Протеородопсин
Эта статья может быть слишком технической для понимания большинства читателей . ( Май 2016 г. ) |
| Протеородопсин | |
|---|---|
 Визуализация мультфильма о протеородопсине от EViture | |
| Идентификаторы | |
| Символ | Бак_родопсин |
| ИнтерПро | ИПР017402 |
| ОБЛАСТЬ ПРИМЕНЕНИЯ 2 | 2сп / СКОПе / СУПФАМ |
| TCDB | 3.E.1 |
| Суперсемейство OPM | 6 |
| белок OPM | 4hyj |
Протеородопсин (также известный как pRhodopsin ) представляет собой семейство , трансмембранных белков которые используют ретиналь в качестве хромофора для светоопосредованной функциональности, в данном случае протонного насоса . рРодопсин обнаружен у морских планктонных бактерий , архей и эукариотов ( протов ), но впервые был обнаружен у бактерий. [ 1 ] [ 2 ] [ 3 ] [ 4 ]
Его название происходит от протеобактерий (теперь называемых Pseudomonadota ), которые были названы в честь древнегреческого Πρωτεύς ( Протея ), раннего морского бога, упомянутого Гомером как « Морской старик », Ῥόδος (rhódon) означающего « роза », из-за его розоватый цвет и ὄψις ( opsis ) — «зрение». Некоторые члены семейства гомологичных родопсин -подобных пигментов , то есть бактериородопсин (которых насчитывается более 800 типов), обладают сенсорными функциями, подобными опсинам , необходимым для зрительной фототрансдукции . Многие из этих сенсорных функций неизвестны – например, функция нейропсина в сетчатке человека. [ 5 ] Известно, что члены имеют разные спектры поглощения , включая зеленый и синий видимый свет . [ 6 ] [ 7 ] [ 8 ] [ 9 ] [ 10 ] [ 11 ]
История
[ редактировать ]Протеородопсин (PR или pRhodopsin) был впервые обнаружен в 2000 году в бактериальной искусственной хромосоме ранее некультивируемых морских гаммапротеобактерий , которые до сих пор упоминаются только по метагеномным данным их риботипа , SAR86. Было обнаружено, что больше видов гаммапротеобактерий, как грамположительных , так и грамотрицательных , экспрессируют этот белок. [ 1 ]
Распределение
[ редактировать ]Образцы бактерий, экспрессирующих протеородопсин, были получены из восточной части Тихого океана , центральной части северной части Тихого океана и Южного океана , Антарктиды . [ 12 ] Впоследствии гены вариантов протеородопсина были идентифицированы в образцах Средиземного , Красного , Саргассова , Японского и Северного морей . [ 4 ] [ 6 ]
Варианты протеородопсина не распространяются случайным образом, а рассредоточиваются по градиентам глубины на основе максимальной настройки поглощения конкретной голопротеиновой последовательности ; в основном это происходит из-за электромагнитного поглощения водой , которое создает градиенты длин волн относительно глубины. Oxyris marina — это динофлагеллятный протист с поглощающим зеленый цвет протеородопсином (продукт группы L109), который обитает в основном в мелких приливных водоемах и на берегах, где все еще доступен зеленый свет. Karlodinium micrum , еще один динолагелят, экспрессирует протеородопсин синего цвета (E109), что может быть связано с его глубоководными вертикальными миграциями . [ 3 ] O. marina Первоначально считалось, что является гетеротрофом , однако протеородопсин вполне может участвовать функционально значимым образом, поскольку он