Кандида Pelagibacter communis
| " Кандидат Pelagibacter communis " | |
|---|---|
| |
| Научная классификация ( Кандидат ) | |
| Домен: | |
| Тип: | |
| Сорт: | |
| Подкласс: | |
| Заказ: | |
| Семья: | « Пелагибактеровые »
|
| Род: | « Кандидат Пелагибактер». Раппе и др . 2002 г.
|
| Разновидность: | " Кап . Обыкновенный"
|
| Биномиальное имя | |
| " Кандидат Pelagibacter communis " правильный. Раппе и др . 2002 г.
| |
| Синонимы [ 1 ] | |
| |
« Candidatus Pelagibacter» с единственным видом « Ca.P. communis» был выделен в 2002 году и получил особое название, [ 2 ] хотя он еще не описан в соответствии с требованиями бактериологического кодекса . [ 3 ] Это распространенный представитель клады SAR11 типа Alphaproteobacteria . Члены SAR11 являются высокодоминантными организмами, обитающими как в соленой, так и в пресной воде по всему миру, и первоначально были известны только по их генам рРНК , впервые выявленным в Саргассовом море в 1990 году лабораторией Стивена Джованнони в Университете штата Орегон , а затем обнаруженным в океанах по всему миру. [ 4 ] « Ca. P. communis» и его родственники могут быть самыми многочисленными организмами в океане и, вполне возможно, самыми многочисленными бактериями во всем мире. Они могут составлять около 25% всех клеток микробного планктона , а летом они могут составлять примерно половину клеток, присутствующих в поверхностных водах океана умеренного пояса. Общая численность Ca. P. communis и его родственников оценивается примерно в 2 × 10. 28 микробы. [ 5 ]
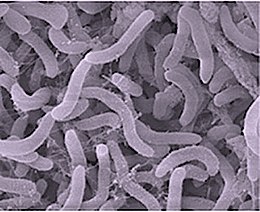
Это палочковидная или серповидная форма и одна из самых маленьких известных самовоспроизводящихся клеток: длина 0,37–0,89 мкм и диаметр всего 0,12–0,20 мкм. Геном Pelagibacter занимает около 30% объема клетки. [ 6 ] Это грамотрицательный . [ 7 ] Он перерабатывает растворенный органический углерод . Его численность претерпевает регулярные сезонные циклы - летом достигает ~ 50% клеток в поверхностных водах океана умеренного пояса. Земли Таким образом, он играет важную роль в углеродном цикле .
Его открытие стало предметом пятого эпизода «Океана микробов» канала PBS «Близкие незнакомцы: невидимая жизнь на Земле» . [ 8 ]
Выращивание
[ редактировать ]Несколько штаммов Candidatus Pelagibacter communis были выращены благодаря усовершенствованным методам выделения. [ 9 ] Наиболее изученным штаммом является HTCC1062 (коллекция высокопроизводительного культивирования). [ 2 ]
Факторы, регулирующие популяцию SAR11, до сих пор в значительной степени неизвестны. У них есть датчики ограничения азота , фосфатов и железа , а также очень необычная потребность в восстановленных соединениях серы . [ 10 ] Предполагается, что они сформировались в результате эволюции в экосистеме с низким содержанием питательных веществ, такой как Саргассово море, где он был впервые обнаружен. [ 11 ]
Популяция клеток « Ca.P. communis» может удваиваться каждые 29 часов, что довольно медленно, но они могут размножаться в условиях недостатка питательных веществ. [ 12 ]
« Ca. P. communis» можно выращивать на определенной искусственной среде с добавлением восстановленной серы, глицина, пирувата и витаминов. [ 13 ]
Геном
[ редактировать ]Геном штамма HTCC1062 « Ca.P. communis» был полностью секвенирован в 2005 году, показав, что « Ca.P. communis» имеет самый маленький геном (1 308 759 пар оснований) среди всех свободноживущих организмов. [ 6 ] кодирующий только 1354 открытых рамки считывания (всего 1389 генов). [ 14 ] Единственные виды с меньшими геномами — это симбионты и паразиты, такие как Mycoplasmagentium или Nanoarchaeum equitans. [ 6 ] Он имеет наименьшее количество открытых рамок считывания среди всех свободно живых организмов и самые короткие межгенные спейсеры, но у него все еще есть метаболические пути для всех 20 аминокислот и большинства кофакторов. [ 6 ] Его геном был оптимизирован . Эта концепция оптимизации важна, поскольку она уменьшает количество энергии, необходимой для репликации клеток. [ 7 ] « Ca. P. communis» экономит энергию, используя пары оснований A и T (≈70,3% всех пар оснований), поскольку они содержат меньше азота — ресурса, который организму трудно получить. [ 7 ]
