Бактериородопсин
| Бактериородопсин | |||
|---|---|---|---|
 Гомотример бактериородопсина Halobacterium salinarum , вид из цитоплазмы. Связанная сетчатка показана черным цветом. ПДБ : 6RQP | |||
| Идентификаторы | |||
| Организм | |||
| Символ | боп | ||
| ЮниПрот | P02945 | ||
| |||
Бактериородопсин (Боп) — белок, используемый , особенно галоархей , классом эвриархей архей . [1] Он действует как протонный насос ; то есть он улавливает энергию света и использует ее для перемещения протонов через мембрану из клетки. [2] Полученный протонный градиент впоследствии преобразуется в химическую энергию. [3]
Функция [ править ]

Бактериородопсин представляет собой светоуправляемый H + переносчик ионов обнаружен у некоторых галоархей, в первую очередь у Halobacterium salinarum (ранее известного как Syn. H. halobium ). генерируемая Протондвижущая сила, белком, используется АТФ-синтазой для выработки аденозинтрифосфата (АТФ) . Экспрессируя бактериородопсин, клетки архей способны синтезировать АТФ в отсутствие источника углерода. [4] [5]
Структура [ править ]

Бактериородопсин представляет собой массой 27 кДа, интегральный мембранный белок обычно встречающийся в двумерных кристаллических участках, известных как «фиолетовая мембрана», которые могут занимать почти 50% площади поверхности клетки архей. Повторяющийся элемент гексагональной решетки состоит из трех одинаковых белковых цепей, каждая из которых повернута на 120 градусов относительно остальных. [6] Каждый мономер имеет семь трансмембранных альфа-спиралей и обращенный к внеклеточному двухцепочечному бета-листу . [7] [8]
Бактериородопсин синтезируется как белок-предшественник , известный как бактериоопсин, который сильно модифицируется после трансляции . [9] [10] Модификации:
- Ковалентное конъюгирование молекулы ретиналя с остатком Lys216 через основание Шиффа для создания ретинилиденового хромофора. [11]
- Расщепление , сигнального пептида первых 13 аминокислот на N-конце и превращение остатка Gln14 в пироглутамат. [12]
- Удаление остатка Asp262 на С-конце [12]
Спектральные свойства [ править ]
Молекула бактериородопсина имеет фиолетовый цвет и наиболее эффективно поглощает зеленый свет (в диапазоне длин волн 500–650 нм ). В нативной мембране белок имеет максимальное поглощение при 553 нм, однако добавление детергента разрушает тримерную форму, что приводит к потере экситонной связи между хромофорами, и, следовательно, мономерная форма имеет максимум поглощения 568 нм. [13] [14]
Бактериородопсин имеет широкий спектр возбуждения. Для длины волны обнаружения от 700 до 800 нм он имеет заметное обнаруженное излучение для длин волн возбуждения от 470 до 650 нм (с пиком при 570 нм). [15] При накачке длиной волны 633 нм спектр излучения имеет заметную интенсивность в диапазоне от 650 до 850 нм. [16]
Механизм [ править ]
Обзор фотоцикла [ править ]
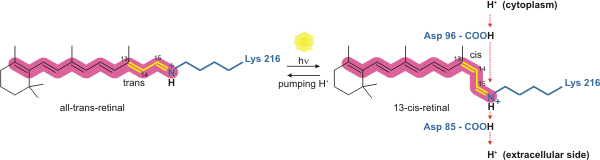
Бактериородопсин – это протонный насос, управляемый светом. Именно молекула сетчатки меняет свое состояние изомеризации с полностью транс на 13- цис при поглощении фотона . Окружающий белок реагирует на изменение формы хромофора, претерпевая упорядоченную последовательность конформационных изменений (известных под общим названием фотоцикл). [17] Конформационные изменения изменяют p K a значения консервативных аминокислот в ядре белка, включая Asp85, Asp96 и атом N основания Шиффа (Lys216). Эти последовательные изменения константы диссоциации кислоты приводят к переносу одного протона с внутриклеточной стороны на внеклеточную сторону мембраны на каждый фотон, поглощенный хромофором.