был наиболее широко экспрессируемым ядерным геном и, кроме того, неравномерно распределен в организме, что указывает на некоторую органеллы мембранную функцию . Ранее единственными известными эукариотическими белками, передающими солнечную энергию, были Фотосистема I и Фотосистема II . Была выдвинута гипотеза, что латеральный перенос генов - это метод, с помощью которого протеородопсин проник во многие типы. Бактерии, археи и эукарии колонизируют фотозону , где они выходят на свет; Протеородопсин смог распространиться через эту зону, но не в другие части водной толщи. [ 3 ] [ 4 ] [ 9 ] [ 13 ] [ 14 ]
Таксономия
[ редактировать ]Протеородопсин принадлежит к семейству сходных ретинилиденовых белков, наиболее сходных со своими архейными гомологами галородопсином и бактериородопсином. Сенсорный родопсин был открыт Францем Кристианом Боллом в 1876 году. [ 11 ] [ 15 ] Бактериородопсин был открыт в 1971 году и назван в 1973 году, и в настоящее время известно, что он существует только у архей, а не у бактерий. [ 16 ] Галородопсин был впервые обнаружен и назван в 1977 году. [ 17 ] Бактериородопсин и галородопсин существуют только у архей, тогда как протеородопсин встречается у бактерий, архей и эукариот. Протеородопсин разделяет семь трансмембранных α-спиралей сетчатки, ковалентно связанных по механизму основания Шиффа с остатком лизина в седьмой спирали (спираль G). Бактериородопсин, как и протеородопсин, представляет собой протонный насос, управляемый светом. Сенсорный родопсин — это G-связанный белок, участвующий в зрении. [ 1 ] [ 17 ]
Активный сайт
[ редактировать ]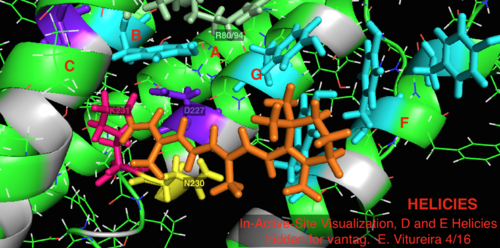
По сравнению с его более известным архейным гомологом бактериородопсином, большинство остатков активного центра, имеющих известное значение для механизма бактериородопсина, консервативны в протеородопсине. Однако сходство последовательностей существенно не сохраняется ни для гало-, ни для бактериородопсина. Гомологи активного центра остатков Arg 82, Asp 85 (первичный акцептор протонов ), Asp 212 и Lys 216 ( сетчатки сайт связывания оснований Шиффа ) в бактериородопсине консервативны как Arg94, Asp97, Asp227 и Lys231 в протеородопсине. Однако в протеородопсине нет остатков карбоновой кислоты , непосредственно гомологичных Glu 194 или Glu204 бактериородопсина (или Glu 108 и 204 в зависимости от варианта бакродопсина), которые, как полагают, участвуют в пути высвобождения протонов на внеклеточной поверхности. Однако Asp97 и Arg94 могут заменять эту функциональность без близкого соседства остатков, как в бактериородопсине. Факультет химии Сиракузского университета решительно доказал, что Asp97 не может быть группой, высвобождающей протоны, поскольку высвобождение произошло в условиях, при которых группа аспарагиновой кислоты оставалась протонированной. [ 18 ] [ 19 ] [ 20 ] [ 21 ]
Лиганд
[ редактировать ]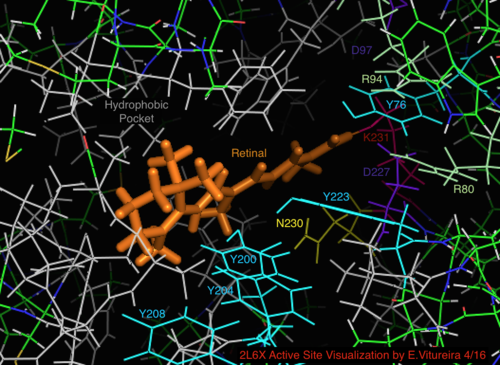
Семейство галопротеинов родопсина имеет общий лиганд ретиналь, один из многих типов витамина А. Ретиналь представляет собой конъюгированный полиненасыщенный β-каротин - хромофор ( полиен ), получаемый в результате диеты плотоядных животных или каротиновым путем ( 15,15'-моноксигеназа ). .
