ФМР1
| ФМР1 | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Идентификаторы | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | FMR1 , FMRP, FRAXA, POF, POF1, ломкий X умственная отсталость 1, регулятор трансляции FMRP 1, ломкий X-мессенджер рибонуклеопротеин 1 | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | Опустить : 309550 ; МГИ : 95564 ; Гомологен : 1531 ; Генные карты : FMR1 ; ОМА : FMR1 — ортологи | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||






FMR1 ( хрупкий рибонуклеопротеин 1-мессенджер ) — человеческий ген. [ 5 ] который кодирует белок, называемый хрупким рибонуклеопротеином-мессенджером X , или FMRP. [ 6 ] Этот белок, чаще всего встречающийся в мозге, необходим для нормального когнитивного развития и женской репродуктивной функции. Мутации этого гена могут привести к синдрому хрупкой Х-хромосомы , умственной отсталости , преждевременной недостаточности яичников , аутизму , болезни Паркинсона , задержке развития и другим когнитивным нарушениям. [ 7 ] Премутация FMR1 связана с широким спектром клинических фенотипов , от которых страдают более двух миллионов человек во всем мире. [ 8 ]
Функция
[ редактировать ]Синаптическая пластичность
[ редактировать ]FMRP имеет разнообразный набор функций в разных областях нейрона; однако эти функции не были полностью охарактеризованы. Было высказано предположение, что FMRP играет роль в нуклеоцитоплазматическом перемещении мРНК , дендритной локализации мРНК и синтезе синаптического белка . [ 9 ] Исследования синдрома хрупкой X-хромосомы значительно помогли понять функциональность FMRP благодаря наблюдаемым эффектам потери FMRP на нейроны. Мышиная модель Fragile X Messenger Ribonucleoprotein показала участие FMRP в синаптической пластичности . [ 10 ] Синаптическая пластичность требует производства новых белков в ответ на активацию синаптических рецепторов . Предполагается, что именно производство белков в ответ на стимуляцию обеспечивает постоянные физические изменения и измененные синаптические связи, связанные с процессами обучения и памяти.
метаботропного глутаматного рецептора группы 1 Передача сигналов (mGluR) играет важную роль в FMRP-зависимой синаптической пластичности. Постсинаптическая стимуляция mGluR приводит к усилению синтеза белка через систему вторичных мессенджеров . [ 11 ] Роль mGluR в синаптической пластичности дополнительно подтверждается наблюдением удлинения дендритных шипов после стимуляции mGluR. [ 12 ] Кроме того, активация mGluR приводит к синтезу FMRP вблизи синапсов . Образующийся FMRP связывается с полирибосомальными комплексами после стимуляции mGluR, что позволяет предположить участие Fragile X Messenger Ribonucleoprotein в процессе трансляции . Это еще раз подтверждает роль FMRP в синтезе синаптических белков и росте синаптических связей. [ 13 ] Потеря FMRP приводит к аномальному фенотипу дендритных шипов . В частности, удаление гена FMR1 у мышей привело к увеличению количества синапсов в позвоночнике. [ 14 ]
Роль в переводе
[ редактировать ]Предлагаемый механизм влияния FMRP на синаптическую пластичность заключается в его роли негативного регулятора трансляции. FMRP представляет собой РНК-связывающий белок, который связывается с полирибосомами . [ 13 ] [ 15 ] Способность FMRP связываться с РНК зависит от его доменов KH и боксов RGG . Домен KH представляет собой консервативный мотив, который характеризует многие РНК-связывающие белки. Мутагенез этого домена привел к нарушению связывания FMRP с РНК. [ 16 ]
