EIF4E
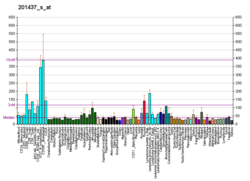
| EIF4E | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Идентификаторы | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | EIF4E , эукариотический фактор инициации трансляции 4E, AUTS19, CBP, EIF4E1, EIF4EL1, EIF4F, eIF-4E | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | Опустить : 133440 ; МГИ : 95305 ; Гомологен : 123817 ; Генные карты : EIF4E ; OMA : EIF4E — ортологи | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||








Эукариотический фактор инициации трансляции 4E , также известный как eIF4E , представляет собой белок , который у человека кодируется EIF4E геном . [ 5 ] [ 6 ]
Структура и функции
[ редактировать ]Большинство эукариотических клеточных мРНК блокируются на своих 5'-концах структурой с 7- метилгуанозина пятью штрихами , m7GpppX (где X — любой нуклеотид). Эта структура участвует в нескольких клеточных процессах, включая повышенную эффективность трансляции, сплайсинг, стабильность мРНК и ядерный экспорт РНК. eIF4E представляет собой эукариотический фактор инициации трансляции, участвующий в направлении рибосом к кэп-структуре мРНК, а также в других этапах метаболизма РНК, требующих кэп-связывания. Это полипептид массой 24 кДа , который существует как в свободной форме, так и в составе eIF4F . преинициаторного комплекса [ 7 ] Многим клеточным мРНК требуется eIF4E для трансляции в белок. Полипептид eIF4E некоторые считают лимитирующим компонентом эукариотического аппарата трансляции и участвует в стадии связывания мРНК-рибосомы при синтезе эукариотического белка.
Другие субъединицы eIF4F представляют собой полипептид массой 47 кДа, называемый eIF4A . [ 8 ] который обладает АТФазной и РНК- хеликазной активностью, а также каркасным полипептидом массой 220 кДа, eIF4G . [ 9 ] [ 10 ] [ 11 ]
Некоторые вирусы разрезают eIF4G таким образом, что сайт связывания eIF4E удаляется, и вирус может транслировать свои белки без eIF4E. Кроме того, некоторые клеточные белки, наиболее примечательными из которых являются белки теплового шока, не нуждаются в eIF4E для трансляции. И вирусы, и клеточные белки достигают этого посредством внутреннего сайта входа в рибосому в РНК или с помощью других механизмов трансляции РНК, например, через eIF3d. [ 12 ] [ 13 ]
eIF4E играет роль вне трансляции, и другие кэп-связывающие белки могут участвовать в кэп-зависимой трансляции независимым от eIF4E образом, включая такие факторы, как eIF3D, eIF3I, PARN, ядерный кэп-связывающий комплекс CBC. [ 14 ] [ 15 ] [ 12 ] [ 16 ] [ 17 ] [ 13 ] [ 18 ] Многие из них, по-видимому, зависят как от специфических особенностей транскриптов, так и от клеточного контекста.
eIF4E обнаружен в ядре многих типов клеток млекопитающих, а также у других видов, включая дрожжи, дрозофилу и человека. [ 19 ] [ 20 ] [ 21 ] [ 22 ] [ 23 ] [ 24 ] [ 25 ] [ 26 ] eIF4E обнаруживается в ядерных тельцах, подмножество которых колокализуется с ядерными тельцами PML, а eIF4E дополнительно обнаруживается диффузно в частях нуклеоплазмы млекопитающих. [ 23 ] [ 21 ] [ 24 ] [ 25 ] [ 27 ] [ 28 ] [ 26 ] В ядре eIF4E играет четко определенную роль в экспорте выбранных РНК, что способствует его онкогенным фенотипам. [ 29 ] [ 21 ] [ 24 ] [ 25 ] [ 27 ] [ 28 ] [ 30 ] [ 31 ] [ 32 ] [ 33 ] Это зависит от способности eIF4E связывать m 7 G-кэп РНК и наличие 50-нуклеотидного чувствительного элемента eIF4E (4ESE) в 3'UTR чувствительных транскриптов; хотя другие элементы также могут играть роль. Эта форма экспорта основана на пути CRM1/XPO1. [ 21 ] [ 24 ] [ 27 ] [ 34 ] [ 35 ] [ 26 ] Было показано, что ядерный eIF4E играет и другие роли в процессинге РНК, в том числе в м 7 G-кэпирование, альтернативное полиаденилирование и сплайсинг. [ 36 ] [ 37 ] [ 38 ]
Увеличение ядерного накопления eIF4E, а также увеличение eIF4E-зависимого экспорта РНК, м 7 G-кэпирование и сплайсинг выбранных транскриптов характерны для образцов пациентов с ОМЛ с высоким содержанием eIF4E. [ 25 ] [ 30 ] [ 38 ] [ 37 ] РНК выбираются на основе кодов USER или цис-действующих элементов внутри их РНК для определенных уровней процессинга РНК; таким образом, не все транскрипты чувствительны ко всем уровням регуляции (включая трансляцию). [ 35 ] [ 39 ] [ 18 ] Для своей функции экспорта РНК eIF4E напрямую связывается с богатым лейцином белком пентатрикопептидного повтора (LRPPRC), который напрямую связывается с дорсальной поверхностью eIF4E и одновременно с РНК 4ESE, тем самым действуя в качестве платформы для сборки комплекса экспорта РНК. [ 26 ] [ 35 ] Текущая модель заключается в том, что LRPPRC связывается с CRM1/XPO1, захватывая ядерную пору и транспортируя РНК 4ESE в цитоплазму. [ 28 ] [ 26 ] [ 35 ] В целом, ядерные функции eIF4E могут оказывать мощное воздействие на протеом, позволяя eIF4E как перезаписывать сообщение, так и увеличивать выработку белков за счет повышенного накопления в цитоплазме из-за увеличения экспорта, а также увеличения количества белков. в некоторых случаях рибосомы на транскрипт. Его многочисленные роли в процессинге РНК требуют ассоциации с РНК через кэп m7G, и, таким образом, eIF4E можно считать белком кэп-шаперона.
Регулирование
[ редактировать ]Поскольку eIF4E является фактором инициации, который относительно невелик, eIF4E можно контролировать на нескольких уровнях. [ 40 ] [ 18 ] Регуляция eIF4E может достигаться на уровнях транскрипции, стабильного фосфорилирования РНК, субклеточной локализации и белков-партнеров. [ 41 ]
а. Регуляция eIF4E посредством экспрессии генов и стабильности РНК
Механизмы, ответственные за регуляцию транскрипции eIF4E, до конца не изучены. Однако в нескольких отчетах предполагается корреляция между уровнями myc и уровнями мРНК eIF4E во время клеточного цикла. [ 42 ] Основа этой связи была дополнительно установлена путем характеристики двух сайтов связывания myc (повторов E-бокса CACGTG) в промоторной области гена eIF4E. [ 43 ] Этот мотив последовательности является общим с другими мишенями in vivo для myc, а мутации в повторах E-бокса eIF4E инактивируют промоторную область, тем самым уменьшая ее экспрессию.
Недавние исследования показали, что уровни eIF4E могут регулироваться на уровне транскрипции с помощью NFkB и C/EBP. [ 44 ] [ 45 ] Трансдукция первичных клеток ОМЛ IkB-SR приводила не только к снижению уровня мРНК eIF4E, но и к релокализации белка eIF4E. [ 25 ] Стабильность мРНК eIF4E также регулируется белками HuR и TIAR. [ 46 ] [ 47 ] Амплификация гена eIF4E наблюдалась в подмножестве образцов рака головы и шеи и молочной железы. [ 48 ]
б. Регуляция eIF4E путем фосфорилирования
Стимулы, такие как гормоны, факторы роста и митогены, которые способствуют пролиферации клеток, также повышают скорость трансляции за счет фосфорилирования eIF4E. [ 49 ] Хотя скорости фосфорилирования и трансляции eIF4E не всегда коррелируют, постоянные закономерности фосфорилирования eIF4E наблюдаются на протяжении всего клеточного цикла; при этом низкое фосфорилирование наблюдается во время фаз G 0 и M и при этом высокое фосфорилирование наблюдается во время фаз G 1 и S. [ 50 ] Эти данные дополнительно подтверждаются кристаллической структурой eIF4E, которая предполагает, что фосфорилирование по остатку серина 209 может увеличивать сродство eIF4E к кэпированной мРНК.
Фосфорилирование eIF4E также связано с его способностью подавлять экспорт РНК и его онкогенным потенциалом, что впервые было показано на клеточных линиях. [ 51 ]
в. Регуляция eIF4E белками-партнерами
Сборка комплекса eIF4F ингибируется белками, известными как eIF4E-связывающие белки (4E-BP), которые представляют собой небольшие термостабильные белки, которые блокируют кэп-зависимую трансляцию. [ 41 ] Нефосфорилированные 4E-BP сильно взаимодействуют с eIF4E, тем самым предотвращая трансляцию; тогда как фосфорилированные 4E-BP слабо связываются с eIF4E и, таким образом, не мешают процессу трансляции. [ 52 ] Более того, связывание 4E-BP ингибирует фосфорилирование Ser209 на eIF4E. [ 53 ] Следует отметить, что 4E-BP1 обнаруживается как в ядре, так и в цитоплазме, что указывает на то, что он, вероятно, также модулирует функции ядерного eIF4Es eIF4E. [ 54 ] Недавнее исследование показало, что 4E-BP3 регулирует eIF4E-зависимый ядерно-цитоплазматический экспорт мРНК. [ 55 ] Существует также множество цитоплазматических регуляторов eIF4E, которые связываются с тем же сайтом, что и 4E-BP1.