Функция
[ редактировать ]Протеородопсин функционирует во всех океанах Земли как управляемый светом H+-насос по механизму, аналогичному механизму бактериородопсина. Как и в бактериородопсине, хромофор сетчатки протеородопсина ковалентно связан с апопротеином через протонированное основание Шиффа по адресу Lys231. Конфигурация хромофора сетчатки в нефотолизованном протеородопсине преимущественно полностью транс-трансгендерная. [ 18 ] и изомеризуется в 13-цис при освещении светом. Было предложено несколько моделей полного фотоцикла протеородопсина, основанных на FTIR и УФ-видимой спектроскопии ; они напоминают устоявшиеся модели фотоцикла бактериородопсина. [ 18 ] [ 20 ] [ 21 ] [ 22 ] Полные фотосистемы на основе протеородопсина были обнаружены и экспрессированы в E. coli, что придает им дополнительную способность опосредованного светом градиента энергии для генерации АТФ без внешней необходимости в сетчатке или предшественниках; вместе с геном PR пять других белков кодируют путь биосинтеза фотопигментов. [ 23 ]
Генная инженерия
[ редактировать ]Если ген протеородопсина вставить в E. coli дать ретиналь и этим модифицированным бактериям , то они встроят пигмент в свою клеточную мембрану и будут перекачивать H+ в присутствии света. Темно-фиолетовый цвет соответствует явно трансформированным колониям из-за поглощения света. Протонные градиенты можно использовать для питания других мембранных белковых структур или для подкисления органеллы типа везикул. [ 1 ] Далее было продемонстрировано, что протонный градиент, создаваемый протеородопсином, можно использовать для генерации АТФ. [ 23 ]
См. также
[ редактировать ]Галерея
[ редактировать ]-
 Голофермент (зеленый) со спиралями, помеченными AG (фиолетовый), а также лигандом сетчатки (оранжевый)
Голофермент (зеленый) со спиралями, помеченными AG (фиолетовый), а также лигандом сетчатки (оранжевый) -
 Визуализация поверхности протеородопсина с указанием терминалей
Визуализация поверхности протеородопсина с указанием терминалей -
 Визуализация связанного с сетчаткой активного сайта белковой структуры 2L6X pRhodopsin, остатки закодированы цветом и помечены по активности, лиганд окрашен в оранжевый цвет.
Визуализация связанного с сетчаткой активного сайта белковой структуры 2L6X pRhodopsin, остатки закодированы цветом и помечены по активности, лиганд окрашен в оранжевый цвет. -
 2L6x Визуализация с мультяшной цветовой кодировкой и маркировкой в активном сайте, спирали D и E, скрытые для обзора, сайт связывания лигандов в сетчатке
2L6x Визуализация с мультяшной цветовой кодировкой и маркировкой в активном сайте, спирали D и E, скрытые для обзора, сайт связывания лигандов в сетчатке




Ссылки
[ редактировать ]- ^ Jump up to: а б с д Бежа О, Аравинд Л, Кунин Е.В., Сузуки М.Т., Хадд А., Нгуен Л.П., Йованович С.Б., Гейтс К.М., Фельдман Р.А., Спудич Дж.Л., Спудич Е.Н., Делонг Э.Ф. (сентябрь 2000 г.). «Бактериальный родопсин: свидетельства нового типа фототрофии в море». Наука . 289 (5486): 1902–6. Бибкод : 2000Sci...289.1902B . дои : 10.1126/science.289.5486.1902 . ПМИД 10988064 .
- ^ Линь С., Чжан Х., Чжуан Ю., Тран Б., Гилл Дж. (ноябрь 2010 г.). «Метатранскриптомный анализ на основе сращивания лидеров приводит к распознаванию скрытых геномных особенностей динофлагеллят» . Труды Национальной академии наук Соединенных Штатов Америки . 107 (46): 2003–8. Бибкод : 2010PNAS..10720033L . дои : 10.1073/pnas.1007246107 . ПМЦ 2993343 . ПМИД 21041634 .
- ^ Jump up to: а б с Сламовиц Ч., Окамото Н., Бурри Л., Джеймс Э.Р., Килинг П.Дж. (2011). «Бактериальный протонный насос протеородопсина у морских эукариот» . Природные коммуникации . 2 (2): 183. Бибкод : 2011NatCo...2..183S . дои : 10.1038/ncomms1188 . ПМИД 21304512 .