Некодирующие РНК были идентифицированы у « Ca. P. communis» посредством биоинформационного анализа опубликованного генома и метагеномных данных. Примеры нкРНК, обнаруженные в этих организмах, включают рибопереключатель SAM-V и другие цис-регуляторные элементы, такие как мотив rpsB . [ 15 ] [ 16 ] Другим примером важной нкРНК у « Ca. P. communis» и других членов клады SAR11 является консервативный, глицин-активируемый рибопереключатель на малат-синтазе, предположительно приводящий к «функциональной ауксотрофии» глицина или его предшественников для достижения оптимального роста. [ 17 ]
Обнаружено, что у него есть гены протеородопсина , которые помогают питать опосредованные светом протонные насосы . Незначительные различия возникают в экспрессии последовательностей его кодонов , когда он подвергается воздействию света или темноты. Больше генов окислительного фосфорилирования экспрессируется в темноте. [ 18 ]
Имя
[ редактировать ]Название рода ( Pelagibacter ) происходит от латинского существительного среднего рода pelagus («море») в сочетании с суффиксом -bacter (палочка, бактерия), что означает «морская бактерия». Соединительная гласная — это «i», а не «o», поскольку первый термин — это латинское «pelagus», а не греческий оригинал πέλαγος (pelagos) (слово «pelagus» — греческое слово, используемое в латинской поэзии, это Существительное 2-го склонения с греческим неправильным именительным падежом множественного числа pelagē , а не pelagi , греческое слово является средним в 3-м склонении на -ος (мн. -η), не связанным с латинскими словами 2-го склонения на -us. [ 19 ] ). Название видового эпитета ( ubique ) — латинское наречие, означающее «везде»; виды со статусом Candidatus не опубликованы на законных основаниях, поэтому не обязательно должны быть грамматически правильными, например, наличие конкретных эпитетов должно быть прилагательными или существительными в приложении в именительном падеже или существительными в родительном падеже в соответствии с правилом 12c IBCN. [ 20 ]
Термин « Candidatus » используется для предлагаемых видов, по которым из-за отсутствия информации [ 21 ] не позволяет ему быть признанным видом согласно бактериологическому коду, [ 22 ] [ 23 ] например, хранение в двух общедоступных хранилищах ячеек или отсутствие анализа FAME , [ 24 ] [ 25 ] тогда как « Candidatus Pelagibacter communis» не находится в АТСС и ДСМЗ , анализ липидов и хинонов не проводился.
HTTC1062 является типовым штаммом вида « Ca.P. communis», который, в свою очередь, является типовым видом рода « Candidatus Pelagibacter». [ 2 ] который, в свою очередь, является типовым родом клады SAR11 или семейства Pelagibacteraceae . [ 26 ]
Бактериофаг
[ редактировать ]сообщил, В феврале 2013 года журнал Nature что был обнаружен бактериофаг HTVC010P , атакующий « Ca.P. communis», и «вероятно, это действительно самый распространенный организм на планете». [ 27 ] [ 28 ]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Орен А. (2017). «Призыв к лингвистической точности - в том числе и в отношении Candidatus таксонов » . Int J Syst Evol Microbiol . 67 (4): 1085–1094. дои : 10.1099/ijsem.0.001715 . ПМИД 27926819 .
- ^ Jump up to: а б с Майкл С. Раппе; Стефани А. Коннон; Кевин Л. Вергин; Стивен Дж. Джованнони (2002). «Культивирование повсеместно распространенной клады морского бактериопланктона SAR11». Природа . 418 (6898): 630–633. Бибкод : 2002Natur.418..630R . дои : 10.1038/nature00917 . ПМИД 12167859 . S2CID 4352877 .