Было показано, что FMRP ингибирует трансляцию мРНК. Мутация белка FMRP привела к неспособности подавлять трансляцию, в отличие от аналога дикого типа, который был способен это делать. [ 17 ] Как упоминалось ранее, стимуляция mGluR связана с повышением уровня белка FMRP. Кроме того, стимуляция mGluR приводит к увеличению уровней мРНК-мишеней FMRP. Исследование показало, что базальные уровни белков, кодируемых этими целевыми мРНК, значительно повышены и неправильно регулируются у мышей с дефицитом FMRP. [ 18 ]
Репрессия трансляции FMRP действует путем ингибирования инициации трансляции. FMRP напрямую связывает CYFIP1 , который, в свою очередь, связывает фактор инициации трансляции eIF4E . Комплекс FMRP-CYFIP1 запрещает eIF4E-зависимую инициацию, тем самым подавляя трансляцию. [ 19 ] Применительно к наблюдаемому фенотипу при синдроме хрупкой Х-хромосомы избыточные уровни белка и снижение контроля трансляции можно объяснить потерей репрессии трансляции с помощью FMRP при синдроме хрупкой Х-хромосомы. [ 19 ] [ 20 ] FMRP контролирует трансляцию большой группы целевых мРНК; однако степень трансляционного контроля FMRP неизвестна. Было показано, что белок подавляет трансляцию мРНК-мишеней в синапсах, в том числе тех, которые кодируют белки цитоскелета Arc/Arg3.1 и MAP1B , а также киназу CaM II . [ 21 ] Кроме того, FMRP связывает мРНК PSD-95 и GluR1/2. Важно отметить, что эти FMRP-связывающие мРНК играют значительную роль в пластичности нейронов.
Было показано, что контроль трансляции FMRP регулируется передачей сигналов mGluR. Стимуляция mGluR может привести к транспортировке комплексов мРНК в синапсы для локального синтеза белка. Было показано, что гранулы FMRP локализуются с мРНК MAP1B и рибосомальной РНК в дендритах, что позволяет предположить, что этот комплекс в целом, возможно, потребуется транспортировать в дендриты для локального синтеза белка. Кроме того, было обнаружено, что микротрубочки являются необходимым компонентом для mGluR-зависимой транслокации FMRP в дендриты. [ 9 ] FMRP может играть дополнительную роль в локальном синтезе белка, помогая в ассоциации груза мРНК и микротрубочек. [ 22 ] Таким образом, FMRP способен регулировать эффективность транспорта, а также репрессию трансляции во время транспорта. Наконец, синтез FMRP, убиквитинирование и протеолиз происходят быстро в ответ на передачу сигналов mGluR, что указывает на чрезвычайно динамичную роль регулятора трансляции. [ 18 ]
Экспрессия генов
[ редактировать ]Ген FMR1 расположен на Х-хромосоме и содержит повторяющийся тринуклеотид CGG. У большинства людей сегмент CGG повторяется примерно 5–44 раза. Более высокое количество повторов сегмента CGG связано с нарушением когнитивной и репродуктивной функции. Если у человека имеется 45-54 повтора, это считается «серой зоной» или пограничным риском, 55-200 повторов называется премутацией, а более 200 повторов считается полной мутацией гена FMR1 по данным Американского колледжа медицинской генетики. и геномика. [ 23 ] Первая полная последовательность ДНК расширения повтора у человека с полной мутацией была получена учеными в 2012 году с помощью секвенирования SMRT . [ 24 ] Это пример нарушения тринуклеотидных повторов . Расширение тринуклеотидных повторов , вероятно, является следствием проскальзывания цепи либо во время репарации ДНК , либо во время репликации ДНК . [ 25 ]
FMRP представляет собой белок, связывающий хроматин , который участвует в ответной реакции на повреждение ДНК . [ 26 ] [ 27 ] FMRP занимает участки на мейотических хромосомах и регулирует динамику механизма реакции на повреждение ДНК во время сперматогенеза . [ 26 ]
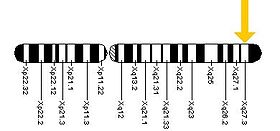
Ген FMR1 можно найти на длинном (q) плече Х-хромосомы в положении 27,3, от пары оснований 146 699 054 до пары оснований 146 738 156.
