eIF2
Фактор инициации эукариот 2 ( eIF2 ) является фактором инициации эукариот . Он необходим для большинства форм инициации эукариотической трансляции . eIF2 опосредует связывание тРНК i Из к рибосоме GTP -зависимым образом. eIF2 представляет собой гетеротример, состоящий из альфа- субъединицы (также называемой субъединицей 1, EIF2S1), бета- субъединицы (субъединицы 2, EIF2S2) и гамма- субъединицы (субъединицы 3, EIF2S3).
После завершения фазы инициации eIF2 высвобождается из рибосомы, связанной с GDP, в виде неактивного бинарного комплекса. Для участия в следующем раунде инициации трансляции этот ВВП необходимо обменять на GTP.
Функция
[ редактировать ]
eIF2 является важным фактором синтеза белка, образующего тройной комплекс (ТС) с GTP и инициатором Met - тРНК i. Из . После своего образования ТК связывается с субъединицей рибосомы 40S, образуя преинициаторный комплекс 43S (43S PIC). , сборка 43S PIC, как полагают, стимулируется факторами инициации eIF1 , eIF1A и комплексом eIF3 По данным экспериментов in vitro . Затем 43S PIC связывает мРНК , которая ранее была раскручена комплексом eIF4F . Белки 43S PIC и eIF4F образуют новый комплекс 48S на мРНК, который начинает поиск по мРНК стартового кодона (AUG). При спаривании оснований кодона AUG с Met-тРНК eIF5 (который представляет собой белок, активирующий ГТФазу , или GAP) рекрутируется в комплекс и индуцирует eIF2 гидролизовать свой GTP. Это приводит к высвобождению eIF2-GDP из этого комплекса 48S, и трансляция начинается после рекрутирования рибосомальной субъединицы 60S и образования инициирующего комплекса 80S . Наконец, с помощью фактора обмена гуаниновых нуклеотидов (GEF) eIF2B , [ 1 ] ВВП в eIF2 обменивается на GTP, а тройные комплексные реформы — на новый раунд инициации трансляции. [ 2 ] [ 3 ] [ 4 ]
Структура
[ редактировать ]eIF2 представляет собой гетеротример с общей молярной массой 126 кДа , который состоит из трех субъединиц: α (субъединица 1), β (субъединица 2) и γ (субъединица 3). Последовательности всех трех субъединиц высококонсервативны (парные аминокислотные идентичности для каждой субъединицы составляют от 47 до 72% при сравнении белков Homo sapiens и Saccharomyces cerevisiae ).
| подразделение | Альфа | Бета | Гамма |
|---|---|---|---|
| Молекулярный вес / кДа | 36 | 38 | 52 |
| Сходство | семейство eIF2-альфа ИПР011488 |
семейство eIF2-бета / eIF5 ИПР002735 |
Семейство факторов элонгации, связывающих GTP (и другие) P41091 |
| Взаимодействия | Связывание eIF5, eIF2B и РНК | Связывание GTP и РНК |
α -субъединица содержит основную мишень фосфорилирования — серин в положении 51. Она также содержит домен мотива S1, который является потенциальным сайтом связывания РНК. Следовательно, α-субъединицу можно считать регуляторной субъединицей тримера.
содержит β-субъединица несколько сайтов фосфорилирования (остатки 2, 13, 67, 218). Важно учитывать, что в N-концевом домене (NTD) также имеются три лизиновых кластера, которые важны для взаимодействия с eIF2B. Более того, последовательность белка содержит мотив цинкового пальца , который, как было показано, играет роль как в образовании тройного комплекса, так и в образовании преинициаторного комплекса 43S. Существуют также две последовательности, связывающие гуаниновые нуклеотиды, которые, как было показано, не участвуют в регуляции активности eIF2. Считается также, что β-субъединица взаимодействует как с тРНК, так и с мРНК.