Было обнаружено множество других белков-партнеров, которые могут как стимулировать, так и подавлять активность eIF4E, например, белки, содержащие гомеодомен, включая HoxA9, Hex/PRH, Hox 11, Bicoid, Emx-2 и Engrailed 2. [ 56 ] [ 24 ] [ 57 ] [ 58 ] [ 59 ] В то время как HoxA9 способствует экспорту мРНК и активности трансляции eIF4E, Hex/PRH ингибирует ядерные функции eIF4E. [ 25 ] [ 60 ] [ 61 ] РНК-хеликаза DDX3 напрямую связывается с eIF4E, модулирует трансляцию и потенциально выполняет функции P-телец и экспорта мРНК. [ 62 ] [ 26 ]
Домены RING также связывают eIF4E. Белок промиелоцитарного лейкоза PML является мощным супрессором как экспорта ядерной РНК, так и онкогенной активности eIF4E, при этом домен RING PML напрямую связывает eIF4E на его дорсальной поверхности, подавляя онкогенную активность eIF4E; и, более того, подмножество ядерных тел PML и eIF4E локализуются совместно. [ 63 ] [ 21 ] [ 64 ] [ 23 ] [ 24 ] [ 28 ] Комплексы РНК-eIF4E никогда не наблюдаются в тельцах PML, что соответствует наблюдению, что PML подавляет функцию связывания кэпа m7G eIF4E. [ 21 ] [ 64 ] [ 28 ] Структурные исследования показывают, что родственный белок RING-пальца аренавируса, белок Lassa Fever Z, может аналогичным образом связывать eIF4E на дорсальной поверхности. [ 64 ] [ 65 ] [ 66 ]
Вход eIF4E в ядро опосредован его прямым взаимодействием с импортином 8, где импортин 8 связывается с кэп-связывающим сайтом m7G eIF4E. [ 32 ] Действительно, снижение уровня импортина 8 снижает онкогенный потенциал клеток, сверхэкспрессирующих eIF4E, и их функцию экспорта РНК. Импортин 8 связывается с кэп-сайтом связывания eIF4E и конкурирует с избытком m. 7 Аналоги G cap по данным ЯМР. eIF4E также стимулирует экспорт РНК импортина 8, тем самым производя больше белка импортина 8. В зависимости от типа клеток могут существовать дополнительные импортины, которые играют эту роль. Хотя первоначальное исследование показало, что белок-транспортер eIF4E 4E-T (eIF4ENIF1) облегчает проникновение в ядро, более поздние исследования показали, что этот фактор скорее изменяет локализацию eIF4E в цитоплазматических процессинговых тельцах (P-тельцах) и подавляет трансляцию. [ 67 ]
Было обнаружено, что связанный с геномом вирусного белка потивируса (VPg) напрямую связывает eIF4E в своем сайте связывания кэпа. VPg ковалентно связан со своей геномной РНК, и это взаимодействие позволяет VPg действовать как «кэп». [ 68 ] [ 69 ] [ 16 ] [ 70 ] VPg потивируса не имеет последовательности или структурной гомологии с другими VPg, например, VPg полиовируса. In vitro конъюгаты VPg-РНК транслировались с эффективностью, аналогичной m. 7 РНК с G-кепами, указывающие на то, что VPg связывает eIF4E и задействует механизм трансляции; в то время как свободный VPg (в отсутствие конъюгированной РНК) успешно конкурирует за все кэп-зависимые активности eIF4E в клетке, ингибируя трансляцию и экспорт РНК. [ 70 ]
д. Регуляция клеточной локализации eIF4E
Несколько факторов, которые регулируют функции eIF4E, также модулируют субклеточную локализацию eIF4E. Например, сверхэкспрессия PRH/Hex приводит к удержанию в цитоплазме eIF4E и, таким образом, к потере его экспортной активности мРНК и подавлению трансформации. [ 24 ] Сверхэкспрессия PML приводит к секвестрации eIF4E в ядерных тельцах с PML и уменьшению количества ядерных телец eIF4E, содержащих РНК, что коррелирует с подавлением eIF4E-зависимого экспорта мРНК и может модулироваться стрессом. [ 21 ] [ 23 ] [ 25 ] Сверхэкспрессия LRPPRC снижает совместную локализацию eIF4E с PML в ядре и приводит к увеличению активности экспорта мРНК eIF4E. Как обсуждалось выше, импортин 8 переносит eIF4E в ядро, и его сверхэкспрессия стимулирует экспорт РНК и активность онкогенной трансформации eIF4E в клеточных линиях. Трансдукция первичных клеток ОМЛ IkB-SR приводила не только к снижению уровня мРНК eIF4E, но и к релокализации белка eIF4E. [ 25 ]
Роль eIF4E при раке
[ редактировать ]Роль eIF4E в развитии рака была установлена после того, как Lazaris-Karatzas et al. сделали открытие, что сверхэкспрессия eIF4E вызывает онкогенную трансформацию фибробластов. [ 71 ] С момента этого первоначального наблюдения многочисленные группы повторили эти результаты на различных клеточных линиях. [ 72 ] В результате активность eIF4E участвует в развитии нескольких видов рака, включая рак молочной железы, легких и простаты. Фактически, транскрипционное профилирование метастатических опухолей человека выявило отчетливую метаболическую характеристику, при которой eIF4E, как известно, постоянно активируется. [ 73 ]
Уровни eIF4E повышаются при многих видах рака, включая острый миелоидный лейкоз (ОМЛ), множественную миелому, детский ОЛЛ, диффузную крупноклеточную В-клеточную лимфому, рак молочной железы, рак предстательной железы, рак головы и шеи, и его повышение обычно коррелирует с плохим прогнозом. [ 30 ] [ 74 ] [ 75 ] [ 76 ] [ 77 ] [ 78 ] [ 79 ] [ 80 ] При многих из этих видов рака, таких как ОМЛ, eIF4E обогащен ядрами, и обнаружено, что некоторые активности eIF4E повышены в первичных образцах пациентов, включая кэпирование, сплайсинг, экспорт РНК и трансляцию.
В первых клинических испытаниях eIF4E в качестве конкурента m7G использовался старый противовирусный препарат рибавирин, который обладал значительной активностью в линиях раковых клеток и на животных моделях, связанных с нарушенной регуляцией eIF4E. [ 81 ] [ 82 ] [ 75 ] [ 31 ] [ 83 ] [ 84 ] [ 85 ] [ 78 ] [ 86 ] [ 87 ] [ 88 ] [ 89 ] [ 90 ] [ 80 ] В первом исследовании, направленном на eIF4E, было продемонстрировано, что монотерапия рибавирином ингибирует активность eIF4E, что приводит к объективным клиническим ответам, включая полную ремиссию у пациентов с ОМЛ. [ 30 ] Интересно, что релокализация eIF4E из ядра в цитоплазму коррелирует с клиническими ремиссиями, что указывает на значимость его ядерной активности для прогрессирования заболевания. [ 30 ] Последующие исследования рибавирина при ОМЛ в сочетании с противолейкемическими препаратами снова показали объективные клинические ответы, включая ремиссии и молекулярное воздействие на eIF4E. [ 76 ] [ 91 ] В этих исследованиях ОМЛ клинические ответы коррелировали со снижением ядерного eIF4E, а клинический рецидив — с повторным появлением ядерного eIF4E eIF4E и его активностью по экспорту РНК. Другие исследования, в которых использовалась комбинация рибавирина, показали аналогичные многообещающие результаты при раке головы и шеи. [ 79 ] Рибавирин нарушает все изученные на сегодняшний день активности eIF4E (сплайсинг, кэпирование, экспорт и трансляция РНК). Таким образом, eIF4E оказался успешно терапевтически целенаправленным для людей; однако лекарственная устойчивость к рибавирину является новой проблемой для долгосрочного контроля заболевания. [ 84 ] [ 76 ] [ 91 ]
На eIF4E также воздействовали антисмысловые олигонуклеотиды, которые оказались очень эффективными на мышиных моделях рака простаты. [ 92 ] но в исследованиях монотерапии на людях не было получено клинической пользы, вероятно, из-за неэффективности снижения уровней eIF4E у людей по сравнению с мышами. [ 93 ] Существует также аллостерический ингибитор eIF4E, который связывается между местом связывания кэпа и дорсальной поверхностью, который используется экспериментально. [ 94 ]
FMRP подавляет трансляцию посредством связывания EIF4E
[ редактировать ]Белок умственной отсталости Fragile X ( FMR1 ) регулирует трансляцию специфических мРНК посредством связывания eIF4E. FMRP действует путем связывания CYFIP1 , который напрямую связывает eIF4e с доменом, который структурно аналогичен доменам, обнаруженным в 4E-BP, включая EIF4EBP3, EIF4EBP1 и EIF4EBP2. Комплекс FMRP/CYFIP1 связывается таким образом, чтобы предотвратить взаимодействие eIF4E-eIF4G, необходимое для трансляции осуществления . Взаимодействие FMRP/CYFIP1/eIF4E усиливается наличием мРНК (ов). В частности, РНК BC1 обеспечивает оптимальное взаимодействие FMRP и CYFIP1. [ 95 ] РНК-BC1 представляет собой нетранслируемую дендритную мРНК, которая связывает FMRP, обеспечивая его ассоциацию со специфической целевой мРНК. BC1 может функционировать, регулируя взаимодействия FMRP и мРНК в синапсе (ах) посредством рекрутирования FMRP на соответствующую мРНК. [ 96 ]
Кроме того, FMRP может рекрутировать CYFIP1 на специфические мРНК для подавления трансляции. Ингибитор трансляции FMRP-CYFIP1 регулируется стимуляцией нейрона (ов). Повышенная синаптическая стимуляция привела к диссоциации eIF4E и CYFIP1, что позволило инициировать трансляцию. [ 95 ]
Взаимодействия
[ редактировать ]Было показано, что EIF4E взаимодействует с:
. Другие непосредственные участники: PML; [ 21 ] [ 64 ] белок Z аренавируса; [ 64 ] [ 63 ] [ 65 ] [ 66 ] Импортин 8; [ 32 ] белок VPg потивируса, [ 70 ] ЛРППРК, [ 35 ] [ 26 ] РНМТ [ 117 ] и другие.