Сопутствующие условия
[ редактировать ]Синдром ломкой Х-хромосомы
[ редактировать ]Почти все случаи синдрома ломкой Х-хромосомы вызваны экспансией тринуклеотидного повтора CGG в гене FMR1 . В этих случаях CGG аномально повторяется от 200 до более 1000 раз. В результате эта часть гена FMR1 метилируется, что заставляет ген молчать (он отключается и не производит никакого белка). Без адекватного FMR1 могут развиться серьезные нарушения обучаемости или интеллектуальные нарушения, а также физические отклонения, наблюдаемые при синдроме хрупкой Х-хромосомы.
Менее 1% всех случаев синдрома хрупкой Х-хромосомы вызваны мутациями , которые удаляют часть или весь ген FMR1 или изменяют пару оснований, что приводит к изменению одной из аминокислот в гене. Эти мутации нарушают трехмерную форму FMRP или препятствуют синтезу белка, что приводит к появлению признаков и симптомов синдрома ломкой Х-хромосомы.
Последовательность CGG в гене FMR1 , повторяющаяся от 55 до 200 раз, называется премутацией. Хотя большинство людей с премутацией интеллектуально нормальны, некоторые из этих людей имеют легкие версии физических особенностей, наблюдаемых при синдроме хрупкой Х-хромосомы (например, оттопыренные уши), и могут испытывать проблемы с психическим здоровьем, такие как тревога или депрессия.
Синдром тремора/атаксии, связанный с ломкой Х-хромосомой
[ редактировать ]Премутации связаны с повышенным риском синдрома хрупкого Х-ассоциированного тремора/атаксии (FXTAS). FXTAS характеризуется атаксией (потерей координации), тремором , потерей памяти , потерей чувствительности в нижних конечностях ( периферическая невропатия ), а также психическими и поведенческими изменениями. Заболевание обычно развивается в позднем возрасте.
Преждевременное старение яичников
[ редактировать ]Ген FMR1 играет очень важную роль в функции яичников, независимо от когнитивных/неврологических эффектов. Незначительные расширения повторов CGG, которые не вызывают синдром ломкой Х-хромосомы , связаны с повышенным риском преждевременного старения яичников , также называемого скрытой первичной недостаточностью яичников, состояния, при котором женщины преждевременно истощают свою функцию яичников. [ 28 ] [ 29 ] [ 30 ]
Синдром поликистозных яичников
[ редактировать ]Было обнаружено, что очень специфический субгенотип FMR1 связан с синдромом поликистозных яичников (СПКЯ). Экспрессия гена, называемая гетерозиготной-нормальной/низкой, может вызывать чрезмерную активность фолликулов, подобную СПКЯ, и гиперактивную функцию яичников в более молодом возрасте.
Взаимодействия
[ редактировать ]FMR1 Было показано, что взаимодействует с:
- CYFIP1 , [ 31 ]
- CYFIP2 , [ 31 ] [ 32 ]
- FXR1 , [ 33 ] [ 34 ] и
- ФСР2 , [ 33 ] [ 34 ] [ 35 ]
- НУФИП1 , [ 32 ] [ 36 ] и
- НУФИП2 . [ 32 ]
Ссылки
[ редактировать ]- ^ Jump up to: а б с GRCh38: Версия Ensembl 89: ENSG00000102081 – Ensembl , май 2017 г.
- ^ Jump up to: а б с GRCm38: выпуск Ensembl 89: ENSMUSG00000000838 – Ensembl , май 2017 г.
