Альфа-синуклеин
| СНКА | |||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| Идентификаторы | |||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | SNCA , NACP, PARK1, PARK4, PD1, синуклеин альфа | ||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | ОМИМ : 163890 ; МГИ : 1277151 ; Гомологены : 293 ; GeneCards : SNCA ; OMA : SNCA – ортологи | ||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||





Альфа-синуклеин ( aSyn ) — это белок , который у человека кодируется SNCA геном . [5] Альфа-синуклеин представляет собой нейрональный белок, который регулирует движение синаптических пузырьков и последующее высвобождение нейромедиаторов . [6] [7]
Его много в мозге, тогда как меньшие количества обнаруживаются в сердце, мышцах и других тканях. В головном мозге альфа-синуклеин обнаруживается главным образом в окончаниях аксонов пресинаптических нейронов . [5] Внутри этих терминалей альфа-синуклеин взаимодействует с фосфолипидами. [8] и белки. [5] [9] [10] Пресинаптические терминали высвобождают химические посланники, называемые нейротрансмиттерами, из отсеков, известных как синаптические пузырьки . Высвобождение нейротрансмиттеров передает сигналы между нейронами и имеет решающее значение для нормальной функции мозга. [5]
При болезни Паркинсона и других синуклеинопатиях нерастворимые формы альфа-синуклеина накапливаются в виде включений в тельцах Леви . [11]
Семейная болезнь Паркинсона связана с мутациями в гене -синуклеина ( SNCA ). В процессе зародышевой нуклеации альфа-синуклеин приобретает поперечно-листовую структуру, аналогичную другим амилоидам. [12]
Белок альфа-синуклеина человека состоит из 140 аминокислот. [13] [14] [15] Фрагмент альфа-синуклеина, известный как неамилоидный бета -компонент (NAC) болезни Альцгеймера амилоида , первоначально обнаруженный во фракции, обогащенной амилоидом, оказался фрагментом белка-предшественника NACP. [13] Позже было установлено, что NACP является человеческим гомологом синуклеина электрических скатов рода Torpedo . Поэтому NACP теперь называют человеческим альфа-синуклеином. [16]
Экспрессия тканей [ править ]
Альфа-синуклеин — это синуклеин белок- , который в основном встречается в нервной ткани и составляет до одного процента всех белков в цитозоле клеток головного мозга . [17] Он экспрессируется в высокой степени в нейронах лобной коры , гиппокампа , полосатого тела и обонятельной луковицы . [17] но также может быть обнаружен в ненейрональных глиальных клетках . [18] В меланоцитах экспрессия белка SNCA может регулироваться транскрипционным фактором, ассоциированным с микрофтальмией (MITF). [19]
Установлено, что альфа-синуклеин широко локализован в ядре нейронов головного мозга млекопитающих, что позволяет предположить роль альфа-синуклеина в ядре. [20] Однако синуклеин обнаруживается преимущественно в пресинаптических окончаниях как в свободной, так и в мембраносвязанной форме. [21] при этом примерно 15% синуклеина в любой момент нейронов находится в мембраносвязанном состоянии. [22]
Было также показано, что альфа-синуклеин локализуется в митохондриях нейронов . [23] [24] Альфа-синуклеин высоко экспрессируется в митохондриях обонятельной луковицы , гиппокампе, полосатом теле и таламусе, где также богат цитозольный альфа-синуклеин. Однако кора головного мозга и мозжечокдва исключения: они содержат богатый цитозольный альфа-синуклеин, но очень низкие уровни митохондриального альфа-синуклеина. Показано, что альфа-синуклеин локализуется во внутренней мембране митохондрий и что ингибирующее действие альфа-синуклеина на активность комплекса I дыхательной цепи митохондрий носит дозозависимый характер. Таким образом, предполагается, что альфа-синуклеин в митохондриях по-разному экспрессируется в разных областях мозга, а фоновые уровни митохондриального альфа-синуклеина могут быть потенциальным фактором, влияющим на функцию митохондрий и предрасполагающим некоторые нейроны к дегенерации. [24]
По крайней мере три изоформы синуклеина производятся путем альтернативного сплайсинга . [25] Основная форма белка и наиболее изученная — это полноразмерный белок, состоящий из 140 аминокислот. Другими изоформами являются альфа-синуклеин-126, в котором отсутствуют остатки 41–54 из-за потери экзона 3; и альфа-синуклеин-112, [26] в котором отсутствуют остатки 103-130 из-за потери экзона 5. [25]
В кишечной нервной системе (ЭНС) [ править ]
Первые характеристики агрегатов aSyn в ЭНС пациентов с БП были выполнены на аутопсийных образцах в конце 1980-х годов. [27] Пока неизвестно, являются ли изменения микробиома, связанные с БП, следствием процесса заболевания или основной патофизиологии, или того и другого. [28]
У людей с диагнозом различных синуклеинопатий часто наблюдаются запоры и другие желудочно-кишечные дисфункции за несколько лет до появления двигательной дисфункции. [29]
Альфа-синуклеин потенциально соединяет ось кишечник-мозг у пациентов с болезнью Паркинсона . Распространенная наследственная болезнь Паркинсона связана с мутациями гена альфа-синуклеина (SNCA). В процессе зародышевой нуклеации альфа-синуклеин приобретает поперечно-листовую структуру, аналогичную другим амилоидам. [27]
Enterobacteriaceae , которые представляют , которые довольно часто встречаются в кишечнике человека, могут создавать курли собой функциональные амилоидные белки. Развернутый амилоид CsgA, который секретируется бактериями и позже агрегируется внеклеточно с образованием биопленок, опосредует адгезию к эпителиальным клеткам и способствует защите от бактериофагов, образует курчавые волокна. Пероральная инъекция бактерий, продуцирующих курли, также может стимулировать образование и агрегацию амилоидного белка Syn у старых крыс и нематод . Воспалительные реакции хозяина в кишечном тракте и на периферии модулируются воздействием курли. Биохимические исследования показывают, что эндогенные бактериальные шапероны кёрли способны кратковременно взаимодействовать с Сином и контролировать его агрегацию. [29]
Клинические и патологические данные подтверждают гипотезу о том, что болезнь aSyn при БП возникает через кишечник-мозг. Поэтому для ранней диагностики и раннего лечения на этапе создания и распространения aSyn крайне важно идентифицировать патогенный aSyn в пищеварительной системе, например, с помощью биопсии желудочно-кишечного тракта (ЖКТ). [27]
Согласно растущему числу исследований, дисбиоз кишечника может быть основным фактором развития болезни Паркинсона, способствуя кишечной проницаемости, воспалению желудочно-кишечного тракта, а также агрегации и распространению асин. [27]
Не только ЦНС, но и другие периферические ткани, такие как ЖКТ, имеют физиологическую экспрессию aSyn, а также его фосфорилированные варианты. [30] Как предположили Боргаммер и Ван Ден Берге (2019), один из подходов заключается в признании возможности возникновения подтипов БП с различными методами распространения aSyn, включая путь «сначала периферическая нервная система» (ПНС) или «сначала ЦНС». [31]
Хотя желудочно-кишечный тракт связан с другими неврологическими расстройствами, такими как расстройства аутистического спектра , депрессия , тревога и болезнь Альцгеймера , агрегация белков и/или воспаление в кишечнике представляют собой новую тему исследований синуклеинопатий . [29]
Структура [ править ]
Альфа-синуклеин в растворе считается внутренне неупорядоченным белком , т.е. у него отсутствует единая стабильная трехмерная структура. [32] [33] Однако по состоянию на 2014 год все больше сообщений предполагают наличие частичных структур или в основном структурированных олигомерных состояний в структуре раствора альфа-синуклеина даже в отсутствие липидов. Эта тенденция также подтверждается большим количеством измерений одиночных молекул ( оптическим пинцетом ) на отдельных копиях мономерного альфа-синуклеина, а также ковалентно связанных димеров или тетрамеров альфа-синуклеина. [34]
Альфа-синуклеин специфически активируется в дискретной популяции пресинаптических окончаний мозга в период синаптической перестройки, связанной с приобретением. [35] Показано, что альфа-синуклеин существенно взаимодействует с тубулином , [36] и что альфа-синуклеин может обладать активностью потенциального белка, ассоциированного с микротрубочками , такого как тау . [37] Данные свидетельствуют о том, что альфа-синуклеин действует как молекулярный шаперон при образовании комплексов SNARE . [38] [39] В частности, он одновременно связывается с фосфолипидами плазматической мембраны через свой N-концевой домен и с синаптобревином -2 через свой С-концевой домен, что имеет повышенное значение во время синаптической активности. [40] Действительно, появляется все больше доказательств того, что альфа-синуклеин участвует в функционировании нейронального аппарата Гольджи и транспорте пузырьков . [41]
По-видимому, альфа-синуклеин необходим для нормального развития когнитивных функций. У нокаутных мышей с целевой инактивацией экспрессии альфа-синуклеина наблюдаются нарушения пространственного обучения и рабочей памяти. [42]
Взаимодействие с липидными мембранами [ править ]
Были собраны экспериментальные данные о взаимодействии альфа-синуклеина с мембраной и его участии в составе и обмене мембран. Скрининг дрожжевого генома показал, что несколько генов, отвечающих за метаболизм липидов и слияние митохондрий, играют роль в токсичности альфа-синуклеина. [43] [44] И наоборот, уровни экспрессии альфа-синуклеина могут влиять на вязкость и относительное количество жирных кислот в липидном бислое. [45]
Известно, что альфа-синуклеин напрямую связывается с липидными мембранами, связываясь с отрицательно заряженными поверхностями фосфолипидов . [45] Альфа-синуклеин образует вытянутую спиральную структуру на небольших однослойных везикулах. [46] Было обнаружено преимущественное связывание с небольшими везикулами. [47] Связывание альфа-синуклеина с липидными мембранами оказывает на последние комплексное воздействие, изменяя структуру бислоя и приводя к образованию мелких везикул. [48] Было показано, что альфа-синуклеин изгибает мембраны отрицательно заряженных фосфолипидных везикул и образует канальцы из крупных липидных везикул. [49] С помощью криоЭМ было показано, что это мицеллярные трубочки диаметром ~5-6 нм. [50] Также было показано, что альфа-синуклеин образует липидные дискообразные частицы, подобные аполипопротеинам . [51] Исследования ЭПР показали, что структура альфа-синуклеина зависит от связывающей поверхности. [52] Белок принимает разорванную спиральную конформацию на липопротеиновых частицах, в то время как он образует вытянутую спиральную структуру на липидных везикулах и мембранных трубках. [52] Исследования также предположили возможную антиоксидантную активность альфа-синуклеина в мембране. [53]

Мембранное взаимодействие альфа-синуклеина модулирует или влияет на скорость его агрегации. [54] Мембранно-опосредованная модуляция агрегации очень похожа на ту, которая наблюдается для других амилоидных белков, таких как IAPP и абета. [54] Агрегированные состояния альфа-синуклеина пронизывают мембрану липидных везикул. [55] Они образуются при взаимодействии со склонными к перекисному окислению полиненасыщенными жирными кислотами (ПНЖК), но не с мононенасыщенными жирными кислотами. [56] а связывание автоокислению способствующих переходных металлов, липидов, таких как железо или медь, провоцирует олигомеризацию альфа-синуклеина. [57] Агрегированный альфа-синуклеин обладает специфической активностью в отношении перекисных липидов и индуцирует автоокисление липидов в богатых ПНЖК мембранах как нейронов, так и астроцитов, снижая устойчивость к апоптозу. [58] Аутоокисление липидов ингибируется, если клетки предварительно инкубировать с изотопно-армированными ПНЖК (D-ПНЖК). [59]
Функция [ править ]
Хотя функция альфа-синуклеина недостаточно изучена, исследования показывают, что он играет роль в ограничении подвижности синаптических везикул, что, как следствие, ослабляет рециркуляцию синаптических везикул и высвобождение нейромедиаторов. [60] [61] [62] [63] [64] [65] [9] Альтернативная точка зрения состоит в том, что альфа-синуклеин связывается с VAMP2 ( синаптобревином ) и стабилизирует комплексы SNARE ; [40] [66] [67] [68] [69] хотя недавние исследования показывают, что связывание альфа-синуклеина с VAMP2 имеет решающее значение для опосредованного альфа-синуклеином ослабления рециркуляции синаптических пузырьков, соединяя две, казалось бы, разные точки зрения. [9] Это также может помочь регулировать высвобождение дофамина — типа нейромедиатора, который имеет решающее значение для контроля начала и остановки произвольных и непроизвольных движений. [5]
Альфа-синуклеин модулирует процессы репарации ДНК , включая восстановление двухцепочечных разрывов (DSB). [70] Маркеры ответа на повреждение ДНК локализуются совместно с альфа-синуклеином, образуя дискретные фокусы в клетках человека и мозге мыши. Истощение альфа-синуклеина в клетках человека вызывает повышенное введение DSB ДНК после воздействия блеомицина и снижение способности восстанавливать эти DSB. Кроме того, мыши с нокаутом альфа-синуклеина демонстрируют более высокий уровень DSB, и эту проблему можно облегчить путем трансгенного повторного введения человеческого альфа-синуклеина. Альфа-синуклеин стимулирует путь восстановления DSB, называемый негомологичным соединением концов . [70] Функция восстановления ДНК альфа-синуклеина, по-видимому, нарушена в нейронах, несущих включения с тельцами Леви , и это может вызвать гибель клеток.