- ^ Jump up to: а б с Фригаард Н.У., Мартинес А., Минсер Т.Дж., Делонг Э.Ф. (февраль 2006 г.). «Боковой перенос гена протеородопсина между морскими планктонными бактериями и архей». Природа . 439 (7078): 847–50. Бибкод : 2006Natur.439..847F . дои : 10.1038/nature04435 . ПМИД 16482157 . S2CID 4427548 .
- ^ Бур ЭД, Юэ В.В., Рен Х, Цзян З, Ляо Х.В., Мэй Икс, Вемараджу С., Нгуен М.Т., Рид Р.Р., Ланг Р.А., Яу К.В., Ван Гелдер Р.Н. (20 октября 2015 г.). " "Нейропсин (OPN5)-опосредованный фотоувлечение локальных циркадных осцилляторов в сетчатке и роговице млекопитающих" " . Труды Национальной академии наук . 112 (42): 13093–13098. Бибкод : 2015PNAS..11213093B . дои : 10.1073/pnas.1516259112 . ПМЦ 4620855 . ПМИД 26392540 .
- ^ Jump up to: а б Бежа О, Спудич Э.Н., Спудич Дж.Л., Леклерк М., Делонг Э.Ф. (июнь 2001 г.). «Фототрофия протеородопсина в океане». Природа . 411 (6839): 786–9. Бибкод : 2001Natur.411..786B . дои : 10.1038/35081051 . ПМИД 11459054 . S2CID 4428964 .
- ^ Ман Д., Ван В., Сабехи Г., Аравинд Л., Пост А.Ф., Массана Р., Спудич Э.Н., Спудич Дж.Л., Бежа О (апрель 2003 г.). «Диверсификация и спектральная настройка морских протеородопсинов» . Журнал ЭМБО . 22 (8): 1725–31. дои : 10.1093/emboj/cdg183 . ПМК 154475 . ПМИД 12682005 .
- ^ Келемен Б.Р., Ду М., Дженсен Р.Б. (декабрь 2003 г.). «Протеородопсин в живом цвете: разнообразие спектральных свойств живых бактериальных клеток» . Biochimica et Biophysical Acta (BBA) – Биомембраны . 1618 (1): 25–32. дои : 10.1016/j.bbamem.2003.10.002 . ПМИД 14643930 .
- ^ Jump up to: а б Сабехи Г, Киркуп БК, Розенберг М, Стамблер Н, Польц МФ, Бежа О (май 2007 г.). «Адаптация и спектральная настройка расходящихся морских протеородопсинов из восточного Средиземноморья и Саргассова морей» . Журнал ISME . 1 (1): 48–55. дои : 10.1038/ismej.2007.10 . ПМИД 18043613 .
- ^ Энциклопедия неврологических наук . Академическая пресса. 29 апреля 2014 г. с. 441. ИСБН 978-0-12-385158-1 .
- ^ Jump up to: а б Гизе, Артур С. (сентябрь 2013 г.). Фотофизиология: Общие принципы; Действие света на растения . Эльзевир. п. 9. ISBN 978-1-4832-6227-7 .
- ^ Вентер Дж.К., Ремингтон К., Гейдельберг Дж.Ф., Халперн А.Л., Раш Д., Эйзен Дж.А., Ву Д., Полсен И., Нельсон К.Е., Нельсон В., Фаутс Д.Е., Леви С., Кнап А.Х., Ломас М.В., Нилсон К., Уайт О., Петерсон Дж. , Хоффман Дж., Парсонс Р., Баден-Тилсон Х., Пфанкох С., Роджерс Ю.Х., Смит Х.О. (апрель 2004 г.). «Дробовиковое секвенирование экологического генома Саргассова моря». Наука . 304 (5667): 66–74. Бибкод : 2004Sci...304...66V . CiteSeerX 10.1.1.124.1840 . дои : 10.1126/science.1093857 . ПМИД 15001713 . S2CID 1454587 .
- ^ Джованнони, С.Дж.; Биббс, Л; Чо, Джей Си; Стейпелс, доктор медицины; Дезидерио, Р; Вергин, КЛ; Раппе, М.С.; Лэйни, С; Вильгельм, LJ; Трипп, HJ; Матур, Э.Дж.; Барофски, Д.Ф. (3 ноября 2005 г.). «Протеородопсин в вездесущей морской бактерии SAR11». Природа . 438 (7064): 82–5. Бибкод : 2005Natur.438...82G . дои : 10.1038/nature04032 . ПМИД 16267553 . S2CID 4414677 .
