RB1CC1 и ATG16L1. , [149] [150] [151] (ii) Комплекс ULK1 -ATG13- FIP200/RB1CC1 связывается с Beclin 1 - VPS34 - ATG14 посредством прямых взаимодействий между ATG13 доменом HORMA и ATG14 , [152] (iii) ATG16L1 взаимодействует с WIPI2 , который связывается с PI3P , ферментативным продуктом класса III PI3K Beclin 1-VPS34-ATG14. [153] Таким образом, инактивация mTOR, инициированная через GALTOR [140] при повреждении лизосом плюс одновременная активация посредством галектина-9 (который также распознает нарушение лизосомальной мембраны). AMPK [140] который непосредственно фосфорилирует и активирует ключевые компоненты ( ULK1 , [154] Беклин 1 [155] ) перечисленных выше систем аутофагии и дополнительно инактивирует mTORC1, [156] [157] позволяет добиться сильной индукции аутофагии и аутофагического удаления поврежденных лизосом.
Кроме того, несколько типов событий убиквитинирования параллельны и дополняют процессы, управляемые галектином: убиквитинирование TRIM16-ULK1-Beclin-1 стабилизирует эти комплексы, способствуя активации аутофагии, как описано выше. [148] ATG16L1 обладает внутренней аффинностью связывания с убиквитином. [151] ); тогда как убиквитинирование с помощью убиквитинлигазы, специфичной для гликопротеина FBXO27, нескольких подвергающихся повреждению гликозилированных лизосомальных мембранных белков, таких как LAMP1 , LAMP2 , GNS/ N-ацетилглюкозамин-6-сульфатаза , TSPAN6/ тетраспанин-6 , PSAP/ просапозин и TMEM192/ трансмембранный белок 192 [158] может способствовать осуществлению лизофагии через аутофагические рецепторы, такие как p62/ SQSTM1 , который рекрутируется во время лизофагии, [151] или другие подлежащие определению функции.
Склеродермия
[ редактировать ]Склеродермия , также известная как системный склероз , представляет собой хроническое системное аутоиммунное заболевание , характеризующееся уплотнением ( склеро ) кожи ( дермы ), поражающее внутренние органы в более тяжелых формах. [159] [160] mTOR играет роль в фиброзных заболеваниях и аутоиммунитете, а блокада пути mTORC исследуется как средство лечения склеродермии. [9]
Ингибиторы mTOR в качестве терапии
[ редактировать ]Трансплантация
[ редактировать ]Ингибиторы mTOR, например рапамицин , уже используются для предотвращения отторжения трансплантата .
Болезнь накопления гликогена
[ редактировать ]В некоторых статьях сообщалось, что рапамицин может ингибировать mTORC1, что позволяет увеличить фосфорилирование GS (гликогенсинтазы) в скелетных мышцах. Это открытие представляет собой потенциально новый терапевтический подход к болезни накопления гликогена , которая включает накопление гликогена в мышцах.
Противораковый
[ редактировать ]Для лечения рака человека используются два основных ингибитора mTOR: темсиролимус и эверолимус . Ингибиторы mTOR нашли применение при лечении различных злокачественных новообразований, включая почечно-клеточный рак (темсиролимус), рак поджелудочной железы , рак молочной железы и почечно-клеточный рак (эверолимус). [161] Полный механизм действия этих агентов не ясен, но считается, что они действуют, нарушая ангиогенез опухоли и вызывая нарушение перехода G1/S . [162]
Антивозрастной
[ редактировать ]Ингибиторы mTOR могут быть полезны для лечения/профилактики некоторых возрастных состояний. [163] включая нейродегенеративные заболевания, такие как болезнь Альцгеймера и болезнь Паркинсона . [164] После кратковременного лечения ингибиторами mTOR дактолисибом и эверолимусом у пожилых людей (65 лет и старше) у пациентов, получавших лечение, наблюдалось снижение количества инфекций в течение года. [165]
различные природные соединения, в том числе галлат эпигаллокатехина (EGCG), кофеин , куркумин , берберин , кверцетин , ресвератрол и птеростильбен , ингибируют mTOR при применении к изолированным клеткам в культуре. Сообщалось, что [166] [167] [168] Пока не существует качественных доказательств того, что эти вещества ингибируют передачу сигналов mTOR или продлевают продолжительность жизни, когда люди принимают их в качестве пищевых добавок , несмотря на обнадеживающие результаты на животных, таких как плодовые мухи и мыши. Различные судебные разбирательства продолжаются. [169] [170]
Взаимодействия
[ редактировать ]Было показано, что механическая мишень рапамицина взаимодействует с: [171]
- АБЛ1 , [172]
- АКТ1 , [52] [173] [174]
- ИГФ-ИР , [12]
- ИнсР , [12]
- КЛИП1 , [175]
- ЭИФ3Ф [176]
- EIF4EBP1 , [45] [177] [178] [179] [180] [181] [182] [183]
- ФКБП1А , [13] [50] [184] [185] [186] [187]
- ГПХН , [188]
- КИАА1303 , [13] [45] [49] [50] [85] [177] [178] [179] [189] [190] [191] [192] [193] [194] [195] [196] [197] [198] [199] [200]
- ПРКЦД , [201]
- РЕБ , [180] [202] [203] [204]
- РИКТОР , [13] [49] [50] [191] [197] [199] [200]
- РПС6КБ1 , [45] [178] [180] [181] [182] [196] [199] [205] [206] [207] [208] [209] [210] [211] [212]
- СТАТ1 , [213]
- СТАТ3 , [214] [215]
- Двухпоровые каналы : TPCN1 ; ТКПН2 , [216] и
- УБКЛН1 . [217]
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с GRCh38: Версия Ensembl 89: ENSG00000198793 – Ensembl , май 2017 г.
- ^ Перейти обратно: а б с GRCm38: выпуск Ensembl 89: ENSMUSG00000028991 – Ensembl , май 2017 г.
- ^ «Ссылка на Human PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Ссылка на Mouse PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ Сейбрс С.Дж., Мартин М.М., Брунн Г.Дж. и др. (январь 1995 г.). «Выделение белка-мишени комплекса FKBP12-рапамицин в клетках млекопитающих» . Ж. Биол. Хим . 270 (2): 815–22. дои : 10.1074/jbc.270.2.815 . ПМИД 7822316 .
- ^ Перейти обратно: а б с д Браун Э.Дж., Альберс М.В., Шин Т.Б. и др. (июнь 1994 г.). «Белок млекопитающих, на который нацелен комплекс G1-арестант рапамицин-рецептор». Природа . 369 (6483): 756–8. Бибкод : 1994Natur.369..756B . дои : 10.1038/369756a0 . ПМИД 8008069 . S2CID 4359651 .
- ^ Перейти обратно: а б с д Сабатини Д.М., Эрджюмент-Бромаж Х., Луи М. и др. (июль 1994 г.). «RAFT1: белок млекопитающих, который связывается с FKBP12 рапамицин-зависимым образом и гомологичен дрожжевым TOR». Клетка . 78 (1): 35–43. дои : 10.1016/0092-8674(94)90570-3 . ПМИД 7518356 . S2CID 33647539 .
- ^ Перейти обратно: а б Сейбрс С.Дж., Мартин М.М., Брунн Г.Дж. и др. (январь 1995 г.). «Выделение белка-мишени комплекса FKBP12-рапамицин в клетках млекопитающих» . Журнал биологической химии . 270 (2): 815–22. дои : 10.1074/jbc.270.2.815 . ПМИД 7822316 .
- ^ Перейти обратно: а б Митра А., Луна Дж.И., Марусина А.И. и др. (ноябрь 2015 г.). «Двойное ингибирование mTOR необходимо для предотвращения TGF-β-опосредованного фиброза: последствия для склеродермии» . Журнал исследовательской дерматологии . 135 (11): 2873–6. дои : 10.1038/jid.2015.252 . ПМЦ 4640976 . ПМИД 26134944 .
- ^ Перейти обратно: а б с д и ж Липтон Дж.О., Шахин М. (октябрь 2014 г.). «Неврология mTOR» . Нейрон . 84 (2): 275–291. дои : 10.1016/j.neuron.2014.09.034 . ПМК 4223653 . ПМИД 25374355 .
Сигнальный путь mTOR действует как интегратор молекулярных систем, поддерживая взаимодействие организма и клеток с окружающей средой. Путь mTOR регулирует гомеостаз, напрямую влияя на синтез белка, транскрипцию, аутофагию, метаболизм, а также биогенез и поддержание органелл. Неудивительно, что передача сигналов mTOR участвует во всей иерархии функций мозга, включая пролиферацию нервных стволовых клеток, сборку и поддержание цепей, пластичность, зависящую от опыта, и регуляцию сложного поведения, такого как питание, сон и циркадные ритмы. ...
Функция mTOR опосредована двумя большими биохимическими комплексами, определяемыми соответствующим белковым составом и широко рассмотренными в других источниках (Dibble and Manning, 2013; Laplante and Sabatini, 2012) (Figure 1B). Вкратце, общими для комплекса mTOR 1 (mTORC1) и комплекса mTOR 2 (mTORC2) являются: сам mTOR, летальный для млекопитающих с белком 8 sec13 (mLST8; также известный как GβL), и ингибирующий домен DEP, содержащий белок, взаимодействующий с mTOR ( ДЕПТОР). Специфичным для mTORC1 является связанный с регулятором белок мишени рапамицина (Raptor) млекопитающих и богатый пролином субстрат Akt массой 40 кДа (PRAS40) (Kim et al., 2002; Laplante and Sabatini, 2012). Raptor необходим для активности mTORC1. Комплекс mTORC2 включает нечувствительный к рапамицину компаньон mTOR (Rictor), белок 1, взаимодействующий с киназой MAP млекопитающих (mSIN1), и белки, наблюдаемые с rictor 1 и 2 (PROTOR 1 и 2) (Jacinto et al., 2006; Jacinto и др., 2004; Пирс и др., 2007; Сарбасов и др., 2004) (рис. 1Б). Rictor и mSIN1 имеют решающее значение для функции mTORC2.
Рисунок 1: Доменная структура киназы mTOR и компонентов mTORC1 и mTORC2.
Рисунок 2. Сигнальный путь mTOR - ^ Перейти обратно: а б с Хэй Н., Соненберг Н. (август 2004 г.). «Вверх и вниз по течению от mTOR» . Гены и развитие . 18 (16): 1926–45. дои : 10.1101/gad.1212704 . ПМИД 15314020 .
- ^ Перейти обратно: а б с д Инь Ю, Хуа Х, Ли М и др. (январь 2016 г.). «mTORC2 способствует активации рецептора инсулиноподобного фактора роста I типа и активации рецептора инсулина посредством тирозинкиназной активности mTOR» . Клеточные исследования . 26 (1): 46–65. дои : 10.1038/cr.2015.133 . ПМЦ 4816127 . ПМИД 26584640 .