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Перейти обратно: а б с GRCh38: Версия Ensembl 89: ENSG00000151247 – Ensembl , май 2017 г.
- ^ Перейти обратно: а б с GRCm38: выпуск Ensembl 89: ENSMUSG00000028156 – Ensembl , май 2017 г.
- ^ «Ссылка на Human PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Ссылка на Mouse PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ Пеллетье Дж., Брук Дж.Д., Хаусман Д.Э. (август 1991 г.). «Присвоение двух генов фактора инициации трансляции-4E (EIF4EL1 и EIF4EL2) хромосомам 4 и 20 человека». Геномика . 10 (4): 1079–82. дои : 10.1016/0888-7543(91)90203-Q . ПМИД 1916814 .
- ^ Джонс Р.М., Макдональд М.Э., Бранда Дж., Альтерр М.Р., Луис Д.Н., Шмидт Е.В. (май 1997 г.). «Присвоение человеческого гена, кодирующего фактор инициации эукариот 4E (EIF4E), в область q21-25 на хромосоме 4». Соматическая клетка и молекулярная генетика . 23 (3): 221–223. дои : 10.1007/BF02721373 . ПМИД 9330633 . S2CID 10683455 .
- ^ Соненберг Н., Рупрехт К.М., Хехт С.М., Шаткин А.Ю. (сентябрь 1979 г.). «Белок, связывающий кэп-связывающий эукариотический мРНК: очистка с помощью аффинной хроматографии на m7GDP, связанном с сефарозой» . Труды Национальной академии наук Соединенных Штатов Америки . 76 (9): 4345–9. Бибкод : 1979PNAS...76.4345S . дои : 10.1073/pnas.76.9.4345 . ПМК 411571 . ПМИД 291969 .
- ^ Хатчинс А.П., Робертс Г.Р., Ллойд К.В., Дунан Дж.Х. (2004). «Взаимодействие между CDKA и eIF4A in vivo: возможный механизм, связывающий трансляцию и пролиферацию клеток» . ФЭБС Летт . 556 (1–3): 91–4. дои : 10.1016/S0014-5793(03)01382-6 . ПМИД 14706832 . S2CID 35343626 .
- ^ Се AC, Руджеро Д (11 августа 2010 г.). «Нацеливание на эукариотический фактор инициации трансляции 4E (eIF4E) при раке» . Клинические исследования рака . 16 (20): 4914–4920. дои : 10.1158/1078-0432.CCR-10-0433 . ПМЦ 7539621 . ПМИД 20702611 .
- ^ Рыхлик В., Домье Л.Л., Гарднер П.Р., Хеллманн Г.М., Роудс Р.Э. (февраль 1987 г.). «Аминокислотная последовательность кэп-связывающего белка мРНК из тканей человека» . Труды Национальной академии наук Соединенных Штатов Америки . 84 (4): 945–9. Бибкод : 1987ПНАС...84..945Р . дои : 10.1073/pnas.84.4.945 . ПМК 304336 . ПМИД 3469651 .
- ^ «Ген Энтрез: eIF4E Эукариотический фактор инициации трансляции 4E» .
- ^ Перейти обратно: а б де ла Парра К, Эрнлунд А, Алард А, Рагглс К, Юберхайде Б, Шнайдер Р.Дж. (03 августа 2018 г.). «Распространенная альтернативная форма кэп-зависимой инициации трансляции мРНК» . Природные коммуникации . 9 (1): 3068. Бибкод : 2018NatCo...9.3068D . дои : 10.1038/s41467-018-05539-0 . ISSN 2041-1723 . ПМК 6076257 . ПМИД 30076308 .
- ^ Перейти обратно: а б Ли А.С., Кранцуш П.Дж., Дудна Дж.А., Кейт Дж.Х. (04 августа 2016 г.). «eIF3d — это белок, связывающий кэп мРНК, который необходим для специализированной инициации трансляции» . Природа . 536 (7614): 96–99. Бибкод : 2016Natur.536...96L . дои : 10.1038/nature18954 . ISSN 0028-0836 . ПМК 5003174 . ПМИД 27462815 .
- ^ Бухари С.И., Трусделл С.С., Ли С., Коллу С., Классон А., Бухали М., Джайн Э., Мортенсен Р.Д., Янагия А., Садреев Р.И., Хаас В., Васудеван С. (март 2016 г.). «Специализированный механизм трансляции, опосредованный FXR1a-ассоциированной микроРНП в клеточном покое» . Молекулярная клетка . 61 (5): 760–773. doi : 10.1016/j.molcel.2016.02.013 . ПМЦ 4811377 . ПМИД 26942679 .
- ^ Кумар П., Хеллен С.У., Пестова Т.В. (01.07.2016). «К механизму eIF4F-опосредованного прикрепления рибосом к мРНК млекопитающих» . Гены и развитие . 30 (13): 1573–1588. дои : 10.1101/gad.282418.116 . ISSN 0890-9369 . ПМЦ 4949329 . ПМИД 27401559 .
- ^ Перейти обратно: а б Борден К.Л., Вольпон Л. (01.09.2020). «Разнообразие, пластичность и адаптируемость кэп-зависимой инициации трансляции и связанного с ней механизма» . Биология РНК . 17 (9): 1239–1251. дои : 10.1080/15476286.2020.1766179 . ISSN 1547-6286 . ПМЦ 7549709 . ПМИД 32496897 .
- ^ Рамбут Икс, Макват Л.Е. (01 сентября 2020 г.). «Комплекс, связывающий ядерный кэп, как хореограф транскрипции генов и процессинга пре-мРНК» . Гены и развитие . 34 (17–18): 1113–1127. дои : 10.1101/gad.339986.120 . ISSN 0890-9369 . ПМК 7462061 . ПМИД 32873578 .
- ^ Перейти обратно: а б с Марс Дж.К., Грам М., Чулкович-Кралячич Б., Борден К.Л. (08 декабря 2021 г.). «Кэп-связывающий комплекс CBC и эукариотический фактор трансляции eIF4E: сообщники в созревании и трансляции кэп-зависимой РНК» . Раки . 13 (24): 6185. doi : 10.3390/cancers13246185 . ISSN 2072-6694 . ПМЦ 8699206 . ПМИД 34944805 .
- ^ Лейбкович Ф., Гойер С., Дарво А., Нерон С., Лемье Р., Зоненберг Н. (15 октября 1992 г.). «Фракция 5'-кэп-связывающего белка мРНК, эукариотического фактора инициации 4E, локализуется в ядре» . Труды Национальной академии наук . 89 (20): 9612–9616. Бибкод : 1992PNAS...89.9612L . дои : 10.1073/pnas.89.20.9612 . ISSN 0027-8424 . ПМК 50182 . ПМИД 1384058 .
- ^ Дости Дж., Лейбкович Ф., Зоненберг Н. (24 января 2000 г.). «Ядерный фактор инициации эукариот 4e (Eif4e) колокализуется с факторами сплайсинга в спеклах» . Журнал клеточной биологии . 148 (2): 239–246. дои : 10.1083/jcb.148.2.239 . ISSN 0021-9525 . ПМК 2174286 . ПМИД 10648556 .
- ^ Перейти обратно: а б с д и ж г час Коэн Н. (15 августа 2001 г.). «PML RING подавляет онкогенную трансформацию за счет снижения сродства eIF4E к мРНК» . Журнал ЭМБО . 20 (16): 4547–4559. дои : 10.1093/emboj/20.16.4547 . ПМК 125576 . ПМИД 11500381 .
- ^ Иборра Ф.Дж., Джексон Д.А., Кук PR (10 августа 2001 г.). «Совместная транскрипция и трансляция в ядрах клеток млекопитающих» . Наука . 293 (5532): 1139–1142. дои : 10.1126/science.1061216 . ISSN 0036-8075 . ПМИД 11423616 . S2CID 17404294 .
- ^ Перейти обратно: а б с д Тописирович И., Капили А.Д., Борден К.Л. (1 сентября 2002 г.). «Обработка гамма-интерфероном и кадмием модулирует эукариотический фактор инициации 4E-зависимый транспорт мРНК циклина D1 PML-зависимым способом» . Молекулярная и клеточная биология . 22 (17): 6183–6198. дои : 10.1128/MCB.22.17.6183-6198.2002 . ISSN 1098-5549 . ПМК 134012 . ПМИД 12167712 .
- ^ Перейти обратно: а б с д и ж г Тописирович I (3 февраля 2003 г.). «Богатый пролином гомеодомен, PRH, является тканеспецифичным ингибитором eIF4E-зависимого транспорта и роста мРНК циклина D1» . Журнал ЭМБО . 22 (3): 689–703. дои : 10.1093/emboj/cdg069 . ПМК 140753 . ПМИД 12554669 .
- ^ Перейти обратно: а б с д и ж г час Тописирович И., Гузман М.Л., МакКоннелл М.Дж., Лихт Дж.Д., Цулькович Б., Неринг С.Дж., Джордан К.Т., Борден К.Л. (01 декабря 2003 г.). «Аберрантный эукариотический фактор инициации трансляции 4E-зависимый транспорт мРНК препятствует гемопоэтической дифференцировке и способствует лейкемогенезу» . Молекулярная и клеточная биология . 23 (24): 8992–9002. дои : 10.1128/MCB.23.24.8992-9002.2003 . ISSN 1098-5549 . ПМК 309660 . ПМИД 14645512 .