Фотоцикл бактериородопсина состоит из девяти различных стадий, начиная с основного состояния или состояния покоя, которое обозначается «bR». Промежуточные соединения обозначаются отдельными буквами и могут различаться по спектрам поглощения . [18] Девять стадий таковы:
- БР + фотон → К ⇌ Л ⇌ М 1 ⇌ М 2 ⇌ М 2 ' ⇌ Н ⇌ Н' ⇌ О ⇌ БР [18]
Основное состояние + фотон → состояние K → состояние L [ править ]

Бактериородопсин в основном состоянии поглощает фотон, и сетчатка меняет изомеризацию с полностью транс- 15- анти на напряженное 13- цис -15- анти в состоянии K. Реакция изомеризации быстрая и протекает менее чем за 1 пс. Сетчатка принимает менее напряженную конформацию с образованием интермедиата L.
Состояние L → M 1 Состояние [ править ]
Asp85 принимает протон от атома N основания Шиффа. В промежуточном продукте M1 ни основание Шиффа, ни Asp85 не заряжены.
М 1 Состояние М 2 → Состояние [ править ]
Основание Шиффа поворачивается от внеклеточной стороны белка к цитоплазматической стороне, готовясь принять новый протон.
М 2 Состояние М 2 → Состояние ' [ править ]
Протон высвобождается из Glu204 и Glu194 во внеклеточную среду.
M 2 'состояние → N состояние [ править ]
Шиффовское основание сетчатки принимает протон от Asp96. В состоянии N заряжены как Asp96, так и база Шиффа.
Состояние N → Состояние N [ править ]
Asp96 принимает протон с цитоплазматической стороны мембраны и становится незаряженным.
Состояние N → Состояние O [ править ]
Ретиналь реизомеризуется в полностью транс- состояние.
Состояние О → основное состояние [ править ]
Asp85 переносит протон на Glu194 и Glu204. [19] [20] на внеклеточной стороне белка.
и другие Гомологи подобные белки
Бактериородопсин принадлежит к семейству микробных родопсинов . К его гомологам относятся археродопсины , [21] светоуправляемый хлоридный насос галородопсин (кристаллическая структура которого также известна) и некоторые непосредственно активируемые светом каналы, такие как каналродопсин .
Бактериородопсин подобен позвоночных родопсинам , пигментам , которые воспринимают свет в сетчатке . Родопсины также содержат ретиналь; последовательностей ограничено однако функции родопсина и бактериородопсина различны, и сходство их аминокислотных . И родопсин, и бактериородопсин принадлежат к семейству белков рецепторов 7TM , но родопсин является рецептором, связанным с G-белком , а бактериородопсин - нет. При первом использовании электронной кристаллографии на атомном уровне для получения структуры белка структура бактериородопсина была решена в 1990 году. [22] Затем его использовали в качестве шаблона для создания моделей рецепторов, связанных с G-белком, прежде чем кристаллографические структуры стали доступны для этих белков . Он был тщательно изучен как на слюде, так и на слюде. [23] [24] и стеклянные подложки с использованием атомно-силовой микроскопии и фемтосекундной кристаллографии. [25]
Все остальные фототрофные системы бактерий, водорослей и растений используют хлорофиллы или бактериохлорофиллы, а не бактериородопсин. Они также создают протонный градиент, но совершенно другим и более косвенным путем, задействуя цепь переноса электронов, состоящую из нескольких других белков. Кроме того, хлорофиллам помогают улавливать световую энергию другие пигменты, известные как «антенны»; их нет в системах на основе бактериородопсина. Возможно, фототрофия независимо развивалась как минимум дважды: один раз у бактерий и один раз у архей.
Галерея [ править ]
![Одиночный мономер бактериородопсина с молекулой сетчатки между 7 вертикальными альфа-спиралями (идентификатор PDB: 1X0S [26][27][28]). Еще одна маленькая спираль — светло-голубая, бета-лист — желтый.](//upload.wikimedia.org/wikipedia/commons/2/21/Bacteriorhodopsin_subunit_1X0S.png) Одиночный мономер бактериородопсина с молекулой сетчатки между 7 вертикальными альфа-спиралями ( PDB : 1X0S). ID [26] [27] [28] ). Еще одна маленькая спираль — светло-голубая, бета-лист — желтый.