- ^ Перейти обратно: а б с д Хасинто Э., Ловит Р., Шмидт А. и др. (ноябрь 2004 г.). «Комплекс 2 TOR млекопитающих контролирует актиновый цитоскелет и нечувствителен к рапамицину». Природная клеточная биология . 6 (11): 1122–8. дои : 10.1038/ncb1183 . ПМИД 15467718 . S2CID 13831153 .
- ^ Пауэрс Т (ноябрь 2022 г.). Келлог Д. (ред.). «История происхождения рапамицина: системная предвзятость в биомедицинских исследованиях и политика холодной войны» . Молекулярная биология клетки . 33 (13). doi : 10.1091/mbc.E22-08-0377 . ПМЦ 9634974 . ПМИД 36228182 .
- ^ Сегал С.Н., Бейкер Х., Везина С. (октябрь 1975 г.). «Рапамицин (AY-22,989), новый противогрибковый антибиотик. II. Ферментация, выделение и характеристика». Журнал антибиотиков . 28 (10): 727–732. дои : 10.7164/антибиотики.28.727 . ПМИД 1102509 .
- ^ Перейти обратно: а б с Хейтман Дж., Мовва Н.Р., Холл Миннесота (август 1991 г.). «Цели для остановки клеточного цикла иммунодепрессантом рапамицином в дрожжах». Наука . 253 (5022): 905–9. Бибкод : 1991Sci...253..905H . дои : 10.1126/science.1715094 . ПМИД 1715094 . S2CID 9937225 .
- ^ Кунц Дж., Энрикес Р., Шнайдер У. и др. (май 1993 г.). «Мишень рапамицина в дрожжах, TOR2, является важным гомологом фосфатидилинозитолкиназы, необходимым для прогрессирования G1». Клетка . 73 (3): 585–596. дои : 10.1016/0092-8674(93)90144-F . ПМИД 8387896 . S2CID 42926249 .
- ^ Кафферки Р., Янг П.Р., Маклафлин М.М. и др. (октябрь 1993 г.). «Доминантные миссенс-мутации в новом дрожжевом белке, связанном с фосфатидилинозитол-3-киназой млекопитающих и VPS34, устраняют цитотоксичность рапамицина» . Мол Клеточная Биол . 13 (10): 6012–23. дои : 10.1128/MCB.13.10.6012 . ПМЦ 364661 . ПМИД 8413204 .
- ^ Перейти обратно: а б Магнусон Б., Эким Б., Фингар, округ Колумбия (январь 2012 г.). «Регуляция и функция рибосомальной протеинкиназы S6 (S6K) в сигнальных сетях mTOR». Биохимический журнал . 441 (1): 1–21. дои : 10.1042/BJ20110892 . ПМИД 22168436 . S2CID 12932678 .
- ^ Перейти обратно: а б Авраам Р.Т., Видеррехт Г.Дж. (1996). «Иммунофармакология рапамицина». Ежегодный обзор иммунологии . 14 : 483–510. doi : 10.1146/annurev.immunol.14.1.483 . ПМИД 8717522 .
- ^ Бирер Б.Е., Маттила П.С., Штандарт Р.Ф. и др. (декабрь 1990 г.). «Два различных пути передачи сигнала в Т-лимфоцитах ингибируются комплексами, образующимися между иммунофилином и FK506 или рапамицином» . Труды Национальной академии наук Соединенных Штатов Америки . 87 (23): 9231–5. Бибкод : 1990PNAS...87.9231B . дои : 10.1073/pnas.87.23.9231 . ПМЦ 55138 . ПМИД 2123553 .
- ^ Bierer BE, Somers PK, Wandless TJ и др. (октябрь 1990 г.). «Исследование иммунодепрессивного действия с помощью неприродного иммунофилинового лиганда». Наука . 250 (4980): 556–9. Бибкод : 1990Sci...250..556B . дои : 10.1126/science.1700475 . ПМИД 1700475 . S2CID 11123023 .
- ^ Дюмон Ф.Дж., Мелино М.Р., Старух М.Дж. и др. (февраль 1990 г.). «Имуносупрессивные макролиды FK-506 и рапамицин действуют как реципрокные антагонисты в мышиных Т-клетках» . Дж Иммунол . 144 (4): 1418–24. дои : 10.4049/jimmunol.144.4.1418 . ПМИД 1689353 . S2CID 44256944 .
- ^ Дюмон Ф.Дж., Старух М.Ю., Копрак С.Л. и др. (январь 1990 г.). «Отличные механизмы подавления активации мышиных Т-клеток родственными макролидами FK-506 и рапамицином» . Дж Иммунол . 144 (1): 251–8. дои : 10.4049/jimmunol.144.1.251 . ПМИД 1688572 . S2CID 13201695 .
- ^ Хардинг М.В., Галат А., Юлинг Д.Е. и др. (октябрь 1989 г.). «Рецептор иммунодепрессанта FK506 представляет собой цис-транс-пептидилпролилизомеразу». Природа . 341 (6244): 758–60. Бибкод : 1989Natur.341..758H . дои : 10.1038/341758a0 . ПМИД 2477715 . S2CID 4349152 .
- ^ Фретц Х., Альберс М.В., Галат А. и др. (февраль 1991 г.). «Рапамицин и белки, связывающие FK506 (иммунофилины)». Журнал Американского химического общества . 113 (4): 1409–1411. дои : 10.1021/ja00004a051 .
- ^ Лю Дж., Фармер Дж.Д., Лейн В.С. и др. (август 1991 г.). «Кальцинеурин является общей мишенью комплексов циклофилин-циклоспорин А и FKBP-FK506». Клетка . 66 (4): 807–15. дои : 10.1016/0092-8674(91)90124-H . ПМИД 1715244 . S2CID 22094672 .
- ^ Кунц Дж., Энрикес Р., Шнайдер У. и др. (май 1993 г.). «Мишень рапамицина в дрожжах, TOR2, является важным гомологом фосфатидилинозитолкиназы, необходимым для прогрессирования G1». Клетка . 73 (3): 585–596. дои : 10.1016/0092-8674(93)90144-F . ПМИД 8387896 . S2CID 42926249 .
- ^ Хейтман Дж. (ноябрь 2015 г.). «Об открытии TOR как мишени рапамицина» . ПЛОС Патогены . 11 (11): e1005245. дои : 10.1371/journal.ppat.1005245 . ПМЦ 4634758 . ПМИД 26540102 .
- ^ Кеннеди Б.К., Ламминг Д.В. (2016). «Механистическая цель рапамицина: главный проводник метаболизма и старения» . Клеточный метаболизм . 23 (6): 990–1003. дои : 10.1016/j.cmet.2016.05.009 . ПМЦ 4910876 . ПМИД 27304501 .
- ^ «Символьный отчет для МТОР» . Данные HGNC для MTOR . Комитет по генной номенклатуре Хьюго . 1 сентября 2020 г. Проверено 17 декабря 2020 г.
- ^ Токунага С., Ёсино К., Ёнезава К. (январь 2004 г.). «mTOR объединяет пути восприятия аминокислот и энергии». Связь с биохимическими и биофизическими исследованиями . 313 (2): 443–6. дои : 10.1016/j.bbrc.2003.07.019 . ПМИД 14684182 .
- ^ Випперман М.Ф., Монтроуз, округ Колумбия, Готто А.М. и др. (2019). «Цель рапамицина для млекопитающих: метаболический реостат для регулирования функции жировой ткани и здоровья сердечно-сосудистой системы» . Американский журнал патологии . 189 (3): 492–501. дои : 10.1016/j.ajpath.2018.11.013 . ПМК 6412382 . ПМИД 30803496 .
- ^ Биверс К.С., Ли Ф., Лю Л. и др. (август 2006 г.). «Куркумин ингибирует мишень опосредованных рапамицином сигнальных путей в раковых клетках, воздействующую на млекопитающих». Международный журнал рака . 119 (4): 757–64. дои : 10.1002/ijc.21932 . ПМИД 16550606 . S2CID 25454463 .
- ^ Кеннеди Б.К., Ламминг Д.В. (июнь 2016 г.). «Механистическая цель рапамицина: главный проводник метаболизма и старения» . Клеточный метаболизм . 23 (6): 990–1003. дои : 10.1016/j.cmet.2016.05.009 . ПМЦ 4910876 . ПМИД 27304501 .
- ^ Хуан С., Хоутон П.Дж. (декабрь 2001 г.). «Механизмы устойчивости к рапамицинам». Обновления по лекарственной устойчивости . 4 (6): 378–91. дои : 10.1054/drup.2002.0227 . ПМИД 12030785 .
- ^ Перейти обратно: а б Хуанг С., Бьёрнсти М.А., Хоутон П.Дж. (2003). «Рапамицины: механизм действия и клеточная резистентность» . Биология и терапия рака . 2 (3): 222–32. дои : 10.4161/cbt.2.3.360 . ПМИД 12878853 .
- ^ Перейти обратно: а б Ингарджиола С., Туркето Дуарте Г., Робалья С. и др. (октябрь 2020 г.). «Растительная мишень рапамицина: обеспечение питания и обмена веществ в фотосинтетических организмах» . Гены . 11 (11): 1285. doi : 10.3390/genes11111285 . ПМЦ 7694126 . ПМИД 33138108 .
- ^ Перейти обратно: а б Ши Л., Ву Ю, Шин Дж. (июль 2018 г.). «TOR-сигнализация у растений: сохранение и инновации» . Разработка . 145 (13). дои : 10.1242/dev.160887 . ПМК 6053665 . ПМИД 29986898 .
- ^ Сюн Ю, Шин Дж (февраль 2014 г.). «Роль мишени сигнальных сетей рапамицина в росте и метаболизме растений» . Физиология растений . 164 (2): 499–512. дои : 10.1104/стр.113.229948 . ПМЦ 3912084 . ПМИД 24385567 .
- ^ Вулшлегер С., Ловит Р., Холл Миннесота (февраль 2006 г.). «Передача сигналов TOR в росте и метаболизме» . Клетка . 124 (3): 471–84. дои : 10.1016/j.cell.2006.01.016 . ПМИД 16469695 .
- ^ Бетц С., Холл Миннесота (ноябрь 2013 г.). «Где mTOR и что он там делает?» . Журнал клеточной биологии . 203 (4): 563–74. дои : 10.1083/jcb.201306041 . ПМЦ 3840941 . ПМИД 24385483 .
- ^ Груневуд М.Дж., Зварткруис Ф.Дж. (август 2013 г.). «Реб и Рэгс объединяются в лизосоме, чтобы активировать mTORC1». Труды Биохимического общества . 41 (4): 951–5. дои : 10.1042/bst20130037 . ПМИД 23863162 . S2CID 8237502 .
