Халоквадратум Уолсби
| Халоквадратум Уолсби | |
|---|---|
 | |
| Haloquadratum walsbyi (рисунок, 2008 г.) | |
 | |
| Haloquadratum walsbyi ( СЭМ- изображение, 2023 г.) | |
| Научная классификация | |
| Домен: | |
| Тип: | |
| Сорт: | |
| Заказ: | |
| Семья: | |
| Род: | |
| Разновидность: | Х. Уолсби |
| Биномиальное имя | |
| Халоквадратум Уолсби Бернс и др. 2007 год | |
Haloquadratum walsbyi — вид архей рода Haloquadratum , известный своей квадратной формой и галофильной природой. [1]
Впервые обнаруженный в соляном бассейне на Синайском полуострове в Египте , H. walsbyi известен своими плоскими клетками квадратной формы и необычной способностью выживать в водной среде с высокими концентрациями хлорида натрия и хлорида магния . [2] [1] вида Родовое название Haloquadratum переводится с греческого и латыни как «соляной квадрат». Эту архею также часто называют «квадратной бактерией Уолсби» из-за ее характерной квадратной формы, которая делает ее уникальной. [3] В соответствии со своим названием Haloquadratum walsbyi наиболее часто встречается в соленых средах.
Haloquadratum walsbyi — фототрофный галофильный архей. Это был единственный признанный вид рода Haloquadratum до 1999 года, когда сообщалось, что Haloarculaquadrata была извлечена из бассейна с рассолом. [2] Haloquadratum walsbyi очень необычен из-за своей уникальной клеточной структуры, напоминающей почти идеально плоскую фигуру.
Этот род впервые был обнаружен в 1980 году британским микробиологом профессором Энтони Э. Уолсби из образцов, взятых из Сабха Гавиш, соленого озера на юге Синая, Египет. Это открытие официально описано в 2007 году Бернсом и др . Попытки культивировать археи были безуспешными до 2004 года и привели к идентификации Haloarculaquadrata , другого вида квадратных архей рода Haloarcula , отличного от H. walsbyi , менее многочисленного и генетически совершенно отличного.
Описание
[ редактировать ]Размер клеток Haloquadratum walsbyi составляет от 2 до 5 микрон и толщина от 100 до 200 нанометров . Археи обычно содержат гранулы полигидроксиалканоатов , и содержат ряд преломляющих вакуолей, заполненных газом которые обеспечивают плавучесть в водной среде и позволяют максимально поглощать свет . Эти газовые вакуоли были открыты Уоллаби в 1980 году при определении принадлежности внутриклеточных преломляющих тел в структуре архей. [3] Они собираются в листы шириной до 40 мкм, но связи между клетками хрупкие и легко разрываются. [4]
Эти организмы можно найти на любом участке очень соленой воды. При испарении морской воды сначала выпадают в осадок карбонат кальция (CaCO 3 ) и сульфат кальция (CaSO 4 ), что приводит к образованию рассола, богатого хлоридом натрия NaCl. Если испарение продолжается, NaCl выпадает в осадок в виде галита , оставляя рассол, богатый хлоридом магния (MgCl 2 ). H. walsbyi процветает во время заключительной фазы осаждения галита и может составлять 80% биомассы этой среды. [ нужна ссылка ] Клетки Haloquadratum walsbyi были признаны грамотрицательными посредством окрашивания, и при выращивании в лаборатории наилучшими условиями для роста является среда с 18% солей при нейтральном pH . [5]
Геном , что был H. walsbyi полностью секвенирован позволяет лучше понять филогенетическую и таксономическую классификацию этого организма и его роль в экосистеме . Геномное сравнение испанских и австралийских изолятов ( штаммы HBSQ001 и C23). Т ) убедительно свидетельствует о быстром глобальном расселении, поскольку они удивительно похожи и сохранили порядок генов . [ нужна ссылка ]
Его рост в лаборатории был получен в среде с очень высокими концентрациями хлоридов (более 2 моль · л). −1 MgCl 2 и более 3 моль·л −1 NaCl), что делает этот организм одним из наиболее галорезистентных из известных. Оптимальная роста температура составляет 40 °C (104 °F), что делает эту архей мезофилом .