Одиночный мономер бактериородопсина с молекулой сетчатки между 7 вертикальными альфа-спиралями ( PDB : 1X0S). ID [26] [27] [28] ). Еще одна маленькая спираль — светло-голубая, бета-лист — желтый.![Тример бактериородопсина с одной молекулой сетчатки в каждой субъединице, видимой с внеклеточной стороны EC (ID PDB: 1X0S [26][27][28]).](//upload.wikimedia.org/wikipedia/commons/c/c4/Bacteriorhodopsin_trimer_1X0S.png)
![Одиночный мономер бактериородопсина с молекулой сетчатки между 7 вертикальными альфа-спиралями (идентификатор PDB: 1X0S [26][27][28]). Еще одна маленькая спираль — светло-голубая, бета-лист — желтый.](http://upload.wikimedia.org/wikipedia/commons/2/21/Bacteriorhodopsin_subunit_1X0S.png)
![Тример бактериородопсина с одной молекулой сетчатки в каждой субъединице, видимой с внеклеточной стороны EC (ID PDB: 1X0S [26][27][28]).](http://upload.wikimedia.org/wikipedia/commons/c/c4/Bacteriorhodopsin_trimer_1X0S.png)
См. также [ править ]
Литература [ править ]
- ^ Шох С.Л., Чуфо С., Домрачев М., Хоттон С.Л., Каннан С., Хованская Р., Лейпе Д., Маквей Р., О'Нил К., Робберце Б., Шарма С., Суссов В., Салливан Дж.П., Сан Л., Тернер С., Карш-Мизрачи (2020). «Галобактерии» . Таксономия NCBI: комплексная обновленная информация о курировании, ресурсах и инструментах . Национальный центр биотехнологической информации . Проверено 31 марта 2021 г.
- ^ Воэт, Джудит Г .; Воэт, Дональд (2004). Биохимия . Нью-Йорк: Дж. Уайли и сыновья. ISBN 978-0-471-19350-0 .
- ^ «Бактериородопсин: перекачка ионов» .
- ^ Николлс Д.Г .; Фергюсон С.Дж. (1992). Биоэнергетика 2 (2-е изд.). Сан-Диего: Академическая пресса. ISBN 9780125181242 .
- ^ Страйер, Люберт (1995). Биохимия (четвертое изд.). Нью-Йорк - Бейзингсток: WH Freeman and Company. ISBN 978-0716720096 .
- ^ Эссен Л.О., Зигерт Р., Леман В.Д., Остерхельт Д. (1998). «Липидные пятна в олигомерах мембранных белков: кристаллическая структура бактериородопсина-липидного комплекса» . Труды Национальной академии наук Соединенных Штатов Америки . 95 (20): 11673–11678. Бибкод : 1998PNAS...9511673E . дои : 10.1073/pnas.95.20.11673 . ПМК 21699 . ПМИД 9751724 .
- ^ Пебай-Перуа Э., Раммель Г., Розенбуш Дж. П., Ландау Э. М. (1997). «Рентгеновская структура бактериородопсина при 2,5 Å из микрокристаллов, выращенных в липидных кубических фазах». Наука . 277 (5332): 1676–1681. дои : 10.1126/science.277.5332.1676 . ПМИД 9287223 .
- ^ Люке Х., Шоберт Б., Рихтер Х.Т., Картайллер Дж.П., Лани Дж.К. (1999). «Структура бактериородопсина с разрешением 1,55 Å». Журнал молекулярной биологии . 291 (4): 899–911. дои : 10.1006/jmbi.1999.3027 . ПМИД 10452895 .
- ^ Остерхельт, Дитер ; Стокениус, Вальтер (1971). «Родопсинподобный белок из пурпурной мембраны Halobacterium halobium». Новая биология природы . 233 (39): 149–152. дои : 10.1038/newbio233149a0 . ПМИД 4940442 .
- ^ Остерхельт, Дитер (1982). «[3] Восстановление белков сетчатки бактериородопсина и галородопсина». Восстановление белков сетчатки бактериородопсина и галородопсина . Методы энзимологии. Том. 88. стр. 10–17. дои : 10.1016/0076-6879(82)88006-3 . ISBN 9780121819880 .