включает γ-субъединица три сайта связывания гуаниновых нуклеотидов и, как известно, является основным сайтом стыковки GTP/GDP. Он также содержит полость, связывающую тРНК, что было обнаружено с помощью рентгеновской кристаллографии . Цинковая костяшка способна связывать один Zn. 2+ катион. [ 4 ] [ 6 ] [ 7 ] Это связано с некоторыми факторами удлинения, такими как EF-Tu . [ 8 ]
Регулирование
[ редактировать ]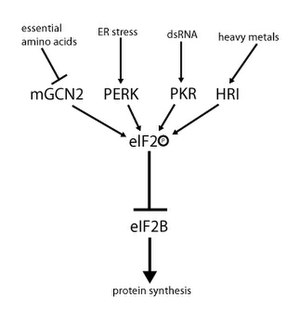
Активность eIF2 регулируется механизмом, включающим как обмен гуаниновых нуклеотидов, так и фосфорилирование. Фосфорилирование происходит в α-субъединице, которая является мишенью для ряда серинкиназ , фосфорилирующих серин 51. Эти киназы действуют в результате стресса, такого как недостаток аминокислот ( GCN2 ), стресс ER ( PERK ), присутствие dsRNA ( PKR ) дефицит гема ( HRI ) или интерферона . [ 10 ] После фосфорилирования eIF2 проявляет повышенное сродство к eIF2B, его GEF. Однако eIF2B способен обменивать GDP на GTP только в том случае, если eIF2 находится в нефосфорилированном состоянии. Однако фосфорилированный eIF2 из-за более сильного связывания действует как ингибитор собственного GEF (eIF2B). Поскольку клеточная концентрация eIF2B намного ниже, чем у eIF2, даже небольшое количество фосфорилированного eIF2 может полностью отменить активность eIF2B путем секвестрации. Без GEF eIF2 больше не может быть возвращен в активное (связанное с GTP) состояние. Как следствие, трансляция останавливается, поскольку инициация больше невозможна без наличия тройного комплекса. Кроме того, низкая концентрация тройного комплекса позволяет экспрессировать GCN4 (голодное состояние), что, в свою очередь, приводит к повышенной активации генов синтеза аминокислот. [ 2 ] [ 3 ] [ 4 ] [ 9 ] [ 11 ]
Болезнь
[ редактировать ]Поскольку eIF2 необходим для большинства форм инициации трансляции и, следовательно, синтеза белка, дефекты eIF2 часто приводят к летальному исходу. Белок высококонсервативен среди эволюционно отдаленных видов, что указывает на большое влияние мутаций на жизнеспособность клеток. Следовательно, никаких заболеваний, непосредственно связанных с мутациями eIF2, наблюдаться не может. Однако существует множество заболеваний, вызванных пониженной регуляцией eIF2 через его вышестоящие киназы. Например, повышенные концентрации активной PKR и неактивного (фосфорилированного) eIF2 были обнаружены у пациентов с нейродегенеративными заболеваниями, такими как болезнь Альцгеймера , Паркинсона и болезнь Хантингтона . Существует также один доказанный пример заболевания, связанного с ГЭФ eIF2B. Мутации во всех пяти субъединицах eIF2B связаны с болезнью исчезновения белого вещества (VWM), генетической лейкодистрофией, которая приводит к дегенерации и исчезновению белого вещества мозга. [ 12 ] [ 13 ] До сих пор не до конца понятно, почему эти дефекты поражают только клетки мозга. Потенциально сниженные уровни нестабильных регуляторных белков могут играть роль в развитии упомянутых заболеваний. [ 4 ] [ 14 ]
См. также
[ редактировать ]- Факторы инициации эукариот
- Киназы eIF2
- eIF2A
- eIF2D
- Лейкоэнцефалопатия с исчезновением белого вещества
Ссылки
[ редактировать ]- ^ eIF2B состоит из субъединиц EIF2B1 , EIF2B2 , EIF2B3 , EIF2B4 , EIF2B5.
- ^ Jump up to: а б Кимбалл С.Р. (1999). «Фактор инициации эукариот eIF2». Межд. Дж. Биохим. Клеточная Биол. 31 (1): 25–9. дои : 10.1016/S1357-2725(98)00128-9 . ПМИД 10216940 .
- ^ Jump up to: а б с Херши Дж.В. (1989). «Фосфорилирование белков контролирует скорость трансляции» (PDF) . Ж. Биол. хим. 264 (35): 20823–6. дои : 10.1016/S0021-9258(19)30005-5 . ПМИД 2687263 .
- ^ Jump up to: а б с д Хиннебуш АГ (2005). «Регуляция трансляции GCN4 и общий аминокислотный контроль дрожжей». Анну. Преподобный Микробиол. 59 : 407–50. дои : 10.1146/annurev.micro.59.031805.133833 . ПМИД 16153175 .
