Бактериальная систематика
Бактериальная систематика — это раздел систематики, посвященный классификации образцов бактерий по таксономическим рангам.
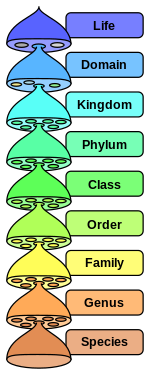
В научной классификации, установленной Карлом Линнеем , [1] каждый вид отнесен к роду, в результате чего получается название, состоящее из двух частей. Это название обозначает два самых низких уровня в иерархии рангов , все более крупные группы видов, основанные на общих чертах. Из этих рангов домены представляют собой наиболее общий уровень категоризации. В настоящее время ученые делят всю жизнь всего на три домена : эукариоты , бактерии и археи . [2]
Бактериальная таксономия — это классификация штаммов внутри домена «Бактерии» по иерархии сходства. Эта классификация аналогична классификации растений , млекопитающих и других таксономий. Однако биологи, специализирующиеся в разных областях, со временем разработали разные таксономические соглашения. Например, систематики бактерий называют типы на основе описаний штаммов. Вместо этого зоологи, в частности, используют типовой образец.
Разнообразие [ править ]
Бактерии ( прокариоты , вместе с археями) имеют много общих черт. Эти общие черты включают отсутствие ядерной мембраны, одноклеточность, деление бинарным делением и, как правило, небольшой размер. Различные виды можно дифференцировать путем сравнения нескольких характеристик, что позволяет их идентифицировать и классифицировать. Примеры включают в себя:
- Филогения : Все бактерии произошли от общего предка и с тех пор разнородны и, следовательно, обладают разными уровнями эволюционного родства (см. Типы бактерий и Хронология эволюции ).
- Метаболизм : разные бактерии могут иметь разные метаболические способности (см. Микробный метаболизм ).
- Окружающая среда : разные бактерии процветают в разных средах, таких как высокая/низкая температура и соль (см. Экстремофилы ).
- Морфология : между бактериями существует множество структурных различий, таких как форма клеток, окраска по Граму (количество липидных бислоев) или состав бислоев (см. Морфология бактериальных клеток , Структура бактериальных клеток ).
История [ править ]
Первые описания [ править ]
Бактерии впервые наблюдал Антони ван Левенгук в 1676 году с помощью однолинзового микроскопа его собственной конструкции. [3] Он назвал их « животными » и опубликовал свои наблюдения в серии писем в Королевское общество . [4] [5] [6]
Ранние описанные роды бактерий включают Vibrio и Monas , О. Ф. Мюллером (1773, 1786), классифицированные тогда как Infusoria (однако многие виды, ранее входившие в эти роды, сегодня считаются протистами); Polyangium , созданный HF Link (1809 г.), первая бактерия, известная и сегодня; Серратия Бицио ; (1823 г.) и Spirillum , Spirochaeta and Bacterium Эренберга ( 1838 ). [7] [8]
Термин Bacterium , введенный как род Эренбергом в 1838 году, [9] стал универсальным для палочковидных клеток. [7]
Ранние классификации формальные
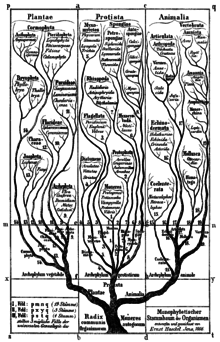
Бактерии были впервые классифицированы как растения , составляющие класс Schizomycetes , которые вместе с Schizophyceae (сине-зеленые водоросли/ цианобактерии ) образовали тип Schizophyta . [11]
Геккель в 1866 году отнес группу к типу Moneres (от μονήρης: простые) в царстве Протисты и определил их как совершенно бесструктурные и однородные организмы, состоящие лишь из кусочка плазмы. [10] Он разделил тип на две группы: [10]
- die Gymnomoneren (без конверта)
- Протогены – такие как Protogenes primordialis , теперь классифицируемые как эукариоты, а не бактерии.
- Протамеба - теперь классифицируется как эукариот, а не бактерия.
- Вибрионы - род бактерий в форме запятой, впервые описанный в 1854 году. [12] )
- Бактерия - род палочковидных бактерий, впервые описанный в 1828 году, который позже дал название членам Monera, ранее называвшемуся «moneron» (множественное число «monera») на английском языке и « eine Moneren » (жен. мн.). « Монерес ») на немецком языке .
- Bacillus - род спорообразующих палочковидных бактерий, впервые описанный в 1835 году. [13]
- Спирохеты - тонкие бактерии спиралевидной формы, впервые описанные в 1835 году. [13]
- Spirillum - бактерии спиралевидной формы, впервые описанные в 1832 году. [14]
- и т. д.
- die Lepomoneren (с конвертом)
- Протомонада - теперь классифицируется как эукариот, а не бактерия. Название было повторно использовано в 1984 году для несвязанного рода бактерий. [15]
- Вампирелла - теперь классифицируется как эукариот, а не бактерия.
Классификация Фердинанда Кона (1872) имела большое влияние в девятнадцатом веке и признавала шесть родов: Micrococcus , Bacterium , Bacillus , Vibrio , Spirillum и Spirochaeta . [7]
группу как прокариотов реклассифицировал Позднее Чаттон . [16]
За классификацию цианобактерий (в просторечии «сине-зеленые водоросли») ведутся споры между водорослями и бактериями (например, Геккель отнес Nostoc к типу водорослей Archephyta). [10] ).
в 1905 году Эрвин Ф. Смит принял 33 действительных различных названия родов бактерий и более 150 недействительных названий. [17] и Виймен в исследовании 1913 года [18] пришел к выводу, что все виды бактерий должны относиться к родам Planococcus , Streptococcus , Klebsiella , Merista , Planomerista , Neisseria , Sarcina , Planosarcina , Metabacterium , Clostridium , Serratia , Bacterium и Spirillum .
Кон [19] выделил четыре трибы : сферобактерии, микробактерии, десмобактерии и спиробактерии. Станье и Ван Нил [20] признал царство Monera с двумя типами: Myxophyta и Schizomycetae, причем последний включает классы Eubacteriae (три отряда), Myxobacteriae (один отряд) и Spirochetae (один отряд). Биссет [21] выделяют 1 класс и 4 порядка: Eubacteriales, Actinomycetales, Streptomycetales и Flexibacteriales. Вальтера Мигулы , Система [22] которая была наиболее широко принятой системой своего времени и включала все известные на тот момент виды, но была основана только на морфологии, содержала три основные группы Coccaceae, Bacillaceae и Spirillaceae, а также Trichobacterinae для нитчатых бактерий. Орла-Йенсен [23] установили два отряда: Cephalotrichinae (семь семейств) и Peritrichinae (предположительно, насчитывающее только одно семейство). Берги и др. [24] представила классификацию, которая в целом следовала Заключительному отчету Комитета Общества американских бактериологов 1920 года (Уинслоу и др.), Который разделил класс Schizomycetes на четыре отряда: Myxobacteriales, Thiobacteriales, Chlamydobacteriales и Eubacteriales, причем пятая группа включает четыре рода, считающихся промежуточными. между бактериями и простейшими: Spirocheta , Cristospira , Saprospira и Treponema .