- ^ «Ссылка на Human PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Ссылка на Mouse PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ Веркерк А.Дж., Пьеретти М., Сатклифф Дж.С., Фу Ю.Х., Куль Д.П., Пиццути А., Райнер О., Ричардс С., Виктория М.Ф., Чжан Ф.П. (май 1991 г.). «Идентификация гена (FMR-1), содержащего повтор CGG, совпадающий с областью кластера точки разрыва, демонстрирующей изменение длины при синдроме хрупкой X». Клетка . 65 (5): 905–14. дои : 10.1016/0092-8674(91)90397-H . ПМИД 1710175 . S2CID 21463845 .
- ^ Верхей К., Баккер С.Э., де Грааф Э., Кеулеманс Дж., Виллемсен Р., Веркерк А.Дж., Гальяард Х., Ройзер А.Дж., Хогевен А.Т., Оостра Б.А. (июнь 1993 г.). «Характеристика и локализация продукта гена FMR-1, связанного с синдромом ломкой Х-хромосомы» . Природа . 363 (6431): 722–4. Бибкод : 1993Natur.363..722V . дои : 10.1038/363722a0 . hdl : 1765/56659 . ПМИД 8515814 . S2CID 4331494 .
- ^ «Хрупкий X-мессенджер рибонуклеопротеин 1» Справочник генов человека
- ^ Мила М., Родригес-Ревенга Л., Матилья-Дуэньяс А. (октябрь 2016 г.). «Премутация FMR1: основные механизмы и клиническое участие». Мозжечок . 15 (5): 543–5. дои : 10.1007/s12311-016-0808-7 . ПМИД 27338822 . S2CID 16002209 .
- ^ Jump up to: а б Антар Л.Н., Диктенберг Дж.Б., Плочиняк М., Афроз Р., Басселл Г.Дж. (август 2005 г.). «Локализация гранул мРНК, связанных с FMRP, и потребность микротрубочек для зависимого от активности транспорта в нейронах гиппокампа» . Гены, мозг и поведение . 4 (6): 350–9. дои : 10.1111/j.1601-183X.2005.00128.x . ПМИД 16098134 .
- ^ Хубер К.М., Галлахер С.М., Уоррен С.Т., Медведь М.Ф. (май 2002 г.). «Измененная синаптическая пластичность в мышиной модели хрупкой умственной отсталости X» . Труды Национальной академии наук Соединенных Штатов Америки . 99 (11): 7746–50. Бибкод : 2002PNAS...99.7746H . дои : 10.1073/pnas.122205699 . ПМЦ 124340 . ПМИД 12032354 .
- ^ Вейлер И.Дж., Гриноф В.Т. (август 1993 г.). «Метаботропные рецепторы глутамата запускают постсинаптический синтез белка» . Труды Национальной академии наук Соединенных Штатов Америки . 90 (15): 7168–71. Бибкод : 1993PNAS...90.7168W . дои : 10.1073/pnas.90.15.7168 . ПМК 47097 . ПМИД 8102206 .
- ^ Вандерклиш П.В., Эдельман Г.М. (февраль 2002 г.). «Дендритные шипы удлиняются после стимуляции метаботропных глутаматных рецепторов группы 1 в культивируемых нейронах гиппокампа» . Труды Национальной академии наук Соединенных Штатов Америки . 99 (3): 1639–44. Бибкод : 2002PNAS...99.1639V . дои : 10.1073/pnas.032681099 . ПМК 122243 . ПМИД 11818568 .
- ^ Jump up to: а б Вейлер И.Дж., Ирвин С.А., Клинцова А.Я., Спенсер С.М., Бразелтон А.Д., Мияширо К., Комери Т.А., Патель Б., Эбервин Дж., Гриноу В.Т. (май 1997 г.). «Хрупкий белок умственной отсталости X транслируется вблизи синапсов в ответ на активацию нейромедиатора» . Труды Национальной академии наук Соединенных Штатов Америки . 94 (10): 5395–400. Бибкод : 1997PNAS...94.5395W . дои : 10.1073/pnas.94.10.5395 . ПМК 24689 . ПМИД 9144248 .