- ^ Список видов-кандидатов в LPSN ; Парте, Эйдан К.; Сарда Карбасс, Хоаким; Мейер-Колтхофф, Ян П.; Реймер, Лоренц К.; Гёкер, Маркус (1 ноября 2020 г.). «Список названий прокариот, имеющих номенклатурное положение (ЛПСН), перемещается в ДСМЗ» . Международный журнал систематической и эволюционной микробиологии . 70 (11): 5607–5612. дои : 10.1099/ijsem.0.004332 .
- ^ Р.М. Моррис и др. (2002). «Клада SAR11 доминирует в сообществах бактериопланктона на поверхности океана». Природа . 420 (6917): 806–810. Бибкод : 2002Natur.420..806M . дои : 10.1038/nature01240 . ПМИД 12490947 . S2CID 4360530 .
- ^ « Candidatus Pelagibacter ubique». Европейский институт биоинформатики. Европейский институт биоинформатики, 2011. Интернет. 8 января 2012 г. http://www.ebi.ac.uk/2can/genomes/bacteria/Candidatus_Pelagibacter_ubique.html. Архивировано 1 декабря 2008 г. в Wayback Machine.
- ^ Jump up to: а б с д Стивен Дж. Джованнони, Х. Джеймс Трипп и др. (2005). «Оптимизация генома космополитической океанической бактерии». Наука . 309 (5738): 1242–1245. Бибкод : 2005Sci...309.1242G . дои : 10.1126/science.1114057 . ПМИД 16109880 . S2CID 16221415 .
- ^ Jump up to: а б с «Архивная копия» (PDF) . Архивировано (PDF) из оригинала 5 марта 2006 г. Проверено 2 февраля 2012 г.
{{cite web}}: CS1 maint: архивная копия в названии ( ссылка ) , Готье, Николас; Зинман, Гай; Д'Антонио, Маттео; Авраам, Майкл. Курс DTU «Сравнительная микробная геномика». 2005. - ^ Посмотреть «Океаны микробов» http://www.podcastdirectory.com/podshows/4339749. Архивировано 17 февраля 2012 г. на Wayback Machine.
- ^ Стингл, У.; Трипп, HJ; Джованнони, SJ (2007). «Усовершенствование высокопроизводительного культивирования привело к появлению новых штаммов SAR11 и других многочисленных морских бактерий с побережья Орегона и с участка исследования атлантических временных рядов на Бермудских островах» . Журнал ISME . 1 (4): 361–71. Бибкод : 2007ISMEJ...1..361S . дои : 10.1038/ismej.2007.49 . ПМИД 18043647 .
- ^ Х. Джеймс Трипп; Джошуа Б. Китнер; Майкл С. Швальбах; Джон В. Х. Дейси; и др. (апрель 2008 г.). «Морским бактериям SAR11 для роста требуется экзогенная восстановленная сера». Природа . 452 (7188): 741–4. Бибкод : 2008Natur.452..741T . дои : 10.1038/nature06776 . ПМИД 18337719 . S2CID 205212536 .
- ^ Лаборатория Джованнони http://giovannonilab.science.oregonstate.edu/. Архивировано 20 июля 2011 г. в Wayback Machine.
- ^ Джованнони Стивен Дж.; Стингл Ульрих (2005). «Молекулярное разнообразие и экология микробного планктона». Природа . 437 (7057): 343–348. Бибкод : 2005Natur.437..343G . дои : 10.1038/nature04158 . ПМИД 16163344 . S2CID 4349881 .
- ^ Карини, Пол; и др. (2012). «Потребности в питательных веществах для роста крайнего олиготрофа Candidatus Pelagibacter ubique» HTCC1062 на определенной среде» . Журнал ISME . 7 (3): 592–602. дои : 10.1038/ismej.2012.122 . ПМЦ 3578571 . ПМИД 23096402 .
- ^ «Геном Pelagibacter ubique» . НКБИ . Проверено 27 ноября 2012 г.
- ^ Мейер М.М., Эймс Т.Д., Смит Д.П. и др. (2009). «Идентификация кандидатных структурированных РНК в морском организме Candidatus Pelagibacter ubique » . БМК Геномика . 10 :268. дои : 10.1186/1471-2164-10-268 . ПМК 2704228 . ПМИД 19531245 .