Однако разные авторы часто переклассифицировали роды из-за отсутствия видимых признаков, что приводило к плохому состоянию, которое было резюмировано в 1915 году Робертом Эрлом Бьюкененом. [25] К тому времени вся группа получила от разных авторов разные звания и названия, а именно:
- Шизомицеты (Нагели, 1857 г.) [11]
- Бактерии (Кон 1872 а) [26]
- Бактерии (Кон 1872 б) [27]
- Schizomycetaceae (ДеТони и Тревизан, 1889 г.) [28]
Более того, семьи, на которые был разделен класс, менялись от автора к автору, а для некоторых, таких как Ципф, имена были на немецком, а не на латыни. [29]
Первое издание Бактериологического кодекса 1947 г. решило ряд проблем. [30] [ нужен пример ]
Система А.Р. Прево [31] [32] ) имел четыре подтипа и восемь классов, а именно:
- Эубактериальные (классы Asporulales и Sporulales)
- Микобактерии (классы Actinomycetales, Myxobacteriales и Azotobacteriales)
- Algobacteriales (классы Siderobacteriales и Thiobacteriales)
- Протозообактерии (класс Spirochetales)
| Линней 1735 [33] | Геккель 1866 [34] | Чаттон 1925 [35] | Коупленд 1938 [36] | Уиттакер 1969 [37] | Вёзе и др. 1990 [38] | Кавалер-Смит 1998, [39] 2015 [40] |
|---|---|---|---|---|---|---|
| 2 королевства | 3 королевства | 2 империи | 4 королевства | 5 королевств | 3 домена | 2 империи, 6/7 королевств |
| (не лечится) | Протиста | Прокариоты | Монера | Монера | Бактерии | Бактерии |
| Архея | Архея (2015) | |||||
| Эукариоты | проктист | Протиста | Эукария | « Простейшие » | ||
| " Хромиста " | ||||||
| Овощи | Растения | Растения | Растения | Растения | ||
| Грибы | Грибы | |||||
| животное | животное | животное | животное | животное |
группы на основе окрашивания Граму по Неформальные
Несмотря на отсутствие единого мнения относительно основных подгрупп бактерий , результаты окрашивания по Граму чаще всего использовались в качестве инструмента классификации. Следовательно, до появления молекулярной филогении царство прокариот было разделено на четыре отдела: [41] Схема классификации, формально до сих пор соблюдаемая руководством Берджи по систематической бактериологии для порядка томов. [42]
- Грациликуты (грамотрицательные)
- Фотобактерии (фотосинтезирующие): класс Oxyphotobacteriae (вода как донор электронов, включает порядок Cyanobacteriales = сине-зеленые водоросли, ныне тип Cyanobacteria ) и класс Anoxyphotobacteriae (анаэробные фототрофы, отряды: Rhodospirillales и Chlorobiales).
- Скотобактерии (нефотосинтезирующие, теперь протеобактерии и другие грамотрицательные нефотосинтетические типы)
- Firmacutes [так в оригинале] (грамположительные, впоследствии исправленные на Firmicutes [43] )
- несколько отрядов, таких как Bacillales и Actinomycetales (теперь в типе Actinobacteria )
- Молликуты (граммовая переменная, например, микоплазма )
- Mendocutes (неравномерное окрашивание по Граму, «метаногенные бактерии», ныне известные как археи )
Молекулярная эра [ править ]
Вёзе реклассификация и «Архаические бактерии »

Вёзе утверждал, что бактерии, археи и эукариоты представляют собой отдельные линии происхождения, которые на раннем этапе отделились от наследственной колонии организмов. [45] [46] Однако некоторые биологи утверждают, что археи и эукариоты произошли от группы бактерий. [47] В любом случае считается, что вирусы и археи начали взаимоотношения примерно два миллиарда лет назад и что совместная эволюция . между членами этих групп могла происходить [48] Вполне возможно, что последний общий предок бактерий и архей был термофилом, что повышает вероятность того, что более низкие температуры являются «экстремальными условиями» с точки зрения архей, а организмы, живущие в более прохладной среде, появились лишь позже. [49] Поскольку археи и бактерии связаны друг с другом не больше, чем с эукариотами, единственное сохранившееся значение термина «прокариот » - «не эукариот», что ограничивает его ценность. [50]
Благодаря усовершенствованным методологиям стало ясно, что метаногенные бактерии сильно отличались друг от друга и (ошибочно) считались остатками древних бактерий. [51] таким образом, Карл Вёзе , считающийся предшественником революции молекулярной филогении, определил три основные линии происхождения: архебактерии , эубактерии и уркариоты , последние теперь представлены нуклеоцитоплазматическим компонентом эукариотов . [52] Эти линии были формализованы в ранг Домена ( regio на латыни), который делил Жизнь на 3 домена: Эукариоты , Археи и Бактерии . [2]
Подразделения [ править ]
В 1987 году Карл Вёзе разделил эубактерии на 11 подразделений на основе последовательностей 16S рибосомальной РНК (SSU), которые с некоторыми дополнениями используются до сих пор. [53] [54]
Оппозиция [ править ]
Хотя трехдоменная система широко распространена, [55] некоторые авторы выступили против этого по разным причинам.
Одним из выдающихся ученых, выступающих против трехдоменной системы, является Томас Кавальер-Смит , который предположил, что археи и эукариоты ( неомура ) произошли от грамположительных бактерий ( позибактерий ), которые, в свою очередь, произошли от грамотрицательных бактерий ( негибактерий ), основанных на нескольких логические аргументы, [56] [57] которые являются весьма противоречивыми и обычно игнорируются сообществом молекулярной биологии ( см. комментарии рецензентов по поводу [57] например, Эрик Баптест является «агностиком» в отношении выводов) и часто не упоминаются в обзорах ( например . [58] ) ввиду субъективного характера сделанных предположений. [59]
Однако, несмотря на множество статистически подтвержденных исследований по укоренению древа жизни между бактериями и неомурами с помощью различных методов, [60] включая некоторые из них, невосприимчивые к ускоренной эволюции, что, по утверждению Кавальер-Смита, является источником предполагаемой ошибки в молекулярных методах. [56] - есть несколько исследований, которые пришли к разным выводам, некоторые из которых относят корень к типу Firmicutes с гнездящимися архей. [61] [62] [63]
Молекулярная таксономия Рэди Гупты, основанная на консервативных сигнатурных последовательностях белков, включает монофилетическую грамотрицательную кладу, монофилетическую грамположительную кладу и полифилетическую археоту, полученную из грамположительных. [64] [65] [66] Молекулярный анализ Хори и Осавы указал на связь между метабактериями (= археотами) и эукариотами. [67] Единственный кладистический анализ бактерий, основанный на классических данных, в значительной степени подтверждает результаты Гупты (см. Комплексную мега-таксономию ).
Джеймс Лейк представил схему из двух первичных царств (паркариоты + эукариоты и эоциты + кариоты) и предложил схему из пяти первичных царств (эукариоты, эоциты, метанобактерии, галобактерии и эубактерии), основанную на рибосомной структуре, и схему из четырех первичных царств (эукариоты, эоциты) . , Methanobacteria и Photocyta ), бактерии классифицируются в соответствии с 3 основными биохимическими инновациями: фотосинтез (Photocyta), метаногенез (Methanobacteria) и серное дыхание ( Eocyta ). [68] [69] [70] Он также обнаружил доказательства того, что грамотрицательные бактерии возникли в результате симбиоза двух грамположительных бактерий. [71]
Власти [ править ]
Классификация — это группировка организмов во все более инклюзивные группы на основе филогении и фенотипа, а номенклатура — это применение формальных правил для наименования организмов. [72]
Номенклатурный орган [ править ]
Несмотря на отсутствие официальной и полной классификации прокариот, названия (номенклатура), данные прокариотам, регулируются Международным кодексом номенклатуры бактерий ( Бактериологическим кодексом ), книгой, содержащей общие соображения, принципы, правила и различные примечания, а также советует [73] аналогично номенклатурным кодам других групп.
Классификационные органы [ править ]
По мере распространения таксонов были разработаны компьютерные таксономические системы. Раннее программное обеспечение для несетевой идентификации, получившее широкое распространение, было создано Эдвардсом 1978, Келлоггом 1979, Шиндлером, Дубеном и Лысенко 1979, Бирсом и Локхардом 1962, Гилленбергом 1965, Холмсом и Хиллом 1985, Лапажем и др. 1970 и Лапажем и др. 1973. [74] : 63
Сегодня таксоны, которые были правильно описаны, рассматриваются в руководстве Берджи по систематической бактериологии , которое призвано помочь в идентификации видов и считается высшим авторитетом. [42] Доступна онлайн-версия таксономического описания бактерий и архей (TOBA) [1] .
Список названий прокариот с номенклатурой (LPSN) представляет собой онлайн-базу данных, которая в настоящее время содержит более двух тысяч принятых названий с их ссылками, этимологией и различными примечаниями. [75]
Описание новых видов [ править ]
Международный журнал систематической бактериологии/Международный журнал систематической и эволюционной микробиологии (IJSB/IJSEM) — это рецензируемый журнал, который выступает в качестве официального международного форума для публикации новых таксонов прокариот. Если вид опубликован в другом рецензируемом журнале, автор может подать запрос в IJSEM с соответствующим описанием, и, если оно правильное, новый вид будет включен в валидационный список IJSEM.