- ^ Кимбалл С.Р., Джефферсон Л.С. (2004). «Аминокислоты как регуляторы экспрессии генов» . Нутр. Метаб. 1 (1): 3. дои : 10.1186/1743-7075-1-3 . ПМК 524028 . ПМИД 15507151 .
- ^ Ролл-Мекак А., Один П., Цао С., Девер Т.Е., Берли С.К. (2004). «Рентгеновская структура фактора инициации трансляции eIF2gamma: значение для связывания тРНК и eIF2alpha» . Ж. Биол. хим. 279 (11): 10634–42. дои : 10.1074/jbc.M310418200 . PMID 14688270 .
- ^ Ито Т, Маринчев А, Вагнер Г (2004). «Структура раствора человеческого фактора инициации eIF2alpha обнаруживает гомологию с фактором элонгации eEF1B» . Структура . 12 (9): 1693–704. дои : 10.1016/j.str.2004.07.010 . ПМИД 15341733 .
- ^ Шмитт, Э; Бланке, С; Мечулам, Ю. (2 апреля 2002 г.). «Большая субъединица фактора инициации aIF2 является близким структурным гомологом факторов элонгации» . Журнал ЭМБО . 21 (7): 1821–32. дои : 10.1093/emboj/21.7.1821 . ПМК 125960 . ПМИД 11927566 .
- ^ Jump up to: а б Ника Дж., Риппель С., Ханниг Э.М. (2001). «Биохимический анализ гамма-комплекса eIF2beta выявил структурную функцию eIF2alpha в катализируемом обмене нуклеотидов» . Ж. Биол. хим. 276 (2): 1051–6. дои : 10.1074/jbc.M007398200 . ПМИД 11042214 .
- ^ Сэмюэл CE (1979). «Механизм действия интерферона: фосфорилирование фактора инициации синтеза белка eIF-2 в обработанных интерфероном клетках человека за счет специфичности сайта процессинга рибосом-ассоциированной киназы, аналогичной киназе ретикулоцитов кролика, регулируемой гемином» . Proc Natl Acad Sci США . 76 (2): 600–4. Бибкод : 1979ПНАС...76..600С . дои : 10.1073/pnas.76.2.600 . ПМК 382996 . ПМИД 284384 .
- ^ Хоуп А, Струл К (1987). «GCN4, белок-активатор транскрипции эукариот, связывается в виде димера с целевой ДНК» . Журнал ЭМБО . 6 (9): 2781–2784. дои : 10.1002/j.1460-2075.1987.tb02573.x . ПМЦ 553703 . ПМИД 3678204 .
- ^ Кнаап, Марджо С. ван дер; Пронк, Ян К.; Шепер, Герт К. (1 мая 2006 г.). «Исчезающая болезнь белого вещества» . Ланцет Неврология . 5 (5): 413–423. дои : 10.1016/S1474-4422(06)70440-9 . ISSN 1474-4422 . ПМИД 16632312 . S2CID 44301370 .
- ^ Лигуотер, Питер Эй Джей; Вермюлен, Герр; Кёнст, Андреа AM; Найду, Саккубай; Малдерс, Джойс; Виссер, Аллердин; Керсберген, Паула; Мобах, Драгош; Фондс, Дафна; ван Беркель, Карола ГМ; Леммерс, Ричард JLF (декабрь 2001 г.). «Субъединицы фактора инициации трансляции eIF2B являются мутантными при лейкоэнцефалопатии с исчезновением белого вещества» . Природная генетика . 29 (4): 383–388. дои : 10.1038/ng764 . ISSN 1546-1718 . ПМИД 11704758 . S2CID 20313523 .
- ^ Чанг Р.К., Ю М.С., Лай К.С. (2006). «Значение молекулярной передачи сигналов для контроля трансляции белков при нейродегенеративных заболеваниях» . Нейросигналы . 15 (5): 249–58. дои : 10.1159/000102599 . ПМИД 17496426 .
Внешние ссылки
[ редактировать ]- EIF-2 в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)
- Кэп-зависимая инициация трансляции от Nature Reviews Microbiology. Хорошее изображение и обзор функции факторов инициации