альфа- функция Пронейрогенная синуклеина
При некоторых нейродегенеративных заболеваниях альфа-синуклеин продуцирует нерастворимые тельца включения . Эти заболевания, известные как синуклеинопатии , связаны либо с более высоким уровнем нормального альфа-синуклеина, либо с его мутантными вариантами. [71] Однако нормальная физиологическая роль Snca еще полностью не объяснена. Фактически было продемонстрировано, что физиологический Snca оказывает нейропротекторное воздействие путем ингибирования апоптоза, индуцированного несколькими типами апоптотических стимулов, или путем регулирования экспрессии белков, участвующих в апоптотических путях.Недавно было продемонстрировано, что активация альфа-синуклеина в зубчатой извилине (нейрогенная ниша, где новые нейроны генерируются на протяжении всей жизни) активирует стволовые клетки в модели преждевременного нервного старения. Эта модель показывает снижение экспрессии альфа-синуклеина и снижение пролиферации стволовых клеток, что физиологически наблюдается при старении. Экзогенный альфа-синуклеин в зубчатой извилине способен устранить этот дефект. Более того, альфа-синуклеин также усиливает пролиферацию нервных клеток-предшественников зубчатой извилины у молодых мышей дикого типа. Таким образом, альфа-синуклеин представляет собой эффектор активации нервных стволовых клеток и клеток-предшественников. [72] Аналогичным образом было обнаружено, что альфа-синуклеин необходим для поддержания стволовых клеток СВЗ (субвентрикулярной зоны, т.е. другой нейрогенной ниши) в циклическом состоянии. [73]
Последовательность [ править ]
альфа-синуклеина Первичная структура обычно делится на три отдельных домена:
- Остатки 1–60: амфипатическая N-концевая область, в которой доминируют четыре повтора по 11 остатков, включая консенсусную последовательность KTKEGV. Эта последовательность имеет структурную склонность к альфа-спирали, аналогичную доменам, связывающим аполипопротеины. [74] Это высококонсервативный терминал, взаимодействующий с кислыми липидными мембранами, и все обнаруженные точечные мутации гена SNCA расположены внутри этого терминала. [75]
- Остатки 61-95: центральная гидрофобная область, которая включает область неамилоидного β-компонента (NAC), участвующую в агрегации белка. [13] Этот домен уникален для альфа-синуклеина среди семейства синуклеинов. [76]
- Остатки 96–140: высококислотная и богатая пролином область, не имеющая отчетливой структурной склонности. Этот домен играет важную роль в функции, растворимости и взаимодействии альфа-синуклеина с другими белками . [77] [40]
Аутопротеолитическая активность [ править ]
Использование масс-спектрометрии ионной подвижности высокого разрешения (IMS-MS) на альфа-синуклеине, очищенном с помощью ВЭЖХ, in vitro показало, что альфа-синуклеин является аутопротеолитическим (самопротеолитическим ) , генерируя при инкубации множество фрагментов с малой молекулярной массой . [78] Было обнаружено, что белок массой 14,46 кДа генерирует множество более мелких фрагментов, в том числе фрагменты массой 12,16 кДа ( аминокислоты 14–133) и 10,44 кДа (40–140), образующиеся в результате усечения С- и N-концев , а также C-концевой фрагмент массой 7,27 кДа (72 -140). Фрагмент массой 7,27 кДа, который содержит большую часть области NAC, агрегировал значительно быстрее, чем полноразмерный альфа-синуклеин. Возможно, что эти аутопротеолитические продукты играют роль промежуточных продуктов или кофакторов в агрегации альфа-синуклеина in vivo .
Клиническое значение
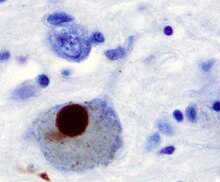
Альфа-синуклеин, не имеющий единой, четко определенной третичной структуры, представляет собой внутренне неупорядоченный белок . [79] [80] со значением pI 4,7, [81] который при определенных патологических условиях может неправильно сворачиваться таким образом, что его основные гидрофобные остатки подвергаются воздействию внутриклеточной среды, обеспечивая тем самым возможность возникновения гидрофобных взаимодействий с аналогичным, в равной степени экспонированным белком. [80] Это может привести к самосборке и последующей агрегации в крупные нерастворимые фибриллы, известные как амилоиды . [80] Превращение растворимого альфа-синуклеина в высокоупорядоченные фибриллярные структуры, пересекающие β-листы, не происходит, как считалось ранее, по двухэтапному механизму, а скорее происходит через ряд временных растворимых олигомерных промежуточных продуктов. [82] [83] В 2011 году две группы опубликовали свои выводы о том, что немутированный α-синуклеин образует стабильно свернутый тетрамер, который сопротивляется агрегации , утверждая, что этот свернутый тетрамер представляет собой соответствующую структуру in vivo в клетках. [84] [85] тем самым освобождая альфа-синуклеин от его нарушенного состояния. Сторонники гипотезы тетрамеров утверждали, что перекрестные связи in vivo в бактериях, первичных нейронах и клетках эритролейкемии человека подтверждают наличие лабильных тетрамерных видов. [86] [87] [88] Однако, несмотря на многочисленные отчеты внутриклеточного ЯМР, демонстрирующие, что альфа-синуклеин действительно является мономерным и неупорядоченным в интактных E. coli , клетках [89] [90] [91] [92] [93] [94] [95] [96] этот вопрос до сих пор остается предметом дискуссий на местах, несмотря на постоянно растущую гору противоречивых сообщений. [93] [97] [98] Тем не менее, альфа-синуклеин агрегирует с образованием нерастворимых фибрилл при патологических состояниях, характеризующихся тельцами Леви , таких как болезнь Паркинсона , деменция с тельцами Леви и множественная системная атрофия . [99] [100] Эти расстройства известны как синуклеинопатии . Модели синуклеинопатий in vitro показали, что агрегация альфа-синуклеина может приводить к различным клеточным нарушениям, включая повреждение микротрубочек, синаптические и митохондриальные дисфункции, окислительный стресс, а также нарушение регуляции передачи сигналов кальция, протеасомальных и лизосомальных путей. [101] Альфа-синуклеин является основным структурным компонентом фибрилл телец Леви. Иногда тельца Леви содержат тау-белок ; [102] однако альфа-синуклеин и тау составляют два различных подмножества филаментов в одних и тех же телецах включения. [103] Патология альфа-синуклеина также встречается как в спорадических, так и в семейных случаях болезни Альцгеймера. [104]
Механизм агрегации альфа-синуклеина неясен. Есть свидетельства существования структурированного промежуточного продукта, богатого бета-структурой , который может быть предшественником агрегации и, в конечном итоге, телец Леви. [105] Исследование одной молекулы, проведенное в 2008 году, предполагает, что альфа-синуклеин существует в виде смеси неструктурированных конформеров, богатых альфа-спиралями и бета-листами, в равновесии. Мутации или буферные условия, которые, как известно, улучшают агрегацию, сильно увеличивают популяцию бета-конформера, что позволяет предположить, что это может быть конформация, связанная с патогенной агрегацией. [106] Одна из теорий состоит в том, что большинство агрегатов альфа-синуклеина расположены в пресинапсе в виде более мелких отложений, что вызывает синаптическую дисфункцию. [107] Среди стратегий лечения синуклеинопатий есть соединения, ингибирующие агрегацию альфа-синуклеина. Было показано, что небольшая молекула куминальдегида ингибирует фибрилляцию альфа-синуклеина. [108] Вирус Эпштейна-Барра причастен к этим расстройствам. [109]
В редких случаях семейных форм болезни Паркинсона наблюдается мутация гена, кодирующего альфа-синуклеин. пять точечных мутаций На данный момент идентифицировано : A53T , [110] А30П, [111] Е46К, [112] H50Q, [113] и G51D; [114] однако всего с паркинсонизмом связано девятнадцать мутаций в гене SNCA: A18T, A29S, A53E, A53V, E57A, V15A, T72M, L8I, V15D, M127I, P117S, M5T, G93A, E83Q и A30G. [115]
Сообщалось, что некоторые мутации влияют на этапы инициации и амплификации процесса агрегации. [116] [117] Геномная дупликация и трипликация гена, по-видимому, является редкой причиной болезни Паркинсона в других линиях, хотя и более распространена, чем точечные мутации. [118] [119] Следовательно, определенные мутации альфа-синуклеина могут привести к образованию амилоидных фибрилл, которые способствуют развитию болезни Паркинсона. Сверхэкспрессия человеческого альфа-синуклеина дикого типа или A53T-мутанта у приматов приводит к отложению альфа-синуклеина в вентральной части среднего мозга, дегенерации дофаминергической системы и нарушению двигательных функций. [120]