- ^ Эфеян А., Зонджу Р., Сабатини Д.М. (сентябрь 2012 г.). «Аминокислоты и mTORC1: от лизосом к болезни» . Тенденции молекулярной медицины . 18 (9): 524–33. doi : 10.1016/j.molmed.2012.05.007 . ПМЦ 3432651 . ПМИД 22749019 .
- ^ Перейти обратно: а б с д и ж Ким Д.Х., Сарбасов Д.Д., Али С.М. и др. (июль 2002 г.). «mTOR взаимодействует с хищником, образуя чувствительный к питательным веществам комплекс, который передает сигналы механизму роста клеток» . Клетка . 110 (2): 163–75. дои : 10.1016/S0092-8674(02)00808-5 . ПМИД 12150925 .
- ^ Ким Д.Х., Сарбасов Д.Д., Али С.М. и др. (апрель 2003 г.). «GbetaL, положительный регулятор рапамицин-чувствительного пути, необходимый для чувствительного к питательным веществам взаимодействия между хищником и mTOR» . Молекулярная клетка . 11 (4): 895–904. дои : 10.1016/S1097-2765(03)00114-X . ПМИД 12718876 .
- ^ Фанг Ю., Вилелла-Бах М., Бахманн Р. и др. (ноябрь 2001 г.). «Митогенная активация передачи сигналов mTOR, опосредованная фосфатидной кислотой». Наука . 294 (5548): 1942–5. Бибкод : 2001Sci...294.1942F . дои : 10.1126/science.1066015 . ПМИД 11729323 . S2CID 44444716 .
- ^ Бонд П. (март 2016 г.). «Краткий обзор регуляции mTORC1 с помощью факторов роста, энергетического статуса, аминокислот и механических раздражителей» . Дж. Межд. Соц. Спортивный Нутр . 13 :8. дои : 10.1186/s12970-016-0118-y . ПМЦ 4774173 . ПМИД 26937223 .
- ^ Перейти обратно: а б с Фриас М.А., Торин С.С., Яффе Дж.Д. и др. (сентябрь 2006 г.). «mSin1 необходим для фосфорилирования Akt/PKB, и его изоформы определяют три различных mTORC2» . Современная биология . 16 (18): 1865–70. Бибкод : 2006CBio...16.1865F . дои : 10.1016/j.cub.2006.08.001 . ПМИД 16919458 .
- ^ Перейти обратно: а б с д и Сарбасов Д.Д., Али С.М., Ким Д.Х. и др. (июль 2004 г.). «Rictor, новый партнер по связыванию mTOR, определяет нечувствительный к рапамицину и независимый от хищников путь, который регулирует цитоскелет» . Современная биология . 14 (14): 1296–302. Бибкод : 2004CBio...14.1296D . дои : 10.1016/j.cub.2004.06.054 . ПМИД 15268862 .
- ^ Бетц С., Страка Д., Прешианотто-Башонг С. и др. (июль 2013 г.). «Особенная статья: передача сигналов комплекса mTOR 2-Akt на мембранах эндоплазматического ретикулума, связанных с митохондриями (MAM), регулирует физиологию митохондрий» . Труды Национальной академии наук Соединенных Штатов Америки . 110 (31): 12526–34. дои : 10.1073/pnas.1302455110 . ПМК 3732980 . ПМИД 23852728 .
- ^ Перейти обратно: а б Сарбасов Д.Д., Гертин Д.А., Али С.М. и др. (февраль 2005 г.). «Фосфорилирование и регуляция Akt/PKB комплексом rictor-mTOR». Наука . 307 (5712): 1098–101. Бибкод : 2005Sci...307.1098S . дои : 10.1126/science.1106148 . ПМИД 15718470 . S2CID 45837814 .
- ^ Стивенс Л., Андерсон К., Стоко Д. и др. (январь 1998 г.). «Киназы протеинкиназы B, которые опосредуют фосфатидилинозитол-3,4,5-трифосфат-зависимую активацию протеинкиназы B». Наука . 279 (5351): 710–4. Бибкод : 1998Sci...279..710S . дои : 10.1126/science.279.5351.710 . ПМИД 9445477 .
- ^ Карози Дж.М., Фурье С., Бенсалем Дж. и др. (2022). «Ось mTOR-лизосома в центре старения» . Открытая биография FEBS . 12 (4): 739–757. дои : 10.1002/2211-5463.13347 . ПМЦ 8972043 . ПМИД 34878722 .
- ^ Чжоу С, Тан X, Чен Х (2018). «Сиртуины и инсулинорезистентность» . Границы эндокринологии . 9 : 748. дои : 10.3389/fendo.2018.00748 . ПМК 6291425 . ПМИД 30574122 .
- ^ Бэкл Дж.Дж., Чен Н., Винер Д.А. (2023). «Хроническое воспаление и признаки старения» . Молекулярный метаболизм . 74 : 101755. doi : 10.1016/j.molmet.2023.101755 . ПМЦ 10359950 . ПМИД 37329949 .
- ^ Перейти обратно: а б Ламминг Д.В., Йе Л., Катаджисто П. и др. (март 2012 г.). «Рапамицин-индуцированная инсулинорезистентность опосредована потерей mTORC2 и не связана с долголетием» . Наука . 335 (6076): 1638–43. Бибкод : 2012Sci...335.1638L . дои : 10.1126/science.1215135 . ПМК 3324089 . ПМИД 22461615 .
- ^ Зинзалла В., Страчка Д., Опплигер В. и др. (март 2011 г.). «Активация mTORC2 путем ассоциации с рибосомой» . Клетка . 144 (5): 757–68. дои : 10.1016/j.cell.2011.02.014 . ПМИД 21376236 .
- ^ Чжан Ф., Чжан Х., Ли М. и др. (ноябрь 2010 г.). «Компонент комплекса mTOR Rictor взаимодействует с PKCzeta и регулирует метастазирование раковых клеток» . Исследования рака . 70 (22): 9360–70. дои : 10.1158/0008-5472.CAN-10-0207 . ПМИД 20978191 .
- ^ Гертин Д.А., Стивенс Д.М., Торин CC и др. (декабрь 2006 г.). «Абляция у мышей компонентов mTORC raptor, rictor или mLST8 показывает, что mTORC2 необходим для передачи сигналов Akt-FOXO и PKCalpha, но не S6K1» . Развивающая клетка . 11 (6): 859–71. дои : 10.1016/j.devcel.2006.10.007 . ПМИД 17141160 .
- ^ Гу Ю, Линднер Дж, Кумар А и др. (март 2011 г.). «Rictor/mTORC2 необходим для поддержания баланса между пролиферацией бета-клеток и их размером» . Диабет . 60 (3): 827–37. дои : 10.2337/db10-1194 . ПМК 3046843 . ПМИД 21266327 .
- ^ Ламминг Д.В., Демиркан Г., Бойлан Дж.М. и др. (январь 2014 г.). «Передача сигналов в печени посредством механистической мишени комплекса рапамицина 2 (mTORC2)» . Журнал ФАСЭБ . 28 (1): 300–15. дои : 10.1096/fj.13-237743 . ПМЦ 3868844 . ПМИД 24072782 .
- ^ Кумар А., Лоуренс Дж.К., Юнг Д.И. и др. (июнь 2010 г.). «Специфическое для жировых клеток удаление риктора у мышей нарушает регулируемый инсулином метаболизм жировых клеток и глюкозы и липидов во всем организме» . Диабет . 59 (6): 1397–406. дои : 10.2337/db09-1061 . ПМК 2874700 . ПМИД 20332342 .
- ^ Ламминг Д.В., Михайлова М.М., Катаджисто П. и др. (октябрь 2014 г.). «Истощение Rictor, важного белкового компонента mTORC2, сокращает продолжительность жизни мужчин» . Стареющая клетка . 13 (5): 911–7. дои : 10.1111/acel.12256 . ПМК 4172536 . ПМИД 25059582 .
- ^ Фельдман М.Е., Апсель Б., Уотила А. и др. (февраль 2009 г.). «Ингибиторы активного центра mTOR нацелены на устойчивые к рапамицину результаты mTORC1 и mTORC2» . ПЛОС Биология . 7 (2): е38. дои : 10.1371/journal.pbio.1000038 . ПМЦ 2637922 . ПМИД 19209957 .
- ^ Ву Дж., Лю Дж., Чен Э.Б. и др. (сентябрь 2013 г.). «Увеличение продолжительности жизни млекопитающих, а также сегментарное и тканеспецифическое замедление старения после генетического снижения экспрессии mTOR» . Отчеты по ячейкам . 4 (5): 913–20. дои : 10.1016/j.celrep.2013.07.030 . ПМЦ 3784301 . ПМИД 23994476 .
- ^ Лоулор М.А., Мора А., Эшби П.Р. и др. (июль 2002 г.). «Основная роль PDK1 в регуляции размера и развития клеток у мышей» . Журнал ЭМБО . 21 (14): 3728–38. дои : 10.1093/emboj/cdf387 . ПМК 126129 . ПМИД 12110585 .
- ^ Ян З.З., Чопп О., Бодри А. и др. (апрель 2004 г.). «Физиологические функции протеинкиназы В/Акт». Труды Биохимического общества . 32 (Часть 2): 350–4. дои : 10.1042/BST0320350 . ПМИД 15046607 .
- ^ Нодзима А., Ямашита М., Ёсида Ю. и др. (01.01.2013). «Гаплонедостаточность akt1 продлевает продолжительность жизни мышей» . ПЛОС ОДИН . 8 (7): e69178. Бибкод : 2013PLoSO...869178N . дои : 10.1371/journal.pone.0069178 . ПМЦ 3728301 . ПМИД 23935948 .
- ^ Креспо Дж.Л., Холл Миннесота (декабрь 2002 г.). «Выяснение передачи сигналов TOR и действия рапамицина: уроки Saccharomyces cerevisiae» . Обзоры микробиологии и молекулярной биологии . 66 (4): 579–91, оглавление. doi : 10.1128/mmbr.66.4.579-591.2002 . ПМК 134654 . ПМИД 12456783 .
- ^ Питер Г.Дж., Дюринг Л., Ахмед А. (март 2006 г.). «Репрессия углеродных катаболитов регулирует проникновение аминокислот в Saccharomyces cerevisiae через сигнальный путь TOR» . Журнал биологической химии . 281 (9): 5546–52. дои : 10.1074/jbc.M513842200 . ПМИД 16407266 .
- ^ Перейти обратно: а б Пауэрс Р.В., Каберлейн М., Колдуэлл С.Д. и др. (январь 2006 г.). «Продление хронологической продолжительности жизни дрожжей за счет снижения передачи сигналов пути TOR» . Гены и развитие . 20 (2): 174–84. дои : 10.1101/gad.1381406 . ПМЦ 1356109 . ПМИД 16418483 .