![Изображение квадратной клетки Haloquadratum walsbyi, полученное оптической фазово-контрастной микроскопией. Многочисленные светлые точки представляют собой газовые пузырьки, которые позволяют всплывать на поверхность, скорее всего, для приобретения кислорода.[4] Масштабная линейка 1 мкм](//upload.wikimedia.org/wikipedia/commons/thumb/1/14/Optical_phase-contrast_microscopy_image_of_a_Haloquadratum_walsbyi_square_cell_-_PLoS_ONE.png/374px-Optical_phase-contrast_microscopy_image_of_a_Haloquadratum_walsbyi_square_cell_-_PLoS_ONE.png) Изображение квадратной клетки Haloquadratum walsbyi , полученное оптической фазово-контрастной микроскопией . Многочисленные светлые точки представляют собой газовые пузырьки , которые позволяют всплывать на поверхность, скорее всего, для приобретения кислорода. [4]Масштабная линейка 1 мкм
Изображение квадратной клетки Haloquadratum walsbyi , полученное оптической фазово-контрастной микроскопией . Многочисленные светлые точки представляют собой газовые пузырьки , которые позволяют всплывать на поверхность, скорее всего, для приобретения кислорода. [4]Масштабная линейка 1 мкм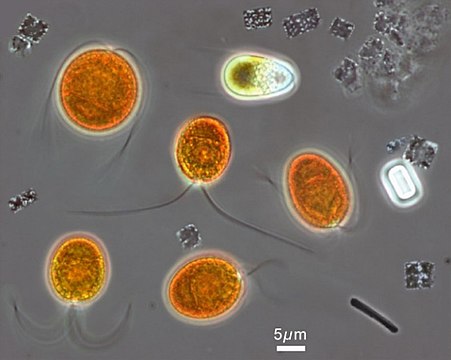 Микроскопическое изображение гиперсоленого озера Тиррелл , на котором можно предварительно идентифицировать оранжевый хлорофит Dunaliella salina в сопровождении ряда более мелких Haloquadratum walsbyi , демонстрирующих их плоские клетки квадратной формы.
Микроскопическое изображение гиперсоленого озера Тиррелл , на котором можно предварительно идентифицировать оранжевый хлорофит Dunaliella salina в сопровождении ряда более мелких Haloquadratum walsbyi , демонстрирующих их плоские клетки квадратной формы.
![Изображение квадратной клетки Haloquadratum walsbyi, полученное оптической фазово-контрастной микроскопией. Многочисленные светлые точки представляют собой газовые пузырьки, которые позволяют всплывать на поверхность, скорее всего, для приобретения кислорода.[4] Масштабная линейка 1 мкм](http://upload.wikimedia.org/wikipedia/commons/thumb/1/14/Optical_phase-contrast_microscopy_image_of_a_Haloquadratum_walsbyi_square_cell_-_PLoS_ONE.png/374px-Optical_phase-contrast_microscopy_image_of_a_Haloquadratum_walsbyi_square_cell_-_PLoS_ONE.png)

Разнообразие
[ редактировать ]Удивительно большое количество клеток в соляных рассолах по всему миру составляет Haloquadratum walsbyi, до 80%. Были проведены эксперименты по изучению генетического разнообразия в среде соленого рассола. семь различных типов геномного острова H. walsbyi . В естественной среде обнаружено [6] После изучения метогеномной фосмид библиотеки H. walsbyi были идентифицированы два типа островков, связанных с клеточной стенкой. Гены в этих островках включают гены, ответственные за синтез структур поверхностного слоя, такие как гликопротеины , и гены, ответственные за синтез клеточных оболочек . [3] Гомологичная рекомбинация отвечает за сохранение упомянутых выше генов, а также за разнообразие метагенома в его естественной среде. Поверхностные структуры разных клеток H. walsbyi помогают дифференцировать источники происхождения для популяции в целом. Эти различные структуры также увеличивают разнообразие клеток в их естественной среде. Эти изменения в клеточной структуре могут быть связаны с попытками клеток снизить свою восприимчивость к атакам вирусов . [6] В 2009 году в Австралии был проведен эксперимент по определению разнообразия H. walsbyi в трех разных прудах-кристаллизаторах с соленой водой. Во всех трех пулах, которые были расположены в разных регионах, все они имели по две 97%-OTU как Haloquadratum , так и Halorubrum -подобных последовательностей. [7]