Определенные участки белка альфа-синуклеина могут играть роль в таупатиях . [121] [122] [123]
Прионная форма белка альфа-синуклеина может быть возбудителем заболевания множественной системной атрофией . [124] [125] [126]

Описаны самореплицирующиеся «прионоподобные» амилоидные сборки альфа-синуклеина, невидимые для амилоидного красителя тиофлавина Т и способные быстро распространяться в нейронах in vitro и in vivo. [128]
Этот раздел нуждается в дополнительных цитатах для проверки . ( Ноябрь 2015 г. ) |
Антитела против альфа-синуклеина заменили антитела против убиквитина в качестве золотого стандарта иммуноокрашивания телец Леви. [129] Центральная панель на рисунке справа показывает основной путь агрегации белка. Мономерный α-синуклеин изначально разворачивается в растворе, но также может связываться с мембранами в α-спиральной форме. Кажется вероятным, что эти два вида существуют в равновесии внутри клетки, хотя это не доказано. Из работ in vitro становится ясно, что развернутый мономер может сначала агрегироваться в небольшие олигомерные частицы, которые можно стабилизировать за счет взаимодействий, подобных β-листам, а затем в нерастворимые фибриллы с более высокой молекулярной массой. В клеточном контексте есть некоторые доказательства того, что присутствие липидов может способствовать образованию олигомеров: α-синуклеин также может образовывать кольцевые пороподобные структуры, которые взаимодействуют с мембранами. Отложение α-синуклеина в патологических структурах, таких как тельца Леви, вероятно, является поздним событием, которое происходит в некоторых нейронах. Слева приведены некоторые из известных модификаторов этого процесса. Электрическая активность нейронов изменяет ассоциацию α-синуклеина с везикулами, а также может стимулировать поло-подобная киназа 2 (PLK2), которая, как было показано, фосфорилирует α-синуклеин по Ser 129. Предполагается, что в этом участвуют и другие киназы. Помимо фосфорилирования, усечения с помощью протеаз, таких как кальпаины , и нитрования, вероятно, за счет оксида азота (NO) или других активных форм азота, которые присутствуют во время воспаления, все они модифицируют синуклеин, так что он имеет более высокую склонность к агрегации. Добавление убиквитина (показано черным пятном) к тельцам Леви, вероятно, является вторичным процессом отложения. Справа показаны некоторые из предполагаемых клеточных мишеней для токсичности, опосредованной альфа-синуклеином, которые включают (сверху вниз) транспорт ЭР-Гольджи, синаптические везикулы, митохондрии и лизосомы и другие протеолитические механизмы. Предполагается, что в каждом из этих случаев α-синуклеин оказывает вредное воздействие, перечисленное под каждой стрелкой, хотя в настоящее время неясно, является ли какое-либо из них необходимым или достаточным для токсичности нейронов.
Белково-белковые взаимодействия [ править ]
Было показано, что альфа-синуклеин взаимодействует с
- Переносчик дофамина , [130] [131]
- Паркин (лигаза) , [132] [133]
- фосфолипаза D1 , [134]
- СНКАИП , [135] [136] [137] [138]
- Тау-белок . [139] [140] [141]
- Бета-амилоид [142]
См. также [ править ]
- Синуклеин
- Контурси Терме — деревня в Италии, где мутация гена альфа-синуклеина привела к семейному анамнезу болезни Паркинсона.
Ссылки [ править ]
- ↑ Перейти обратно: Перейти обратно: а б с GRCh38: Версия Ensembl 89: ENSG00000145335 – Ensembl , май 2017 г.
- ↑ Перейти обратно: Перейти обратно: а б с GRCm38: выпуск Ensembl 89: ENSMUSG00000025889 – Ensembl , май 2017 г.
- ^ «Ссылка на Human PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Ссылка на Mouse PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ↑ Перейти обратно: Перейти обратно: а б с д и «Домашний справочник генетики: SNCA» . Национальная медицинская библиотека США. 12 ноября 2013 г. Проверено 14 ноября 2013 г.
- ^ Бонини Н.М., Гиассон Б.И. (ноябрь 2005 г.). «Подавление функции альфа-синуклеина» . Клетка . 123 (3): 359–361. дои : 10.1016/j.cell.2005.10.017 . ПМИД 16269324 . S2CID 18772904 .
- ^ Чандра С., Галлардо Г., Фернандес-Чакон Р., Шлютер О.М., Зюдхоф ТЦ (ноябрь 2005 г.). «Альфа-синуклеин сотрудничает с CSPalpha в предотвращении нейродегенерации» . Клетка . 123 (3): 383–396. дои : 10.1016/j.cell.2005.09.028 . ПМИД 16269331 . S2CID 18173864 .
- ^ Чандра С., Чен X, Ризо Дж., Ян Р., Südhof TC (апрель 2003 г.). «Разорванная альфа-спираль в свернутом альфа-синуклеине» . Журнал биологической химии . 278 (17): 15313–15318. дои : 10.1074/jbc.M213128200 . ПМИД 12586824 . S2CID 27116894 .
- ↑ Перейти обратно: Перейти обратно: а б с Сунь Дж., Ван Л., Бао Х., Преми С., Дас Ю, Чепмен Э.Р., Рой С. (июнь 2019 г.). «Функциональное сотрудничество α-синуклеина и VAMP2 в рециркуляции синаптических пузырьков» . Труды Национальной академии наук Соединенных Штатов Америки . 116 (23): 11113–11115. Бибкод : 2019PNAS..11611113S . дои : 10.1073/pnas.1903049116 . ПМК 6561242 . ПМИД 31110017 .
- ^ Атиас М., Тевет Ю., Сан Дж., Ставски А., Таль С., Кан Дж. и др. (июнь 2019 г.). «Синапсины регулируют функции α-синуклеина» . Труды Национальной академии наук Соединенных Штатов Америки . 116 (23): 11116–11118. Бибкод : 2019PNAS..11611116A . дои : 10.1073/pnas.1903054116 . ПМК 6561288 . ПМИД 31110014 .
- ^ Спиллантини М.Г., Шмидт М.Л., Ли В.М., Трояновски Дж.К., Джейкс Р., Гедерт М. (28 августа 1997 г.). «α-Синуклеин в тельцах Леви» . Природа 388 (6645): 839–840. Бибкод : 1997Nature.388..839G . дои : 10.1038/42166 . ПМИД 9278044 . S2CID 4419837 .
- ^ Зигмонд М.Дж., Койл Дж.Т., Роуленд Дж.П. (2015). Нейробиология заболеваний головного мозга: биологические основы неврологических и психических расстройств . Лондон. ISBN 978-0-12-398280-3 . OCLC 896232309 .
{{cite book}}: CS1 maint: отсутствует местоположение издателя ( ссылка ) - ↑ Перейти обратно: Перейти обратно: а б с Уэда К., Фукусима Х., Маслия Э., Ся Ю., Иваи А., Ёшимото М. и др. (декабрь 1993 г.). «Молекулярное клонирование кДНК, кодирующей нераспознанный компонент амилоида при болезни Альцгеймера» . Труды Национальной академии наук Соединенных Штатов Америки . 90 (23): 11282–11286. Бибкод : 1993PNAS...9011282U . дои : 10.1073/pnas.90.23.11282 . ПМК 47966 . ПМИД 8248242 .
- ^ Ся Ю., Сайто Т., Уэда К., Танака С., Чен Х., Хашимото М. и др. (октябрь 2001 г.). «Характеристика гена альфа-синуклеина человека: структура генома, сайт начала транскрипции, промоторная область и полиморфизмы» . Журнал болезни Альцгеймера . 3 (5): 485–494. дои : 10.3233/JAD-2001-3508 . ПМИД 12214035 . Архивировано из оригинала 14 мая 2016 г. Проверено 19 февраля 2009 г.
- ^ Ся Ю, Сайто Т, Уэда К, Танака С, Чен X, Хашимото М, Сюй Л, Конрад С, Сундсмо М, Ёшимото М, Тал Л, Кацман Р, Маслия Э (2002). «Характеристика гена альфа-синуклеина человека: геномная структура, сайт начала транскрипции, промоторная область и полиморфизмы: ошибка p489, рис. 3» . Дж. Альцгеймерс Дис . 4 (4): 337. Архивировано из оригинала 14 мая 2016 г. Проверено 19 февраля 2009 г.
- ^ Джейкс Р., Спиллантини М.Г., Гедерт М. (май 1994 г.). «Идентификация двух различных синуклеинов человеческого мозга» . Письма ФЭБС . 345 (1): 27–32. дои : 10.1016/0014-5793(94)00395-5 . ПМИД 8194594 . S2CID 36840279 .
- ↑ Перейти обратно: Перейти обратно: а б Иваи А., Маслия Э., Ёсимото М., Ге Н., Фланаган Л., де Сильва Х.А. и др. (февраль 1995 г.). «Белок-предшественник не-А-бета-компонента амилоида болезни Альцгеймера является пресинаптическим белком центральной нервной системы» . Нейрон . 14 (2): 467–475. дои : 10.1016/0896-6273(95)90302-X . ПМИД 7857654 . S2CID 17941420 .
- ^ Филиппини А, Дженнарелли М, Руссо I (март 2019 г.). «А-синуклеин и глия при болезни Паркинсона: полезный или вредный дуэт для эндолизосомальной системы?». Клеточная и молекулярная нейробиология . 39 (2): 161–168. дои : 10.1007/s10571-019-00649-9 . ПМИД 30637614 . S2CID 58006790 .