- ^ Перейти обратно: а б Каберляйн М., Пауэрс Р.В., Штеффен К.К. и др. (ноябрь 2005 г.). «Регуляция репликативной продолжительности жизни дрожжей с помощью TOR и Sch9 в ответ на питательные вещества». Наука . 310 (5751): 1193–6. Бибкод : 2005Sci...310.1193K . дои : 10.1126/science.1115535 . ПМИД 16293764 . S2CID 42188272 .
- ^ Цзя К., Чен Д., Риддл Д.Л. (август 2004 г.). «Путь TOR взаимодействует с сигнальным путем инсулина, регулируя развитие, метаболизм и продолжительность жизни личинок C. elegans». Разработка . 131 (16): 3897–906. дои : 10.1242/dev.01255 . ПМИД 15253933 . S2CID 10377667 .
- ^ Капахи П., Зид Б.М., Харпер Т. и др. (май 2004 г.). «Регуляция продолжительности жизни дрозофилы путем модуляции генов сигнального пути TOR» . Современная биология . 14 (10): 885–90. Бибкод : 2004CBio...14..885K . дои : 10.1016/j.cub.2004.03.059 . ПМК 2754830 . ПМИД 15186745 .
- ^ Харрисон Д.Э., Стронг Р., Шарп З.Д. и др. (июль 2009 г.). «Рапамицин, вводимый в позднем возрасте, продлевает продолжительность жизни генетически гетерогенных мышей» . Природа . 460 (7253): 392–5. Бибкод : 2009Natur.460..392H . дои : 10.1038/nature08221 . ПМК 2786175 . ПМИД 19587680 .
- ^ Миллер Р.А., Харрисон Д.Э., Астл К.М. и др. (июнь 2014 г.). «Увеличение продолжительности жизни у мышей, опосредованное рапамицином, зависит от дозы и пола и метаболически отличается от диетических ограничений» . Стареющая клетка . 13 (3): 468–77. дои : 10.1111/acel.12194 . ПМК 4032600 . ПМИД 24341993 .
- ^ Фок В.К., Чен Ю., Боков А. и др. (01.01.2014). «У мышей, которых кормили рапамицином, продолжительность жизни увеличивается, что связано с серьезными изменениями в транскриптоме печени» . ПЛОС ОДИН . 9 (1): e83988. Бибкод : 2014PLoSO...983988F . дои : 10.1371/journal.pone.0083988 . ПМЦ 3883653 . ПМИД 24409289 .
- ^ Арриола Апело С.И., Пампер С.П., Баар Э.Л. и др. (июль 2016 г.). «Периодическое введение рапамицина продлевает продолжительность жизни самок мышей C57BL/6J» . Журналы геронтологии. Серия А, Биологические и медицинские науки . 71 (7): 876–81. дои : 10.1093/gerona/glw064 . ПМЦ 4906329 . ПМИД 27091134 .
- ^ Попович ИГ, Анисимов ВН, Забежинский МА и др. (май 2014 г.). «Продление продолжительности жизни и профилактика рака у трансгенных мышей HER-2/neu, получавших низкие прерывистые дозы рапамицина» . Биология и терапия рака . 15 (5): 586–92. дои : 10.4161/cbt.28164 . ПМК 4026081 . ПМИД 24556924 .
- ^ Баар Э.Л., Карбахал К.А., Онг И.М. и др. (февраль 2016 г.). «Поло- и тканеспецифичные изменения в передаче сигналов mTOR с возрастом у мышей C57BL/6J» . Стареющая клетка . 15 (1): 155–66. дои : 10.1111/acel.12425 . ПМЦ 4717274 . ПМИД 26695882 .
- ^ Кэрон А., Ричард Д., Лапланте М. (июль 2015 г.). «Роль комплексов mTOR в метаболизме липидов». Ежегодный обзор питания . 35 : 321–48. doi : 10.1146/annurev-nutr-071714-034355 . ПМИД 26185979 .
- ^ Кота Д., Пру К., Смит К.А. и др. (май 2006 г.). «Передача сигналов mTOR в гипоталамусе регулирует потребление пищи». Наука . 312 (5775): 927–30. Бибкод : 2006Sci...312..927C . дои : 10.1126/science.1124147 . ПМИД 16690869 . S2CID 6526786 .
- ^ Перейти обратно: а б Криете А., Босл В.Дж., Букер Г. (июнь 2010 г.). «Модель старения клеточных систем, основанная на правилах, с использованием мотивов петли обратной связи, опосредованных реакциями на стресс» . PLOS Вычислительная биология . 6 (6): e1000820. Бибкод : 2010PLSCB...6E0820K . дои : 10.1371/journal.pcbi.1000820 . ПМЦ 2887462 . ПМИД 20585546 .
- ^ Перейти обратно: а б Шике С.М., Филлипс Д., Маккой Дж.П. и др. (сентябрь 2006 г.). «Мишень пути рапамицина (mTOR) у млекопитающих регулирует потребление кислорода и окислительную способность митохондрий» . Журнал биологической химии . 281 (37): 27643–52. дои : 10.1074/jbc.M603536200 . ПМИД 16847060 .
- ^ Есенкызы А, Салиев Т, Жаналиева М и др. (2020). «Полифенолы как миметики ограничения калорий и индукторы аутофагии в исследованиях старения» . Питательные вещества . 12 (5): 1344. дои : 10.3390/nu12051344 . ПМЦ 7285205 . ПМИД 32397145 .
- ^ Перейти обратно: а б Лаберж Р., Сан Ю., Орьяло А.В. и др. (2015). «MTOR регулирует секреторный фенотип, связанный с протуморогенным старением, путем стимулирования трансляции IL1A» . Природная клеточная биология . 17 (8): 1049–1061. дои : 10.1038/ncb3195 . ПМЦ 4691706 . ПМИД 26147250 .
- ^ Ван Р., Ю З., Сунчу Б. и др. (2017). «Рапамицин ингибирует секреторный фенотип стареющих клеток с помощью Nrf2-независимого механизма» . Стареющая клетка . 16 (3): 564–574. дои : 10.1111/acel.12587 . ПМК 5418203 . ПМИД 28371119 .
- ^ Ван Р., Сунчу Б., Перес VI (2017). «Рапамицин и ингибирование секреторного фенотипа». Экспериментальная геронтология . 94 : 89–92. дои : 10.1016/j.exger.2017.01.026 . ПМИД 28167236 . S2CID 4960885 .
- ^ Вайххарт Т (2018). «mTOR как регулятор продолжительности жизни, старения и клеточного старения: мини-обзор» . Геронтология . 84 (2): 127–134. дои : 10.1159/000484629 . ПМК 6089343 . ПМИД 29190625 .
- ^ Сюй К., Лю П., Вэй В. (декабрь 2014 г.). «Передача сигналов mTOR в онкогенезе» . Biochimica et Biophysical Acta (BBA) - Обзоры о раке . 1846 (2): 638–54. дои : 10.1016/j.bbcan.2014.10.007 . ПМК 4261029 . ПМИД 25450580 .
- ^ Гертен Д.А., Сабатини Д.М. (август 2005 г.). «Расширяющаяся роль mTOR при раке». Тенденции молекулярной медицины . 11 (8): 353–61. doi : 10.1016/j.molmed.2005.06.007 . ПМИД 16002336 .
- ^ Популо Х., Лопес Х.М., Соареш П. (2012). «Сигнальный путь mTOR при раке человека» . Международный журнал молекулярных наук . 13 (2): 1886–918. дои : 10.3390/ijms13021886 . ПМК 3291999 . ПМИД 22408430 .
- ^ Истон Дж. Б., Хоутон П. Дж. (октябрь 2006 г.). «mTOR и терапия рака». Онкоген . 25 (48): 6436–46. дои : 10.1038/sj.onc.1209886 . ПМИД 17041628 . S2CID 19250234 .
- ^ Зонджу Р., Эфеян А., Сабатини Д.М. (январь 2011 г.). «mTOR: от интеграции сигналов роста к раку, диабету и старению» . Nature Reviews Молекулярно-клеточная биология . 12 (1): 21–35. дои : 10.1038/nrm3025 . ПМК 3390257 . ПМИД 21157483 .
- ^ Томас Г.В., Тран С., Меллингхофф И.К. и др. (январь 2006 г.). «Фактор, индуцируемый гипоксией, определяет чувствительность к ингибиторам mTOR при раке почки». Природная медицина . 12 (1): 122–7. дои : 10.1038/nm1337 . ПМИД 16341243 . S2CID 1853822 .
- ^ Немазаный И., Эспейяк С., Пенде М. и др. (август 2013 г.). «Роль передачи сигналов PI3K, mTOR и Akt2 в онкогенезе печени посредством контроля экспрессии PKM2». Труды Биохимического общества . 41 (4): 917–22. дои : 10.1042/BST20130034 . ПМИД 23863156 .
- ^ Тан Г., Гудснук К., Куо Ш. и др. (сентябрь 2014 г.). «Утрата mTOR-зависимой макроаутофагии вызывает аутистический дефицит синаптической обрезки» . Нейрон . 83 (5): 1131–43. дои : 10.1016/j.neuron.2014.07.040 . ПМК 4159743 . ПМИД 25155956 .
- ^ Рознер М., Ханнедер М., Сигел Н. и др. (июнь 2008 г.). «Путь mTOR и его роль в генетических заболеваниях человека». Мутационные исследования . 659 (3): 284–92. дои : 10.1016/j.mrrev.2008.06.001 . ПМИД 18598780 .
- ^ Ли X, Алафузофф I, Сойнинен Х и др. (август 2005 г.). «Уровни mTOR и его нижестоящих мишеней для киназы 4E-BP1, eEF2 и eEF2 во взаимосвязи с тау в мозге при болезни Альцгеймера». Журнал ФЭБС . 272 (16): 4211–20. дои : 10.1111/j.1742-4658.2005.04833.x . ПМИД 16098202 . S2CID 43085490 .
- ^ Чано Т., Окабе Х., Хулетт С.М. (сентябрь 2007 г.). «Недостаточность RB1CC1 вызывает атрофию нейронов за счет изменения передачи сигналов mTOR и участвует в патологии болезни Альцгеймера». Исследования мозга . 1168 (1168): 97–105. дои : 10.1016/j.brainres.2007.06.075 . ПМИД 17706618 . S2CID 54255848 .
- ^ Селькое DJ (сентябрь 2008 г.). «Растворимые олигомеры бета-амилоидного белка ухудшают синаптическую пластичность и поведение» . Поведенческие исследования мозга . 192 (1): 106–13. дои : 10.1016/j.bbr.2008.02.016 . ПМК 2601528 . ПМИД 18359102 .