Геномика и структура
[ редактировать ]H. walsbyi классифицируется как олиготрофный микроорганизм, поскольку он растет в условиях дефицита питательных веществ, когда концентрации органических веществ минимальны. Высокое соотношение поверхности к объему H. walsbyi из-за его уплощенной формы помогает максимизировать поглощение питательных веществ и преодолеть это ограничение. Из-за своей квадратной формы они более способны к сплющиванию, чем сферической формы. микроорганизмы [1] H. walsbyi может сглаживать очень большую величину - около 0,1-0,5 мкм. Общий размер клеточной структуры колеблется от 1,5 до 11 мкм. Однако наблюдались более крупные клетки. Размер самой крупной зарегистрированной клетки H. walsbyi составлял 40 x 40 мкм. [8]
Квадратная форма H. walsbyi была в центре внимания многих исследований. Он способен поддерживать эту структуру благодаря своим адаптивным свойствам . [1] Эти черты можно найти как в H. walsbyi составе генома , так и в его белковых последовательностях. Например, у H.walsbyi экспрессия белка галомуцина создает водный защитный слой, который помогает предотвратить высыхание клеток. [9] Эти адаптации позволяют H. walsbyi процветать в таких средах, как насыщенные рассолы, сохраняя при этом определенную квадратную структуру. [1]
Клеточная структура H. walsbyi состоит из газовых везикул с высокой преломляющей способностью , гранул поли-β-гидроксиалканоата и уникальной клеточной стенки. [9] У этого микроба клеточные стенки имеют толщину от 15 до 25 нм. Геном H.walsbyi кодирует S-слоистые гликопротеины клеточной стенки. Кроме того, фотоактивные белки сетчатки. мембрана также кодирует [9] Штамм HBSQ001, обнаруженный в 2004 году, продемонстрировал те же внутренние клеточные структуры. Однако этот конкретный штамм показал сложную трихотомическую структуру клеточной стенки. [9]
Штамм HBSQ001, DSM 16790 H. walsbyi имеет хромосому длиной 3 132 494 п.н. H. walsbyi отличается аномально низким содержанием гуанина-цитозина (ГЦ) по сравнению с другими галоархей. H. walsbyi имеет среднее содержание GC 47,9% по сравнению с ожидаемыми 60-70%. Кроме того, кодируемые белки высококонсервативны по аминокислотной последовательности. Предполагается, что H. walsbyi произошел от типичного предка, богатого GC и умеренно консервативного. [1]
История
[ редактировать ]Архея Haloquadratum walsbyi была впервые обнаружена в 1980 году профессором микробиологии Энтони Э. Уолсби . [10] Первоначально микроб был назван в его честь как «квадратная бактерия Уолсби», поскольку он был обнаружен до того, как домен архей был полностью признан. [11] Сейчас он официально известен как Haloquadratum walsbyi и считается хорошо известной галофильной архей. Кроме того, он считается одной из первых обнаруженных архей с квадратной формой клеток. [12]
После наблюдения уникальной формы H. walsbyi его выращивание стало целью учёных, изучающих этот вид. Было обнаружено, что гиперсоленая среда является важной средой для поддержания чистых культур. [13] H. walsbyi остается одним из крупнейших известных сегодня прокариотов и содержит около 3 миллионов пар оснований. [13]