- ^ Антар Л.Н., Ли С., Чжан Х., Кэрролл Р.К., Басселл Г.Дж. (2006). «Локальные функции FMRP в подвижности конуса роста аксонов и зависимой от активности регуляции филоподий и синапсов позвоночника». Молекулярная и клеточная нейронауки . 32 (1–2): 37–48. дои : 10.1016/j.mcn.2006.02.001 . ПМИД 16631377 . S2CID 18257268 .
- ^ Браун В., Смолл К., Лаккис Л., Фэн Ю., Гюнтер С., Уилкинсон К.Д., Уоррен С.Т. (июнь 1998 г.). «Очищенный рекомбинантный Fmrp демонстрирует селективное связывание РНК как внутреннее свойство хрупкого X-белка умственной отсталости» . Журнал биологической химии . 273 (25): 15521–7. дои : 10.1074/jbc.273.25.15521 . ПМИД 9624140 .
- ^ Сиоми Х., Чой М., Сиоми М.К., Нуссбаум Р.Л., Дрейфусс Дж. (апрель 1994 г.). «Основная роль доменов KH в связывании РНК: нарушение связывания РНК из-за мутации в домене KH FMR1, которая вызывает синдром хрупкой X». Клетка . 77 (1): 33–9. дои : 10.1016/0092-8674(94)90232-1 . PMID 8156595 . S2CID 35339859 .
- ^ Лаггербауэр Б., Остарек Д., Кейдель Э.М., Остарек-Ледерер А., Фишер Ю. (февраль 2001 г.). «Доказательства того, что хрупкий белок умственной отсталости X является негативным регулятором трансляции» . Молекулярная генетика человека . 10 (4): 329–38. дои : 10.1093/хмг/10.4.329 . ПМИД 11157796 .
- ^ Jump up to: а б Хоу Л., Антион, доктор медицинских наук, Ху Д., Спенсер СМ, Пэйлор Р., Кланн Э. (август 2006 г.). «Динамическая трансляционная и протеасомная регуляция хрупкого X-белка умственной отсталости контролирует mGluR-зависимую долговременную депрессию» . Нейрон . 51 (4): 441–54. дои : 10.1016/j.neuron.2006.07.005 . ПМИД 16908410 . S2CID 13915959 .
- ^ Jump up to: а б Наполи I, Меркальдо В, Бойл П.П., Элеутери Б, Зальфа Ф, Де Рубейс С, Ди Марино Д, Мор Е, Массими М, Фалькони М, Витке В, Коста-Маттиоли М, Зоненберг Н, Аксель Т, Баньи К (сентябрь) 2008). «Белок синдрома хрупкой X подавляет зависимую от активности трансляцию через CYFIP1, новый 4E-BP» . Клетка . 134 (6): 1042–54. дои : 10.1016/j.cell.2008.07.031 . ПМИД 18805096 . S2CID 14123165 .
- ^ Муддашетти Р.С., Келич С., Гросс С., Сюй М., Басселл Г.Дж. (май 2007 г.). «Дисрегулированная метаботропная глутамат-зависимая трансляция рецептора AMPA и мРНК постсинаптической плотности-95 в синапсах на мышиной модели синдрома ломкой Х-хромосомы» . Журнал неврологии . 27 (20): 5338–48. doi : 10.1523/JNEUROSCI.0937-07.2007 . ПМЦ 6672337 . ПМИД 17507556 .
- ^ Зальфа Ф, Георгий М, Примерано Б, Моро А, Ди Пента А, Рейс С, Оостра Б, Баньи С (февраль 2003 г.). «Белок синдрома хрупкой Х-хромосомы FMRP связывается с РНК BC1 и регулирует трансляцию специфических мРНК в синапсах» . Клетка . 112 (3): 317–27. дои : 10.1016/S0092-8674(03)00079-5 . ПМИД 12581522 . S2CID 14892764 .