- ^ Перейти обратно: а б Оддо С. (январь 2012 г.). «Роль передачи сигналов mTOR при болезни Альцгеймера» . Границы бионауки . 4 (1): 941–52. дои : 10.2741/s310 . ПМЦ 4111148 . ПМИД 22202101 .
- ^ Перейти обратно: а б Ан У.Л., Коуберн Р.Ф., Ли Л. и др. (август 2003 г.). «Повышающая регуляция фосфорилированной/активированной киназы S6 p70 и ее связь с нейрофибриллярной патологией при болезни Альцгеймера» . Американский журнал патологии . 163 (2): 591–607. дои : 10.1016/S0002-9440(10)63687-5 . ПМК 1868198 . ПМИД 12875979 .
- ^ Чжан Ф., Бехарри З.М., Харрис Т.Е. и др. (май 2009 г.). «Протеинкиназа PIM1 регулирует фосфорилирование PRAS40 и активность mTOR в клетках FDCP1». Биология и терапия рака . 8 (9): 846–53. дои : 10.4161/cbt.8.9.8210 . ПМИД 19276681 . S2CID 22153842 .
- ^ Ку Э.Х., Скваццо С.Л. (июль 1994 г.). «Доказательства того, что производство и высвобождение бета-амилоидного белка осуществляется эндоцитарным путем» . Журнал биологической химии . 269 (26): 17386–9. дои : 10.1016/S0021-9258(17)32449-3 . ПМИД 8021238 .
- ^ Перейти обратно: а б с Каккамо А., Маджумдер С., Ричардсон А. и др. (апрель 2010 г.). «Молекулярное взаимодействие между мишенью рапамицина (mTOR), бета-амилоида и тау у млекопитающих: влияние на когнитивные нарушения» . Журнал биологической химии . 285 (17): 13107–20. дои : 10.1074/jbc.M110.100420 . ПМК 2857107 . ПМИД 20178983 .
- ^ Лафай-Шебасье С., Паккалин М., Пейдж G и др. (июль 2005 г.). «Изменение передачи сигналов mTOR/p70S6k под воздействием Абета, а также в трансгенных моделях APP-PS1 и у пациентов с болезнью Альцгеймера» . Журнал нейрохимии . 94 (1): 215–25. дои : 10.1111/j.1471-4159.2005.03187.x . ПМИД 15953364 . S2CID 8464608 .
- ^ Перейти обратно: а б с д Каккамо А., Мальдонадо М.А., Маджумдер С. и др. (март 2011 г.). «Естественно секретируемый бета-амилоид увеличивает активность рапамицина (mTOR) у млекопитающих посредством механизма, опосредованного PRAS40» . Журнал биологической химии . 286 (11): 8924–32. дои : 10.1074/jbc.M110.180638 . ПМК 3058958 . ПМИД 21266573 .
- ^ Санчак Ю., Торин С.С., Петерсон Т.Р. и др. (март 2007 г.). «PRAS40 представляет собой регулируемый инсулином ингибитор протеинкиназы mTORC1» . Молекулярная клетка . 25 (6): 903–15. doi : 10.1016/j.molcel.2007.03.003 . ПМИД 17386266 .
- ^ Ван Л., Харрис Т.Э., Рот Р.А. и др. (июль 2007 г.). «PRAS40 регулирует активность киназы mTORC1, действуя как прямой ингибитор связывания субстрата» . Журнал биологической химии . 282 (27): 20036–44. дои : 10.1074/jbc.M702376200 . ПМИД 17510057 .
- ^ Пей Джей-Джей, Хьюгон Дж (декабрь 2008 г.). «mTOR-зависимая передача сигналов при болезни Альцгеймера» . Журнал клеточной и молекулярной медицины . 12 (6Б): 2525–32. дои : 10.1111/j.1582-4934.2008.00509.x . ПМЦ 3828871 . ПМИД 19210753 .
- ^ Меске В., Альберт Ф., Ом Т.Г. (январь 2008 г.). «Сочетание мишени рапамицина у млекопитающих с сигнальным путем фосфоинозитид-3-киназы регулирует фосфорилирование тау-киназы-3, зависимое от протеинфосфатазы 2А и гликогенсинтазы-3» . Журнал биологической химии . 283 (1): 100–9. дои : 10.1074/jbc.M704292200 . ПМИД 17971449 .
- ^ Янссенс В., Горис Дж. (февраль 2001 г.). «Протеинфосфатаза 2А: высокорегулируемое семейство серин/треонинфосфатаз, участвующих в росте клеток и передаче сигналов» . Биохимический журнал . 353 (Часть 3): 417–39. дои : 10.1042/0264-6021:3530417 . ПМЦ 1221586 . ПМИД 11171037 .
- ^ Морита Т., Собу К. (октябрь 2009 г.). «Спецификация нейрональной полярности, регулируемой локальной трансляцией CRMP2 и тау по пути mTOR-p70S6K» . Журнал биологической химии . 284 (40): 27734–45. дои : 10.1074/jbc.M109.008177 . ПМЦ 2785701 . ПМИД 19648118 .
- ^ Пуйгерманаль Э., Марсикано Г., Бускетс-Гарсия А. и др. (сентябрь 2009 г.). «Каннабиноидная модуляция долговременной памяти гиппокампа опосредуется передачей сигналов mTOR». Природная неврология . 12 (9): 1152–8. дои : 10.1038/nn.2369 . ПМИД 19648913 . S2CID 9584832 .
- ^ Тишмейер В., Шикник Х., Краус М. и др. (август 2003 г.). «Чувствительная к рапамицину передача сигналов в долгосрочной консолидации памяти, зависимой от слуховой коры». Европейский журнал неврологии . 18 (4): 942–50. дои : 10.1046/j.1460-9568.2003.02820.x . ПМИД 12925020 . S2CID 2780242 .
- ^ Хёффер Калифорния, Кланн Э (февраль 2010 г.). «Сигнализация mTOR: на перекрестке пластичности, памяти и болезней» . Тенденции в нейронауках . 33 (2): 67–75. дои : 10.1016/j.tins.2009.11.003 . ПМК 2821969 . ПМИД 19963289 .
- ^ Келлехер Р.Дж., Говиндараджан А., Юнг Х.И. и др. (февраль 2004 г.). «Трансляционный контроль посредством передачи сигналов MAPK в долгосрочной синаптической пластичности и памяти» . Клетка . 116 (3): 467–79. дои : 10.1016/S0092-8674(04)00115-1 . ПМИД 15016380 .
- ^ Энингер Д., Хан С., Шилянски С. и др. (август 2008 г.). «Обращение дефицита обучения в мышиной модели туберозного склероза Tsc2 +/-» . Природная медицина . 14 (8): 843–8. дои : 10.1038/nm1788 . ПМК 2664098 . ПМИД 18568033 .
- ^ Морено Дж.А., Рэдфорд Х., Перетти Д. и др. (май 2012 г.). «Устойчивая репрессия трансляции eIF2α-P опосредует прионную нейродегенерацию» . Природа . 485 (7399): 507–11. Бибкод : 2012Natur.485..507M . дои : 10.1038/nature11058 . ПМК 3378208 . ПМИД 22622579 .
- ^ Диас-Троя С., Перес-Перес М.Э., Флоренсио Ф.Дж. и др. (октябрь 2008 г.). «Роль TOR в регуляции аутофагии от дрожжей до растений и млекопитающих» . Аутофагия . 4 (7): 851–65. дои : 10.4161/авто.6555 . ПМИД 18670193 .
- ^ МакКрей Б.А., Тейлор Дж.П. (декабрь 2008 г.). «Роль аутофагии в возрастной нейродегенерации». Нейросигналы . 16 (1): 75–84. дои : 10.1159/000109761 . ПМИД 18097162 . S2CID 13591350 .
- ^ Недельский Н.Б., Тодд П.К., Тейлор Дж.П. (декабрь 2008 г.). «Аутофагия и система убиквитин-протеасома: участники нейропротекции» . Biochimica et Biophysica Acta (BBA) - Молекулярные основы болезней . 1782 (12): 691–9. дои : 10.1016/j.bbadis.2008.10.002 . ПМЦ 2621359 . ПМИД 18930136 .
- ^ Рубинштейн, округ Колумбия (октябрь 2006 г.). «Роль путей внутриклеточной деградации белков в нейродегенерации». Природа . 443 (7113): 780–6. Бибкод : 2006Natur.443..780R . дои : 10.1038/nature05291 . ПМИД 17051204 . S2CID 4411895 .
- ^ Оддо С. (апрель 2008 г.). «Убиквитин-протеасомная система при болезни Альцгеймера» . Журнал клеточной и молекулярной медицины . 12 (2): 363–73. дои : 10.1111/j.1582-4934.2008.00276.x . ПМЦ 3822529 . ПМИД 18266959 .
- ^ Ли X, Ли Х, Ли XJ (ноябрь 2008 г.). «Внутриклеточная деградация неправильно свернутых белков при полиглутаминовых нейродегенеративных заболеваниях» . Обзоры исследований мозга . 59 (1): 245–52. дои : 10.1016/j.brainresrev.2008.08.003 . ПМЦ 2577582 . ПМИД 18773920 .
- ^ Каккамо А., Маджумдер С., Денг Дж.Дж. и др. (октябрь 2009 г.). «Рапамицин спасает неправильную локализацию TDP-43 и связанную с ней низкомолекулярную нестабильность нейрофиламентов» . Журнал биологической химии . 284 (40): 27416–24. дои : 10.1074/jbc.M109.031278 . ПМЦ 2785671 . ПМИД 19651785 .
- ^ Равикумар Б., Вашер С., Бергер З. и др. (июнь 2004 г.). «Ингибирование mTOR вызывает аутофагию и снижает токсичность экспансии полиглутамина на моделях болезни Хантингтона на мухах и мышах» . Природная генетика . 36 (6): 585–95. дои : 10.1038/ng1362 . ПМИД 15146184 .
- ^ Рами А. (октябрь 2009 г.). «Обзор: аутофагия при нейродегенерации: пожарный и/или поджигатель?» . Невропатология и прикладная нейробиология . 35 (5): 449–61. дои : 10.1111/j.1365-2990.2009.01034.x . ПМИД 19555462 .