Как упоминалось ранее, место открытия этого уникального микроба находилось в трансконтинентальной стране Египет на Синайском полуострове . [4] Однако с этим открытием наступил длительный период, состоявший из интенсивных попыток проб и ошибок добиться полной изоляции H. walsbyi. Из-за того, насколько сложно было полностью изолировать этот микроорганизм, существовал огромный пробел в известной информации о H. walsbyi . физиологических процессах и геномном составе [4] Однако в 2004 году два штамма H. walsbyi были успешно выделены и пригодны для секвенирования . [10] Вторым штаммом был австралийский изолят под названием C23. [10] Дополнительно выделено пять штаммов, всего семь изолятов H. walsbyi. [14] В особой гиперсоленой среде, озере Тиррелл, Haloquadratum walsbyi составлял почти 38% сообщества архей , обнаруженных при культивировании экосистемы. [15]
Нормальная микробиота
[ редактировать ]Archaeon Haloquadratum walsbyi изобилует красными рассолами, солеными озерами и прудами-кристаллизаторами солнечной соли. [16] мелкие водоемы, которые сообщаются друг с другом и повышают соленость. [17] Бактериородопсин , мембранный белок, который использует энергию света для приведения в действие водородно-ионного насоса. [18] которые обнаружены в Haloquadratum walsbyi, поглощают энергию света и встречаются в сообществах в этих рассолах. [16] Использование этих бактериородопсинов показывает фотогетеротрофную природу Haloquadratum walsbyi. Насыщенная солью среда, в которой обитает этот архей, помимо богатства хлоридом магния, имеет очень низкую активность в воде, что вызывает от высыхания стресс . Эти насыщенные солью среды могут иметь соленость, более чем в десять раз превышающую соленость средней морской воды. Насыщенные магнием в этих экосистемах, так называемые выпи , чаще всего встречаются практически без присутствия жизни. [19] Эта среда очень враждебна, и H. walsbyi способен выжить в ней только благодаря своему уникальному геномному составу, в то время как другие организмы погибли бы в аналогичных условиях. [1]
Ссылки
[ редактировать ]- ↑ Перейти обратно: Перейти обратно: а б с д и ж г Болхейс Х., Палм П., Венде А., Фальб М., Рампп М., Родригес-Валера Ф. и др. (июль 2006 г.). «Геном квадратного архея Haloquadratum walsbyi: жизнь в пределах активности воды» . БМК Геномика . 7 : 169. дои : 10.1186/1471-2164-7-169 . ПМЦ 1544339 . ПМИД 16820047 .
- ↑ Перейти обратно: Перейти обратно: а б Орен А., Вентоса А., Гутьеррес М.К., Камекура М. (июль 1999 г.). «Haloarcula Quadata sp. nov., квадратный подвижный архей, выделенный из соляного бассейна на Синае (Египет)» . Международный журнал систематической бактериологии . 49 (3): 1149–1155. дои : 10.1099/00207713-49-3-1149 . ПМИД 10425773 .
Квадратные бактерии были впервые обнаружены в 1980 году Уолсби в Гавиш-Сабха, прибрежном соляном бассейне на Синайском полуострове в Египте (Parkes & Walsby, 1981; Walsby, 1980). Уолсби признал эти чрезвычайно тонкие, квадратные структуры прокариотами...
- ↑ Перейти обратно: Перейти обратно: а б с Остерхельт Д (2022). «Haloquadratum walsbyi – обзор» . Институт биохимии Макса Планка . Макс-Планк-Гезельшафт . Проверено 16 ноября 2022 г.