- ^ Эстес П.С., О'Ши М., Класен С., Зарнеску, округ Колумбия (октябрь 2008 г.). «Хрупкий белок X контролирует эффективность транспорта мРНК в нейронах дрозофилы». Молекулярная и клеточная нейронауки . 39 (2): 170–9. дои : 10.1016/j.mcn.2008.06.012 . ПМИД 18655836 . S2CID 24629325 .
- ^ «Технические стандарты и рекомендации для хрупкого X» . Американский колледж медицинской генетики . 2000-10-02 . Проверено 29 марта 2013 г.
- ^ Лумис Э.В., Ид Дж.С., Пелусо П., Инь Дж., Хики Л., Ранк D, Маккалмон С., Хагерман Р.Дж., Тассоне Ф., Хагерман П.Дж. (январь 2013 г.). «Секвенирование несеквенируемого: расширенные аллели CGG-повторения хрупкого гена X» . Геномные исследования . 23 (1): 121–8. дои : 10.1101/гр.141705.112 . ПМК 3530672 . ПМИД 23064752 .
- ^ Усдин К., House NC, Freudenreich CH (2015). «Повторяющаяся нестабильность во время восстановления ДНК: данные модельных систем» . Крит. Преподобный Биохим. Мол. Биол . 50 (2): 142–67. дои : 10.3109/10409238.2014.999192 . ПМЦ 4454471 . ПМИД 25608779 .
- ^ Jump up to: а б Алпатов Р., Леш Б.Дж., Накамото-Киношита М., Бланко А., Чен С., Штютцер А., Армаш К.Дж., Саймон М.Д., Сюй С., Али М., Мурн Дж., Присич С., Кутателадзе Т.Г., Вакоч Ч.Р., Мин Дж., Кингстон Р.Э., Фишл В., Уоррен С.Т., Пейдж Д.К., Ши Ю. (май 2014 г.). «Зависимая от хроматина роль хрупкого X-белка умственной отсталости FMRP в реакции на повреждение ДНК» . Клетка . 157 (4): 869–81. дои : 10.1016/j.cell.2014.03.040 . ПМК 4038154 . ПМИД 24813610 .
- ^ Dockendorff TC, Лабрадор М (май 2018 г.). «Хрупкий белок X и функция генома». Мол. Нейробиол . 56 (1): 711–721. дои : 10.1007/s12035-018-1122-9 . ПМИД 29796988 . S2CID 44159474 .
- ^ Глейхер Н. , Барад Д.Х. (август 2010 г.). «Ген FMR1 как регулятор рекрутирования яичников и овариального резерва». Акушерско-гинекологический осмотр . 65 (8): 523–30. doi : 10.1097/OGX.0b013e3181f8bdda . ПМИД 20955631 . S2CID 26409068 .
- ^ Чаттерджи С., Майтра А., Кадам С., Патель З., Гокрал Дж., Мехерджи П. (август 2009 г.). «Определение размера повтора CGG в гене FMR1 у индийских женщин с преждевременной недостаточностью яичников» . Репродуктивная биомедицина онлайн . 19 (2): 281–6. дои : 10.1016/s1472-6483(10)60086-7 . ПМИД 19712568 .
- ^ Стреули I, Фрайсс Т., Ибечеол В., Муа И., Моррис М.А., де Зиглер Д. (август 2009 г.). «Промежуточные и премутационные аллели FMR1 у женщин со скрытой первичной недостаточностью яичников» . Фертильность и бесплодие . 92 (2): 464–70. doi : 10.1016/j.fertnstert.2008.07.007 . ПМИД 18973899 .
- ^ Jump up to: а б Шенк А., Бардони Б., Моро А., Баньи С., Мандель Дж.Л. (июль 2001 г.). «Высококонсервативное семейство белков, взаимодействующее с хрупким X-белком умственной отсталости (FMRP) и демонстрирующее избирательное взаимодействие с FMRP-родственными белками FXR1P и FXR2P» . Труды Национальной академии наук Соединенных Штатов Америки . 98 (15): 8844–9. Бибкод : 2001PNAS...98.8844S . дои : 10.1073/pnas.151231598 . ПМЦ 37523 . ПМИД 11438699 .