- ^ Перейти обратно: а б с д и ж г Тописирович И., Сиддики Н., Лапуант В.Л., Трост М., Тибо П., Бангеранье С., Пиньоль-Рома С., Борден К.Л. (22 апреля 2009 г.). «Молекулярное исследование эукариотического фактора инициации 4E (eIF4E) экспортно-компетентного RNP» . Журнал ЭМБО . 28 (8): 1087–1098. дои : 10.1038/emboj.2009.53 . ISSN 0261-4189 . ПМК 2683702 . ПМИД 19262567 .
- ^ Перейти обратно: а б с Цулькович Б, Тописирович И, Скрабанек Л, Руис-Гутьеррес М, Борден К.Л. (25 апреля 2005 г.). «eIF4E способствует ядерному экспорту мРНК циклина D1 через элемент в 3'UTR» . Журнал клеточной биологии . 169 (2): 245–256. дои : 10.1083/jcb.200501019 . ISSN 1540-8140 . ПМК 2171863 . ПМИД 15837800 .
- ^ Перейти обратно: а б с д и Цулькович Б, Тописирович И, Скрабанек Л, Руис-Гутьеррес М, Борден К.Л. (06 ноября 2006 г.). «eIF4E является центральным узлом регулона РНК, который управляет клеточной пролиферацией» . Журнал клеточной биологии . 175 (3): 415–426. дои : 10.1083/jcb.200607020 . ISSN 1540-8140 . ПМК 2064519 . PMID 17074885 .
- ^ Руссо Д., Каспар Р., Розенвальд И., Герке Л., Зоненберг Н. (6 февраля 1996 г.). «Инициация трансляции орнитиндекарбоксилазы и нуклеоцитоплазматический транспорт мРНК циклина D1 увеличиваются в клетках, сверхэкспрессирующих эукариотический фактор инициации 4E» . Труды Национальной академии наук . 93 (3): 1065–1070. Бибкод : 1996PNAS...93.1065R . дои : 10.1073/pnas.93.3.1065 . ISSN 0027-8424 . ПМК 40031 . ПМИД 8577715 .
- ^ Перейти обратно: а б с д и Ассулин С., Цулкович Б., Коколакис Э., Руссо С., Беслу Н., Амри А., Каплан С., Лебер Б., Рой Д.С., Миллер В.Х., Борден К.Л. (09.07.2009). «Молекулярное воздействие на онкоген eIF4E при остром миелолейкозе (ОМЛ): клиническое исследование, подтверждающее принцип действия рибавирина» . Кровь . 114 (2): 257–260. дои : 10.1182/кровь-2009-02-205153 . ISSN 0006-4971 . ПМИД 19433856 . S2CID 28957125 .
- ^ Перейти обратно: а б Боллманн Ф., Фешир К., Новаг С., Кох К., Арт Дж., Кляйнерт Х., Паутц А. (апрель 2013 г.). «Экспрессия индуцируемой синтазы оксида азота человека (iNOS) зависит от сохранения участка хромосомы 1 (CRM1) и транспорта мРНК нуклеоцитоплазмы, опосредованного эукариотическим фактором инициации трансляции 4E (elF4E)» . Оксид азота . 30 : 49–59. дои : 10.1016/j.niox.2013.02.083 . ПМИД 23471078 .
- ^ Перейти обратно: а б с Вольпон Л., Чулкович-Кралячич Б., Осборн М.Дж., Рамтеке А., Сан К., Нисман А., Чук Ю.М., Борден К.Л. (10 мая 2016 г.). «Импортин 8 опосредует чувствительный к кэпу m 7 G ядерный импорт эукариотического фактора инициации трансляции eIF4E» . Труды Национальной академии наук . 113 (19): 5263–5268. Бибкод : 2016PNAS..113.5263V . дои : 10.1073/pnas.1524291113 . ISSN 0027-8424 . ПМЦ 4868427 . ПМИД 27114554 .
- ^ Захреддин Х.А., Кулькович-Кралячич Б., Эмонд А., Петтерссон Ф., Мидура Р., Лауэр М., Дель Ринкон С., Кали В., Ассулин С., Миллер В.Х., Хаскалл В., Борден К.Л. (07.11.2017). «Эукариотический фактор инициации трансляции eIF4E использует выработку гиалуронана для управления своей злокачественной активностью» . электронная жизнь . 6 : е29830. doi : 10.7554/eLife.29830 . ISSN 2050-084X . ПМК 5705209 . ПМИД 29111978 .
- ^ Кулжкович-Кралячич Б, Фернандо ТМ, Марулло Р, Кальво-Видал Н, Верма А, Ян С, Таббо Ф, Гаудиано М, Захреддин Х, Гольдштейн РЛ, Патель Дж, Талдоне Т, Хиосис Г, Ладетто М, Гионе П (2016) -02-18). «Комбинаторное нацеливание ядерного экспорта и трансляции РНК ингибирует агрессивные В-клеточные лимфомы» . Кровь . 127 (7): 858–868. doi : 10.1182/blood-2015-05-645069 . ISSN 0006-4971 . ПМК 4760090 . ПМИД 26603836 .
- ^ Перейти обратно: а б с д и Вольпон Л., Цулькович-Кралячич Б., Зон Х.С., Бланше-Коэн А., Осборн М.Дж., Борден К.Л. (июнь 2017 г.). «Биохимическая основа eIF4E-зависимого экспорта мРНК и ядерной переработки экспортного оборудования» . РНК . 23 (6): 927–937. дои : 10.1261/rna.060137.116 . ISSN 1355-8382 . ПМЦ 5435865 . ПМИД 28325843 .
- ^ Дэвис М.Р., Делало М., Борден К.Л. (апрель 2019 г.). «Ядерный eIF4E стимулирует 3'-концевое расщепление целевых РНК» . Отчеты по ячейкам . 27 (5): 1397–1408.e4. дои : 10.1016/j.celrep.2019.04.008 . ПМК 6661904 . ПМИД 31042468 .
- ^ Перейти обратно: а б Кулькович-Кралячич Б, Скрабанек Л, Ревуэльта М.В., Гасиорек Дж, Коулинг В.Х., Черкьетти Л., Борден К.Л. (27 октября 2020 г.). «Эукариотический фактор инициации трансляции eIF4E повышает стационарное кэпирование m 7 G кодирующих и некодирующих транскриптов» . Труды Национальной академии наук . 117 (43): 26773–26783. Бибкод : 2020PNAS..11726773C . дои : 10.1073/pnas.2002360117 . ISSN 0027-8424 . ПМЦ 7604501 . ПМИД 33055213 .
- ^ Перейти обратно: а б Грам М., Моррис Г., Кулкович-Кралячич Б., Марс Дж.К., Гендрон П., Скрабанек Л., Ревуэльта М.В., Черкьетти Л., Гузман М.Л., Борден К.Л. (03.04.2023). «Эукариотический фактор инициации трансляции eIF4E перепрограммирует альтернативный сплайсинг» . Журнал ЭМБО . 42 (7): e110496. дои : 10.15252/embj.2021110496 . ISSN 0261-4189 . ПМЦ 10068332 . ПМИД 36843541 .
- ^ Вольпон Л., Осборн М.Дж., Борден К.Л. (20 мая 2019 г.). «Биохимические и структурные данные об эукариотическом факторе инициации трансляции eIF4E» . Современная наука о белках и пептидах . 20 (6): 525–535. дои : 10.2174/1389203720666190110142438 . ПМИД 30636602 . S2CID 58587801 .
- ^ Дункан Р., Милберн СК, Херши Дж.В. (5 января 1987 г.). «Регулируемое фосфорилирование и низкое содержание фактора инициации клеток HeLa eIF-4F предполагают его роль в контроле трансляции. Влияние теплового шока на eIF-4F» . Журнал биологической химии . 262 (1): 380–388. дои : 10.1016/S0021-9258(19)75938-9 . ISSN 0021-9258 . ПМИД 3793730 .
- ^ Перейти обратно: а б Рихтер Дж. Д., Зоненберг Н. (3 февраля 2005 г.). «Регуляция кэп-зависимой трансляции с помощью белков, ингибирующих eIF4E» . Природа . 433 (7025): 477–480. Бибкод : 2005Natur.433..477R . дои : 10.1038/nature03205 . ISSN 1476-4687 . ПМИД 15690031 . S2CID 4347657 .
- ^ Розенвальд И.Б., Роудс Д.Б., Калланан Л.Д., Иссельбахер К.Дж., Шмидт Э.В. (1 июля 1993 г.). «Повышение экспрессии эукариотических факторов инициации трансляции eIF-4E и eIF-2 альфа в ответ на индукцию роста c-myc» . Труды Национальной академии наук Соединенных Штатов Америки . 90 (13): 6175–6178. Бибкод : 1993PNAS...90.6175R . дои : 10.1073/pnas.90.13.6175 . ISSN 0027-8424 . ПМК 46890 . ПМИД 8327497 .
- ^ Джонс Р.М., Бранда Дж., Джонстон К.А., Полименис М., Гадд М., Рустги А., Калланан Л., Шмидт Е.В. (сентябрь 1996 г.). «Необходимый E-бокс в промоторе гена, кодирующего кэп-связывающий белок мРНК (эукариотический фактор инициации 4E), является мишенью для активации c-myc» . Молекулярная и клеточная биология . 16 (9): 4754–4764. дои : 10.1128/mcb.16.9.4754 . ISSN 0270-7306 . ПМК 231476 . ПМИД 8756633 .