- ^ Хук К.С., Шлегель Н.К., Эйххофф О.М., Видмер Д.С., Преториус С., Эйнарссон С.О. и др. (декабрь 2008 г.). «Новые цели MITF идентифицированы с использованием двухэтапной стратегии микрочипов ДНК» . Исследование пигментных клеток и меланомы . 21 (6): 665–676. дои : 10.1111/j.1755-148X.2008.00505.x . ПМИД 19067971 . S2CID 24698373 .
- ^ Ю С., Ли Х, Лю Г, Хань Дж, Чжан С, Ли Ю и др. (март 2007 г.). «Обширная ядерная локализация альфа-синуклеина в нейронах головного мозга нормальных крыс, выявленная с помощью нового моноклонального антитела». Нейронаука . 145 (2): 539–555. doi : 10.1016/j.neuroscience.2006.12.028 . ПМИД 17275196 . S2CID 37294944 .
- ^ Маклин П.Дж., Кавамата Х., Рибич С., Хайман Б.Т. (март 2000 г.). «Мембранная ассоциация и конформация белка альфа-синуклеина в интактных нейронах. Влияние мутаций, связанных с болезнью Паркинсона» . Журнал биологической химии . 275 (12): 8812–8816. дои : 10.1074/jbc.275.12.8812 . ПМИД 10722726 .
- ^ Ли Х.Дж., Чой С., Ли С.Дж. (январь 2002 г.). «Мембраносвязанный альфа-синуклеин обладает высокой склонностью к агрегации и способностью вызывать агрегацию цитозольной формы» . Журнал биологической химии . 277 (1): 671–678. дои : 10.1074/jbc.M107045200 . ПМИД 11679584 . S2CID 10438997 .
- ^ Чжан Л., Чжан С., Чжу Ю, Цай Ц, Чан П., Уэда К. и др. (декабрь 2008 г.). «Полуколичественный анализ альфа-синуклеина в субклеточных пулах нейронов головного мозга крыс: электронно-микроскопическое исследование иммунозолота с использованием С-концевого специфического моноклонального антитела». Исследования мозга . 1244 : 40–52. дои : 10.1016/j.brainres.2008.08.067 . ПМИД 18817762 . S2CID 1737088 .
- ↑ Перейти обратно: Перейти обратно: а б Лю Г, Чжан С, Инь Дж, Ли Х, Ченг Ф, Ли Ю и др. (май 2009 г.). «Альфа-синуклеин по-разному экспрессируется в митохондриях разных областей мозга крыс и в зависимости от дозы снижает активность комплекса I». Письма по неврологии . 454 (3): 187–192. дои : 10.1016/j.neulet.2009.02.056 . ПМИД 19429081 . S2CID 45120745 .
- ↑ Перейти обратно: Перейти обратно: а б Бейер К. (сентябрь 2006 г.). «Структура альфа-синуклеина, посттрансляционная модификация и альтернативный сплайсинг как усилители агрегации». Акта Нейропатологика . 112 (3): 237–251. дои : 10.1007/s00401-006-0104-6 . ПМИД 16845533 . S2CID 1367846 .
- ^ Уэда К., Сайто Т., Мори Х. (декабрь 1994 г.). «Тканезависимый альтернативный сплайсинг мРНК для NACP, предшественника не-А-бета-компонента амилоида болезни Альцгеймера». Связь с биохимическими и биофизическими исследованиями . 205 (2): 1366–1372. дои : 10.1006/bbrc.1994.2816 . ПМИД 7802671 .
- ↑ Перейти обратно: Перейти обратно: а б с д Шеффер Э., Клюге А., Беттнер М., Цунке Ф., Коссайс Ф., Берг Д., Арнольд П. (2020). «Альфа-синуклеин соединяет ось кишечник-мозг у пациентов с болезнью Паркинсона - взгляд на клинические аспекты, клеточную патологию и аналитическую методологию» . Границы клеточной биологии и биологии развития . 8 : 573696. doi : 10.3389/fcell.2020.573696 . ПМК 7509446 . ПМИД 33015066 .
- ^ Анис Э., Се А., Брундин Л., Брундин П. (февраль 2022 г.). «Обработка последних результатов: кишечный альфа-синуклеин, изменения микробиома при болезни Паркинсона». Тенденции в эндокринологии и обмене веществ . 33 (2): 147–157. дои : 10.1016/j.tem.2021.11.005 . ПМИД 34949514 . S2CID 245351514 .
- ↑ Перейти обратно: Перейти обратно: а б с Сэмпсон Т.Р., Чаллис С., Джайн Н., Моисеенко А., Ладинский М.С., Шастри Г.Г. и др. (февраль 2020 г.). Чиу И.М., Гарретт В.С., Дежарден М. (ред.). «Кишечный бактериальный амилоид способствует агрегации альфа-синуклеина и двигательным нарушениям у мышей» . электронная жизнь . 9 : е53111. дои : 10.7554/eLife.53111 . ПМК 7012599 . ПМИД 32043464 .
- ^ Барренши М., Зоренков Д., Беттнер М., Ланге С., Коссаис Ф., Шарф А.Б. и др. (январь 2017 г.). «Отличительная структура кишечных агрегатов фосфо-альфа-синуклеина и профили экспрессии генов у пациентов с болезнью Паркинсона» . Acta Neuropathologica Communications . 5 (1): 1. дои : 10.1186/s40478-016-0408-2 . ПМК 5217296 . ПМИД 28057070 .
- ^ Борхаммер П., Ван Ден Берге Н. (30 октября 2019 г.). «Болезнь Паркинсона, связанная с поражением мозга и кишечника: гипотеза» . Журнал болезни Паркинсона . 9 (с2): С281–С295. дои : 10.3233/jpd-191721 . ПМК 6839496 . ПМИД 31498132 .
- ^ ван Ройен Б.Д., ван Лейенхорст-Гренер К.А., Классенс М.М., Субраманиам В. (декабрь 2009 г.). «Флуоресценция триптофана выявляет структурные особенности олигомеров альфа-синуклеина» (PDF) . Журнал молекулярной биологии . 394 (5): 826–833. дои : 10.1016/j.jmb.2009.10.021 . ПМИД 19837084 . S2CID 36085937 .
- ^ Вейнреб П.Х., Чжэнь В., Пун А.В., Конвей К.А., Лэнсбери П.Т. (октябрь 1996 г.). «NACP, белок, участвующий в болезни Альцгеймера и обучении, развернут в природе». Биохимия . 35 (43): 13709–13715. дои : 10.1021/bi961799n . ПМИД 8901511 .
- ^ Неупане К., Соланки А., Сосова И., Белов М., Вудсайд М.Т. (январь 2014 г.). «Разнообразные метастабильные структуры, образованные небольшими олигомерами α-синуклеина, исследованные с помощью силовой спектроскопии» . ПЛОС ОДИН . 9 (1): e86495. Бибкод : 2014PLoSO...986495N . дои : 10.1371/journal.pone.0086495 . ПМК 3901707 . ПМИД 24475132 .
- ^ Джордж Дж.М., Джин Х., Вудс В.С., Клейтон Д.Ф. (август 1995 г.). «Характеристика нового белка, регулируемого в критический период обучения пению у зебры» . Нейрон . 15 (2): 361–372. дои : 10.1016/0896-6273(95)90040-3 . PMID 7646890 . S2CID 11421888 .
- ^ Алим М.А., Хоссейн М.С., Арима К., Такеда К., Изумияма Ю., Накамура М. и др. (январь 2002 г.). «Формирование фибрилл альфа-синуклеина семян тубулина» . Журнал биологической химии . 277 (3): 2112–2117. дои : 10.1074/jbc.M102981200 . ПМИД 11698390 . S2CID 84374030 .
- ^ Алим М.А., Ма К.Л., Такеда К., Айзава Т., Мацубара М., Накамура М. и др. (август 2004 г.). «Демонстрация роли альфа-синуклеина как функционального белка, связанного с микротрубочками». Журнал болезни Альцгеймера . 6 (4): 435–42, обсуждение 443–9. дои : 10.3233/JAD-2004-6412 . ПМИД 15345814 .
- ^ Бонини Н.М., Гиассон Б.И. (ноябрь 2005 г.). «Подавление функции альфа-синуклеина» . Клетка . 123 (3): 359–361. дои : 10.1016/j.cell.2005.10.017 . ПМИД 16269324 . S2CID 18772904 .
- ^ Чандра С., Галлардо Г., Фернандес-Чакон Р., Шлютер О.М., Зюдхоф ТЦ (ноябрь 2005 г.). «Альфа-синуклеин сотрудничает с CSPalpha в предотвращении нейродегенерации» . Клетка . 123 (3): 383–396. дои : 10.1016/j.cell.2005.09.028 . ПМИД 16269331 . S2CID 18173864 .
- ↑ Перейти обратно: Перейти обратно: а б с Бурре Дж., Шарма М., Цеценис Т., Бухман В., Этертон М.Р., Зюдхоф ТЦ (сентябрь 2010 г.). «Альфа-синуклеин способствует сборке SNARE-комплекса in vivo и in vitro» . Наука . 329 (5999): 1663–1667. Бибкод : 2010Sci...329.1663B . дои : 10.1126/science.1195227 . ПМЦ 3235365 . ПМИД 20798282 .
- ^ Купер А.А., Гитлер А.Д., Кашикар А., Хейнс С.М., Хилл К.Дж., Бхуллар Б. и др. (июль 2006 г.). «Альфа-синуклеин блокирует трафик ER-Гольджи, а Rab1 спасает потерю нейронов в моделях Паркинсона» . Наука . 313 (5785): 324–328. Бибкод : 2006Sci...313..324C . дои : 10.1126/science.1129462 . ЧВК 1983366 . ПМИД 16794039 .
- ^ Кохан В.С., Афанасьева М.А., Ванькин Г.И. (май 2012 г.). «Мыши с нокаутом α-синуклеина имеют когнитивные нарушения». Поведенческие исследования мозга . 231 (1): 226–230. дои : 10.1016/j.bbr.2012.03.026 . ПМИД 22469626 . S2CID 205884600 .
- ^ Тауро М (4 февраля 2019 г.). «Токсичность альфа-синуклеина вызвана митохондриальной дисфункцией» . Электронный репозиторий диссертаций и диссертаций .
- ^ Уиллингем С., Отейро Т.Ф., ДеВит М.Дж., Линдквист С.Л., Муховски П.Дж. (декабрь 2003 г.). «Дрожжевые гены, усиливающие токсичность мутантного фрагмента хантингтина или альфа-синуклеина». Наука . 302 (5651): 1769–1772. Бибкод : 2003Sci...302.1769W . дои : 10.1126/science.1090389 . ПМИД 14657499 . S2CID 43221047 .