- ^ Jump up to: а б с Бардони Б., Кастетс М., Хуот М.Е., Шенк А., Адинолфи С., Корбин Ф., Пасторе А., Ханджян Э.В., Мандель Дж.Л. (июль 2003 г.). «82-FIP, новый белок, взаимодействующий с FMRP (хрупким Х-белком умственной отсталости), демонстрирует внутриклеточную локализацию, зависящую от клеточного цикла» . Молекулярная генетика человека . 12 (14): 1689–98. дои : 10.1093/hmg/ddg181 . ПМИД 12837692 .
- ^ Jump up to: а б Сиоми MC, Чжан Ю, Сиоми Х, Дрейфусс Дж (июль 1996 г.). «Специфические последовательности в белке FMR1 синдрома хрупкой X и белках FXR опосредуют их связывание с 60S рибосомальными субъединицами и взаимодействия между ними» . Молекулярная и клеточная биология . 16 (7): 3825–32. дои : 10.1128/mcb.16.7.3825 . ПМК 231379 . ПМИД 8668200 .
- ^ Jump up to: а б Чжан Ю, О'Коннор Дж.П., Сиоми М.К., Шринивасан С., Дутра А., Нуссбаум Р.Л., Дрейфус Г. (ноябрь 1995 г.). «Хрупкий белок синдрома умственной отсталости X взаимодействует с новыми гомологами FXR1 и FXR2» . Журнал ЭМБО . 14 (21): 5358–66. дои : 10.1002/j.1460-2075.1995.tb00220.x . ПМЦ 394645 . ПМИД 7489725 .
- ^ Семан С., Браун В., Уоррен С.Т. (декабрь 1999 г.). «Выделение FMRP-ассоциированной частицы рибонуклеопротеина-мессенджера и идентификация нуклеолина и хрупких X-родственных белков как компонентов комплекса» . Молекулярная и клеточная биология . 19 (12): 7925–32. дои : 10.1128/mcb.19.12.7925 . ПМЦ 84877 . ПМИД 10567518 .
- ^ Бардони Б., Шенк А., Мандель Дж.Л. (декабрь 1999 г.). «Новый РНК-связывающий ядерный белок, который взаимодействует с хрупким белком X умственной отсталости (FMR1)» . Молекулярная генетика человека . 8 (13): 2557–66. дои : 10.1093/hmg/8.13.2557 . ПМИД 10556305 .
Дальнейшее чтение
[ редактировать ]- Бассел Дж.Дж., Уоррен С.Т. (октябрь 2008 г.). «Синдром хрупкой Х-хромосомы: потеря локальной регуляции мРНК изменяет развитие и функцию синапсов» . Нейрон . 60 (2): 201–14. дои : 10.1016/j.neuron.2008.10.004 . ПМК 3691995 . ПМИД 18957214 .
- Хагерман П.Дж., Хагерман Р.Дж. (май 2004 г.). «Премутация хрупкого-X: перспектива зрелости» . Американский журнал генетики человека . 74 (5): 805–16. дои : 10.1086/386296 . ПМК 1181976 . ПМИД 15052536 .
- Хагерман Р.Дж., Ливитт Б.Р., Фарзин Ф., Жакмон С., Греко К.М., Брунберг Дж.А., Тассоне Ф., Хессл Д., Харрис С.В., Чжан Л., Джардини Т., Гейн Л.В., Ферранти Дж., Руис Л., Лихи М.А., Григсби Дж., Хагерман П.Дж. (май 2004 г.). «Синдром тремора/атаксии, связанный с ломким Х-хромосомой (FXTAS) у женщин с премутацией FMR1» . Американский журнал генетики человека . 74 (5): 1051–6. дои : 10.1086/420700 . ПМК 1181968 . ПМИД 15065016 .