- ^ Бэйли Х., Хуанг К.С., Радхакришнан Р., Росс А.Х., Такагаки Ю., Хорана Х.Г. (1981). «Место прикрепления сетчатки в бактериородопсине» . Труды Национальной академии наук Соединенных Штатов Америки . 78 (4): 2225–2229. Бибкод : 1981PNAS...78.2225B . дои : 10.1073/pnas.78.4.2225 . ПМК 319317 . ПМИД 6941281 .
- ↑ Перейти обратно: Перейти обратно: а б Хой К.К., Бада Хуарес Х.Ф., судья П.Дж., Йен ХИ, Ву Д., Виналс Дж., Тейлор Г.Ф., Уоттс А., Робинсон К.В. (2021). «Наночастицы Lipodisq, не содержащие детергентов, облегчают масс-спектрометрию высокого разрешения свернутых интегральных мембранных белков» . Нано-буквы . 21 (7): 2824–2831. Бибкод : 2021NanoL..21.2824H . дои : 10.1021/acs.nanolett.0c04911 . ПМК 8050825 . ПМИД 33787280 .
- ^ Ван Дж., Линк С., Хейес КД, Эль-Сайед М.А. (2002). «Сравнение динамики первичных событий бактериородопсина в его тримерном и мономерном состояниях» . Биофизический журнал . 83 (3): 1557–1566. Бибкод : 2002BpJ....83.1557W . дои : 10.1016/S0006-3495(02)73925-8 . ПМК 1302253 . ПМИД 12202380 .
- ^ Пескителли Дж., Вуди Р.В. (2012). «Экситонное происхождение спектра видимого кругового дихроизма бактериородопсина». Журнал физической химии Б. 116 (23): 6751–6763. дои : 10.1021/jp212166k . ПМИД 22329810 .
- ^ Шенкль, Сельма; Зграблич, Горан; Портуондо-Кампа, Эрвин; Хааке, Стефан; Шерги, Маджед (2007). «О зависимости флуоресценции бактериородопсина от длины волны возбуждения». Письма по химической физике . 441 (4–6): 322–326. Бибкод : 2007CPL...441..322S . дои : 10.1016/j.cplett.2007.04.086 .
- ^ Отани, Х.; Цукамото, Ю.; Сакода, Ю.; Хамагучи, Х. (1995). «Спектры флуоресценции бактериородопсина и промежуточных продуктов O и Q при комнатной температуре». ФЭБС Летт . 359 (1): 65–68. дои : 10.1016/0014-5793(94)01440-c . ПМИД 7851532 .
- ^ Хаяши С., Таджхоршид Э., Шультен К. (сентябрь 2003 г.). «Молекулярно-динамическое моделирование фотоизомеризации бактериородопсина с использованием ab initio сил для возбужденного хромофора» . Биофизический журнал . 85 (3): 1440–9. Бибкод : 2003BpJ....85.1440H . дои : 10.1016/S0006-3495(03)74576-7 . ПМК 1303320 . ПМИД 12944261 .
- ↑ Перейти обратно: Перейти обратно: а б Эрнст О.П., Лодовски Д.Т., Эльстнер М., Хегеман П., Браун Л.С., Кандори Х. (2014). «Микробные и животные родопсины: структуры, функции и молекулярные механизмы» . Химические обзоры . 114 (1): 126–163. дои : 10.1021/cr4003769 . ПМЦ 3979449 . ПМИД 24364740 .
- ^ Дюмаев А.К.; Рихтер, ХТ; Браун, Л.С.; Танио, М.; Тузи, С.; Сайто, Х.; Кимура, Ю.; Нидлман, Р.; Ланьи, Дж. К. (1998). «Существование цепи переноса протонов в бактериородопсине: участие Glu-194 в высвобождении протонов на внеклеточную поверхность» . Биохимия . 37 (8): 2496–2906. дои : 10.1021/bi971842m . ПМИД 9485398 .