Распространение [ править ]
Коллекции микробных культур представляют собой хранилища штаммов, целью которых является их охрана и распространение. Основными из них являются: [72]
Анализы [ править ]
Этот раздел нуждается в расширении . Вы можете помочь, добавив к нему . ( май 2011 г. ) |
Бактерии сначала классифицировались исключительно на основании их формы (вибрион, палочка, кокк и т. д.), наличия эндоспор, окраски по Граму, аэробных условий и подвижности. Эта система изменилась с изучением метаболических фенотипов, где использовались метаболические характеристики. [76] В последнее время, с появлением молекулярной филогении, для идентификации видов используются несколько генов, наиболее важным из которых является ген 16S рРНК , за которым следуют 23S , область ITS , gyrB и другие для подтверждения лучшего разрешения. Самый быстрый способ идентификации на предмет соответствия выделенного штамма виду или роду сегодня осуществляется путем амплификации его гена 16S с помощью универсальных праймеров, секвенирования ампликона размером 1,4 КБ и отправки его в специализированную идентификационную веб-базу данных, а именно в проект базы данных рибосом . 2] Архивировано 19 августа 2020 года в Wayback Machine , которая выравнивает последовательность с другими последовательностями 16S с использованием инфернальной, вторичной структуры, лежащей в основе глобального выравнивания. [77] [78] или ARB SILVA , который выравнивает последовательности с помощью SINA (инкрементальный выравниватель SILVA), который выполняет локальное выравнивание начального числа и расширяет его [3] . [79]
Существует несколько методов идентификации: [72]
- Фенотипический анализ
- анализ жирных кислот
- Условия роста ( агаровый планшет , многолуночные планшеты Biolog )
- Генетические анализы
- ДНК-ДНК-гибридизация
- Профилирование ДНК
- Последовательность
- Соотношения GC
- Филогенетический анализ
- 16S -основанная филогения
- филогения, основанная на других генах
- Анализ мультигенных последовательностей
- Анализ полногеномной последовательности
Новые виды [ править ]
Минимальные стандарты описания нового вида зависят от того, к какой группе принадлежит вид. см. [80]
Кандидат [ править ]
Candidatus является компонентом таксономического названия бактерии, которую нельзя хранить в коллекции бактериологических культур. Это временный таксономический статус для некультивируемых организмов. например, « Candidatus Pelagibacter ubique »
Концепция вида [ править ]
Бактерии делятся бесполым путем и по большей части не проявляют регионализма (« Все есть повсюду »), поэтому концепция вида, которая лучше всего подходит для животных, становится полностью предметом суждения.
Число названных видов бактерий и архей (около 13 000) [81] удивительно мал, учитывая их раннюю эволюцию, генетическое разнообразие и проживание во всех экосистемах. Причиной этого являются различия в концепциях видов между бактериями и макроорганизмами, трудности выращивания/характеристики в чистой культуре (необходимое условие для наименования новых видов, см. выше ) и обширный горизонтальный перенос генов, стирающий различия между видами. [82]
Наиболее общепринятым определением является определение полифазного вида, которое учитывает как фенотипические, так и генетические различия. [83] Однако более быстрый диагностический порог для разделения видов составляет менее 70% гибридизации ДНК-ДНК. [84] что соответствует менее чем 97% идентичности последовательности 16S ДНК. [85] Было отмечено, что если бы это было применено к классификации животных, отряд приматов был бы одним видом. [86] По этой причине были предложены более строгие определения видов, основанные на последовательностях всего генома. [87]
Патология филогении против
В идеале таксономическая классификация должна отражать эволюционную историю таксонов, то есть филогению. Хотя существуют некоторые исключения, когда фенотип различается среди групп, особенно с медицинской точки зрения. Ниже приведены некоторые примеры проблемных классификаций.
coli : слишком крупная и полифилетичная . Escherichia
В семействе Enterobacteriaceae класса Gammaproteobacteria виды рода Shigella ( S.dysenteriae , S.flexneri , S.boydii , S.sonnei ) с эволюционной точки зрения являются штаммами вида Escherichia coli (полифилетическими), но из-за генетических различий патогенные штаммы вызывают различные заболевания. [88] Как ни странно, существуют также штаммы E. coli , которые производят токсин шига, известный как STEC .
Escherichia coli — плохо классифицированный вид, поскольку некоторые штаммы имеют только 20% общего генома. Будучи настолько разнообразным, ему следует присвоить более высокий таксономический рейтинг. [89] Однако из-за медицинских условий, связанных с этим видом, оно не будет изменено во избежание путаницы в медицинском контексте.
cereus : близкая полифилетичная и Группа Bacillus
Подобным же образом виды Bacillus (=тип Firmicutes ), принадлежащие к « группе B. cereus » ( B. anthracis , B. cereus , B. thuringiensis , B. mycoides , B. pseudomycoides , B. weihenstephanensis и B. medusa ) имеют на 99–100% сходную последовательность 16S рРНК (97% — это часто упоминаемый адекватный предел вида) и являются полифилетическими, но по медицинским причинам (сибирская язва и т. д. ) остаются отдельными. [90]
pestis : чрезвычайно вид недавний Yersinia
Yersinia pestis, по сути, является штаммом Yersinia pseudotuberculosis , но с островом патогенности, вызывающим совершенно другую патологию (черная чума и туберкулезоподобные симптомы соответственно), возникшую 15 000–20 000 лет назад. [91]
роды Pseudomonas Вложенные
В отряде гаммапротеобактерий Pseudomonadales род Azotobacter и вид Azomonas macrocytogenes на самом деле являются членами рода Pseudomonas , но были ошибочно классифицированы из-за способности связывать азот и большого размера рода Pseudomonas , что делает классификацию проблематичной. [76] [92] [93] Вероятно, это будет исправлено в ближайшем будущем.
в Bacillus Вложенные роды
Другим примером большого рода с вложенными родами является род Bacillus , в котором роды Paenibacillus и Brevibacillus представляют собой вложенные клады. [94] В настоящее время недостаточно геномных данных для полного и эффективного исправления таксономических ошибок у Bacillus .
Агробактерии устойчивость к названия смене :
На основе молекулярных данных было показано, что род Agrobacterium гнездится в Rhizobium , а виды Agrobacterium переходят в род Rhizobium (в результате чего образовались следующие комп. нов.: Rhizobium radiobacter (ранее известный как A. tumefaciens) , R. rhizogenes , R. Rubi , R. undicola и R. vitis ). [95] Учитывая фитопатогенную природу видов Agrobacterium , было предложено сохранить род Agrobacterium. [96] и последнее было опровергнуто [97]
Номенклатура [ править ]
Таксономические названия пишутся курсивом (или подчеркиваются при написании от руки) с большой первой буквы, за исключением эпитетов видов и подвидов. Несмотря на то, что это распространено в зоологии, таутонимы (например, Bison Bison ) неприемлемы, а названия таксонов, используемые в зоологии, ботанике или микологии, не могут быть повторно использованы для обозначения бактерий (ботаника и зоология действительно имеют общие названия).
Номенклатура — это набор правил и соглашений, регулирующих названия таксонов. Разница в номенклатуре между различными королевствами/доменами рассматривается в . [98]
Для бактерий допустимые имена должны иметь латинское или неолатинское название и могут использовать только основные латинские буквы (w и j включительно, см. Историю латинского алфавита ), следовательно, дефисы, ударения и другие буквы не принимаются и должны быть правильно транслитерированы. (например, ß=сс). [99] Древнегреческий язык, написанный греческим алфавитом, необходимо транслитерировать в латинский алфавит.
При создании составных слов соединительная гласная необходима в зависимости от происхождения предыдущего слова, независимо от следующего слова, если только последнее не начинается с гласной, и в этом случае соединительная гласная не добавляется. Если первое сложное слово латинское, то соединительной гласной будет -i-, тогда как если первое сложное слово греческое, соединительной гласной будет -o-. [100]
По этимологии имен обратитесь в LPSN .