- ^ Поята Э; Мейер ММ; Эймс Т.Д.; Брейкер RR (ноябрь 2009 г.). «Вариант класса аптамеров рибосвитча для S-аденозилметионина, распространенный у морских бактерий» . РНК . 15 (11): 2046–56. дои : 10.1261/rna.1824209 . ПМЦ 2764483 . ПМИД 19776155 .
- ^ Х. Джеймс Трипп; Майкл С. Швальбах; Мишель М. Мейер; Джошуа Б. Китнер; и др. (январь 2009 г.). «Уникальный глицин-активируемый рибопереключатель, связанный с глицин-сериновой ауксотрофией при SAR11» . Экологическая микробиология . 11 (1): 230–8. Бибкод : 2009EnvMi..11..230T . дои : 10.1111/j.1462-2920.2008.01758.x . ПМК 2621071 . ПМИД 19125817 .
- ^ Штайндлер Лаура; Швальбах Майкл С.; Смит Дэниел П.; Чан Фрэнсис; и др. (2011). «Энергетически голодный» Candidatus Pelagibacter ubique заменяет эндогенное углеродное дыхание на производство АТФ, опосредованное светом» . ПЛОС ОДИН . 6 (5): 9999. Бибкод : 2011PLoSO...619725S . дои : 10.1371/journal.pone.0019725 . ПМК 3090418 . ПМИД 21573025 .
- ^ Грегори Р. Крейн. «запись о пелаге в цифровой библиотеке Персея» . Проект цифровой библиотеки «Персей» . Университет Тафтса . Проверено 22 мая 2011 г.
- ^ Лапаж, С.; Снит, П.; Лессель, Э.; Скерман, В.; Силигер, Х.; Кларк, В. (1992). Международный кодекс номенклатуры бактерий: Бактериологический кодекс, редакция 1990 г. Вашингтон, округ Колумбия: ASM Press. ПМИД 21089234 .
- ^ «Минимальные стандарты описания новых таксонов» . Архивировано из оригинала 27 января 2013 г. Проверено 15 декабря 2010 г.
- ^ Мюррей, RGE; Шлейфер, К.Х. (1994). «Таксономические заметки: предложение по регистрации свойств предполагаемых таксонов прокариотов» . Международный журнал систематической и эволюционной микробиологии . 44 (1): 174–176. дои : 10.1099/00207713-44-1-174 . ПМИД 8123559 .
- ^ Судебная комиссия Международного комитета по систематической бактериологии: протоколы заседаний, 2 и 6 июля 1994 г., Прага, Чехия». Международный журнал систематической и эволюционной микробиологии 1995; 45, 195–196.
- ^ Эузеби, JP (2010). "Введение" . Список названий прокариот, стоящих в номенклатуре . Архивировано из оригинала 06 марта 2011 г. Проверено 16 декабря 2010 г.
- ^ Снит, PH A (1992). Лапаж, СП; Снит, PHA; Лессель, Э.Ф.; Скерман, ВБД; Силигер, HPR; Кларк, Вашингтон (ред.). Международный кодекс номенклатуры бактерий . Вашингтон, округ Колумбия: Американское общество микробиологии. ISBN 978-1-55581-039-9 . ПМИД 21089234 .
- ^ Трэш, Джей Си; Бойд, А.; Хаггетт, MJ; Гроте, Дж.; Карини, П.; Йодер, Р.Дж.; Робберце, Б.; Спатафора, JW; Раппе, М.С.; Джованнони, SJ (2011). «Филогеномные доказательства общего предка митохондрий и клады SAR11» . Научные отчеты . 1 : 13. Бибкод : 2011НатСР...1Е..13Т . дои : 10.1038/srep00013 . ПМК 3216501 . ПМИД 22355532 .
- ^ «Блошиный рынок: недавно обнаруженный вирус может оказаться самым распространенным организмом на планете» . Экономист . 16 февраля 2013 года . Проверено 16 февраля 2013 г.
- ^ Чжао, Ю.; Темпертон, Б.; Трэш, Джей Си; Швальбах, М.С.; Вергин, КЛ; Лэндри, ЗК; Эллисман, М.; Диринк, Т.; Салливан, МБ; Джованнони, SJ (2013). «Обилие вирусов SAR11 в океане». Природа . 494 (7437): 357–360. Бибкод : 2013Natur.494..357Z . дои : 10.1038/nature11921 . ПМИД 23407494 . S2CID 4348619 .