- ^ Фёлкль, Саймон и др. «Гиперактивный путь mTOR способствует лимфопролиферации и аномальной дифференцировке при аутоиммунном лимфопролиферативном синдроме». Кровь, Журнал Американского общества гематологии 128.2 (2016): 227-238. https://doi.org/10.1182/blood-2015-11-685024
- ^ Аренас, Дэниел Дж. и др. «Повышенная активация mTOR при идиопатической мультицентрической болезни Кастлемана». Кровь 135.19 (2020): 1673–1684. https://doi.org/10.1182/blood.2019002792
- ^ Эль-Салем, Моуна и др. «Конститутивная активация сигнального пути mTOR при посттрансплантационных лимфопролиферативных заболеваниях». Лабораторные исследования 87.1 (2007): 29-39. https://doi.org/10.1038/labinvest.3700494
- ^ Перейти обратно: а б с д Брук М.С., Уилкинсон Д.Д., Филлипс Б.Е. и др. (январь 2016 г.). «Гомеостаз и пластичность скелетных мышц в молодости и старении: влияние питания и физических упражнений» . Акта Физиологика . 216 (1): 15–41. дои : 10.1111/apha.12532 . ПМЦ 4843955 . ПМИД 26010896 .
- ^ Перейти обратно: а б Бриошь Т., Пагано А.Ф., Пи Дж. и др. (апрель 2016 г.). «Мышечное истощение и старение: экспериментальные модели, жировые инфильтраты и профилактика» (PDF) . Молекулярные аспекты медицины . 50 : 56–87. дои : 10.1016/j.mam.2016.04.006 . ПМИД 27106402 . S2CID 29717535 .
- ^ Драммонд М.Дж., Дрейер Х.К., Фрай К.С. и др. (апрель 2009 г.). «Пищевая и сократительная регуляция синтеза белка скелетных мышц человека и передачи сигналов mTORC1» . Журнал прикладной физиологии . 106 (4): 1374–84. doi : 10.1152/japplphysicalol.91397.2008 . ПМК 2698645 . ПМИД 19150856 .
- ^ Сальто Р., Вильчес Х.Д., Хирон, доктор медицинских наук и др. (2015). «β-гидрокси-β-метилбутират (HMB) способствует росту нейритов в клетках Neuro2a» . ПЛОС ОДИН . 10 (8): e0135614. Бибкод : 2015PLoSO..1035614S . дои : 10.1371/journal.pone.0135614 . ПМЦ 4534402 . ПМИД 26267903 .
- ^ Кугиас Д.Г., Нолан С.О., Косс В.А. и др. (апрель 2016 г.). «Бета-гидрокси-бета-метилбутират смягчает эффекты старения в дендритном дереве пирамидных нейронов медиальной префронтальной коры как самцов, так и самок крыс». Нейробиология старения . 40 : 78–85. doi : 10.1016/j.neurobiolaging.2016.01.004 . ПМИД 26973106 . S2CID 3953100 .
- ^ Перейти обратно: а б Филлипс С.М. (май 2014 г.). «Краткий обзор важнейших процессов мышечной гипертрофии, вызванной физическими упражнениями» . Спорт Мед . 44 (Приложение 1): S71–S77. дои : 10.1007/s40279-014-0152-3 . ПМК 4008813 . ПМИД 24791918 .
- ^ Перейти обратно: а б с д и ж г Цзя Дж., Абуду Ю.П., Клод-Топен А. и др. (апрель 2018 г.). «Галектины контролируют mTOR в ответ на повреждение эндомембраны» . Молекулярная клетка . 70 (1): 120–135.e8. дои : 10.1016/j.molcel.2018.03.009 . ПМК 5911935 . ПМИД 29625033 .
- ^ Нода Т., Осуми Ю. (февраль 1998 г.). «Tor, гомолог фосфатидилинозитолкиназы, контролирует аутофагию у дрожжей» . Журнал биологической химии . 273 (7): 3963–6. дои : 10.1074/jbc.273.7.3963 . ПМИД 9461583 .
- ^ Дубулоз Ф., Делош О., Ванке В. и др. (июль 2005 г.). «Белковые комплексы TOR и EGO управляют микроаутофагией у дрожжей» . Молекулярная клетка . 19 (1): 15–26. doi : 10.1016/j.molcel.2005.05.020 . ПМИД 15989961 .
- ^ Перейти обратно: а б Гэнли И.Г., Лам дю Х., Ван Дж. и др. (май 2009 г.). «Комплекс ULK1.ATG13.FIP200 опосредует передачу сигналов mTOR и необходим для аутофагии» . Журнал биологической химии . 284 (18): 12297–305. дои : 10.1074/jbc.M900573200 . ПМЦ 2673298 . ПМИД 19258318 .
- ^ Перейти обратно: а б Юнг Ч., Джун CB, Ро Ш. и др. (апрель 2009 г.). «Комплексы ULK-Atg13-FIP200 опосредуют передачу сигналов mTOR в механизмы аутофагии» . Молекулярная биология клетки . 20 (7): 1992–2003. дои : 10.1091/mbc.e08-12-1249 . ПМК 2663920 . ПМИД 19225151 .
- ^ Перейти обратно: а б Хосокава Н., Хара Т., Кайдзука Т. и др. (апрель 2009 г.). «Питательно-зависимая ассоциация mTORC1 с комплексом ULK1-Atg13-FIP200, необходимая для аутофагии» . Молекулярная биология клетки . 20 (7): 1981–91. дои : 10.1091/mbc.e08-12-1248 . ПМК 2663915 . ПМИД 19211835 .
- ^ Хасегава Дж., Маэдзима И., Ивамото Р. и др. (март 2015 г.). «Селективная аутофагия: лизофагия» . Методы . 75 : 128–32. дои : 10.1016/j.ymeth.2014.12.014 . ПМИД 25542097 .
- ^ Фрайберг М., Элазар З. (октябрь 2016 г.). «Комплекс TRIM16-галактин3 опосредует аутофагию поврежденных эндомембран» . Развивающая клетка . 39 (1): 1–2. дои : 10.1016/j.devcel.2016.09.025 . ПМИД 27728777 .
- ^ Перейти обратно: а б с Чаухан С., Кумар С., Джайн А. и др. (октябрь 2016 г.). «TRIMs и галектины сотрудничают во всем мире, а TRIM16 и галектин-3 совместно направляют аутофагию в гомеостазе повреждения эндомембраны» . Развивающая клетка . 39 (1): 13–27. дои : 10.1016/j.devcel.2016.08.003 . ПМК 5104201 . ПМИД 27693506 .
- ^ Нисимура Т., Кайдзука Т., Кэдвелл К. и др. (март 2013 г.). «FIP200 регулирует нацеливание Atg16L1 на изолирующую мембрану» . Отчеты ЭМБО . 14 (3): 284–91. дои : 10.1038/embor.2013.6 . ПМК 3589088 . ПМИД 23392225 .
- ^ Гаммо Н., Флори О., Оверхольцер М. и др. (февраль 2013 г.). «Взаимодействие между FIP200 и ATG16L1 различает комплексно-зависимую и независимую аутофагию ULK1» . Структурная и молекулярная биология природы . 20 (2): 144–9. дои : 10.1038/nsmb.2475 . ПМК 3565010 . ПМИД 23262492 .
- ^ Перейти обратно: а б с Фудзита Н., Морита Э., Ито Т. и др. (октябрь 2013 г.). «Привлечение аутофагического механизма в эндосомы во время инфекции опосредовано убиквитином» . Журнал клеточной биологии . 203 (1): 115–28. дои : 10.1083/jcb.201304188 . ПМЦ 3798248 . ПМИД 24100292 .
- ^ Пак Дж.М., Юнг Ч.Х., Со М. и др. (03.03.2016). «Комплекс ULK1 опосредует передачу сигналов MTORC1 механизму инициации аутофагии посредством связывания и фосфорилирования ATG14» . Аутофагия . 12 (3): 547–64. дои : 10.1080/15548627.2016.1140293 . ПМК 4835982 . ПМИД 27046250 .
- ^ Дули Х.К., Рази М., Полсон Х.Э. и др. (июль 2014 г.). «WIPI2 связывает конъюгацию LC3 с PI3P, образованием аутофагосом и выведением патогенов путем рекрутирования Atg12-5-16L1» . Молекулярная клетка . 55 (2): 238–52. doi : 10.1016/j.molcel.2014.05.021 . ПМК 4104028 . ПМИД 24954904 .
- ^ Ким Дж., Кунду М., Виоллет Б. и др. (февраль 2011 г.). «AMPK и mTOR регулируют аутофагию посредством прямого фосфорилирования Ulk1» . Природная клеточная биология . 13 (2): 132–41. дои : 10.1038/ncb2152 . ПМЦ 3987946 . ПМИД 21258367 .
- ^ Ким Дж., Ким Ю.К., Фанг С. и др. (январь 2013 г.). «Дифференциальная регуляция отдельных комплексов Vps34 с помощью AMPK при пищевом стрессе и аутофагии» . Клетка . 152 (1–2): 290–303. дои : 10.1016/j.cell.2012.12.016 . ПМЦ 3587159 . ПМИД 23332761 .
- ^ Гвинн Д.М., Шакелфорд Д.Б., Иган Д.Ф. и др. (апрель 2008 г.). «Фосфорилирование AMPK хищника опосредует метаболическую контрольную точку» . Молекулярная клетка . 30 (2): 214–26. doi : 10.1016/j.molcel.2008.03.003 . ПМК 2674027 . ПМИД 18439900 .
- ^ Иноки К., Чжу Т., Гуань К.Л. (ноябрь 2003 г.). «TSC2 опосредует клеточную энергетическую реакцию, контролируя рост и выживание клеток» . Клетка . 115 (5): 577–90. дои : 10.1016/S0092-8674(03)00929-2 . ПМИД 14651849 .
- ^ Ёсида Ю., Ясуда С., Фудзита Т. и др. (август 2017 г.). «FBXO27 направляет поврежденные лизосомы на аутофагию» . Труды Национальной академии наук Соединенных Штатов Америки . 114 (32): 8574–8579. дои : 10.1073/pnas.1702615114 . ПМК 5559013 . ПМИД 28743755 .
- ^ Хименес С.А., Кронин П.М., Кениг А.С. и др. (15 февраля 2012 г.). Варга Дж., Талавера Ф., Голдберг Э., Мехабер А.Дж., Даймонд Х.С. (ред.). «Склеродермия» . Справочник Медскейп . ВебМД . Проверено 5 марта 2014 г.
- ^ Хадж-Али Р.А. (июнь 2013 г.). «Системный склероз» . Руководство Merck Professional . Мерк Шарп и Доум Корп . Проверено 5 марта 2014 г.
- ^ «Мишень ингибиторов рапамицина (mTOR) у млекопитающих при солидных опухолях» . Фармацевтический журнал . Проверено 18 октября 2018 г.
- ^ Фавр С., Кремер Г., Раймонд Э. (август 2006 г.). «Современная разработка ингибиторов mTOR в качестве противораковых средств». Обзоры природы. Открытие наркотиков . 5 (8): 671–88. дои : 10.1038/nrd2062 . ПМИД 16883305 . S2CID 27952376 .