- ↑ Перейти обратно: Перейти обратно: а б Уверский В.Н. (октябрь 2007 г.). «Нейропатология, биохимия и биофизика агрегации альфа-синуклеина». Журнал нейрохимии . 103 (1): 17–37. дои : 10.1111/j.1471-4159.2007.04764.x . ПМИД 17623039 . S2CID 85334400 .
- ^ Джао CC, Хегде Б.Г., Чен Дж., Хаворт И.С., Ланген Р. (декабрь 2008 г.). «Структура мембраносвязанного альфа-синуклеина на основе сайт-направленного спинового мечения и вычислительной обработки» . Труды Национальной академии наук Соединенных Штатов Америки . 105 (50): 19666–19671. Бибкод : 2008PNAS..10519666J . дои : 10.1073/pnas.0807826105 . ПМК 2605001 . ПМИД 19066219 .
- ^ Чжу М., Ли Дж., Финк А.Л. (октябрь 2003 г.). «Ассоциация альфа-синуклеина с мембранами влияет на структуру бислоя, стабильность и образование фибрилл» . Журнал биологической химии . 278 (41): 40186–40197. дои : 10.1074/jbc.M305326200 . ПМИД 12885775 . S2CID 41555488 .
- ^ Мадин Дж., Дойг А.Дж., Миддлтон Д.А. (май 2006 г.). «Исследование регионального влияния альфа-синуклеина на организацию и стабильность фосфолипидных бислоев». Биохимия . 45 (18): 5783–5792. дои : 10.1021/bi052151q . ПМИД 16669622 .
- ^ Варки Дж., Исас Дж.М., Мизуно Н., Дженсен М.Б., Бхатия В.К., Джао CC и др. (октябрь 2010 г.). «Индукция кривизны мембраны и тубуляция являются общими чертами синуклеинов и аполипопротеинов» . Журнал биологической химии . 285 (42): 32486–32493. дои : 10.1074/jbc.M110.139576 . ПМЦ 2952250 . ПМИД 20693280 .
- ^ Мизуно Н., Варки Дж., Кегулян Н.К., Хегде Б.Г., Ченг Н., Ланген Р., Стивен А.С. (август 2012 г.). «Ремоделирование липидных везикул в цилиндрические мицеллы α-синуклеином в вытянутой α-спиральной конформации» . Журнал биологической химии . 287 (35): 29301–29311. дои : 10.1074/jbc.M112.365817 . ПМЦ 3436199 . ПМИД 22767608 .
- ^ Варки Дж., Мизуно Н., Хегде Б.Г., Ченг Н., Стивен А.С., Ланген Р. (июнь 2013 г.). «Олигомеры α-синуклеина с нарушенной спиральной конформацией образуют наночастицы липопротеинов» . Журнал биологической химии . 288 (24): 17620–17630. дои : 10.1074/jbc.M113.476697 . ПМЦ 3682563 . ПМИД 23609437 .
- ↑ Перейти обратно: Перейти обратно: а б Варки Дж., Ланген Р. (июль 2017 г.). «Ремоделирование мембраны амилоидогенными и неамилоидогенными белками, изученными с помощью ЭПР» . Журнал магнитного резонанса . 280 : 127–139. Бибкод : 2017JMagR.280..127V . дои : 10.1016/j.jmr.2017.02.014 . ПМК 5461824 . ПМИД 28579098 .
- ^ Чжу М, Цинь З.Дж., Ху Д., Мунишкина Л.А., Финк А.Л. (июль 2006 г.). «Альфа-синуклеин может действовать как антиоксидант, предотвращая окисление ненасыщенных липидов в везикулах». Биохимия . 45 (26): 8135–8142. дои : 10.1021/bi052584t . ПМИД 16800638 .
- ↑ Перейти обратно: Перейти обратно: а б Рават А., Ланген Р., Варки Дж. (сентябрь 2018 г.). «Мембраны как модуляторы неправильного сворачивания амилоидного белка и мишень токсичности» . Biochimica et Biophysical Acta (BBA) – Биомембраны . 1860 (9): 1863–1875. дои : 10.1016/j.bbamem.2018.04.011 . ПМК 6203680 . ПМИД 29702073 .
- ^ Флагмейер П., Де С., Виртенсон Д.С., Ли С.Ф., Винке Дж., Мюлдерманс С. и др. (июнь 2017 г.). «Сверхчувствительное измерение Ca 2+ Приток в липидные везикулы, индуцированный белковыми агрегатами» . Angewandte Chemie . 56 (27): 7750–7754. doi : 10.1002/ . PMC 5615231. . PMID 28474754 anie.201700966
- ^ Шэрон Р., Бар-Джозеф I, член парламента Фрош, Уолш Д.М., Гамильтон Дж.А., Селькое DJ (февраль 2003 г.). «Образование хорошо растворимых олигомеров альфа-синуклеина регулируется жирными кислотами и усиливается при болезни Паркинсона» . Нейрон . 37 (4): 583–595. дои : 10.1016/s0896-6273(03)00024-2 . ПМИД 12597857 . S2CID 1604719 .
- ^ Амер Д.А., Ирвин ГБ, Эль-Агнаф О.М. (август 2006 г.). «Ингибиторы олигомеризации и токсичности альфа-синуклеина: будущая терапевтическая стратегия лечения болезни Паркинсона и связанных с ней расстройств». Экспериментальное исследование мозга . 173 (2): 223–233. дои : 10.1007/s00221-006-0539-y . ПМИД 16733698 . S2CID 24760126 .
- ^ Руиперес В., Дариос Ф., Давлетов Б. (октябрь 2010 г.). «Альфа-синуклеин, липиды и болезнь Паркинсона». Прогресс в исследованиях липидов . 49 (4): 420–428. дои : 10.1016/j.plipres.2010.05.004 . ПМИД 20580911 .
- ^ Ангелова П.Р., Хоррокс М.Х., Кленерман Д., Ганди С., Абрамов А.Ю., Щепинов М.С. (май 2015 г.). «Перекисное окисление липидов необходимо для гибели клеток, индуцированной альфа-синуклеином» . Журнал нейрохимии . 133 (4): 582–589. дои : 10.1111/jnc.13024 . ПМЦ 4471127 . ПМИД 25580849 .
- ^ Ларсен К.Е., Шмитц Ю., Тройер М.Д., Мошаров Э., Дитрих П., Квази А.З. и др. (ноябрь 2006 г.). «Сверхэкспрессия альфа-синуклеина в PC12 и хромаффинных клетках ухудшает высвобождение катехоламинов, препятствуя позднему этапу экзоцитоза» . Журнал неврологии . 26 (46): 11915–11922. doi : 10.1523/JNEUROSCI.3821-06.2006 . ПМК 6674868 . ПМИД 17108165 .
- ^ Немани В.М., Лу В., Берге В., Накамура К., Оноа Б., Ли М.К. и др. (январь 2010 г.). «Повышенная экспрессия альфа-синуклеина снижает высвобождение нейромедиаторов за счет ингибирования повторной кластеризации синаптических пузырьков после эндоцитоза» . Нейрон . 65 (1): 66–79. дои : 10.1016/j.neuron.2009.12.023 . ПМЦ 3119527 . ПМИД 20152114 .
- ^ Скотт Д.А., Табарин И., Тан И., Картье А., Маслия Э., Рой С. (июнь 2010 г.). «Патологический каскад, приводящий к синаптической дисфункции при нейродегенерации, индуцированной альфа-синуклеином» . Журнал неврологии . 30 (24): 8083–8095. doi : 10.1523/JNEUROSCI.1091-10.2010 . ПМК 2901533 . ПМИД 20554859 .
- ^ Скотт Д., Рой С. (июль 2012 г.). «α-Синуклеин ингибирует межсинаптическую подвижность везикул и поддерживает гомеостаз рециркуляционного пула» . Журнал неврологии . 32 (30): 10129–10135. doi : 10.1523/JNEUROSCI.0535-12.2012 . ПМЦ 3426499 . ПМИД 22836248 .
- ^ Варгас К.Дж., Макани С., Дэвис Т., Вестфаль CH, Кастильо П.Е., Чандра СС (июль 2014 г.). «Синуклеины регулируют кинетику эндоцитоза синаптических пузырьков» . Журнал неврологии . 34 (28): 9364–9376. doi : 10.1523/JNEUROSCI.4787-13.2014 . ПМК 4087213 . ПМИД 25009269 .
- ^ Ван Л., Дас Ю, Скотт Д.А., Тан Ю, Маклин П.Дж., Рой С. (октябрь 2014 г.). «Мультимеры альфа-синуклеина группируют синаптические пузырьки и ослабляют рециклинг» . Современная биология . 24 (19): 2319–2326. Бибкод : 2014CBio...24.2319W . дои : 10.1016/j.cub.2014.08.027 . ПМК 4190006 . ПМИД 25264250 .
- ^ Бурре Дж., Шарма М., Südhof TC (март 2018 г.). «Клеточная биология и патофизиология α-синуклеина» . Перспективы Колд-Спринг-Харбора в медицине . 8 (3): а024091. doi : 10.1101/cshperspect.a024091 . ПМК 5519445 . ПМИД 28108534 .
- ^ Бурре Дж., Шарма М., Südhof TC (октябрь 2012 г.). «Систематический мутагенез α-синуклеина выявляет различные требования к последовательности для физиологической и патологической активности» . Журнал неврологии . 32 (43): 15227–15242. doi : 10.1523/JNEUROSCI.3545-12.2012 . ПМК 3506191 . ПМИД 23100443 .
- ^ Бурре Дж., Шарма М., Südhof TC (октябрь 2014 г.). «α-синуклеин собирается в мультимеры более высокого порядка при связывании с мембраной, способствуя образованию комплекса SNARE» . Труды Национальной академии наук Соединенных Штатов Америки . 111 (40): Е4274–Е4283. Бибкод : 2014PNAS..111E4274B . дои : 10.1073/pnas.1416598111 . ПМК 4210039 . ПМИД 25246573 .
- ^ Диао Дж., Бурре Дж., Вивона С., Сиприано Дж., Шарма М., Кёнг М. и др. (апрель 2013 г.). «Нативный α-синуклеин индуцирует кластеризацию имитаторов синаптических пузырьков посредством связывания с фосфолипидами и синаптобревином-2/VAMP2» . электронная жизнь . 2 : e00592. doi : 10.7554/eLife.00592 . ПМЦ 3639508 . ПМИД 23638301 .