- ^ Кушваха, Южная Каролина; Кейтс, М. (23 августа 1973 г.). «Выделение и идентификация «бактериородопсина» и второстепенных C40-каротиноидов в Halobacterium Cutirubrum». Biochimica et Biophysical Acta (BBA) - Липиды и липидный обмен . 316 (2): 235–43. дои : 10.1016/0005-2760(73)90013-1 . ПМИД 4741911 .
- ^ Энциклопедия неврологических наук . Академическая пресса. Апрель 2014. с. 441. ИСБН 978-0-12-385158-1 .
- ^ Остерхельт, Д; Стокениус, W (29 сентября 1971 г.). «Родопсинподобный белок пурпурной мембраны Halobacterium halobium». Новая биология природы . 233 (39): 149–52. дои : 10.1038/newbio233149a0 . ПМИД 4940442 .
- ^ Jump up to: а б Мацуно-Яги, А; Мукохата, Ю. (9 сентября 1977 г.). «Две возможные роли бактериородопсина; сравнительное исследование штаммов Halobacterium halobium, различающихся пигментацией». Связь с биохимическими и биофизическими исследованиями . 78 (1): 237–43. дои : 10.1016/0006-291x(77)91245-1 . ПМИД 20882 .
- ^ Jump up to: а б с Диумаев А.К., Браун Л.С., Ши Дж., Спудич Е.Н., Спудич Дж.Л., Лани Дж.К. (апрель 2002 г.). «Перенос протона в фотохимическом цикле реакций протеородопсина». Биохимия . 41 (17): 5348–58. дои : 10.1021/bi025563x . ПМИД 11969395 .
- ^ Парта Р., Кребс Р., Катерино Т.Л., Брайман М.С. (июнь 2005 г.). «Ослабленное соединение консервативного аргинина с хромофором протеородопсина и его противоионом предполагает структурные отличия от бактериородопсина» . Biochimica et Biophysica Acta (BBA) — Биоэнергетика . 1708 (1): 6–12. дои : 10.1016/j.bbabio.2004.12.009 . ПМИД 15949979 .
- ^ Jump up to: а б Диумаев А.К., Ван Дж.М., Балинт З., Варо Г., Ланьи Дж.К. (июнь 2003 г.). «Транспорт протонов с помощью протеородопсина требует, чтобы противоион основания Шиффа Asp-97 сетчатки был анионным». Биохимия . 42 (21): 6582–7. дои : 10.1021/bi034253r . ПМИД 12767242 .
- ^ Jump up to: а б Кребс Р.А., Алексиев У., Парта Р., ДеВита А.М., Брайман М.С. (апрель 2002 г.). «Обнаружение быстрого активируемого светом высвобождения H + и образования промежуточного продукта M из протеородопсина» . Физиология БМК . 2 :5. дои : 10.1186/1472-6793-2-5 . ПМЦ 103662 . ПМИД 11943070 .
- ^ Сяо Ю., Парта Р., Кребс Р., Брайман М. (январь 2005 г.). «Фурье-спектроскопия с временным разрешением фотопромежуточных продуктов, участвующих в быстром кратковременном высвобождении H + протеородопсином». Журнал физической химии Б. 109 (1): 634–41. дои : 10.1021/jp046314g . ПМИД 16851056 .
- ^ Jump up to: а б Мартинес А., Брэдли А.С., Вальдбауэр-младший, Саммонс Р.Э., Делонг Э.Ф. (2007). «Экспрессия генов фотосистемы протеородопсина обеспечивает фотофосфорилирование у гетерологичного хозяина» . ПНАС . 104 (13): 5590–5595. Бибкод : 2007PNAS..104.5590M . дои : 10.1073/pnas.0611470104 . ПМЦ 1838496 . ПМИД 17372221 .
| Оптогенетические актуаторы | |
|---|---|
| Оптогенетические датчики | |
| Связанные методы | |