- ^ Хэсти П. (февраль 2010 г.). «Рапамицин: лекарство от всех недугов» . Журнал молекулярно-клеточной биологии . 2 (1): 17–9. дои : 10.1093/jmcb/mjp033 . ПМИД 19805415 .
- ^ Бове Х., Мартинес-Висенте М., Вила М. (август 2011 г.). «Борьба с нейродегенерацией с помощью рапамицина: механистические идеи». Обзоры природы. Нейронаука . 12 (8): 437–52. дои : 10.1038/nrn3068 . ПМИД 21772323 . S2CID 205506774 .
- ^ Манник Дж.Б., Моррис М., Hockey HP и др. (июль 2018 г.). «Ингибирование TORC1 усиливает иммунную функцию и снижает риск инфекций у пожилых людей» . Наука трансляционной медицины . 10 (449): eaaq1564. doi : 10.1126/scitranslmed.aaq1564 . ПМИД 29997249 .
- ^ Эштрела Дж.М., Ортега А., Мена С. и др. (2013). «Птеростильбен: Биомедицинское применение». Критические обзоры клинических лабораторных наук . 50 (3): 65–78. дои : 10.3109/10408363.2013.805182 . ПМИД 23808710 . S2CID 45618964 .
- ^ МакКубри Дж.А., Лертпирияпонг К., Стилман Л.С. и др. (июнь 2017 г.). «Влияние ресвератрола, куркумина, берберина и других нутрицевтиков на старение, развитие рака, раковые стволовые клетки и микроРНК» . Старение . 9 (6): 1477–1536. дои : 10.18632/aging.101250 . ПМК 5509453 . ПМИД 28611316 .
- ^ Малаволта М., Браччи М., Сантарелли Л. и др. (2018). «Индукторы старения, токсичные соединения и сенолитики: многогранность фитохимических веществ, активирующих Nrf2, в адъювантной терапии рака» . Медиаторы воспаления . 2018 : 4159013. doi : 10.1155/2018/4159013 . ПМЦ 5829354 . ПМИД 29618945 .
- ^ Гомес-Линтон Д.Р., Алавес С., Аларкон-Агилар А. и др. (октябрь 2019 г.). «Некоторые природные соединения, которые увеличивают продолжительность жизни и устойчивость к стрессу у модельных организмов старения». Биогеронтология . 20 (5): 583–603. дои : 10.1007/s10522-019-09817-2 . ПМИД 31187283 . S2CID 184483900 .
- ^ Ли В., Цинь Л., Фэн Р. и др. (июль 2019 г.). «Новые сенолитические агенты, полученные из натуральных продуктов». Механизмы старения и развития . 181 : 1–6. дои : 10.1016/j.mad.2019.05.001 . ПМИД 31077707 . S2CID 147704626 .
- ^ «Белковые интеракторы mTOR» . Справочная база данных по белкам человека . Университет Джонса Хопкинса и Институт биоинформатики. Архивировано из оригинала 28 июня 2015 г. Проверено 6 декабря 2010 г.
- ^ Кумар В., Сабатини Д., Пандей П. и др. (апрель 2000 г.). «Регуляция рапамицина и FKBP-мишени 1/мишени рапамицина для млекопитающих и кэп-зависимой инициации трансляции протеин-тирозинкиназой c-Abl» . Журнал биологической химии . 275 (15): 10779–87. дои : 10.1074/jbc.275.15.10779 . ПМИД 10753870 .
- ^ Секулич А., Хадсон С.С., Хомм Дж.Л. и др. (июль 2000 г.). «Прямая связь между сигнальным путем фосфоинозитид-3-киназы-АКТ и мишенью рапамицина у млекопитающих в митоген-стимулированных и трансформированных клетках». Исследования рака . 60 (13): 3504–13. ПМИД 10910062 .
- ^ Ченг С.В., Фрайер Л.Г., Карлинг Д. и др. (апрель 2004 г.). «Thr2446 является новой мишенью для млекопитающих сайта фосфорилирования рапамицина (mTOR), регулируемого статусом питательных веществ» . Журнал биологической химии . 279 (16): 15719–22. дои : 10.1074/jbc.C300534200 . ПМИД 14970221 .
- ^ Чой Дж.Х., Бертрам П.Г., Дренан Р. и др. (октябрь 2002 г.). «Белок, ассоциированный с FKBP12-рапамицином (FRAP), представляет собой киназу CLIP-170» . Отчеты ЭМБО . 3 (10): 988–94. дои : 10.1093/embo-reports/kvf197 . ПМЦ 1307618 . ПМИД 12231510 .
- ^ Харрис Т.Э., Чи А., Шабановиц Дж. и др. (апрель 2006 г.). «mTOR-зависимая стимуляция ассоциации eIF4G и eIF3 инсулином» . Журнал ЭМБО . 25 (8): 1659–68. дои : 10.1038/sj.emboj.7601047 . ПМЦ 1440840 . ПМИД 16541103 .
- ^ Перейти обратно: а б Шальм С.С., Фингар Д.К., Сабатини Д.М. и др. (май 2003 г.). «Связывание хищника, опосредованное мотивом TOS, регулирует многосайтовое фосфорилирование и функцию 4E-BP1» . Современная биология . 13 (10): 797–806. Бибкод : 2003CBio...13..797S . дои : 10.1016/S0960-9822(03)00329-4 . ПМИД 12747827 .
- ^ Перейти обратно: а б с Хара К., Маруки Й., Лонг Икс и др. (июль 2002 г.). «Раптор, связывающий партнер мишени рапамицина (TOR), опосредует действие TOR» . Клетка . 110 (2): 177–89. дои : 10.1016/S0092-8674(02)00833-4 . ПМИД 12150926 .
- ^ Перейти обратно: а б Ван Л., Роудс С.Дж., Лоуренс Дж.С. (август 2006 г.). «Активация инсулином мишени рапамицина (mTOR) млекопитающих связана со стимуляцией связывания 4EBP1 с димерным комплексом mTOR 1» . Журнал биологической химии . 281 (34): 24293–303. дои : 10.1074/jbc.M603566200 . ПМИД 16798736 .
- ^ Перейти обратно: а б с Лонг X, Лин Ю, Ортис-Вега С и др. (апрель 2005 г.). «Rheb связывает и регулирует киназу mTOR» . Современная биология . 15 (8): 702–13. Бибкод : 2005CBio...15..702L . дои : 10.1016/j.cub.2005.02.053 . ПМИД 15854902 .
- ^ Перейти обратно: а б Такахаши Т., Хара К., Иноуэ Х. и др. (сентябрь 2000 г.). «Карбоксильная область, консервативная среди киназ, связанных с фосфоинозитидкиназой, необходима для функции mTOR in vivo и in vitro». Гены в клетки . 5 (9): 765–75. дои : 10.1046/j.1365-2443.2000.00365.x . ПМИД 10971657 . S2CID 39048740 .
- ^ Перейти обратно: а б Бернетт П.Е., Барроу Р.К., Коэн Н.А. и др. (февраль 1998 г.). «RAFT1 фосфорилирование регуляторов трансляции p70 S6 киназы и 4E-BP1» . Труды Национальной академии наук Соединенных Штатов Америки . 95 (4): 1432–7. Бибкод : 1998PNAS...95.1432B . дои : 10.1073/pnas.95.4.1432 . ЧВК 19032 . ПМИД 9465032 .
- ^ Ван X, Бегнет А, Мураками М и др. (апрель 2005 г.). «Различные сигнальные события, расположенные ниже mTOR, взаимодействуют, опосредуя эффекты аминокислот и инсулина на белки, связывающие фактор инициации 4E» . Молекулярная и клеточная биология . 25 (7): 2558–72. дои : 10.1128/MCB.25.7.2558-2572.2005 . ПМК 1061630 . ПМИД 15767663 .
- ^ Чой Дж., Чен Дж., Шрайбер С.Л. и др. (июль 1996 г.). «Структура комплекса FKBP12-рапамицин, взаимодействующего со связывающим доменом FRAP человека». Наука . 273 (5272): 239–42. Бибкод : 1996Sci...273..239C . дои : 10.1126/science.273.5272.239 . ПМИД 8662507 . S2CID 27706675 .
- ^ Люкер К.Е., Смит М.К., Люкер Г.Д. и др. (август 2004 г.). «Кинетика регулируемых белок-белковых взаимодействий, выявленная с помощью визуализации комплементации люциферазы светлячков в клетках и живых животных» . Труды Национальной академии наук Соединенных Штатов Америки . 101 (33): 12288–93. Бибкод : 2004PNAS..10112288L . дои : 10.1073/pnas.0404041101 . ПМК 514471 . ПМИД 15284440 .
- ^ Банашински Л.А., Лю К.В., Уэндлесс Т.Дж. (апрель 2005 г.). «Характеристика тройного комплекса FKBP.рапамицин.FRB». Журнал Американского химического общества . 127 (13): 4715–21. дои : 10.1021/ja043277y . ПМИД 15796538 .
- ^ Сейбрс С.Дж., Мартин М.М., Брунн Г.Дж. и др. (январь 1995 г.). «Выделение белка-мишени комплекса FKBP12-рапамицин в клетках млекопитающих» . Журнал биологической химии . 270 (2): 815–22. дои : 10.1074/jbc.270.2.815 . ПМИД 7822316 .
- ^ Сабатини Д.М., Барроу Р.К., Блэкшоу С. и др. (май 1999 г.). «Взаимодействие RAFT1 с гефирином, необходимое для передачи сигналов, чувствительных к рапамицину». Наука . 284 (5417): 1161–4. Бибкод : 1999Sci...284.1161S . дои : 10.1126/science.284.5417.1161 . ПМИД 10325225 .
- ^ Ха Ш., Ким Д.Х., Ким И.С. и др. (декабрь 2006 г.). «PLD2 образует функциональный комплекс с mTOR/raptor для передачи митогенных сигналов». Сотовая сигнализация . 18 (12): 2283–91. doi : 10.1016/j.cellsig.2006.05.021 . ПМИД 16837165 .
- ^ Бюргер С., ДеВрис Б., Стамболик В. (июнь 2006 г.). «Локализация Rheb в эндомембране имеет решающее значение для его сигнальной функции». Связь с биохимическими и биофизическими исследованиями . 344 (3): 869–80. дои : 10.1016/j.bbrc.2006.03.220 . ПМИД 16631613 .
- ^ Перейти обратно: а б Хасинто Э., Факкинетти В., Лю Д. и др. (октябрь 2006 г.). «SIN1/MIP1 поддерживает целостность комплекса rictor-mTOR и регулирует фосфорилирование Akt и субстратную специфичность» . Клетка . 127 (1): 125–37. дои : 10.1016/j.cell.2006.08.033 . ПМИД 16962653 .