для высших Правила таксонов
Для прокариотов ( бактерий и архей ) ранговое царство не использовалось до 2024 года. [101] (хотя некоторые авторы называли типы царствами [72] ). Категория царства была включена в Бактериологический кодекс в ноябре 2023 года. [102] первые четыре предложения ( Bacillati , Fusobacteriati , Pseudomonadati , Thermotogati ) были официально опубликованы в январе 2024 года. [103]
В случае помещения нового или измененного вида в новые ранги согласно правилу 9 Бактериологического кодекса название образуется путем добавления соответствующего суффикса к основе названия типового рода. [73] Для подкласса и класса рекомендация от [104] обычно следует, что приводит к нейтральному множественному числу, однако некоторые имена не следуют этому и вместо этого принимают во внимание греко-латинскую грамматику (например, женские множественные числа Thermotogae , Aquificae и Chlamydiae , мужские множественные числа Chloroflexi , Bacilli и Deinococci и греческие множественные числа Spirochaetes , Gemmatimonadetes и Chrysiogenetes ). [105]
| Классифицировать | Суффикс | Пример |
|---|---|---|
| Род | Элюзимикробий | |
| Подтриба (вышла из употребления) | в противном случае | ( Элюзимикробиины ) |
| Племя (вышло из употребления) | -еа | ( Элюзимикробные ) |
| Подсемейство | -oideae | ( Элюзимикробиоидные ) |
| Семья | -aceae | Элюзимикробные |
| Подотряд | -ineae | ( Элюзимикробинеае ) |
| Заказ | -выбранный | Элюзимикробиалы |
| Подкласс | -idae | ( Элюзимикробиды ) |
| Сорт | - это | Элюзимикробия |
| Тип | - заказ | Элюзимикробиота |
| Королевство | -у нас были [102] | Элюзимикробиати |
Окончания типов [ править ]
До 2021 года типы не подпадали под действие Бактериологического кодекса , поэтому назывались неофициально. [105] Это привело к появлению множества подходов к названию типов. Некоторые типы, такие как Firmicutes, были названы в соответствии с общими для всего типа признаками. Другие, такие как Chlamydiae, были названы с использованием названия класса или названия рода в качестве основы (например, Chlamydia ). В 2021 году принято решение включить наименования в Бактериологический кодекс. Следовательно, названия многих типов были обновлены в соответствии с новыми номенклатурными правилами. [106] Высшие таксоны, предложенные Кавалье-Смитом. [56] обычно игнорируются сообществом молекулярной филогении ( например . [58] ) ( см. выше ).
Согласно новым правилам, название типа происходит от типового рода:
- Ацидобактериота (из Acidobacterium )
- Actinomycetota (от Actinomyces )
- Аквификота (от Aquifex )
- Арматимонадота (от Арматимонас )
- Атрибактерита (от Atribacter )
- Бациллота (от Bacillus )
- Бактероидота (от Bacteroides )
- Бальнеолота (из Бальнеолы )
- Бделловибрионота (от Bdellovibrio )
- Кальдисерикота (из Caldisericum )
- Калдитрихота (от Калдитрикса )
- Кампилобактерита (из Campylobacter )
- Хламидиота (из Chlamydia )
- Хлоробиота (из хлоробия )
- Хлорофлексота (от Chloroflexus )
- Хризиогенота (от Chrysiogenes )
- Копротермобактерота (от Coprothermobacter )
- Деферрибактерита (от Deferribacter )
- Дейнококкота (от Deinococcus )
- Диктиогломота (от Dictyoglomus )
- Элюзимикробиота (из Elusimicrobium )
- Фибробактерита (от Fibrobacterota )
- Фузобактериота (из Fusobacterium )
- Гемматимонадота (от Gemmatimonas )
- Игнавибактериота (из Ignavibacterium )
- Киритиматиеллота (от Киритиматиелла )
- Лентисфаэрота (от Lentisphaera )
- Микоплазматота (из Mycoplasma )
- Миксококкота (от Myxococcus )
- Нитроспинота (от Nitrospina )
- Нитроспирота (от Nitrospira )
- Planctomycetota (от Planctomyces )
- Псевдомонадота (от Pseudomonas )
- Родотермота (от Rhodothermus )
- Спирохета (от Spirochaeta )
- Синергистота (от Synergistes )
- Термодесульфобактериота (из Thermodesulfobacterium )
- Термомикробиота (от Thermomicrobium )
- Термотогота (от Термотога )
- Веррукомикробиота (из Verrucomicrobium )
Имена в честь людей [ править ]
Некоторые виды названы в честь людей, либо первооткрывателей, либо известных людей в области микробиологии, например, Salmonella названа в честь Д.Э. Сальмона, который ее открыл (хотя и как «Bacillus typhi»). [107] ). [108]
Для родового эпитета все имена, производные от людей, должны быть в женском именительном падеже путем изменения окончания на -а или уменьшительного -ella, в зависимости от имени. [100]
Для видового эпитета названия могут быть преобразованы либо в форму прилагательного (добавление -nus (м.), -na (ф.), -num (сущ.) в зависимости от пола названия рода), либо в родительный падеж названия рода. Латинизированное имя. [100]
Имена в честь мест [ править ]
Многие виды (видовой эпитет) названы в честь места их обитания или обнаружения (например, Thiospirillum jenense ). Их имена создаются путем образования прилагательного путем соединения названия местности с окончанием -ensis (m. или f.) или ense (n.) в соответствии с полом названия рода, если только для этого места не существует классического латинского прилагательного. . Однако названия мест не должны использоваться как существительные в родительном падеже. [100]
Народные имена [ править ]
Несмотря на то, что некоторые гетеро/гомогенные колонии или биопленки бактерий имеют названия на английском языке (например, зубной налет или звездчатое желе ), ни один вид бактерий не имеет народного/тривиального/распространенного названия на английском языке.
Для имен в единственном числе нельзя образовать множественное число ( single tantum ), поскольку это подразумевает несколько групп с одним и тем же ярлыком, а не несколько членов этой группы (по аналогии, в английском языке стулья и столы — это виды мебели, которые нельзя использовать). во множественном числе «мебель» для описания обоих членов), и наоборот, имена во множественном числе имеют форму множественного числа tantum. Однако частичным исключением из этого правила является использование народных названий.Однако, чтобы избежать повторения таксономических названий, нарушающих ход прозы, часто используются и рекомендуются просторечные названия представителей рода или более высоких таксонов, которые образуются путем написания названия таксонов в римском регистре предложений («стандартные» в MS Office), поэтому имя собственное рассматривается как английское нарицательное существительное (например, сальмонеллы), хотя есть некоторые споры о грамматике множественного числа, которое может быть либо обычным множественным числом, добавляя -(e)s (сальмонеллы), либо использование древнегреческой или латинской формы множественного числа ( неправильное множественное число ) существительного (сальмонеллы); последнее проблематично, так как множественное число от - bacter будет -bacteres, а множественное число слова myces (NL masc. n. от греч. masc. n. mukes) - mycetes. [109]
Для определенных имен присутствуют обычаи, например, имена, оканчивающиеся на -monas, преобразуются в -monad (одна псевдомонада, две аэромонады и не -монады).
Бактерии, являющиеся этиологической причиной заболевания, часто обозначаются по названию заболевания, за которым следует описывающее существительное (бактерия, бацилла, кокк, возбудитель или название их типа), например, бактерия холеры ( Vibrio cholerae ) или спирохета болезни Лайма ( Borrelia ). burgdorferi ), обратите внимание также на риккетсиозную оспа ( Rickettsia akari ) (подробнее см. [110] ).
Трепонема превращается в трепонему, и во множественном числе это трепонемы, а не трепонемы.
Некоторые необычные бактерии имеют особые названия, такие как овал Куина ( Quinella ovalis ) и квадрат Уолсби ( Haloquadratum walsbyi ).
До появления молекулярной филогении многие высшие таксономические группы имели лишь тривиальные названия, которые используются до сих пор, некоторые из которых являются полифилетическими, например, Rhizobacteria. Некоторые более высокие таксономические тривиальные имена:
- Сине-зеленые водоросли относятся к типу « Цианобактерии ».
- Зеленые несерные бактерии относятся к типу Chloroflexota.
- Зеленые серобактерии относятся к группе хлоробиот.
- Пурпурные бактерии — некоторые, но не все, представители типа Pseudomonadota.
- Пурпурные серобактерии относятся к отряду Chromatiales.
- Грамположительные бактерии с низким содержанием G+C относятся к типу Bacillota независимо от содержания GC.
- Грамположительные бактерии с высоким содержанием G+C относятся к типу Actinomycetota независимо от содержания GC.
- Ризобии относятся к различным родам Pseudomonadota.
- Молочнокислые бактерии относятся к отряду Lactobacillales.
- Коринеформные бактерии относятся к семейству Corynebacteriaceae.
- Плодоносящие скользящие бактерии или миксобактерии относятся к типу Myxococcota.
- Энтеробактерии являются членами отряда Enterobacteriales (хотя этот термин следует избегать, если они не живут в кишечнике, например Pectobacterium ).
- Уксуснокислые бактерии относятся к семейству Acetobacteraceae.
Терминология [ править ]
- Аббревиатура вида — sp. (множественное число видов ) и используется после родового эпитета для обозначения вида этого рода. Часто используется для обозначения штамма рода, для которого вид неизвестен либо потому, что организм еще не описан как вид, либо для его идентификации было проведено недостаточно испытаний. Например, Halomonas sp. ГФАД-1
- дается термин Candidatus. Если бактерия известна и хорошо изучена, но не поддается культивированию, в ее названии
- Басоним — это оригинальное название новой комбинации, а именно первое название, данное таксону до его реклассификации.