- ^ Ханна-Гупта А., Абаясекара Н., Левин М., Сан Х., Вирджилио М., Ниа Н., Хален С., Спортолетти П., Чон Дж. Я., Пандольфи П. П., Берлинер Н. (сентябрь 2012 г.). «Повышающая регуляция эукариотического фактора инициации трансляции 4E в клетках с гаплонедостаточностью нуклеофозмина 1 приводит к изменениям активности α-белка, связывающего энхансер CCAAT» . Журнал биологической химии . 287 (39): 32728–32737. дои : 10.1074/jbc.M112.373274 . ПМЦ 3463350 . ПМИД 22851180 .
- ^ Харири Ф, Аргуэлло М, Вольпон Л, Кулкович-Кралячич Б, Нильсен Т.Х., Хискотт Дж., Манн К.К., Борден К.Л. (октябрь 2013 г.). «Эукариотический фактор инициации трансляции eIF4E является прямой мишенью транскрипции NF-κB и аберрантно регулируется при остром миелолейкозе» . Лейкемия . 27 (10): 2047–2055. дои : 10.1038/leu.2013.73 . ISSN 0887-6924 . ПМЦ 4429918 . ПМИД 23467026 .
- ^ Мазан-Мамцарз К., Лал А., Мартиндейл Дж.Л., Каваи Т., Гороспе М. (1 апреля 2006 г.). «Трансляционная репрессия с помощью РНК-связывающего белка TIAR» . Молекулярная и клеточная биология . 26 (7): 2716–2727. дои : 10.1128/MCB.26.7.2716-2727.2006 . ISSN 1098-5549 . ПМК 1430315 . ПМИД 16537914 .
- ^ Тописирович И., Сиддики Н., Оролицкий С., Скрабанек Л.А., Трембле М., Хоанг Т., Борден К.Л. (01.03.2009). «Стабильность мРНК эукариотического фактора инициации трансляции 4E регулируется HuR, и эта активность нарушается при раке» . Молекулярная и клеточная биология . 29 (5): 1152–1162. дои : 10.1128/MCB.01532-08 . ISSN 1098-5549 . ПМЦ 2643828 . ПМИД 19114552 .
- ^ Сорреллс Д.Л., Блэк Д.Р., Мешонат С., Роудс Р., Де Бенедетти А., Гао М., Уильямс Б.Дж., Ли Б.Д. (апрель 1998 г.). «Обнаружение амплификации гена eIF4E при раке молочной железы с помощью конкурентной ПЦР» . Анналы хирургической онкологии . 5 (3): 232–237. дои : 10.1007/BF02303778 . ISSN 1068-9265 . ПМИД 9607624 . S2CID 776478 .
- ^ Морли С.Дж., Трау Дж.А. (25 июня 1990 г.). «Дифференциальная стимуляция фосфорилирования факторов инициации eIF-4F, eIF-4B, eIF-3 и рибосомального белка S6 инсулином и эфирами форбола» . Журнал биологической химии . 265 (18): 10611–10616. дои : 10.1016/S0021-9258(18)86990-3 . ISSN 0021-9258 . ПМИД 2191953 .
- ^ Бонно А.М., Зоненберг Н. (15 августа 1987 г.). «Участие кэп-связывающего белка массой 24 кДа в регуляции синтеза белка при митозе» . Журнал биологической химии . 262 (23): 11134–11139. дои : 10.1016/S0021-9258(18)60935-4 . ISSN 0021-9258 . ПМИД 3038908 .
- ^ Тописирович И., Руис-Гутьеррес М., Борден К.Л. (1 декабря 2004 г.). «Фосфорилирование эукариотического фактора инициации трансляции eIF4E способствует его трансформации и транспортной активности мРНК» . Исследования рака . 64 (23): 8639–8642. дои : 10.1158/0008-5472.CAN-04-2677 . ISSN 0008-5472 . ПМИД 15574771 . S2CID 21104713 .
- ^ Питер Д., Играя С., Вебер Р., Вольболд Л., Вейлер С., Эбертш Л., Вайхенридер О., Изаурральде Е. (19 марта 2015 г.). «Молекулярная архитектура ингибиторов трансляции 4E-BP, связанных с eIF4E» . Молекулярная клетка . 57 (6): 1074–1087. doi : 10.1016/j.molcel.2015.01.017 . ISSN 1097-4164 . ПМИД 25702871 .
- ^ Уэлен С.Г., Гинграс АС, Аманква Л., Мадер С., Брэнтон П.Е., Эберсолд Р., Соненберг Н. (17 мая 1996 г.). «Фосфорилирование eIF-4E на серине 209 протеинкиназой C ингибируется репрессорами трансляции, 4E-связывающими белками» . Журнал биологической химии . 271 (20): 11831–11837. дои : 10.1074/jbc.271.20.11831 . ISSN 0021-9258 . ПМИД 8662663 .
- ^ Ронг Л., Ливингстон М., Сукарие Р., Петрулакис Э., Гинграс А.С., Кросби К., Смит Б., Полакевич Р.Д., Пеллетье Дж., Феррайуоло М.А., Соненберг Н. (июль 2008 г.). «Контроль клеточной локализации eIF4E с помощью eIF4E-связывающих белков, 4E-BP» . РНК . 14 (7): 1318–1327. дои : 10.1261/rna.950608 . ISSN 1355-8382 . ПМК 2441981 . ПМИД 18515545 .
- ^ Чен CC, Ли JC, Чанг MC (30 июля 2012 г.). «4E-BP3 регулирует опосредованный eIF4E экспорт ядерной мРНК и взаимодействует с репликационным белком A2» . Письма ФЭБС . 586 (16): 2260–2266. дои : 10.1016/j.febslet.2012.05.059 . ПМИД 22684010 . S2CID 40980342 .
- ^ Ниссинг Д., Бланке С., Джекль Х (1 октября 2002 г.). «Бикоид связывается с 5'-кэп-связанным комплексом каудальной мРНК и подавляет трансляцию» . Гены и развитие . 16 (19): 2576–2582. дои : 10.1101/gad.240002 . ISSN 0890-9369 . ЧВК 187448 . ПМИД 12368268 .
- ^ Неделек С., Фуше И., Брюне И., Буйо С., Прочианц А., Трембло А. (20 июля 2004 г.). «Фактор транскрипции гомеодомена Emx2 взаимодействует с эукариотическим фактором инициации трансляции 4E (eIF4E) в аксонах обонятельных сенсорных нейронов» . Труды Национальной академии наук . 101 (29): 10815–10820. Бибкод : 2004PNAS..10110815N . дои : 10.1073/pnas.0403824101 . ISSN 0027-8424 . ПМК 490017 . ПМИД 15247416 .
- ^ Брюне И., Вейнл С., Пайпер М., Трембло А., Волович М., Харрис В., Прочианц А., Холт С. (ноябрь 2005 г.). «Фактор транскрипции Engrailed-2 направляет аксоны сетчатки» . Природа . 438 (7064): 94–98. Бибкод : 2005Natur.438...94B . дои : 10.1038/nature04110 . ISSN 0028-0836 . ПМЦ 3785142 . ПМИД 16267555 .
- ^ Тописирович И., Борден К.Л. (октябрь 2005 г.). «Гомеодоменные белки и эукариотический фактор инициации трансляции 4E (eIF4E): неожиданная взаимосвязь» . Гистология и гистопатология . 20 (20): 1275–1284. дои : 10.14670/HH-20.1275 . ISSN 0213-3911 . ПМИД 16136508 .
- ^ Курода Н., Го Л., Миядзаки Э., Хамаузу Т., Той М., Хирои М., Энзан Х. (01.01.2005). «Появление миофибробластов и исчезновение CD34-позитивных стромальных клеток в зоне, прилегающей к ксантогранулематозным очагам хронического холецистита» . Гистология и гистопатология . 20 (20): 127–133. дои : 10.14670/HH-20.127 . ISSN 0213-3911 . ПМИД 15578431 .
- ^ Тописирович И., Кенцис А., Перес Х.М., Гузман М.Л., Джордан К.Т., Борден К.Л. (01 февраля 2005 г.). «Активность эукариотического фактора инициации трансляции 4E модулируется HOXA9 на нескольких уровнях» . Молекулярная и клеточная биология . 25 (3): 11:00–11:12. дои : 10.1128/MCB.25.3.1100-1112.2005 . ISSN 1098-5549 . ПМК 544005 . ПМИД 15657436 .
- ^ Ши Дж.В., Цай Т.Ю., Чао Ч., Ву Ли Ю.Х. (24 января 2008 г.). «Кандидатный супрессор опухоли РНК-хеликаза DDX3 специфически подавляет кэп-зависимую трансляцию, действуя как белок, ингибирующий eIF4E» . Онкоген . 27 (5): 700–714. дои : 10.1038/sj.onc.1210687 . ISSN 0950-9232 . ПМИД 17667941 . S2CID 19781838 .
- ^ Перейти обратно: а б Кенцис А., Гордон Р.Э., Борден К.Л. (26 ноября 2002 г.). «Контроль биохимических реакций посредством самосборки супрамолекулярного домена RING» . Труды Национальной академии наук . 99 (24): 15404–15409. Бибкод : 2002PNAS...9915404K . дои : 10.1073/pnas.202608799 . ISSN 0027-8424 . ПМК 137729 . ПМИД 12438698 .
- ^ Перейти обратно: а б с д и Кентсис А., Дуайер Э.К., Перес Дж.М., Шарма М., Чен А., Пан ЗК, Борден К.Л. (сентябрь 2001 г.). «Домены RING белка PML промиелоцитарного лейкоза и аренавирусного белка Z подавляют трансляцию путем прямого ингибирования фактора инициации трансляции eIF4E 1 1. Под редакцией Д. Дрейпера» . Журнал молекулярной биологии . 312 (4): 609–623. дои : 10.1006/jmbi.2001.5003 . ПМИД 11575918 .