- ^ МакМахон Л.П., Юэ В., Сантэн Р.Дж. и др. (январь 2005 г.). «Фарнезилтиосалициловая кислота ингибирует активность рапамицина (mTOR) у млекопитающих как в клетках, так и in vitro, способствуя диссоциации комплекса mTOR-раптор» . Молекулярная эндокринология . 19 (1): 175–83. дои : 10.1210/me.2004-0305 . ПМИД 15459249 .
- ^ Осиро Н., Ёшино К., Хидаят С. и др. (апрель 2004 г.). «Диссоциация раптора от mTOR представляет собой механизм индуцированного рапамицином ингибирования функции mTOR». Гены в клетки . 9 (4): 359–66. дои : 10.1111/j.1356-9597.2004.00727.x . hdl : 20.500.14094/D1002969 . ПМИД 15066126 . S2CID 24814691 .
- ^ Каваи С., Энзан Х., Хаяши Ю. и др. (июль 2003 г.). «Винкулин: новый маркер покоящихся и активированных звездчатых клеток печени в печени человека и крысы». Архив Вирхова . 443 (1): 78–86. дои : 10.1007/s00428-003-0804-4 . ПМИД 12719976 . S2CID 21552704 .
- ^ Чой К.М., МакМахон Л.П., Лоуренс Дж.К. (май 2003 г.). «Два мотива в репрессоре трансляции PHAS-I необходимы для эффективного фосфорилирования рапамицина мишенью млекопитающих и для распознавания хищником» . Журнал биологической химии . 278 (22): 19667–73. дои : 10.1074/jbc.M301142200 . ПМИД 12665511 .
- ^ Перейти обратно: а б Нодзима Х., Токунага С., Эгучи С. и др. (май 2003 г.). «Мишень партнера рапамицина (mTOR) у млекопитающих, раптор, связывает субстраты mTOR киназу p70 S6 и 4E-BP1 посредством их сигнального мотива TOR (TOS)» . Журнал биологической химии . 278 (18): 15461–4. дои : 10.1074/jbc.C200665200 . ПМИД 12604610 .
- ^ Перейти обратно: а б Сарбасов Д.Д., Али С.М., Сенгупта С. и др. (апрель 2006 г.). «Длительное лечение рапамицином ингибирует сборку mTORC2 и Akt/PKB» . Молекулярная клетка . 22 (2): 159–68. doi : 10.1016/j.molcel.2006.03.029 . ПМИД 16603397 .
- ^ Цацос А., Кандрор К.В. (январь 2006 г.). «Питательные вещества подавляют передачу сигналов фосфатидилинозитол-3-киназы/Akt посредством зависимого от хищника mTOR-опосредованного фосфорилирования субстрата 1 инсулинового рецептора» . Молекулярная и клеточная биология . 26 (1): 63–76. дои : 10.1128/MCB.26.1.63-76.2006 . ПМЦ 1317643 . ПМИД 16354680 .
- ^ Перейти обратно: а б с Сарбасов Д.Д., Сабатини Д.М. (ноябрь 2005 г.). «Окислительно-восстановительная регуляция чувствительного к питательным веществам пути и комплекса раптор-mTOR» . Журнал биологической химии . 280 (47): 39505–9. дои : 10.1074/jbc.M506096200 . ПМИД 16183647 .
- ^ Перейти обратно: а б Ян К., Иноки К., Икеноуэ Т. и др. (октябрь 2006 г.). «Идентификация Sin1 как важного компонента TORC2, необходимого для образования комплекса и активности киназы» . Гены и развитие . 20 (20): 2820–32. дои : 10.1101/gad.1461206 . ПМК 1619946 . ПМИД 17043309 .
- ^ Кумар В., Панди П., Сабатини Д. и др. (март 2000 г.). «Функциональное взаимодействие между RAFT1/FRAP/mTOR и протеинкиназой cdelta в регуляции кэп-зависимой инициации трансляции» . Журнал ЭМБО . 19 (5): 1087–97. дои : 10.1093/emboj/19.5.1087 . ПМК 305647 . ПМИД 10698949 .
- ^ Лонг X, Ортис-Вега С, Лин Ю и др. (июнь 2005 г.). «Связывание Rheb с мишенью рапамицина (mTOR) у млекопитающих регулируется достаточностью аминокислот» . Журнал биологической химии . 280 (25): 23433–6. дои : 10.1074/jbc.C500169200 . ПМИД 15878852 .
- ^ Смит Э.М., Финн С.Г., Ти А.Р. и др. (май 2005 г.). «Белок туберозного склероза TSC2 не требуется для регуляции мишени рапамицина у млекопитающих с помощью аминокислот и определенных клеточных стрессов» . Журнал биологической химии . 280 (19): 18717–27. дои : 10.1074/jbc.M414499200 . ПМИД 15772076 .
- ^ Бернарди Р., Герна И., Джин Д. и др. (август 2006 г.). «PML ингибирует трансляцию HIF-1альфа и неоангиогенез посредством репрессии mTOR». Природа . 442 (7104): 779–85. Бибкод : 2006Natur.442..779B . дои : 10.1038/nature05029 . ПМИД 16915281 . S2CID 4427427 .
- ^ Сайто М., Пуллен Н., Бреннан П. и др. (май 2002 г.). «Регуляция активированного варианта киназы S6 1 обнаруживает новую мишень сайта фосфорилирования рапамицина у млекопитающих» . Журнал биологической химии . 277 (22): 20104–12. дои : 10.1074/jbc.M201745200 . ПМИД 11914378 .
- ^ Чан Г.Г., Авраам RT (июль 2005 г.). «Фосфорилирование мишени рапамицина (mTOR) млекопитающих по Ser-2448 опосредовано киназой p70S6» . Журнал биологической химии . 280 (27): 25485–90. дои : 10.1074/jbc.M501707200 . ПМИД 15899889 .
- ^ Хольц МК, Бленис Дж (июль 2005 г.). «Идентификация киназы S6 1 как новой мишени фосфорилирующей киназы рапамицина (mTOR) у млекопитающих» . Журнал биологической химии . 280 (28): 26089–93. дои : 10.1074/jbc.M504045200 . ПМИД 15905173 .
- ^ Исотани С., Хара К., Токунага С. и др. (ноябрь 1999 г.). «Иммуноочищенная мишень рапамицина для млекопитающих фосфорилирует и активирует альфа-киназу p70 S6 in vitro» . Журнал биологической химии . 274 (48): 34493–8. дои : 10.1074/jbc.274.48.34493 . hdl : 20.500.14094/D1002182 . ПМИД 10567431 .
- ^ Торал-Барза Л., Чжан В.Г., Ламисон С. и др. (июнь 2005 г.). «Характеристика клонированной полноразмерной и усеченной человеческой мишени рапамицина: активность, специфичность и ингибирование ферментов, изученные с помощью анализа высокой емкости». Связь с биохимическими и биофизическими исследованиями . 332 (1): 304–10. дои : 10.1016/j.bbrc.2005.04.117 . ПМИД 15896331 .
- ^ Али С.М., Сабатини Д.М. (май 2005 г.). «Структура киназы 1 S6 определяет, фосфорилирует ли раптор-mTOR или rictor-mTOR свой сайт гидрофобного мотива» . Журнал биологической химии . 280 (20): 19445–8. дои : 10.1074/jbc.C500125200 . ПМИД 15809305 .
- ^ Эдингер А.Л., Линардик К.М., Чан Г.Г. и др. (декабрь 2003 г.). «Дифференциальные эффекты рапамицина на мишени сигнальных функций рапамицина в клетках млекопитающих». Исследования рака . 63 (23): 8451–60. ПМИД 14679009 .
- ^ Леоне М., Кроуэлл К.Дж., Чен Дж. и др. (август 2006 г.). «Домен FRB mTOR: структура раствора ЯМР и конструкция ингибитора». Биохимия . 45 (34): 10294–302. дои : 10.1021/bi060976+ . ПМИД 16922504 .
- ^ Кристоф А.С., Маркс-Кончалик Дж., Биллингс Э. и др. (сентябрь 2003 г.). «Стимуляция преобразователя сигнала и активатора транскрипции-1 (STAT1)-зависимой транскрипции гена липополисахаридом и гамма-интерфероном регулируется мишенью рапамицина у млекопитающих» . Журнал биологической химии . 278 (36): 33637–44. дои : 10.1074/jbc.M301053200 . ПМИД 12807916 .
- ^ Ёкогами К., Вакисака С., Авруч Дж. и др. (январь 2000 г.). «Фосфорилирование серина и максимальная активация STAT3 во время передачи сигналов CNTF опосредуются mTOR-мишенью рапамицина» . Современная биология . 10 (1): 47–50. Бибкод : 2000CBio...10...47Y . дои : 10.1016/S0960-9822(99)00268-7 . ПМИД 10660304 .
- ^ Кусаба Х., Гош П., Дерин Р. и др. (январь 2005 г.). «Индуцированная интерлейкином-12 продукция гамма-интерферона Т-клетками периферической крови человека регулируется мишенью рапамицина (mTOR) у млекопитающих» . Журнал биологической химии . 280 (2): 1037–43. дои : 10.1074/jbc.M405204200 . ПМИД 15522880 .
- ^ Цан С., Чжоу Ю., Наварро Б. и др. (февраль 2013 г.). «mTOR регулирует лизосомальные АТФ-чувствительные двухпоровые Na(+) каналы для адаптации к метаболическому состоянию» . Клетка . 152 (4): 778–90. дои : 10.1016/j.cell.2013.01.023 . ПМК 3908667 . ПМИД 23394946 .
- ^ Ву С., Михайлов А., Калло-Хосейн Х. и др. (январь 2002 г.). «Характеристика убикилина 1, белка, взаимодействующего с mTOR» . Biochimica et Biophysical Acta (BBA) - Исследования молекулярных клеток . 1542 (1–3): 41–56. дои : 10.1016/S0167-4889(01)00164-1 . ПМИД 11853878 .
Дальнейшее чтение
[ редактировать ]- Сакстон Р.А., Сабатини Д.М. (март 2017 г.). «Передача сигналов mTOR в росте, обмене веществ и заболеваниях» . Клетка . 168 (6): 960–976. дои : 10.1016/j.cell.2017.02.004 . ПМЦ 5394987 . ПМИД 28283069 .
Внешние ссылки
[ редактировать ]- mTOR + белок в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)
- «Сигнальный путь mTOR в базе данных взаимодействия путей» . Национальный институт рака. Архивировано из оригинала 18 марта 2013 г. Проверено 18 октября 2015 г.
- Обзор всей структурной информации, доступной в PDB для UniProt : P42345 (серин/треонин-протеинкиназа mTOR) в PDBe-KB .
галерея PDB |
|---|