- Синоним – альтернативное название таксона, т.е. таксон дважды ошибочно описан.
- При переносе таксона он приобретает новую комбинацию (comb. nov.) или новое название (nom. nov.).
- парафилия , монофилия и полифилия
См. также [ править ]
- Порядок ветвления бактериальных типов (Woese, 1987)
- Порядок ветвления бактериальных типов (Гупта, 2001)
- Порядок ветвления бактериальных типов (Кавальер-Смит, 2002)
- Порядок ветвления бактериальных типов (Раппе и Джованони, 2003).
- Порядок ветвления бактериальных типов (Battistuzzi et al.,2004)
- Порядок ветвления бактериальных типов (Ciccarelli et al., 2006)
- Порядок ветвления типов бактерий после живого дерева ARB Silva
- Порядок ветвления типов бактерий (База данных геномной таксономии, 2018)
- Типы бактерий , сложная классификация.
- Список родов архей
- Список родов бактерий
- Список отрядов бактерий
- Список латинских и греческих слов, обычно используемых в систематических названиях
- Список секвенированных геномов архей
- Список секвенированных геномов прокариот
- Список клинически важных бактерий
- Проблема видов
- Эволюционный сорт
- Загадочный видовой комплекс
- Синоним (таксономия)
- Таксономия
- LPSN , список принятых названий бактерий и архей
- Цианобактерии — тип распространенных бактерий, но в настоящее время плохо классифицированный.
- Проект микробиома человека
- Микробная экология
Ссылки [ править ]
- ^ Линней, Карл (1735). Системы природы, или три царства природы, систематически представленные классами, отрядами, родами и видами .
- ^ Перейти обратно: а б Вёзе, ЧР; Кандлер, О.; Уилис, ML (1990). «На пути к естественной системе организмов: предложение по доменам архей, бактерий и эукариев» . Труды Национальной академии наук . 87 (12): 4576–4579. Бибкод : 1990PNAS...87.4576W . дои : 10.1073/pnas.87.12.4576 . ПМК 54159 . ПМИД 2112744 .
- ^ Портер-младший (1976). «Энтони ван Левенгук: трехсотлетие со дня открытия бактерий» . Бактериологические обзоры . 40 (2): 260–9. дои : 10.1128/mmbr.40.2.260-269.1976 . ПМК 413956 . ПМИД 786250 .
- ^ ван Левенгук А (1684 г.). «Резюме письма мистера Энтони Ливвенгука из Делфта от 17 сентября 1683 года, содержащего некоторые микроскопические наблюдения о животных в налете на зубах, о веществе, называемом червями в носу, о кутикуле, состоящей из чешуек. " (PDF) . Философские труды . 14 (155–166): 568–574. Бибкод : 1684RSPT...14..568L . дои : 10.1098/rstl.1684.0030 .
- ^ ван Левенгук А (1700 г.). «Часть письма г-на Антони ван Левенгука о червях в овечьей печени, комарах и животных в экскрементах лягушек» . Философские труды . 22 (260–276): 509–518. Бибкод : 1700RSPT...22..509В . дои : 10.1098/rstl.1700.0013 .
- ^ ван Левенгук А (1702 г.). «Часть письма г-на Энтони ван Левенгука, ФРС, о зеленых сорняках, растущих в воде, и о некоторых животных, обнаруженных вокруг них». Философские труды . 23 (277–288): 1304–11. Бибкод : 1702RSPT...23.1304V . дои : 10.1098/rstl.1702.0042 . S2CID 186209549 .
- ^ Перейти обратно: а б с Мюррей, Р.Г.Е., Холт, Дж.Г. (2005). История « Руководства Бержи» . В: Гаррити, ГМ, Бун, Д.Р. и Кастенхольц, Р.В. (ред., 2001). Руководство Берджи по систематической бактериологии , 2-е изд., том. 1, Спрингер-Верлаг, Нью-Йорк, с. 1-14. связь . [См. стр. 2.]
- ^ Пот Б., Гиллис М. и Де Лей Дж., Род Aquaspirillum. В: Балоуз А., Трюпер Х.Г., Дворкин М., Хардер В., Шлейфер К.-Х. (Ред.). Прокариоты , 2-е изд., т. 3. Шпрингер-Верлаг. Нью-Йорк. 1991 год
- ^ «Этимология слова «бактерии» » . Интернет-словарь этимологии . Архивировано из оригинала 18 ноября 2006 года . Проверено 23 ноября 2006 г.
- ^ Перейти обратно: а б с д Геккель, Эрнст (1867). Общая морфология организмов (на немецком языке). Берлин: Раймер. ISBN 978-1-144-00186-3 – через Интернет-архив.
- ^ Перейти обратно: а б Фон Нэгели, К. (1857). Каспари, Р. (ред.). «Отчет о переговорах на 33-й встрече немецких естествоиспытателей и врачей, проходившей в Бонне с 18 по 24 сентября 1857 года». Ботаническая газета (на немецком языке). 15 : 749–776 — через Интернет-архив .
- ^ Пачини, Ф.: Микроскопические наблюдения и патологические выводы об азиатской холере. Gazette Medicale de Italiana Toscano Florence, 1854, 6, 405–412.
- ^ Перейти обратно: а б Эренберг, К.Г. (1835). «Третий вклад в познание крупной организации в направлении наименьшего пространства». Физические трактаты Королевской академии наук в Берлине за 1833–1835 годы , стр. 143–336.
- ^ Эренберг, К.Г. (1830). «Вклад в знание организации инфузорий и их географического распространения, особенно в Сибири». Трактаты Королевской академии наук в Берлине, 1832 г. , стр. 1–88.
- ^ Протомонада в LPSN ; Парте, Эйдан К.; Сарда Карбасс, Хоаким; Мейер-Колтхофф, Ян П.; Реймер, Лоренц К.; Гёкер, Маркус (1 ноября 2020 г.). «Список названий прокариот, имеющих номенклатурное положение (ЛПСН), перемещается в ДСМЗ» . Международный журнал систематической и эволюционной микробиологии . 70 (11): 5607–5612. дои : 10.1099/ijsem.0.004332 .
- ^ Чаттон, Э. (1925). « Pansporella perplexa : Размышления о биологии и филогении простейших». Анналы естественных наук . 10 (VII): 1–84.
- ^ Смит, Эрвин Ф. (1905). Номенклатура и классификация бактерий в отношении болезней растений , Vol. 1.
- ^ Виймен (1913). «Роды Схизомицетум». Микологические летописи , Том. 11, с. 512–527.
- ^ Кон, Фердинанд (1875). «Исследования о бактериях, II». Вклад в биологию растений , Том 1: стр. 141–207.
- ^ Станье и ван Нил (1941). «Основные положения классификации бактерий». Журнал бактериологии , Vol. 42, стр. 437–466.
- ^ Биссет (1962). Бактерии , 2-е изд.; Лондон: Ливингстон.
- ^ Мигула, Уолтер (1897). Система бактерий . Йена, Германия: Густав Фишер.
- ^ Орла Йенсен (1909). «Основные черты природной бактериальной системы и обзор явлений ферментации». Центральный детектор бактериальных паразитов , Том II, №. 22, стр. 305–346.
- ^ Руководство Берджи по детерминативной бактериологии (1925). Балтимор: Williams & Wilkins Co. (со многими последующими изданиями).
- ^ Бьюкенен, Р.Э. (1916). «Исследования по номенклатуре и классификации бактерий: проблема бактериальной номенклатуры» . Журнал бактериологии . 1 (6): 591–596. дои : 10.1128/jb.1.6.591-596.1916 . ПМЦ 378679 . PMID 16558720 – через Национальный центр биотехнологической информации.
- ^ Кон, Фердинанд (1872a). «Организмы в оспенной лимфе». Вирхов (ред.), Архив , Том 55, с. 237.
- ^ Кон, Фердинанд (1872 б) «Исследования бактерий». Вклад в биологию растений Том 1, №. 1, с. 136.
- ^ Детони, Дж.Б.; Тревизан, В. (1889). «Схизомицетовые». Саккардо (редактор), Sylloge Fungorum , Vol. 8, с. 923
- ^ Бьюкенен, RE (март 1917 г.). «Исследования по номенклатуре и классификации бактерий: II. Первичные подразделения шизомицетов» . Журнал бактериологии . 2 (2): 155–64. дои : 10.1128/JB.2.2.155-164.1917 . ПМК 378699 . ПМИД 16558735 .
- ^ «Международный бактериологический кодекс номенклатуры» . Журнал бактериологии . 55 (3): 287–306. Март 1948 г. doi : 10.1128/JB.55.3.287-306.1948 . ПМК 518444 . ПМИД 16561459 .