- ↑ Перейти обратно: Перейти обратно: а б с д Сублими Сапонетти М, Бобба Ф, Салерно Дж, Скарфато А, Корчелли А, Куколо А (апрель 2011 г.). «Морфологические и структурные аспекты чрезвычайно галофильного архея Haloquadratum walsbyi» . ПЛОС ОДИН 6 (4): e18653. Бибкод : 2011PLoSO...618653S . дои : 10.1371/journal.pone.0018653 . ПМК 3084702 . ПМИД 21559517 .
- ^ Бернс Д.Г., Янссен П.Х., Ито Т., Камекура М., Ли З., Дженсен Г. и др. (февраль 2007 г.). «Haloquadratum walsbyi gen. nov., sp. nov., квадратный галоархей Уолсби, выделенный из кристаллизаторов соленой соли в Австралии и Испании» . Международный журнал систематической и эволюционной микробиологии . 57 (Часть 2): 387–392. дои : 10.1099/ijs.0.64690-0 . ПМИД 17267984 .
- ↑ Перейти обратно: Перейти обратно: а б Мартин-Куадрадо AB, Пашич Л, Родригес-Валера Ф (август 2015 г.). «Разнообразие геномного острова, связанного с клеточной стенкой, археи Haloquadratum walsbyi» . БМК Геномика . 16 (1): 603. doi : 10.1186/s12864-015-1794-8 . ПМЦ 4535781 . ПМИД 26268990 .
- ^ О Д., Портер К., Расс Б., Бернс Д., Дьял-Смит М. (март 2010 г.). «Разнообразие Haloquadratum и других галоархей в трех географически удаленных австралийских прудах-кристаллизаторах» . Экстремофилы . 14 (2): 161–169. дои : 10.1007/s00792-009-0295-6 . ПМЦ 2832888 . ПМИД 20091074 .
- ^ Зенке Р., фон Гронау С., Болхейс Х., Груска М., Пфайффер Ф., Остерхельт Д. (2015). «Визуализация флуоресцентной микроскопии галомуцина, секретируемого белка массой 927 кДа, окружающего клетки Haloquadratum walsbyi» . Границы микробиологии . 6 : 249. дои : 10.3389/fmicb.2015.00249 . ПМЦ 4378361 . ПМИД 25870593 .
- ↑ Перейти обратно: Перейти обратно: а б с д Сублими Сапонетти М, Бобба Ф, Салерно Дж, Скарфато А, Корчелли А, Куколо А (апрель 2011 г.). «Морфологические и структурные аспекты чрезвычайно галофильного архея Haloquadratum walsbyi» . ПЛОС ОДИН 6 (4): e18653. Бибкод : 2011PLoSO...618653S . дои : 10.1371/journal.pone.0018653 . ПМК 3084702 . ПМИД 21559517 .
- ↑ Перейти обратно: Перейти обратно: а б с Болуис Х., Мартин-Куадрадо А.Б., Росселли Р., Пашич Л., Родригес-Валера Ф. (июль 2017 г.). «Транскриптомный анализ Haloquadratum walsbyi: тщеславие — это всего лишь поверхность» . БМК Геномика . 18 (1): 510. doi : 10.1186/s12864-017-3892-2 . ПМЦ 5496347 . ПМИД 28673248 .
- ^ Лего Б.А., Лопес-Лопес А., Альба-Касадо Х.К., Дулиттл В.Ф., Болуис Х., Родригес-Валера Ф., Папке Р.Т. (июль 2006 г.). «Экологическая геномика «Haloquadratum walsbyi» в соляном кристаллизаторе указывает на большой пул дополнительных генов у этого в остальном целостного вида» . БМК Геномика . 7 (1): 171. дои : 10.1186/1471-2164-7-171 . ПМК 1560387 . ПМИД 16820057 .
- ^ Лобассо С., Лопалко П., Масколо Дж., Корчелли А. (декабрь 2008 г.). «Липиды ультратонких квадратных галофильных архей Haloquadratum walsbyi» . Архея . 2 (3): 177–183. дои : 10.1155/2008/870191 . ПМЦ 2685597 . ПМИД 19054744 .