- ↑ Перейти обратно: Перейти обратно: а б Шазер А.Дж., Остерберг В.Р., Дент С.Е., Стэкхаус Т.Л., Уэйкхэм К.М., Бутрос С.В. и др. (июль 2019 г.). «Альфа-синуклеин — это ДНК-связывающий белок, который модулирует восстановление ДНК, что приводит к нарушениям с тельцами Леви» . Научные отчеты . 9 (1): 10919. Бибкод : 2019НатСР...910919С . дои : 10.1038/s41598-019-47227-z . ПМК 6662836 . ПМИД 31358782 .
- ^ Рыскалин Л., Бускети К.Л., Лиманаки Ф., Бьяджиони Ф., Гамбарделла С., Форнаи Ф. (2018). «Акцент на благотворном влиянии альфа-синуклеина и переоценка синуклеинопатий» . Современная наука о белках и пептидах . 19 (6): 598–611. дои : 10.2174/1389203718666171117110028 . ПМЦ 5925871 . ПМИД 29150919 .
- ^ Микели Л., Креанца Т.М., Чеккарелли М., Д'Андреа Г., Джаковаццо Г., Анкона Н. и др. (2021). «Анализ транскриптома на мышиной модели преждевременного старения зубчатой извилины: устранение дефицита альфа-синуклеина с помощью экспрессии, управляемой вирусом, или путем восстановления дефектного нейрогенеза» . Границы клеточной биологии и биологии развития . 9 : 696684. doi : 10.3389/fcell.2021.696684 . ПМЦ 8415876 . ПМИД 34485283 .
- ^ Перес-Вильяльба А., Сирерол-Пикер М.С., Беленгер Г., Сориано-Кантон Р., Муньос-Манчадо А.Б., Вилладьего Дж. и др. (январь 2018 г.). «Синаптический регулятор α-синуклеин в дофаминергических волокнах необходим для поддержания субэпендимальных нервных стволовых клеток» . Журнал неврологии . 38 (4): 814–825. doi : 10.1523/JNEUROSCI.2276-17.2017 . ПМК 6596232 . ПМИД 29217686 .
- ^ Клейтон Д.Ф., Джордж Дж.М. (июнь 1998 г.). «Синуклеины: семейство белков, участвующих в синаптической функции, пластичности, нейродегенерации и заболеваниях». Тенденции в нейронауках . 21 (6): 249–254. дои : 10.1016/S0166-2236(97)01213-7 . ПМИД 9641537 . S2CID 20654921 .
- ^ Бассел Р., Элиезер Д. (июнь 2003 г.). «Структурная и функциональная роль 11-мерных повторов в альфа-синуклеине и других обменных липидсвязывающих белках». Журнал молекулярной биологии . 329 (4): 763–778. дои : 10.1016/S0022-2836(03)00520-5 . ПМИД 12787676 .
- ^ Учихара Т., Гиассон Б.И. (январь 2016 г.). «Распространение патологии альфа-синуклеина: гипотезы, открытия и пока нерешенные вопросы экспериментальных исследований и исследований человеческого мозга» . Акта Нейропатологика . 131 (1): 49–73. дои : 10.1007/s00401-015-1485-1 . ПМЦ 4698305 . ПМИД 26446103 .
- ^ Соррентино З.А., Ся Ю., Горион К.М., Хасс Э., Гиассон Б.И. (апрель 2020 г.). «Усечение карбокси-конца мышиного альфа-синуклеина изменяет агрегацию и прионоподобное засев» . Письма ФЭБС . 594 (8): 1271–1283. дои : 10.1002/1873-3468.13728 . ПМЦ 7188589 . ПМИД 31912891 .
- ^ Влад С., Линднер К., Карреман С., Шильдкнехт С., Лейст М., Томчик Н. и др. (декабрь 2011 г.). «Аутопротеолитические фрагменты являются промежуточными продуктами в олигомеризации/агрегации белка альфа-синуклеина, вызывающего болезнь Паркинсона, как показано с помощью масс-спектрометрии ионной подвижности» . ХимБиоХим . 12 (18): 2740–2744. дои : 10.1002/cbic.201100569 . ПМК 3461308 . ПМИД 22162214 .
- ^ Чити Ф, Добсон СМ (июнь 2017 г.). «Неправильное сворачивание белка, образование амилоида и болезни человека: краткий обзор прогресса за последнее десятилетие». Ежегодный обзор биохимии . 86 : 27–68. doi : 10.1146/annurev-biochem-061516-045115 . hdl : 2158/1117236 . ПМИД 28498720 .
- ↑ Перейти обратно: Перейти обратно: а б с Ке ПК, Чжоу Р., Серпелл Л.С., Риек Р., Ноулз Т.П., Лашуэль Х.А. и др. (август 2020 г.). «Полвека амилоидов: прошлое, настоящее и будущее» . Обзоры химического общества . 49 (15): 5473–5509. дои : 10.1039/c9cs00199a . ПМЦ 7445747 . ПМИД 32632432 .
- ^ Фурукава К., Агирре С., Со М., Сасахара К., Мияноири И., Сакураи К. и др. (2020). «Образование изоэлектрической точки амилоида α-синуклеина расширяет общность механизма, ограниченного растворимостью и перенасыщением» . Текущие исследования в области структурной биологии . 2 : 35–44. doi : 10.1016/j.crstbi.2020.03.001 . ПМЦ 8244297 . ПМИД 34235468 .
- ^ Тейлет Ф.С., Бинольфи А., Фрембген-Кеснер Т., Хингорани К., Саркар М., Кин С. и др. (июль 2014 г.). «Физико-химические свойства клеток и их влияние на внутренне неупорядоченные белки (IDP)» . Химические обзоры . 114 (13): 6661–6714. дои : 10.1021/cr400695p . ПМК 4095937 . ПМИД 24901537 .
- ^ Айзенберг Д., Джакер М. (март 2012 г.). «Амилоидное состояние белков при заболеваниях человека» . Клетка . 148 (6): 1188–1203. дои : 10.1016/j.cell.2012.02.022 . ПМЦ 3353745 . ПМИД 22424229 .
- ^ Бартельс Т., Чой Ю.Г., Селькое DJ (август 2011 г.). «α-синуклеин физиологически представляет собой спирально свернутый тетрамер, который сопротивляется агрегации» . Природа . 477 (7362): 107–110. Бибкод : 2011Natur.477..107B . дои : 10.1038/nature10324 . ПМК 3166366 . ПМИД 21841800 .
- ^ Ван В., Перович И., Читтулуру Дж., Каганович А., Нгуен Л.Т., Ляо Дж. и др. (октябрь 2011 г.). «Растворимая конструкция α-синуклеина образует динамический тетрамер» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (43): 17797–17802. Бибкод : 2011PNAS..10817797W . дои : 10.1073/pnas.1113260108 . ПМК 3203798 . ПМИД 22006323 .
- ^ Деттмер У., Ньюман А.Дж., Лют Э.С., Бартельс Т., Селкое Д. (март 2013 г.). «Сшивание in vivo выявляет преимущественно олигомерные формы α-синуклеина и β-синуклеина в нейронах и ненейральных клетках» . Журнал биологической химии . 288 (9): 6371–6385. дои : 10.1074/jbc.M112.403311 . ПМК 3585072 . ПМИД 23319586 .
- ^ Вестфаль CH, Чандра СС (январь 2013 г.). «Мономерные синуклеины вызывают искривление мембраны» . Журнал биологической химии . 288 (3): 1829–1840. дои : 10.1074/jbc.M112.418871 . ПМЦ 3548493 . ПМИД 23184946 .
- ^ Трекслер А.Дж., Роудс Э. (май 2012 г.). «N-концевое ацетилирование имеет решающее значение для образования α-спирального олигомера α-синуклеина» . Белковая наука . 21 (5): 601–605. дои : 10.1002/pro.2056 . ПМК 3403458 . ПМИД 22407793 .
- ^ Бинольфи А., Тейе FX, Селенко П. (октябрь 2012 г.). «Бактериальный внутриклеточный ЯМР альфа-синуклеина человека: неупорядоченный мономер по природе?». Труды Биохимического общества . 40 (5): 950–954. дои : 10.1042/BST20120096 . ПМИД 22988846 .
- ^ Бертини И., Фелли И.С., Гоннелли Л., Васанта Кумар М.В., Пьераттелли Р. (октябрь 2011 г.). «Характеристика внутреннего беспорядка в белках с высоким разрешением: расширение набора экспериментов по ЯМР-спектроскопии, обнаруживающих (13) C, для определения ключевых наблюдаемых». ХимБиоХим . 12 (15): 2347–2352. дои : 10.1002/cbic.201100406 . ПМИД 23106082 . S2CID 34960247 .
- ^ Уодби Калифорния, Камиллони С., Фицпатрик А.В., Кабрита Л.Д., Добсон С.М., Вендрусколо М., Христодулу Дж. (26 августа 2013 г.). «Внутриклеточная ЯМР-характеристика популяций вторичной структуры неупорядоченной конформации α-синуклеина в клетках E. coli» . ПЛОС ОДИН . 8 (8): е72286. Бибкод : 2013PLoSO...872286W . дои : 10.1371/journal.pone.0072286 . ПМЦ 3753296 . ПМИД 23991082 .
- ^ Ли С, Лю М (апрель 2013 г.). «Динамика белков в живых клетках, изученная методом внутриклеточной ЯМР-спектроскопии» . Письма ФЭБС . 587 (8): 1008–1011. дои : 10.1016/j.febslet.2012.12.023 . ПМИД 23318712 . S2CID 2649589 .
- ↑ Перейти обратно: Перейти обратно: а б Фове Б., Мбефо М.К., Фарес М.Б., Десобри С., Майкл С., Арда М.Т. и др. (май 2012 г.). «α-Синуклеин в центральной нервной системе, эритроцитах, клетках млекопитающих и Escherichia coli существует преимущественно в виде неупорядоченного мономера» . Журнал биологической химии . 287 (19): 15345–15364. дои : 10.1074/jbc.M111.318949 . ПМЦ 3346117 . ПМИД 22315227 .
- ^ Перес А.С., Субрини О., Хессель А., Ладант Д., Ченаль А. (январь 2014 г.). «Молекулярная скученность стабилизирует как внутренне неупорядоченное состояние, свободное от кальция, так и свернутое, связанное с кальцием состояние белка RTX: влияние на секрецию токсина» . Биофизический журнал . 106 (2): 271а. Бибкод : 2014BpJ...106R.271S . дои : 10.1016/j.bpj.2013.11.1589 .