- ^ Балашов, ИП; Лу, М.; Имашева Е.С.; Говинджи, Р.; Эбри, Т.Г.; Озерсен б, 3-й; Чен, Ю.; Крауч, РК; Меник, ДР (1999). «Группа высвобождения протонов бактериородопсина контролирует скорость последней стадии его фотоцикла при низком pH» . Биохимия . 38 (7): 2026–2039. дои : 10.1021/bi981926a . ПМИД 10026285 .
{{cite journal}}: CS1 maint: числовые имена: список авторов ( ссылка ) - ^ Бада Хуарес Дж.Ф., судья П.Дж., Адам С., Аксфорд Д., Виналс Дж., Берч Дж., Кван Т.О., Хой К.К., Йен ХИ, Виал А, Милхиет П.Е., Робинсон К.В., Шапиро I, Мораес И., Уоттс А (2021). «Структуры транспортера археродопсина 3 показывают, что нарушение внутренних водных сетей лежит в основе сенсибилизации рецепторов» . Природные коммуникации . 12 (1): 629. Бибкод : 2021NatCo..12..629B . дои : 10.1038/s41467-020-20596-0 . ПМЦ 7840839 . PMID 33504778 .
- ^ Хендерсон Р., Болдуин Дж. М., Ческа Т. А., Землян Ф., Бекманн Э., Даунинг К. Х. (1990). «Модель структуры бактериородопсина на основе электронной криомикроскопии высокого разрешения». Дж Мол Биол . 213 (4): 899–929. дои : 10.1016/S0022-2836(05)80271-2 . ПМИД 2359127 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Мюллер, Дэниел Дж.; Дюфрен, Ив Ф. (2008). «Атомно-силовая микроскопия как многофункциональный молекулярный инструментарий в нанобиотехнологии». Природные нанотехнологии . 3 (5): 261–269. Бибкод : 2008НатНа...3..261М . дои : 10.1038/nnano.2008.100 . ISSN 1748-3387 . ПМИД 18654521 .
- ^ Сибата, Микихиро; Ямасита, Хаято; Учихаши, Такаюки; Кандори, Хидеки; Андо, Тосио (14 февраля 2010 г.). «Высокоскоростная атомно-силовая микроскопия показывает динамические молекулярные процессы в фотоактивированном бактериородопсине» . Природные нанотехнологии . 5 (3): 208–212. Бибкод : 2010НатНа...5..208С . дои : 10.1038/nnano.2010.7 . hdl : 2297/23872 . ISSN 1748-3387 . ПМИД 20154686 .
- ^ Нанго, Эрико; Руайан, Антуан; Кубо, Минору; Накане, Таканори; Викстранд, Сесилия; Кимура, Тецунари; Танака, Томоюки; Тоно, Кенсуке; Сон, Чангён (23 декабря 2016 г.). «Трехмерный фильм структурных изменений бактериородопсина» . Наука . 354 (6319): 1552–1557. Бибкод : 2016Sci...354.1552N . дои : 10.1126/science.aah3497 . ISSN 0036-8075 . ПМИД 28008064 . S2CID 206651572 .
- ↑ Перейти обратно: Перейти обратно: а б Нисикава, Т.; Мураками, М. (28 марта 2005 г.). «Кристаллическая структура 13-цис-изомера бактериородопсина» . Банк данных белков RCSB (PDB). дои : 10.2210/pdb1x0s/pdb . Идентификатор PDB: 1X0S . Проверено 7 октября 2012 г.
{{cite journal}}: Для цитирования журнала требуется|journal=( помощь ) - ↑ Перейти обратно: Перейти обратно: а б Нисикава, Т.; Мураками, М. (2005). «Кристаллическая структура 13-цис-изомера бактериородопсина в темно-адаптированном состоянии». Дж. Мол. Биол . 352 (2): 319–328. дои : 10.1016/j.jmb.2005.07.021 . ПМИД 16084526 . Идентификатор PDB: 1X0S.
- ↑ Перейти обратно: Перейти обратно: а б Изображение создано с помощью RasTop (программное обеспечение для молекулярной визуализации).
Внешние ссылки [ править ]
- Бактериородопсин: молекула месяца , Дэвид Гудселл, Банк данных белков RCSB
- Производство искусственной сетчатки на основе белков: характеристика функции и стабильности бактериородопсина после воздействия микрогравитационной среды , Николь Вагнер и Джордан Греко