- ↑ Перейти обратно: Перейти обратно: а б Болуис Х., Поэле Э.М., Родригес-Валера Ф. (декабрь 2004 г.). «Изоляция и культивирование квадратного архея Уолсби». Экологическая микробиология . 6 (12): 1287–1291. дои : 10.1111/j.1462-2920.2004.00692.x . ПМИД 15560825 .
- ^ Поделл С., Угальде Дж.А., Нарасингарао П., Банфилд Дж.Ф., Гейдельберг КБ, Аллен Э.Э. (18 апреля 2013 г.). «Коммунальная геномика сообщества гиперсоленой микробной экосистемы» . ПЛОС ОДИН . 8 (4): e61692. Бибкод : 2013PLoSO...861692P . дои : 10.1371/journal.pone.0061692 . ПМК 3630111 . ПМИД 23637883 .
- ^ Гай Р., Пашич Л., Фернандес А.Б., Мартин-Куадрадо А.Б., Мизуно С.М., МакМахон К.Д. и др. (31 октября 2011 г.). «Новые многочисленные группы микробов в водных гиперсоленых средах» . Научные отчеты . 1 : 135. Бибкод : 2011НатСР...1Е.135Г . дои : 10.1038/srep00135 . ПМК 3216616 . ПМИД 22355652 .
- ↑ Перейти обратно: Перейти обратно: а б Орен А (2020). «Микробиология красных рассолов». Достижения прикладной микробиологии . 113 : 57–110. дои : 10.1016/bs.aambs.2020.07.003 . ISBN 978-0-12-820709-3 . ПМИД 32948267 . S2CID 221797864 .
- ^ Антон Х, Росселло-Мора Р., Родригес-Валера Ф., Аманн Р. (июль 2000 г.). «Чрезвычайно галофильные бактерии в прудах-кристаллизаторах солнечных соляных заводов» . Прикладная и экологическая микробиология . 66 (7): 3052–3057. Бибкод : 2000ApEnM..66.3052A . дои : 10.1128/aem.66.7.3052-3057.2000 . ПМК 92110 . ПМИД 10877805 .
- ^ Хендерсон Р., Шертлер Г.Ф. (январь 1990 г.). «Структура бактериородопсина и ее отношение к зрительным опсинам и другим семиспиральным рецепторам, связанным с G-белком». Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 326 (1236): 379–389. Бибкод : 1990RSPTB.326..379H . дои : 10.1098/rstb.1990.0019 . ПМИД 1970644 .
- ^ Орен А. (январь 2002 г.). «Разнообразие галофильных микроорганизмов: среда обитания, филогения, физиология и применение». Журнал промышленной микробиологии и биотехнологии . 28 (1): 56–63. дои : 10.1038/sj/jim/7000176 . ПМИД 11938472 . S2CID 24223243 .
Дальнейшее чтение
[ редактировать ]- Лобассо С., Лопалко П., Витале Р., Сапонетти М.С., Капитанио Г., Манджини В. и др. (9 февраля 2012 г.). «Светоактивируемый протонный насос Bop I археи Haloquadratum walsbyi». Фотохимия и фотобиология . 88 (3): 690–700. дои : 10.1111/j.1751-1097.2012.01089.x . ПМИД 22248212 . S2CID 41934280 .
- Куэбас-Ирисарри МФ, Ирисарри-Каро Р.А., Лопес-Моралес К., Бадилло-Ривера К.М., Родригес-Мингела К.М., Монтальво-Родригес Р. (ноябрь 2017 г.). «Клонирование и молекулярная характеристика альфа-глюкозидазы (MalH) из галофильного архея Haloquadratum walsbyi» . Жизнь . 7 (4): 46. дои : 10.3390/life7040046 . ПМЦ 5745559 . ПМИД 29160840 .