- ^ Воудби, Калифорния, Мантл, доктор медицинских наук, Кабрита, Л.Д., Глэдден Л.Ф., Добсон С.М., Христодулу Дж. (июль 2012 г.). «Быстрое различие внутриклеточных и внеклеточных белков с помощью измерений диффузии ЯМР» . Журнал Американского химического общества . 134 (28): 11312–11315. дои : 10.1021/ja304912c . ПМИД 22694283 .
- ^ Крок Р.Л., Саллум К.О., Уотсон Э., Уотт Э.Д., Александреску А.Т. (август 2008 г.). «Водородный обмен мономерного альфа-синуклеина показывает, что развернутая структура сохраняется при физиологической температуре и не зависит от молекулярной скученности в Escherichia coli» . Белковая наука . 17 (8): 1434–1445. дои : 10.1110/ps.033803.107 . ПМК 2492816 . ПМИД 18493022 .
- ^ Бурре Дж., Вивона С., Диао Дж., Шарма М., Брунгер А.Т., Зюдхоф ТК (июнь 2013 г.). «Свойства нативного α-синуклеина мозга» . Природа . 498 (7453): E4–6, обсуждение E6–7. Бибкод : 2013Natur.498E...4B . дои : 10.1038/nature12125 . ПМЦ 4255827 . ПМИД 23765500 .
- ^ Тейлет Ф.С., Бинольфи А., Бекей Б., Марторана А., Роуз Х.М., Стуивер М. и др. (февраль 2016 г.). «Структурное нарушение мономерного α-синуклеина сохраняется в клетках млекопитающих» . Природа . 530 (7588): 45–50. Бибкод : 2016Natur.530...45T . дои : 10.1038/nature16531 . hdl : 11336/53199 . ПМИД 26808899 . S2CID 4461465 .
- ^ Спиллантини М.Г., Шмидт М.Л., Ли В.М., Трояновски Дж.К., Джейкс Р., Годерт М. (август 1997 г.). «Альфа-синуклеин в тельцах Леви» . Природа . 388 (6645): 839–840. Бибкод : 1997Natur.388..839G . дои : 10.1038/42166 . ПМИД 9278044 . S2CID 4419837 .
- ^ Мезей Э., Дехеджиа А., Харта Г., Папп М.И., Полимеропулос М.Х., Браунштейн М.Дж. (июль 1998 г.). «Альфа-синуклеин при нейродегенеративных заболеваниях: убийца или сообщник?» . Природная медицина . 4 (7): 755–757. дои : 10.1038/nm0798-755 . ПМИД 9662355 . S2CID 46196799 .
- ^ Марвиан А.Т., Косс DJ, Алиакбари Ф., Моршеди Д., Оутейро Т.Ф. (сентябрь 2019 г.). «Модели синуклеинопатий in vitro: информация о молекулярных механизмах и защитных стратегиях» . Журнал нейрохимии . 150 (5): 535–565. дои : 10.1111/jnc.14707 . ПМИД 31004503 . S2CID 125080534 .
- ^ Арима К., Хираи С., Сунохара Н., Аото К., Изумияма Ю., Уэда К. и др. (октябрь 1999 г.). «Клеточная совместная локализация фосфорилированных тау- и NACP/альфа-синуклеиновых эпитопов в тельцах Леви при спорадической болезни Паркинсона и при деменции с тельцами Леви». Исследования мозга . 843 (1–2): 53–61. дои : 10.1016/S0006-8993(99)01848-X . ПМИД 10528110 . S2CID 11144367 .
- ^ Арима К., Мизутани Т., Алим М.А., Тонозука-Уэхара Х., Изумияма Ю., Хираи С., Уэда К. (август 2000 г.). «NACP / альфа-синуклеин и тау составляют два различных подмножества филаментов в одних и тех же нейрональных включениях в мозге из семейства паркинсонизма и деменции с тельцами Леви: флуоресцентные и электронно-микроскопические исследования с двойной иммуномаркировкой». Акта Нейропатологика . 100 (2): 115–121. дои : 10.1007/s004010050002 . ПМИД 10963357 . S2CID 22950302 .
- ^ Ёкота О., Терада С., Исидзу Х., Удзике Х., Исихара Т., Накашима Х. и др. (декабрь 2002 г.). «NACP / альфа-синуклеин, NAC и бета-амилоидная патология семейной болезни Альцгеймера с мутацией пресенилина-1 E184D: клинико-патологическое исследование двух случаев аутопсии». Акта Нейропатологика . 104 (6): 637–648. дои : 10.1007/s00401-002-0596-7 . ПМИД 12410385 . S2CID 42542929 .
- ^ Ким Х.И., Хейзе Х., Фернандес К.О., Бальдус М., Цвекстеттер М. (сентябрь 2007 г.). «Корреляция бета-структуры амилоидных фибрилл с развернутым состоянием альфа-синуклеина». ХимБиоХим . 8 (14): 1671–1674. дои : 10.1002/cbic.200700366 . ПМИД 17722123 . S2CID 41870508 .
- ^ Сандал М., Валле Ф., Тессари И., Мамми С., Бергантино Э., Мусиани Ф. и др. (январь 2008 г.). «Конформационные равновесия в мономерном альфа-синуклеине на уровне одиночных молекул» . ПЛОС Биология . 6 (1): e6. дои : 10.1371/journal.pbio.0060006 . ПМК 2174973 . ПМИД 18198943 .
- ^ Шульц-Шеффер WJ (август 2010 г.). «Синаптическая патология агрегации альфа-синуклеина при деменции с тельцами Леви, болезни Паркинсона и деменции при болезни Паркинсона» . Акта Нейропатологика . 120 (2): 131–143. дои : 10.1007/s00401-010-0711-0 . ПМК 2892607 . ПМИД 20563819 .
- ^ Моршеди Д., Алиакбари Ф. (весна 2012 г.). «Ингибирующее действие куминальдегида на амилоидную фибрилляцию и цитотоксичность альфа-синуклеина». Журнал медицинских наук Модареса: патобиология . 15 (1): 45–60.
- ^ Вульф Дж., Хугендорн Х., Тарнопольски М., Муньос Д.Г. (ноябрь 2000 г.). «Моноклональные антитела против вируса Эпштейна-Барра перекрестно реагируют с альфа-синуклеином в мозге человека». Неврология . 55 (9): 1398–1401. дои : 10.1212/WNL.55.9.1398 . ПМИД 11087792 . S2CID 84387269 .
- ^ Полимеропулос М.Х., Лаведан С., Лерой Э., Иде С.Е., Дехеджиа А., Дутра А. и др. (июнь 1997 г.). «Мутация гена альфа-синуклеина выявлена в семьях с болезнью Паркинсона» . Наука . 276 (5321): 2045–2047. дои : 10.1126/science.276.5321.2045 . ПМИД 9197268 .
- ^ Крюгер Р., Кун В., Мюллер Т., Войталла Д., Гребер М., Кёзель С. и др. (февраль 1998 г.). «Мутация Ala30Pro в гене, кодирующем альфа-синуклеин, при болезни Паркинсона». Природная генетика . 18 (2): 106–108. дои : 10.1038/ng0298-106 . ПМИД 9462735 . S2CID 40777043 .
- ^ Зарранс Дж., Алегре Дж., Гомес-Эстебан Х.К., Лескано Э., Рос Р., Ампуэро И. и др. (февраль 2004 г.). «Новая мутация альфа-синуклеина E46K вызывает деменцию с тельцами Паркинсона и Леви». Анналы неврологии . 55 (2): 164–173. дои : 10.1002/ана.10795 . ПМИД 14755719 . S2CID 55263 .
- ^ Аппель-Крессвелл С., Виларино-Гуэлл С., Энкарнасьон М., Шерман Х., Ю И., Шах Б. и др. (июнь 2013 г.). «Альфа-синуклеин p.H50Q, новая патогенная мутация болезни Паркинсона». Двигательные расстройства . 28 (6): 811–813. дои : 10.1002/mds.25421 . ПМИД 23457019 . S2CID 13508258 .
- ^ Лесаж С., Анхайм М., Летурнель Ф., Буссе Л., Оноре А., Розас Н. и др. (апрель 2013 г.). «Мутация α-синуклеина G51D вызывает новый паркинсонически-пирамидный синдром». Анналы неврологии . 73 (4): 459–471. дои : 10.1002/ana.23894 . ПМИД 23526723 . S2CID 43305127 .
- ^ Февга С., Парк Ю., Ломанн Е., Киевит А.Дж., Бридвелд Г.Дж., Ферраро Ф. и др. (август 2021 г.). «Новый миссенс-вариант альфа-синуклеина (Thr72Met) в двух турецких семьях с болезнью Паркинсона» . Паркинсонизм и связанные с ним расстройства . 89 : 63–72. doi : 10.1016/j.parkreldis.2021.06.023 . ПМЦ 8607441 . ПМИД 34229155 .
- ^ Гиассон Б.И., Урю К., Трояновски Дж.К., Ли В.М. (март 1999 г.). «Мутантные альфа-синуклеины человека и дикого типа собираются в удлиненные нити с различной морфологией in vitro» . Журнал биологической химии . 274 (12): 7619–7622. дои : 10.1074/jbc.274.12.7619 . ПМИД 10075647 .
- ^ Флагмейер П., Мейсл Г., Вендруколо М., Ноулз Т.П., Добсон К.М., Бьюэлл А.К., Гальвагнион С. (сентябрь 2016 г.). «Мутации, связанные с семейной болезнью Паркинсона, изменяют этапы инициации и амплификации агрегации альфа-синуклеина» . Труды Национальной академии наук Соединенных Штатов Америки . 113 (37): 10328–10333. Бибкод : 2016PNAS..11310328F . дои : 10.1073/pnas.1604645113 . ПМК 5027465 . ПМИД 27573854 .
- ^ Синглтон А.Б., Фаррер М., Джонсон Дж., Синглтон А., Гаага С., Качергус Дж. и др. (октябрь 2003 г.). «Тройка локуса альфа-синуклеина вызывает болезнь Паркинсона» . Наука . 302 (5646): 841. doi : 10.1126/science.1090278 . ПМИД 14593171 . S2CID 85938327 .
- ^ Шартье-Харлин М.К., Качергус Дж., Румье С., Муру В., Дуэ Х., Линкольн С. и др. (2004). «Дупликация локуса альфа-синуклеина как причина семейной болезни Паркинсона». Ланцет . 364 (9440): 1167–1169. дои : 10.1016/S0140-6736(04)17103-1 . ПМИД 15451224 . S2CID 54419671 .