- ^ Перейти обратно: а б Кенцис А., Гордон Р.Э., Борден К.Л. (22 января 2002 г.). «Свойства самосборки модельного домена RING» . Труды Национальной академии наук . 99 (2): 667–672. дои : 10.1073/pnas.012317299 . ISSN 0027-8424 . ПМЦ 117363 . ПМИД 11792829 .
- ^ Перейти обратно: а б Вольпон Л., Осборн М.Дж., Капул А.А., де ла Торре Дж.К., Борден К.Л. (23 марта 2010 г.). «Структурная характеристика комплекса Z RING-eIF4E показывает особый способ контроля eIF4E» . Труды Национальной академии наук . 107 (12): 5441–5446. Бибкод : 2010PNAS..107.5441V . дои : 10.1073/pnas.0909877107 . ISSN 0027-8424 . ПМЦ 2851782 . ПМИД 20212144 .
- ^ Феррайуоло М.А., Басак С., Дости Дж., Мюррей Э.Л., Шенберг Д.Р., Соненберг Н. (12 сентября 2005 г.). «Роль eIF4E-связывающего белка 4E-T в формировании P-телец и распаде мРНК» . Журнал клеточной биологии . 170 (6): 913–924. дои : 10.1083/jcb.200504039 . ISSN 1540-8140 . ПМК 2171455 . ПМИД 16157702 .
- ^ Герман-Ретана С., Вальтер Дж., Ле Галль О. (март 2008 г.). «Вирус мозаики салата: от разнообразия патогенов до взаимодействия с хозяином» . Молекулярная патология растений . 9 (2): 127–136. дои : 10.1111/j.1364-3703.2007.00451.x . ISSN 1464-6722 . ПМК 6640324 . ПМИД 18705846 .
- ^ Коутиньо де Оливейра Л., Вольпон Л., Осборн М.Дж., Борден К.Л. (апрель 2019 г.). «Назначение химического сдвига связанного с геномом вирусного белка (VPg) вируса Y картофеля» . Задания по биомолекулярному ЯМР . 13 (1): 9–13. дои : 10.1007/s12104-018-9842-3 . ISSN 1874-2718 . ПМК 6428624 . ПМИД 30242622 .
- ^ Перейти обратно: а б с Коутиньо де Оливейра Л., Вольпон Л., Рахарджо А.К., Осборн М.Дж., Кулкович-Кралячич Б., Трахан С., Оффингер М., Квок Б.Х., Борден К.Л. (26 ноября 2019 г.). «Структурные исследования комплекса eIF4E-VPg выявили прямую конкуренцию за кэпированную РНК: последствия для трансляции» . Труды Национальной академии наук . 116 (48): 24056–24065. Бибкод : 2019PNAS..11624056C . дои : 10.1073/pnas.1904752116 . ISSN 0027-8424 . ПМК 6883836 . ПМИД 31712417 .
- ^ Лазарис-Карацас А., Монтин К.С., Зоненберг Н. (7 июня 1990 г.). «Злокачественная трансформация субъединицей фактора инициации эукариот, которая связывается с 5'-кэпом мРНК». Природа . 345 (6275): 544–547. Бибкод : 1990Natur.345..544L . дои : 10.1038/345544a0 . ISSN 0028-0836 . ПМИД 2348862 . S2CID 4366949 .
- ^ Пеллетье Дж., Графф Дж., Руджеро Д., Соненберг Н. (15 января 2015 г.). «НАЦЕЛЕННОСТЬ НА КОМПЛЕКС ИНИЦИАЦИИ ТРАНСЛЯЦИИ eIF4F: ВАЖНАЯ СВЯЗЬ ДЛЯ РАЗВИТИЯ РАКА» . Исследования рака . 75 (2): 250–263. дои : 10.1158/0008-5472.CAN-14-2789 . ISSN 0008-5472 . ПМЦ 4299928 . ПМИД 25593033 .
- ^ Рамасвами С., Росс К.Н., Ландер Э.С., Голуб Т.Р. (январь 2003 г.). «Молекулярная подпись метастазов в первичных солидных опухолях». Природная генетика . 33 (1): 49–54. дои : 10.1038/ng1060 . ISSN 1546-1718 . ПМИД 12469122 . S2CID 12059602 .
- ^ Цулькович Б, Борден К.Л. (2009). «Понимание и нацеливание на эукариотический фактор инициации трансляции eIF4E при раке головы и шеи» . Журнал онкологии . 2009 : 981679. doi : 10.1155/2009/981679 . ISSN 1687-8450 . ПМЦ 2798714 . ПМИД 20049173 .
- ^ Перейти обратно: а б Петтерссон Ф., Яу К., Добокан М.К., Цулькович-Кралячич Б., Ретроувай Х., Пакетт Р., Флорес Л.М., Кроп И.Е., Руссо С., Коколакис Э., Борден К.Л., Бенц CC, Миллер В.Х. (01.05.2011). «Влияние лечения рибавирином на рак молочной железы со сверхэкспрессией eIF4E, биомаркера с прогностической специфичностью для люминального рака молочной железы B-типа» . Клинические исследования рака . 17 (9): 2874–2884. дои : 10.1158/1078-0432.CCR-10-2334 . ISSN 1078-0432 . ПМК 3086959 . ПМИД 21415224 .
- ^ Перейти обратно: а б с Ассулин С., Кулкович-Кралячич Б., Бержерон Дж., Каплан С., Коколакис Э., Ламберт С., Лау С.Дж., Захреддин Х.А., Миллер В.Х., Борден К.Л. (01.01.2015). «Фаза I исследования рибавирина и низких доз цитарабина для лечения рецидивирующего и рефрактерного острого миелолейкоза с повышенным уровнем eIF4E» . Гематологическая . 100 (1): e7–e9. дои : 10.3324/haematol.2014.111245 . ISSN 0390-6078 . ПМЦ 4281321 . ПМИД 25425688 .
- ^ Аттар-Шнайдер О, Пасманик-Чор М, Тартаковер-Маталон С, Друкер Л, Лишнер М (28 февраля 2015 г.). «eIF4E и eIF4GI имеют различные и дифференциальные отпечатки на протеоме и передаче сигналов множественной миеломы» . Онкотаргет . 6 (6): 4315–4329. дои : 10.18632/oncotarget.3008 . ISSN 1949-2553 . ПМЦ 4414192 . ПМИД 25717031 .
- ^ Перейти обратно: а б Зисманов В., Аттар-Шнайдер О., Лишнер М., Айзенфельд Р.Х., Маталон С.Т., Друкер Л. (февраль 2015 г.). «На протеостаз множественной миеломы можно воздействовать с помощью фактора инициации трансляции eIF4E» . Международный журнал онкологии . 46 (2): 860–870. дои : 10.3892/ijo.2014.2774 . ISSN 1019-6439 . ПМИД 25422161 .
- ^ Перейти обратно: а б Данн Л.А., Фьюри М.Г., Шерман Э.Дж., Хо А.А., Катаби Н., Хак С.С., Пфистер Д.Г. (февраль 2018 г.). «Исследование фазы I индукционной химиотерапии афатинибом, рибавирином и еженедельным приемом карбоплатина и паклитаксела при плоскоклеточном раке ротоглотки, ассоциированном с вирусом папилломы человека стадии IVA/IVB» . Голова и шея . 40 (2): 233–241. дои : 10.1002/hed.24938 . ПМК 6760238 . ПМИД 28963790 .
- ^ Перейти обратно: а б Уртишак К.А., Ван Л.С., Чулкович-Кралячич Б., Давенпорт Дж.В., Порацци П., Винсент Т.Л., Тичи Д.Т., Тасиан С.К., Мур Дж.С., Сейф А.Е., Джин С., Барретт Дж.С., Робинсон Б.В., Чен И.М., Харви Р.К. (2019–03) -28). «Нацеливание на передачу сигналов EIF4E с помощью рибавирина при остром лимфобластном лейкозе у младенцев» . Онкоген . 38 (13): 2241–2262. дои : 10.1038/s41388-018-0567-7 . ISSN 0950-9232 . ПМК 6440839 . ПМИД 30478448 .
- ^ Кенцис А, Тописирович И, Цулькович Б, Шао Л, Борден К.Л. (28 декабря 2004 г.). «Рибавирин подавляет опосредованную eIF4E онкогенную трансформацию путем физической мимикрии кэпа мРНК 7-метилгуанозина» . Труды Национальной академии наук . 101 (52): 18105–18110. Бибкод : 2004PNAS..10118105K . дои : 10.1073/pnas.0406927102 . ISSN 0027-8424 . ПМК 539790 . ПМИД 15601771 .
- ^ Кенцис А., Вольпон Л., Тописирович И., Солл С.Э., Цулкович Б., Шао Л., Борден К.Л. (декабрь 2005 г.). «Дополнительные доказательства того, что рибавирин взаимодействует с eIF4E» . РНК . 11 (12): 1762–1766. дои : 10.1261/rna.2238705 . ISSN 1355-8382 . ПМК 1370864 . ПМИД 16251386 .
- ^ Вольпон Л., Осборн М.Дж., Захреддин Х., Ромео А.А., Борден К.Л. (май 2013 г.). «Конформационные изменения, вызванные эукариотическим фактором инициации трансляции eIF4E клинически значимым ингибитором рибавирина трифосфатом» . Связь с биохимическими и биофизическими исследованиями . 434 (3): 614–619. дои : 10.1016/j.bbrc.2013.03.125 . ПМЦ 3659399 . ПМИД 23583375 .
- ^ Перейти обратно: а б Захреддин Х.А., Кулкович-Кралячич Б., Ассулин С., Гендрон П., Ромео А.А., Моррис С.Дж., Кормак Г., Жакит Дж.Б., Черкьетти Л., Коколакис Е., Амри А., Бержерон Дж., Лебер Б., Беккер М.В., Пей С. (2014–07 гг.) -03). «Фактор звукового ежа GLI1 придает устойчивость к лекарствам посредством индуцируемой глюкуронидации» . Природа . 511 (7507): 90–93. Бибкод : 2014Natur.511...90Z . дои : 10.1038/nature13283 . ISSN 0028-0836 . ПМК 4138053 . ПМИД 24870236 .