- Жакмон С., Хагерман Р.Дж., Лихи М.А., Холл Д.А., Левин Р.А., Брунберг Дж.А., Чжан Л., Джардини Т., Гейн Л.В., Харрис С.В., Херман К., Григсби Дж., Греко СМ, Берри-Кравис Е., Тассоне Ф., Хагерман П.Дж. ( январь 2004 г.). «Пенетрантность синдрома хрупкого Х-ассоциированного тремора/атаксии в популяции носителей премутации» . ДЖАМА 291 (4): 460–9. дои : 10.1001/public.291.4.460 . ПМИД 14747503 .
- Джин П., Алиш Р.С., Уоррен С.Т. (ноябрь 2004 г.). «РНК и микроРНК при хрупкой умственной отсталости X». Природная клеточная биология . 6 (11): 1048–53. дои : 10.1038/ncb1104-1048 . ПМИД 15516998 . S2CID 7315938 .
- Джин П., Уоррен С.Т. (март 2003 г.). «Новое понимание синдрома хрупкой Х: от молекул к нейроповедению». Тенденции биохимических наук . 28 (3): 152–8. CiteSeerX 10.1.1.532.4162 . дои : 10.1016/S0968-0004(03)00033-1 . ПМИД 12633995 .
- О'Доннелл В.Т., Уоррен С.Т. (2002). «Десятилетие молекулярных исследований синдрома хрупкой Х». Ежегодный обзор неврологии . 25 : 315–38. дои : 10.1146/annurev.neuro.25.112701.142909 . ПМИД 12052912 .
- Остра Б.А., Кьюрацци П. (декабрь 2001 г.). «Хрупкий ген X и его функция». Клиническая генетика . 60 (6): 399–408. дои : 10.1034/j.1399-0004.2001.600601.x . ПМИД 11846731 . S2CID 40128119 .
- Оостра Б.А., Виллемсен Р. (октябрь 2003 г.). «Хрупкий баланс: уровни экспрессии FMR1» . Молекулярная генетика человека . 12 Спецификация № 2 (90002): R249–57. дои : 10.1093/hmg/ddg298 . ПМИД 12952862 .
- Никола Н.А., Меткалф Д. (октябрь 1991 г.). «Разнообразие субъединиц среди рецепторов гемопоэтического фактора роста». Клетка . 67 (1): 1–4. дои : 10.1016/0092-8674(91)90564-F . ПМИД 1913811 . S2CID 12696714 .
- Сильска Д., Милевски М., Бал Дж. (2003). «[Молекулярный патогенез синдрома ломкой Х-хромосомы]». Медицина возраста развития . 6 (4): 295–308. ПМИД 12810982 .
- Баньи С., Гриноф WT (май 2005 г.). «От торговли мРНП к дисморфогенезу позвоночника: корни синдрома хрупкой Х-хромосомы» . Обзоры природы. Нейронаука . 6 (5): 376–87. дои : 10.1038/nrn1667 . ПМИД 15861180 . S2CID 17374547 .
- Хубер К.М. (апрель 2006 г.). «Хрупкая связь X-мозжечка». Тенденции в нейронауках . 29 (4): 183–5. doi : 10.1016/j.tins.2006.02.001 . ПМИД 16500716 . S2CID 8674692 .
- Леш Д.З., Буй К.М., Диссанаяке С., Клиффорд С., Гулд Э., Булхак-Патерсон Д., Тассоне Ф., Тейлор А.К., Хессл Д., Хагерман Р., Хаггинс Р.М. (2007). «Молекулярные и когнитивные предикторы континуума аутистического поведения у хрупкого Х» . Неврологические и биоповеденческие обзоры . 31 (3): 315–26. doi : 10.1016/j.neubiorev.2006.09.007 . ПМК 2145511 . ПМИД 17097142 .
Внешние ссылки
[ редактировать ]галерея PDB |
|---|