- ^ Эсламболи А., Ромеро-Рамос М., Бургер С., Бьорклунд Т., Музычка Н., Мандель Р.Дж. и др. (март 2007 г.). «Долгосрочные последствия сверхэкспрессии альфа-синуклеина человека в вентральной части среднего мозга приматов» . Мозг . 130 (Часть 3): 799–815. дои : 10.1093/brain/awl382 . ПМИД 17303591 .
- ^ Гиассон Б.И., Форман М.С., Хигучи М., Гольбе Л.И., Грейвс К.Л., Коцбауэр П.Т. и др. (апрель 2003 г.). «Инициация и синергическая фибриллизация тау и альфа-синуклеина». Наука . 300 (5619): 636–640. Бибкод : 2003Sci...300..636G . дои : 10.1126/science.1082324 . ПМИД 12714745 . S2CID 20223000 .
- ^ Такеда А., Хасимото М., Мэллори М., Сундсумо М., Хансен Л., Маслия Э. (март 2000 г.). «С-концевая иммунореактивность альфа-синуклеина в структурах, отличных от телец Леви, при нейродегенеративных заболеваниях». Акта Нейропатологика . 99 (3): 296–304. дои : 10.1007/PL00007441 . ПМИД 10663973 . S2CID 27393027 .
- ^ Уильямс Т., Соррентино З., Вайнрих М., Гиассон Б.И., Чакрабарти П. (01.07.2020). «Дифференциальные свойства перекрестного посева тау и α-синуклеина в мышиных моделях таупатии и синуклеинопатии» . Мозговые коммуникации . 2 (2): fcaa090. doi : 10.1093/braincomms/fcaa090 . ПМЦ 7567170 . ПМИД 33094280 .
- ^ Прусинер С.Б., Верман А.Л., Мордес Д.А., Уоттс Дж.К., Рамперсауд Р., Берри Д.Б. и др. (сентябрь 2015 г.). «Доказательства того, что прионы α-синуклеина вызывают множественную системную атрофию у людей с паркинсонизмом» . Труды Национальной академии наук Соединенных Штатов Америки . 112 (38): Е5308–Е5317. Бибкод : 2015PNAS..112E5308P . дои : 10.1073/pnas.1514475112 . ПМК 4586853 . ПМИД 26324905 .
- ^ Вейлер Н. (31 августа 2015 г.). «Новый тип приона может вызывать нейродегенерацию» .
- ^ Реттнер Р. (31 августа 2015 г.). «Еще одна смертельная болезнь мозга может возникнуть из-за распространения «прионных» белков» . Проводная наука .
- ^ Куксон М.Р. (февраль 2009 г.). «Альфа-синуклеин и гибель нейрональных клеток» . Молекулярная нейродегенерация . 4 (1): 9. дои : 10.1186/1750-1326-4-9 . ПМЦ 2646729 . ПМИД 19193223 .
- ^ Де Джорджи Ф., Лаферьер Ф., Зингирино Ф., Фаджиани Э., Лендс А., Бертони М. и др. (октябрь 2020 г.). «Новые самореплицирующиеся полиморфы альфа-синуклеина, которые ускользают от мониторинга ThT, могут спонтанно возникать и быстро распространяться в нейронах» . Достижения науки . 6 (40): eabc4364. Бибкод : 2020SciA....6.4364D . дои : 10.1126/sciadv.abc4364 . ПМЦ 7852382 . ПМИД 33008896 .
- ^ Фудзивара Х., Хасегава М., Домаэ Н., Кавасима А., Маслия Э., Голдберг М.С. и др. (февраль 2002 г.). «Альфа-синуклеин фосфорилируется при поражениях синуклеинопатии». Природная клеточная биология . 4 (2): 160–164. дои : 10.1038/ncb748 . ПМИД 11813001 . S2CID 40155547 .
- ^ Версингер С., Сидху А (апрель 2003 г.). «Ослабление активности переносчика дофамина альфа-синуклеином». Письма по неврологии . 340 (3): 189–192. дои : 10.1016/S0304-3940(03)00097-1 . ПМИД 12672538 . S2CID 54381509 .
- ^ Ли Ф.Дж., Лю Ф., Приступа З.Б., Низник Х.Б. (апрель 2001 г.). «Прямое связывание и функциональное соединение альфа-синуклеина с переносчиками дофамина ускоряют апоптоз, индуцированный дофамином» . Журнал ФАСЭБ . 15 (6): 916–926. дои : 10.1096/fj.00-0334com . ПМИД 11292651 . S2CID 3406798 .
- ^ Чой П., Гольц Н., Снайдер Х., Чонг М., Петручелли Л., Харди Дж. и др. (сентябрь 2001 г.). «Коассоциация паркина и альфа-синуклеина». НейроОтчет . 12 (13): 2839–2843. дои : 10.1097/00001756-200109170-00017 . ПМИД 11588587 . S2CID 83941655 .
- ^ Кавахара К., Хасимото М., Бар-Он П., Хо Г.Дж., Крюс Л., Мизуно Х. и др. (март 2008 г.). «Агрегаты альфа-синуклеина мешают растворимости и распределению Паркина: роль в патогенезе болезни Паркинсона» . Журнал биологической химии . 283 (11): 6979–6987. дои : 10.1074/jbc.M710418200 . ПМИД 18195004 .
- ^ Ан Б.Х., Рим Х., Ким С.Ю., Сунг Ю.М., Ли М.Ю., Чхве Дж.Ю. и др. (апрель 2002 г.). «Альфа-синуклеин взаимодействует с изоферментами фосфолипазы D и ингибирует индуцированную перванадатом активацию фосфолипазы D в клетках эмбриональной почки-293 человека» . Журнал биологической химии . 277 (14): 12334–12342. дои : 10.1074/jbc.M110414200 . ПМИД 11821392 . S2CID 85695661 .
- ^ Нейстат М., Ржецкая М., Холодилов Н., Берк Р.Э. (июнь 2002 г.). «Анализ взаимодействий синфилина-1 и синуклеина с помощью жидкостного анализа дрожжевой двухгибридной бета-галактозидазы». Письма по неврологии . 325 (2): 119–123. дои : 10.1016/S0304-3940(02)00253-7 . ПМИД 12044636 . S2CID 11517781 .
- ^ Рид Дж.К., Мейстер Л., Танака С., Кадди М., Юм С., Гейер С., Pleasure D (декабрь 1991 г.). «Дифференциальная экспрессия протоонкогена bcl2 в нейробластоме и других линиях опухолевых клеток человека нервного происхождения». Исследования рака . 51 (24): 6529–6538. ПМИД 1742726 .
- ^ Кавамата Х., Маклин П.Дж., Шарма Н., Хайман Б.Т. (май 2001 г.). «Взаимодействие альфа-синуклеина и синфилина-1: эффект мутаций, связанных с болезнью Паркинсона» . Журнал нейрохимии . 77 (3): 929–934. дои : 10.1046/j.1471-4159.2001.00301.x . ПМИД 11331421 . S2CID 83885937 .
- ^ Энгелендер С., Каминский З., Го Икс, Шарп А.Х., Амарави Р.К., Кляйдерляйн Дж.Дж. и др. (май 1999 г.). «Синфилин-1 связывается с альфа-синуклеином и способствует образованию цитозольных включений». Природная генетика . 22 (1): 110–114. дои : 10.1038/8820 . ПМИД 10319874 . S2CID 2611127 .
- ^ Ли В.М., Гиассон Б.И., Трояновский JQ (март 2004 г.). «Больше, чем две горошины в стручке: общие амилоидогенные свойства тау и альфа-синуклеина при нейродегенеративных заболеваниях». Тенденции в нейронауках . 27 (3): 129–134. doi : 10.1016/j.tins.2004.01.007 . ПМИД 15036877 . S2CID 9545889 .
- ^ Йенсен П.Х., Хагер Х., Нильсен М.С., Хойруп П., Глиманн Дж., Джейкс Р. (сентябрь 1999 г.). «Альфа-синуклеин связывается с тау и стимулирует катализируемое протеинкиназой А тау-фосфорилирование остатков серина 262 и 356» . Журнал биологической химии . 274 (36): 25481–25489. дои : 10.1074/jbc.274.36.25481 . ПМИД 10464279 . S2CID 23877061 .
- ^ Гиассон Б.И., Ли В.М., Трояновский JQ (2003). «Взаимодействия амилоидогенных белков». Нейромолекулярная медицина . 4 (1–2): 49–58. дои : 10.1385/НММ:4:1-2:49 . ПМИД 14528052 . S2CID 9086733 .
- ^ Оно К., Такахаши Р., Икеда Т., Ямада М. (сентябрь 2012 г.). «Перекрестное влияние амилоидного β-белка и α-синуклеина» . Журнал нейрохимии . 122 (5): 883–890. дои : 10.1111/j.1471-4159.2012.07847.x . hdl : 2297/34736 . ПМИД 22734715 . S2CID 17593306 .
Дальнейшее чтение [ править ]
- Блейксли С (27 мая 2002 г.). «Сворачивание белков — ключ ко многим заболеваниям» . Нью-Йорк Таймс .
- Сидеровф А, Конча-Марамбио Л, Лафонтан Д.Э., Фаррис К.М., Ма Ю., Урения П.А., Нгуен Х., Алкалай Р.Н., Шахин Л.М., Форуд Т., Галаско Д., Кибурц К., Мерчант К., Молленхауэр Б., Постон К.Л., Сейбил Дж., Симуни Т., Таннер С.М., Вайнтрауб Д., Виденович А., Чой Ш., Курт Р., Каспелл-Гарсия С., Коффи К.С., Фрейзер М., Оливейра Л.М., Хаттен С.Дж., Шерер Т., Марек К., Сото С. (май 2023 г.). «Оценка гетерогенности среди участников когорты Инициативы по маркерам прогрессирования болезни Паркинсона с использованием семенной амплификации альфа-синуклеина: поперечное исследование» . Ланцет Нейрол . 22 (5): 407–417. дои : 10.1016/S1474-4422(23)00109-6 . ПМЦ 10627170 . ПМИД 37059509 . S2CID 258083747 .
Внешние ссылки [ править ]
 СМИ, связанные с альфа-синуклеином, на Викискладе?
СМИ, связанные с альфа-синуклеином, на Викискладе? - альфа-синуклеин в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)
- человека Местоположение генома SNCA и страница сведений о гене SNCA в браузере генома UCSC .
галерея PDB |
|---|