- ^ Ши Ф, Лен Й, Гонг Й, Ши Р, Ян Х, Нарен Д, Ян Т (28 августа 2015 г.). Ивс Си Джей (ред.). «Рибавирин ингибирует активность сигнального пути mTOR/eIF4E, ERK/Mnk1/eIF4E и взаимодействует с ингибитором тирозинкиназы иматинибом, ухудшая опосредованную Bcr-Abl пролиферацию и апоптоз при Ph+ лейкозе» . ПЛОС ОДИН . 10 (8): e0136746. Бибкод : 2015PLoSO..1036746S . дои : 10.1371/journal.pone.0136746 . ISSN 1932-6203 . ПМЦ 4552648 . ПМИД 26317515 .
- ^ Дай Д., Чен Х., Тан Дж., Тан Ю. (январь 2017 г.). «Ингибирование mTOR/eIF4E противовирусным препаратом рибавирином эффективно усиливает эффекты паклитаксела при плоскоклеточном раке полости рта и языка» . Связь с биохимическими и биофизическими исследованиями . 482 (4): 1259–1264. дои : 10.1016/j.bbrc.2016.12.025 . ПМИД 27932243 .
- ^ Вольпин Ф, Касаос Дж, Сесен Дж, Мангравити А, Чой Дж, Горелик Н, Фрикече Дж, Лотт Т, Фелдер Р, Шотландия СДж, Эйзингер-Мэтасон Т.С., Брем Х, Тайлер Б, Скули Н (25 мая 2017 г.) . «Использование противовирусного препарата рибавирина в качестве средства против глиобластомы» . Онкоген . 36 (21): 3037–3047. дои : 10.1038/onc.2016.457 . ISSN 0950-9232 . ПМИД 27941882 . S2CID 21655102 .
- ^ Ван Г, Ли З, Ли З, Хуан Ю, Мао Х, Сюй С, Цуй С (декабрь 2017 г.). «Нацеливание на eIF4E подавляет рост, выживаемость и ангиогенез при ретинобластоме и повышает эффективность химиотерапии» . Биомедицина и фармакотерапия . 96 : 750–756. дои : 10.1016/j.biopha.2017.10.034 . ПМИД 29049978 .
- ^ Си С, Ван Л, Юй Дж, Е Х, Цао Л, Гун Цзи (сентябрь 2018 г.). «Ингибирование эукариотического фактора инициации трансляции 4E эффективно против химиорезистентности рака толстой кишки и шейки матки» . Связь с биохимическими и биофизическими исследованиями . 503 (4): 2286–2292. дои : 10.1016/j.bbrc.2018.06.150 . ПМИД 29959920 . S2CID 49634908 .
- ^ Цзинь Дж, Сян В, Ву С, Ван М, Сяо М, Дэн А (март 2019 г.). «Нацеливание на передачу сигналов eIF4E с помощью рибавирина как сенсибилизирующая стратегия при раке яичников» . Связь с биохимическими и биофизическими исследованиями . 510 (4): 580–586. дои : 10.1016/j.bbrc.2019.01.117 . ПМИД 30739792 . S2CID 73419809 .
- ^ Перейти обратно: а б Ассулин С., Гасиорек Дж., Бержерон Дж., Ламберт С., Чулкович-Кралячич Б., Коколакис Е., Закария С., Шлахтыч Д., Йи К., Борден К.Л. (23 марта 2023 г.). «Молекулярное нацеливание ферментов UDP-глюкуронозилтрансферазы у пациентов с высоким эукариотическим фактором инициации трансляции 4E, рефрактерным / рецидивирующим острым миелолейкозом: рандомизированное исследование II фазы висмодегиба, рибавирина с децитабином или без него» . Гематологическая . 108 (11): 2946–2958. дои : 10.3324/гематол.2023.282791 . ISSN 1592-8721 . ПМЦ 10620574 . ПМИД 36951168 . S2CID 257733013 .
- ^ Графф Дж.Р., Коничек Б.В., Винсент Т.М., Линч Р.Л., Монтейт Д., Вейр С.Н., Швиер П., Кэпен А., Гуд Р.Л., Даулесс М.С., Чен Ю., Чжан Х., Сиссонс С., Кокс К., МакНалти А.М. (04 сентября 2007 г.) ). «Терапевтическое подавление экспрессии фактора инициации трансляции eIF4E снижает рост опухоли без токсичности» . Журнал клинических исследований . 117 (9): 2638–2648. дои : 10.1172/JCI32044 . ISSN 0021-9738 . ЧВК 1957541 . ПМИД 17786246 .
- ^ Хонг Д.С., Курцрок Р., О Й., Уилер Дж., Наинг А., Брэйл Л., Кэллис С., Андре В., Кадам С.К., Насир А., Хольцер Т.Р., Мерик-Бернстам Ф., Фишман М., Саймон Г (15 октября 2011 г.) . «Эскалация дозы, фармакокинетика и фармакодинамическая оценка антисмыслового олигонуклеотида eIF-4E LY2275796 в фазе 1 у пациентов с распространенным раком» . Клинические исследования рака . 17 (20): 6582–6591. дои : 10.1158/1078-0432.CCR-11-0430 . ISSN 1078-0432 . ПМК 5036398 . ПМИД 21831956 .
- ^ Пападопулос Э, Дженни С, Кабха Э, Такрури КДж, Йи Т, Сальви Н, Луна РЭ, Гаватиотис Э, Махалингам П, Артханари Х, Родригес-Миас Р, Ефидофф-Фридман Р, Актас Б.Х., Чорев М, Гальперин Дж.А. (2014) -08-05). «Структура эукариотического фактора инициации трансляции eIF4E в комплексе с 4EGI-1 раскрывает аллостерический механизм диссоциации eIF4G» . Труды Национальной академии наук . 111 (31): Е3187-95. Бибкод : 2014PNAS..111E3187P . дои : 10.1073/pnas.1410250111 . ISSN 0027-8424 . ПМК 4128100 . ПМИД 25049413 .
- ^ Перейти обратно: а б Наполи I, Меркальдо В, Бойл П.П., Элеутери Б, Зальфа Ф, Де Рубейс С, Ди Марино Д, Мор Е, Массими М, Фалькони М, Витке В, Коста-Маттиоли М, Зоненберг Н, Аксель Т, Баньи К (сентябрь) 2008). «Белок синдрома хрупкой X подавляет зависимую от активности трансляцию посредством CYFIP1, нового 4E-BP» . Клетка . 134 (6): 1042–1054. дои : 10.1016/j.cell.2008.07.031 . ПМИД 18805096 . S2CID 14123165 .
- ^ Зальфа Ф, Джорджи М, Примерано Б, Моро А, Ди Пента А, Рейс С, Оостра Б, Баньи С (февраль 2003 г.). «Белок синдрома хрупкой Х-хромосомы FMRP связывается с РНК BC1 и регулирует трансляцию специфических мРНК в синапсах» . Клетка . 112 (3): 317–27. дои : 10.1016/S0092-8674(03)00079-5 . ПМИД 12581522 . S2CID 14892764 .
- ^ Перейти обратно: а б Юинг Р.М., Чу П., Элизма Ф., Ли Х., Тейлор П., Клими С., МакБрум-Серажевски Л., Робинсон М.Д., О'Коннор Л., Ли М., Тейлор Р., Дарси М., Хо Ю, Хейлбут А., Мур Л., Чжан С., Орнацкий О., Бухман Ю.В., Этьер М., Шэн Ю., Василеску Дж., Абу-Фарха М., Ламберт Дж.П., Дювел. Х.С., Стюарт II, Кюль Б., Хог К., Колвилл К., Гладвиш К., Маскат Б., Кинач Р., Адамс С.Л., Моран М.Ф., Морин ГБ, Топалоглу Т., Фигейс Д. (2007). «Крупномасштабное картирование белково-белковых взаимодействий человека методом масс-спектрометрии» . Мол. Сист. Биол . 3 : 89. дои : 10.1038/msb4100134 . ПМЦ 1847948 . ПМИД 17353931 .
- ^ Перейти обратно: а б с Коннолли Э., Браунштейн С., Форменти С., Шнайдер Р.Дж. (май 2006 г.). «Гипоксия ингибирует синтез белка посредством киназы 4E-BP1 и фактора элонгации 2, контролируемой mTOR и не связанной в клетках рака молочной железы» . Мол. Клетка. Биол . 26 (10): 3955–65. дои : 10.1128/MCB.26.10.3955-3965.2006 . ПМК 1489005 . ПМИД 16648488 .
- ^ Руал Дж.Ф., Венкатесан К., Хао Т., Хирозане-Кисикава Т., Дрико А., Ли Н., Берриз Г.Ф., Гиббонс Ф.Д., Дрез М., Айви-Гедехуссу Н., Клитгорд Н., Саймон С., Боксем М., Мильштейн С., Розенберг Дж., Голдберг Д.С., Чжан Л.В., Вонг С.Л., Франклин Г., Ли С., Альбала Дж.С., Лим Дж., Фротон С, Лламосас Е, Чевик С, Бекс С, Ламеш П, Сикорски Р.С., Ванденхаут Дж, Зогби Х.И., Смоляр А, Босак С., Секерра Р., Дусетт-Стамм Л., Кьюсик М.Е., Хилл Д.Е., Рот Ф.П., Видал М. ( октябрь 2005 г.). «К карте сети белок-белковых взаимодействий человека в масштабе протеома». Природа . 437 (7062): 1173–8. Бибкод : 2005Natur.437.1173R . дои : 10.1038/nature04209 . ПМИД 16189514 . S2CID 4427026 .