- ^ Прево, Арканзас (1958). Руководство по классификации и определению анаэробных бактерий . Париж: Массон.
- ^ Превот, Арканзас (1963). Бактерии в Precis de Botanique . Париж: Массон.
- ^ Линней, К. (1735). Системы природы, или три царства природы, систематически представленные классами, отрядами, родами и видами .
- ^ Геккель, Э. (1866). Общая морфология организмов . Раймер, Берлин.
- ^ Чаттон, Э. (1925). « Pansporella perplexa . Размышления о биологии и филогении простейших». Анналы естественных наук — зоология и биология животных . 10-VII: 1–84.
- ^ Коупленд, Х. (1938). «Царства организмов». Ежеквартальный обзор биологии . 13 (4): 383–420. дои : 10.1086/394568 . S2CID 84634277 .
- ^ Уиттакер, Р.Х. (январь 1969 г.). «Новые представления о царствах организмов». Наука . 163 (3863): 150–60. Бибкод : 1969Sci...163..150W . дои : 10.1126/science.163.3863.150 . ПМИД 5762760 .
- ^ Вёзе, К.; Кандлер, О.; Уилис, М. (1990). «На пути к естественной системе организмов: предложение для доменов архей, бактерий и эукариев» . Труды Национальной академии наук Соединенных Штатов Америки . 87 (12): 4576–9. Бибкод : 1990PNAS...87.4576W . дои : 10.1073/pnas.87.12.4576 . ПМК 54159 . ПМИД 2112744 .
- ^ Кавальер-Смит, Т. (1998). «Пересмотренная система жизни шести королевств» . Биологические обзоры . 73 (3): 203–66. дои : 10.1111/j.1469-185X.1998.tb00030.x . ПМИД 9809012 . S2CID 6557779 .
- ^ Руджеро, Майкл А.; Гордон, Деннис П.; Оррелл, Томас М.; Байи, Николя; Бургуэн, Тьерри; Бруска, Ричард К.; Кавалер-Смит, Томас; Гири, Майкл Д.; Кирк, Пол М.; Туесен, Эрик В. (2015). «Классификация всех живых организмов более высокого уровня» . ПЛОС ОДИН . 10 (4): e0119248. Бибкод : 2015PLoSO..1019248R . дои : 10.1371/journal.pone.0119248 . ПМЦ 4418965 . ПМИД 25923521 .
- ^ ГИББОНС NE; Мюррей Р.Г.Е. (1978). «Предложения о высших таксонах бактерий» . Международный журнал систематической бактериологии . 28 : 1–6. дои : 10.1099/00207713-28-1-1 .
- ^ Перейти обратно: а б Джордж М. Гаррити: Руководство Берджи по систематической бактериологии. 2. Ауфляж. Спрингер, Нью-Йорк, 2005 г., Том 2: Протеобактерии, Часть B: Гаммапротеобактерии.
- ^ МЮРРЕЙ (RGE): Высшие таксоны или место для всего...? В: NR KRIEG и JG HOLT (ред.) Руководство Берджи по систематической бактериологии, том. 1, The Williams & Wilkins Co., Балтимор, 1984, с. 31-34
- ^ Сиссарелли Ф.Д., Доркс Т., фон Меринг С., Криви С.Дж., Снел Б., Борк П. (2006). «К автоматической реконструкции высокоразрешенного древа жизни». Наука . 311 (5765): 1283–7. Бибкод : 2006Sci...311.1283C . CiteSeerX 10.1.1.381.9514 . дои : 10.1126/science.1123061 . ПМИД 16513982 . S2CID 1615592 .
- ^ Вёзе Ч.Р., Гупта Р. (1981). «Являются ли архебактерии просто производными от прокариотов?». Природа . 289 (5793): 95–6. Бибкод : 1981Natur.289...95W . дои : 10.1038/289095a0 . ПМИД 6161309 . S2CID 4343245 .
- ^ Вёзе С (1998). «Всемирный предок» . Труды Национальной академии наук Соединенных Штатов Америки . 95 (12): 6854–9. Бибкод : 1998PNAS...95.6854W . дои : 10.1073/pnas.95.12.6854 . ПМК 22660 . ПМИД 9618502 .
- ^ Гупта РС (2000). «Естественные эволюционные отношения между прокариотами». Крит. Преподобный Микробиол . 26 (2): 111–31. CiteSeerX 10.1.1.496.1356 . дои : 10.1080/10408410091154219 . ПМИД 10890353 . S2CID 30541897 .
- ^ К. Майкл Хоган. 2010. Вирус . Энциклопедия Земли . Редакторы: Катлер Кливленд и Сидни Драгган.
- ^ Грибальдо С., Брошье-Армане С (2006). «Происхождение и эволюция архей: современное состояние» . Философские труды Королевского общества Б. 361 (1470): 1007–22. дои : 10.1098/rstb.2006.1841 . ПМЦ 1578729 . ПМИД 16754611 .
- ^ Вёзе ЧР (1 марта 1994 г.). «Где-то должен быть прокариот: микробиология ищет себя» . Микробиол. Преподобный . 58 (1): 1–9. дои : 10.1128/MMBR.58.1.1-9.1994 . ПМЦ 372949 . ПМИД 8177167 .
- ^ Балч, В.; Магрум, Л.; Фокс, Г.; Вулф, Р.; Вёзе, К. (1977). «Древнее расхождение среди бактерий». Журнал молекулярной эволюции . 9 (4): 305–311. Бибкод : 1977JMolE...9..305B . дои : 10.1007/BF01796092 . ПМИД 408502 . S2CID 27788891 .
- ^ Франколини, К.; Эгет, Х. (1979). «Избирательность восприятия зависит от задачи: эффект всплывающего окна вылезает наружу» . Восприятие и психофизика . 25 (2): 99–110. дои : 10.3758/bf03198793 . ПМИД 432104 .
- ^ Холланд Л. (22 мая 1990 г.). «Вёзе, Карл в авангарде революции в области бактериальной эволюции». Ученый . 4 (10).
- ^ Вёзе, ЧР (1987). «Бактериальная эволюция» . Микробиологические обзоры . 51 (2): 221–271. дои : 10.1128/MMBR.51.2.221-271.1987 . ПМК 373105 . ПМИД 2439888 .
- ^ Уорд, Б.Б. (2002). «Сколько существует видов прокариот?» . Труды Национальной академии наук . 99 (16): 10234–10236. Бибкод : 2002PNAS...9910234W . дои : 10.1073/pnas.162359199 . ПМК 124894 . ПМИД 12149517 .
- ^ Перейти обратно: а б с Кавальер-Смит, Т. (2002). «Неомуранское происхождение архебактерий, негибактериальный корень универсального дерева и бактериальная мегаклассификация» . Международный журнал систематической и эволюционной микробиологии . 52 (Часть 1): 7–76. дои : 10.1099/00207713-52-1-7 . ПМИД 11837318 .
- ^ Перейти обратно: а б Кавальер-Смит, Т. (2006). «Укоренение древа жизни посредством анализа переходов» . Биология Директ . 1:19 . дои : 10.1186/1745-6150-1-19 . ПМК 1586193 . ПМИД 16834776 .
- ^ Перейти обратно: а б Пейс, NR (2009). «Картирование Древа Жизни: прогресс и перспективы» . Обзоры микробиологии и молекулярной биологии . 73 (4): 565–576. дои : 10.1128/MMBR.00033-09 . ПМЦ 2786576 . ПМИД 19946133 .
- ^ Фурнье, врач общей практики; Гогартен, JP (2010). «Укоренение рибосомального древа жизни» . Молекулярная биология и эволюция . 27 (8): 1792–1801. дои : 10.1093/molbev/msq057 . ПМИД 20194428 .
- ^ Фурнье, врач общей практики; Гогартен, JP (2010). «Укоренение рибосомального древа жизни» . Молекулярная биология и эволюция . 27 (8): 1792–1801. дои : 10.1093/molbev/msq057 . ПМИД 20194428 .
- ^ Лейк, Дж.; Сервин, Дж.; Гербольд, К.; Скопхаммер, Р. (2008). «Доказательства нового корня Древа Жизни» . Систематическая биология . 57 (6): 835–843. дои : 10.1080/10635150802555933 . ПМИД 19085327 .
- ^ Скопхаммер, Р.Г.; Сервин, Дж.А.; Гербольд, CW; Лейк, JA (2007). «Доказательства существования грамположительного эубактериального корня Древа жизни» . Молекулярная биология и эволюция . 24 (8): 1761–1768. дои : 10.1093/molbev/msm096 . ПМИД 17513883 .
- ^ Валас, Р.Э.; Борн, ЧП (2011). «Происхождение производного суперцарства: как грамположительная бактерия пересекла пустыню и стала археем» . Биология Директ . 6:16 . дои : 10.1186/1745-6150-6-16 . ПМК 3056875 . ПМИД 21356104 .
- ^ Гупта, Р.С. (1998). «Филогения белков и характерные последовательности: переоценка эволюционных взаимоотношений между архебактериями, эубактериями и эукариотами» . Обзоры микробиологии и молекулярной биологии . 62 (4): 1435–1491. дои : 10.1128/MMBR.62.4.1435-1491.1998 . ПМК 98952 . ПМИД 9841678 .
- ^ Гупта, Р.С. (1998). «Третий домен жизни (Архея): установленный факт или находящаяся под угрозой исчезновения парадигма?». Теоретическая популяционная биология . 54 (2): 91–104. дои : 10.1006/tpbi.1998.1376 . ПМИД 9733652 .
- ^ Гупта, Р.С. (2000). «Естественные эволюционные отношения между прокариотами». Критические обзоры по микробиологии . 26 (2): 111–131. CiteSeerX 10.1.1.496.1356 . дои : 10.1080/10408410091154219 . ПМИД 10890353 . S2CID 30541897 .
- ^ Хори, Х.; Осава, С. (1987). «Происхождение и эволюция организмов, выведенная на основе последовательностей 5S рибосомальной РНК» . Молекулярная биология и эволюция . 4 (5): 445–472. doi : 10.1093/oxfordjournals.molbev.a040455 . ПМИД 2452957 .
- ^ Лейк, Дж.А. (1986). «Альтернатива архебактериальной догме» . Природа . 319 (6055): 626. Бибкод : 1986Natur.319..626L . дои : 10.1038/319626b0 . S2CID 4273843 .
- ^ Лейк, Дж.А.; Хендерсон, Э; Оукс, М; Кларк, М.В. (1984). «Эоциты: новая структура рибосом указывает на царство, имеющее близкое родство с эукариотами» . Труды Национальной академии наук Соединенных Штатов Америки . 81 (12): 3786–3790. Бибкод : 1984PNAS...81.3786L . дои : 10.1073/pnas.81.12.3786 . ПМК 345305 . ПМИД 6587394 .
- ^ Лейк, Дж.А.; Хендерсон, Э.; Кларк, Миссури; Шейнман, А.; Оукс, Мичиган (1986). «Картирование эволюции с помощью трехмерной структуры рибосомы». Систематическая и прикладная микробиология . 7 : 131–136. дои : 10.1016/S0723-2020(86)80135-7 .
- ^ Лейк, Дж. (2009). «Доказательства раннего прокариотического эндосимбиоза». Природа . 460 (7258): 967–971. Бибкод : 2009Natur.460..967L . дои : 10.1038/nature08183 . ПМИД 19693078 . S2CID 4413304 .
- ^ Перейти обратно: а б с д Мэдиган, Майкл (2009). Брок Биология микроорганизмов . Сан-Франциско: Пирсон/Бенджамин Каммингс. ISBN 978-0-13-232460-1 .
- ^ Перейти обратно: а б Лапаж, С.; Снит, П.; Лессель, Э.; Скерман, В.; Силигер, Х.; Кларк, В. (1992). Международный кодекс номенклатуры бактерий: Бактериологический кодекс, редакция 1990 г. Вашингтон, округ Колумбия: ASM Press. ПМИД 21089234 .
- ^ Дворкин, Мартин; Фалькоу, Стэнли; Розенберг, Юджин; Шлейфер, Карл-Хайнц; Штакебрандт, Эрко, ред. (2006). Прокариоты: Справочник по биологии бактерий: симбиотические ассоциации, биотехнология, прикладная микробиология . Том. 1 (3-е изд.). Нью-Йорк : Спрингер . стр. li+959. ISBN 978-0-387-25476-0 . OCLC 262687428 .
- ^ Введение в LPSN ; Парте, Эйдан К.; Сарда Карбасс, Хоаким; Мейер-Колтхофф, Ян П.; Реймер, Лоренц К.; Гёкер, Маркус (1 ноября 2020 г.). «Список названий прокариот, имеющих номенклатурное положение (ЛПСН), перемещается в ДСМЗ» . Международный журнал систематической и эволюционной микробиологии . 70 (11): 5607–5612. дои : 10.1099/ijsem.0.004332 .
- ^ Перейти обратно: а б Паллерони, Нью-Джерси (2010). «История Псевдомонады» . Экологическая микробиология . 12 (6): 1377–1383. Бибкод : 2010EnvMi..12.1377P . дои : 10.1111/j.1462-2920.2009.02041.x . ПМИД 20553550 .
- ^ Олсен, Г.; Ларсен, Н.; Вёзе, К. (1991). «Проект базы данных рибосомальных РНК» . Исследования нуклеиновых кислот . 19 Приложение (Дополнение): 2017–2021 гг. дои : 10.1093/nar/19.suppl.2017 . ПМЦ 331344 . ПМИД 2041798 .
- ^ Коул, младший; Ван, К.; Карденас, Э.; Фиш, Дж.; Чай, Б.; Фаррис, Р.Дж.; Кулам-Сайед-Мохидин, А.С.; МакГаррелл, DM; Марш, Т.; Гаррити, генеральный менеджер; Тидже, Дж. М. (2009). «Проект рибосомальной базы данных: улучшенное выравнивание и новые инструменты для анализа рРНК» . Исследования нуклеиновых кислот . 37 (Проблема с базой данных): D141–D145. дои : 10.1093/нар/gkn879 . ПМЦ 2686447 . ПМИД 19004872 .
- ^ Прюсс, Э.; Кваст, К.; Книттель, К.; Фукс, Б.М.; Людвиг, В.; Пеплиес, Дж.; Глокнер, Ф.О. (2007). «SILVA: комплексный онлайн-ресурс для проверенных и согласованных по качеству данных о последовательностях рибосомальных РНК, совместимых с ARB» . Исследования нуклеиновых кислот . 35 (21): 7188–7196. дои : 10.1093/nar/gkm864 . ПМК 2175337 . ПМИД 17947321 .
- ^ минимальные стандарты в LPSN ; Парте, Эйдан К.; Сарда Карбасс, Хоаким; Мейер-Колтхофф, Ян П.; Реймер, Лоренц К.; Гёкер, Маркус (1 ноября 2020 г.). «Список названий прокариот, имеющих номенклатурное положение (ЛПСН), перемещается в ДСМЗ» . Международный журнал систематической и эволюционной микробиологии . 70 (11): 5607–5612. дои : 10.1099/ijsem.0.004332 .
- ^ «Количество опубликованных имен» .
- ^ Стейли, Джей Ти (2006). «Дилемма бактериального вида и геномно-филогенетическая концепция вида» . Философские труды Королевского общества B: Биологические науки . 361 (1475): 1899–1909. дои : 10.1098/rstb.2006.1914 . ПМЦ 1857736 . ПМИД 17062409 .
- ^ Геверс, Д.; Коэн, FM; Лоуренс, JG; Спратт, Б.Г.; Коэнье, Т.; Фейл, Э.Дж.; Стакебрандт, Э.; Ван Де Пер, Ю.В.; Вандам, П.; Томпсон, Флорида; Качели, Дж. (2005). «Мнение: переоценка видов прокариот» . Обзоры природы Микробиология . 3 (9): 733–739. дои : 10.1038/nrmicro1236 . ПМИД 16138101 . S2CID 41706247 .
- ^ Уэйн, Л. (1988). «Международный комитет по систематической бактериологии: объявление об отчете специального комитета по согласованию подходов к бактериальной систематике». Централблатт по бактериологии, микробиологии и гигиене. Серия А: Медицинская микробиология, инфекционные болезни, вирусология, паразитология . 268 (4): 433–434. дои : 10.1016/s0176-6724(88)80120-2 . ПМИД 3213314 .
- ^ Штакебрандт, Э; Гебель, Б.М. (1994). «Таксономическое примечание: место для реассоциации ДНК-ДНК и анализа последовательности 16S рДНК в современном определении вида в бактериологии» . Межд. Дж. Сист. Бактериол . 44 (4): 846–849. дои : 10.1099/00207713-44-4-846 .
- ^ Стейли, Дж. (1997). «Биоразнообразие: находятся ли виды микробов под угрозой?». Современное мнение в области биотехнологии . 8 (3): 340–345. дои : 10.1016/s0958-1669(97)80014-6 . ПМИД 9206017 .