- ^ Перейти обратно: а б с Мадер С., Ли Х., Пауза А., Соненберг Н. (сентябрь 1995 г.). «Фактор инициации трансляции eIF-4E связывается с общим мотивом, общим для фактора трансляции eIF-4 gamma и 4E-связывающих белков-репрессоров трансляции» . Мол. Клетка. Биол . 15 (9): 4990–7. дои : 10.1128/MCB.15.9.4990 . ПМК 230746 . ПМИД 7651417 .
- ^ Рао Р.Д., Младек А.С., Ламонт Дж.Д., Гобл Дж.М., Эрлихман С., Джеймс К.Д., Саркария Дж.Н. (октябрь 2005 г.). «Нарушение параллельных и сближающихся сигнальных путей способствует синергетическому противоопухолевому эффекту одновременного ингибирования mTOR и EGFR в клетках GBM» . Неоплазия . 7 (10): 921–9. дои : 10.1593/neo.05361 . ПМК 1502028 . ПМИД 16242075 .
- ^ Эгучи С., Токунага С., Хидаят С., Осиро Н., Ёсино К., Киккава У., Ёнезава К. (июль 2006 г.). «Различные роли мотивов TOS и RAIP белка-регулятора трансляции 4E-BP1 в ассоциации с хищником и фосфорилированием mTOR в регуляции размера клеток». Генные клетки . 11 (7): 757–66. дои : 10.1111/j.1365-2443.2006.00977.x . ПМИД 16824195 . S2CID 30113895 .
- ^ Ян Д., Брунн Г.Дж., Лоуренс Дж.К. (июнь 1999 г.). «Мутационный анализ сайтов регулятора трансляции PHAS-I, которые избирательно фосфорилируются mTOR» . ФЭБС Летт . 453 (3): 387–90. дои : 10.1016/s0014-5793(99)00762-0 . ПМИД 10405182 . S2CID 5023204 .
- ^ Патель Дж., Маклеод Л.Е., Врис Р.Г., Флинн А., Ван Х., Прауд К.Г. (июнь 2002 г.). «Клеточный стресс глубоко подавляет синтез белка и модулирует состояние фосфорилирования нескольких факторов трансляции» . Евро. Дж. Биохим . 269 (12): 3076–85. дои : 10.1046/j.1432-1033.2002.02992.x . ПМИД 12071973 .
- ^ Перейти обратно: а б Кумар В., Сабатини Д., Панди П., Гинграс А.С., Маджумдер П.К., Кумар М., Юань З.М., Кармайкл Г., Вейхсельбаум Р., Соненберг Н., Куфе Д., Харбанда С. (апрель 2000 г.). «Регуляция рапамицина и FKBP-мишени 1/мишени рапамицина для млекопитающих и кэп-зависимой инициации трансляции протеин-тирозинкиназой c-Abl» . Ж. Биол. Хим . 275 (15): 10779–87. дои : 10.1074/jbc.275.15.10779 . ПМИД 10753870 .
- ^ Кумар В., Панди П., Сабатини Д., Кумар М., Маджумдер П.К., Бхарти А., Кармайкл Г., Куфе Д., Харбанда С. (март 2000 г.). «Функциональное взаимодействие между RAFT1/FRAP/mTOR и протеинкиназой cdelta в регуляции кэп-зависимой инициации трансляции» . Я ЕСМЬ Дж . 19 (5): 1087–97. дои : 10.1093/emboj/19.5.1087 . ПМК 305647 . ПМИД 10698949 .
- ^ Гинграс А.С., Гиги С.П., Раф Б., Полакевич Р.Д., Абрахам Р.Т., Хоекстра М.Ф., Эберсолд Р., Соненберг Н. (июнь 1999 г.). «Регуляция фосфорилирования 4E-BP1: новый двухэтапный механизм» . Генс Дев . 13 (11): 1422–37. дои : 10.1101/gad.13.11.1422 . ПМК 316780 . ПМИД 10364159 .
- ^ Шен X, Томоо К., Утияма С., Кобаяши Ю., Исида Т. (октябрь 2001 г.). «Структурное и термодинамическое поведение эукариотического фактора инициации 4E в супрамолекулярном образовании с 4E-связывающим белком 1 и аналогом кэпа мРНК, изученное спектроскопическими методами» . хим. Фарм. Бык . 49 (10): 1299–303. дои : 10.1248/cpb.49.1299 . ПМИД 11605658 .
- ^ Адегоке О.А., Шевалье С., Мораис Дж.А., Гужон Р., Кимбалл С.Р., Джефферсон Л.С., Винг СС, Марлисс Э.Б. (январь 2009 г.). «Зажим в состоянии сытости стимулирует клеточные механизмы анаболизма мышечных белков и модулирует утилизацию глюкозы у нормальных мужчин» . Являюсь. Дж. Физиол. Эндокринол. Метаб . 296 (1): E105–13. дои : 10.1152/ajpendo.90752.2008 . ПМК 2636991 . ПМИД 18957614 .
- ^ Пауза А., Белшам Г.Дж., Гинграс А.С., Донзе О., Лин Т.А., Лоуренс Дж.К., Соненберг Н. (октябрь 1994 г.). «Инсулинзависимая стимуляция синтеза белка путем фосфорилирования регулятора функции 5'-кэпа». Природа . 371 (6500): 762–7. Бибкод : 1994Natur.371..762P . дои : 10.1038/371762a0 . ПМИД 7935836 . S2CID 4360955 .
- ^ Клейн М., Шепер Г.К., Уилсон М.Л., Ти А.Р., Гордый К.Г. (декабрь 2002 г.). «Локализация и регуляция eIF4E-связывающего белка 4E-BP3». ФЭБС Летт . 532 (3): 319–23. дои : 10.1016/s0014-5793(02)03694-3 . ПМИД 12482586 . S2CID 24527449 .
- ^ Пулен Ф., Гинграс А.С., Олсен Х., Шевалье С., Зоненберг Н. (май 1998 г.). «4E-BP3, новый член семейства белков, связывающих фактор инициации эукариот 4E» . Ж. Биол. Хим . 273 (22): 14002–7. дои : 10.1074/jbc.273.22.14002 . ПМИД 9593750 .
- ^ Дости Дж., Феррайуоло М., Пауза А., Адам С.А., Соненберг Н. (июнь 2000 г.). «Новый челночный белок 4E-T опосредует ядерный импорт белка, связывающего 5'-кэп мРНК, eIF4E» . ЭМБО Дж . 19 (12): 3142–56. дои : 10.1093/emboj/19.12.3142 . ПМК 203362 . ПМИД 10856257 .
- ^ Вари Т.К., Джефферсон Л.С., Кимбалл С.Р. (декабрь 1999 г.). «Аминокислотная стимуляция инициации трансляции в скелетных мышцах крыс». Являюсь. Дж. Физиол . 277 (6, часть 1): E1077–86. дои : 10.1152/ajpendo.1999.277.6.E1077 . ПМИД 10600798 . S2CID 4516850 .
- ^ Харрис Т.Е., Чи А., Шабановиц Дж., Хант Д.Ф., Роудс Р.Э., Лоуренс Дж.К. (апрель 2006 г.). «mTOR-зависимая стимуляция ассоциации eIF4G и eIF3 инсулином» . ЭМБО Дж . 25 (8): 1659–68. дои : 10.1038/sj.emboj.7601047 . ПМЦ 1440840 . ПМИД 16541103 .
- ^ Гради А., Иматака Х., Свиткин Ю.В., Ром Е., Раут Б., Морино С., Соненберг Н. (январь 1998 г.). «Новый функциональный фактор инициации трансляции эукариот человека 4G» . Мол. Клетка. Биол . 18 (1): 334–42. дои : 10.1128/mcb.18.1.334 . ПМЦ 121501 . ПМИД 9418880 .
- ^ Осборн М.Дж., Вольпон Л., Мемарпур-Язди М., Пиллэй С., Тамбипиллай А., Чарнота С., Чулкович-Кралячич Б., Трахан С., Оффингер М., Коулинг В.Х., Борден К.Л. (март 2022 г.). «Идентификация и характеристика взаимодействия между ферментом созревания кэпа метил-7-гуанозина RNMT и кэп-связывающим белком eIF4E» . Журнал молекулярной биологии . 434 (5): 167451. doi : 10.1016/j.jmb.2022.167451 . ПМЦ 9288840 . ПМИД 35026230 .
Дальнейшее чтение
[ редактировать ]- Джайн С., Хури Ф.Р., Шин Д.М. (2004). «Профилактика рака головы и шеи: современное состояние и перспективы». Текущие проблемы рака . 28 (5): 265–86. doi : 10.1016/j.currproblcancer.2004.05.003 . ПМИД 15375804 .
- Цулькович Б, Тописирович И, Борден К.Л. (2007). «Контроль экспрессии генов посредством регулонов РНК: роль эукариотического фактора инициации трансляции eIF4E» . Клеточный цикл . 6 (1): 65–9. дои : 10.4161/cc.6.1.3688 . ПМИД 17245113 .
- Малис Н., Маккарти Дж. Э. (2010). «Инициация перевода: можно предвидеть изменения в механизме» . Клеточные и молекулярные науки о жизни . 68 (6): 991–1003. дои : 10.1007/s00018-010-0588-z . ПМЦ 11115079 . ПМИД 21076851 . S2CID 31720000 .
Внешние ссылки
[ редактировать ]- Кэп-зависимая инициация трансляции от Nature Reviews Microbiology . Хорошее изображение и обзор функции факторов инициации.
галерея PDB |
|---|






Эта статья включает текст из Национальной медицинской библиотеки США , который находится в свободном доступе .