- ^ Райт, Э.; Баум, Д. (2018). «Эксклюзивность предлагает надежный, но практичный критерий вида для бактерий, несмотря на обильный поток генов» . БМК Геномика . 19 (724): 724. doi : 10.1186/s12864-018-5099-6 . ПМК 6171291 . ПМИД 30285620 .
- ^ Лан, Р; Ривз, PR (2002). «Замаскированная Escherichia coli: молекулярное происхождение шигелл». Микробы и инфекция / Институт Пастера . 4 (11): 1125–32. дои : 10.1016/S1286-4579(02)01637-4 . ПМИД 12361912 .
- ^ Мейер-Колтхофф Дж.П., Ханке Р.Л., Петерсен Дж., Шойнер С., Михаэль В., Фибиг А. и др. (2013). «Полная последовательность генома DSM 30083 (T), типового штамма (U5/41 (T)) Escherichia coli, и предложение по определению подвидов в микробной таксономии» . Стандарты в геномных науках . 9 :2. дои : 10.1186/1944-3277-9-2 . ПМЦ 4334874 . ПМИД 25780495 .
- ^ Бавыкин С.Г.; Лысов Ю.П.; Захарьев В.; Келли, Джей-Джей; Джекман, Дж.; Шталь, Д.А.; Черни, А. (2004). «Использование анализа последовательности генов 16S рРНК, 23S рРНК и gyrB для определения филогенетических связей микроорганизмов группы Bacillus cereus» . Журнал клинической микробиологии . 42 (8): 3711–30. doi : 10.1128/JCM.42.8.3711-3730.2004 . ПМК 497648 . ПМИД 15297521 .
- ^ Ахтман, М.; Цурт, К.; Морелли, Г.; Торреа, Г.; Гиюль, А.; Карниэль, Э. (1999). «Yersinia pestis, возбудитель чумы, является недавно появившимся клоном Yersinia pseudotuberculosis» . Труды Национальной академии наук Соединенных Штатов Америки . 96 (24): 14043–14048. Бибкод : 1999PNAS...9614043A . дои : 10.1073/pnas.96.24.14043 . ПМК 24187 . ПМИД 10570195 .
- ^ Янг, Дж. М.; Парк, округ Колумбия -К. (2007). «Вероятная синонимия азотфиксирующего рода Azotobacter и рода Pseudomonas» . Международный журнал систематической и эволюционной микробиологии . 57 (12): 2894–2901. дои : 10.1099/ijs.0.64969-0 . ПМИД 18048745 .
- ^ Редьерс, Х; Вандерлейден, Дж; Де Мот, Р. (2004). «Azotobacter vinelandii: замаскированная Pseudomonas?» . Микробиология . 150 (Часть 5): 1117–9. дои : 10.1099/mic.0.27096-0 . ПМИД 15133068 .
- ^ Сюй, Д; Коте, JC (2003). «Филогенетические взаимоотношения между видами Bacillus и родственными родами, выведенные на основе сравнения 3'-концевых 16S рДНК и 5'-концевых нуклеотидных последовательностей 16S-23S ITS» . Международный журнал систематической и эволюционной микробиологии . 53 (Часть 3): 695–704. дои : 10.1099/ijs.0.02346-0 . ПМИД 12807189 .
- ^ Янг, Дж.; Куикендалл, Л.; Мартинес-Ромеро, Э.; Керр, А.; Савада, Х. (2001). «Пересмотр Rhizobium Frank 1889 с исправленным описанием рода и включением всех видов Agrobacterium Conn 1942 и Allorhizobium undicola de Lajudie et al. 1998 в качестве новых комбинаций: Rhizobium radiobacter, R. Rhizogenes, R. Rubi, Р. Ундикола и Р. Витис» . Международный журнал систематической и эволюционной микробиологии . 51 (Часть 1): 89–103. дои : 10.1099/00207713-51-1-89 . ПМИД 11211278 .
- ^ Фарранд, С.; Ван Беркум, П.; Огер, П. (2003). «Agrobacterium — определяемый род семейства Rhizobiaceae» . Международный журнал систематической и эволюционной микробиологии . 53 (Часть 5): 1681–1687. дои : 10.1099/ijs.0.02445-0 . ПМИД 13130068 .
- ^ Янг, Дж.; Куикендалл, Л.; Мартинес-Ромеро, Э.; Керр, А.; Савада, Х. (2003). «Классификация и номенклатура агробактерий и ризобиумов» . Международный журнал систематической и эволюционной микробиологии . 53 (Часть 5): 1689–1695. дои : 10.1099/ijs.0.02762-0 . ПМИД 13130069 .
- ^ К. Джеффри. 1989. Биологическая номенклатура, 3-е изд. Эдвард Арнольд, Лондон, 86 стр.
- ^ приложение. 9 до нашей эры https://www.ncbi.nlm.nih.gov/books/NBK8808/
- ^ Перейти обратно: а б с д «Помогите! Латынь! Как избежать наиболее распространенных ошибок при присвоении латинских названий вновь открытым прокариотам» . Список названий прокариот, стоящих в номенклатуре . Архивировано из оригинала 7 октября 2011 года . Проверено 14 апреля 2011 г.
- ^ Классификация типов в LPSN ; Парте, Эйдан К.; Сарда Карбасс, Хоаким; Мейер-Колтхофф, Ян П.; Реймер, Лоренц К.; Гёкер, Маркус (1 ноября 2020 г.). «Список названий прокариот, имеющих номенклатурное положение (ЛПСН), перемещается в ДСМЗ» . Международный журнал систематической и эволюционной микробиологии . 70 (11): 5607–5612. дои : 10.1099/ijsem.0.004332 .
- ^ Перейти обратно: а б Орен, Аарон (1 ноября 2023 г.). «Поправка к Принципу 8, Правилам 5b, 8, 15, 33a и Приложению 7 Международного кодекса номенклатуры прокариот с целью включения категорий царства и домена». Международный журнал систематической и эволюционной микробиологии . 73 (11). дои : 10.1099/ijsem.0.006123 . ПМИД 37909283 .
- ^ Гёкер, Маркус; Орен, Аарон (22 января 2024 г.). «Действительная публикация названий двух доменов и семи царств прокариот». Международный журнал систематической и эволюционной микробиологии . 74 (1). дои : 10.1099/ijsem.0.006242 .
- ^ СТЕКЕБРАНДТ, Э.; РЕЙНИ; УОРД-РЕЙНИ, Нидерланды (1997). «Предложение о новой иерархической системе классификации Actinobacteria classis nov» . Межд. Дж. Сист. Бактериол . 47 (2): 479–491. дои : 10.1099/00207713-47-2-479 .
- ^ Перейти обратно: а б Классификация в ЛПСН ; Парте, Эйдан К.; Сарда Карбасс, Хоаким; Мейер-Колтхофф, Ян П.; Реймер, Лоренц К.; Гёкер, Маркус (1 ноября 2020 г.). «Список названий прокариот, имеющих номенклатурное положение (ЛПСН), перемещается в ДСМЗ» . Международный журнал систематической и эволюционной микробиологии . 70 (11): 5607–5612. дои : 10.1099/ijsem.0.004332 .
- ^ Орен А., генеральный менеджер Гаррити. (2021). «Действительная публикация названий сорока двух типов прокариот» . Int J Syst Evol Microbiol . 71 (10): 5056. doi : 10.1099/ijsem.0.005056 . ПМИД 34694987 . S2CID 239887308 .
- ^ ШРЕТЕР (Дж.). В: Ф. КОН (ред.), Криптогамная флора Силезии. Том 3, Выпуск 3, Грибы. Verlag Ю. Ю. Керна, Бреслау, 1885–1889, стр. 1–814.
- ^ Сальмонелла в LPSN ; Парте, Эйдан К.; Сарда Карбасс, Хоаким; Мейер-Колтхофф, Ян П.; Реймер, Лоренц К.; Гёкер, Маркус (1 ноября 2020 г.). «Список названий прокариот, имеющих номенклатурное положение (ЛПСН), перемещается в ДСМЗ» . Международный журнал систематической и эволюционной микробиологии . 70 (11): 5607–5612. дои : 10.1099/ijsem.0.004332 .
- ^ РЕ БЬЮКЕНАН, Таксономия, Анну. Преподобный Микробиол. 1955.9:1-20. http://www.annualreviews.org/doi/pdf/10.1146/annurev.mi.09.100155.000245
- ^ ftp://ftp.ncbi.nih.gov/pub/taxonomy/