Желатиновый зоопланктон

Желатиновый зоопланктон – хрупкие животные, обитающие в толще воды океана. Их хрупкие тела не имеют твердых частей и легко повреждаются или разрушаются. [2] Желатиновый зоопланктон часто прозрачен. [3] Все медузы представляют собой студенистый зоопланктон, но не весь студенистый зоопланктон является медузой. Наиболее часто встречающиеся организмы включают гребневики , медузы , сальпы и Chaetognatha в прибрежных водах . Однако почти все морские типы, включая Annelida , Mollusca и Arthropoda , содержат студенистые виды, но многие из этих странных видов обитают в открытом океане и глубоководных водах и менее доступны для случайного наблюдателя за океаном. [4] Многие студенистые планктеры используют слизистые структуры для фильтрации корма. [5] Желатиновый зоопланктон также называют гелатой . [6]
Как добыча
[ редактировать ]| Часть серии о |
| Планктон |
|---|
 |
Медузы — медленные пловцы, и большинство видов составляют часть планктона. Традиционно медузы рассматривались как трофические тупики, второстепенные участники морской пищевой цепи, студенистые организмы, строение тела которых в основном основано на воде, которая не представляет особой питательной ценности или интереса для других организмов, за исключением нескольких специализированных хищников, таких как океанская солнечная рыба и кожистая морская черепаха . [7] [1] Недавно эта точка зрения была оспорена. Медузы и студенистый зоопланктон в целом, в который входят сальпы и гребневики , очень разнообразны, хрупкие, без твердых частей, их трудно увидеть и контролировать, они подвержены быстрым колебаниям численности и часто живут неудобно далеко от берега или глубоко в океане. Ученым сложно обнаружить и проанализировать медуз в кишках хищников, поскольку при поедании они превращаются в кашу и быстро перевариваются. [7] Но медузы цветут в огромных количествах, и было доказано, что они составляют основные компоненты рациона тунца , морской рыбы и рыбы-меч , а также различных птиц и беспозвоночных, таких как осьминоги , морские огурцы , крабы и амфиподы . [8] [1] «Несмотря на низкую энергетическую плотность, вклад медуз в энергетический бюджет хищников может быть намного больше, чем предполагалось, из-за быстрого пищеварения, низких затрат на отлов, доступности и избирательного питания более богатыми энергией компонентами. морские хищники, восприимчивые к проглатыванию пластика». [1]
Как хищники
[ редактировать ]Согласно исследованию 2017 года, наркомедузы потребляют наибольшее разнообразие мезопелагических жертв, за ними следуют физонектовые сифонофоры , гребневики и головоногие моллюски . [9] Важность так называемой «желейной паутины» только начинает пониматься, но кажется, что медузы, гребневики и сифонофоры могут быть ключевыми хищниками в глубоких пелагических пищевых цепях с экологическим воздействием, аналогичным воздействию хищных рыб и кальмаров. Традиционно студенистые хищники считались неэффективными поставщиками морских трофических путей, но, по-видимому, они играют существенную и неотъемлемую роль в глубоких пелагических пищевых сетях. [9]
Пелагические сифонофоры , разнообразная группа книдарий, встречаются на большинстве глубин океана — от поверхности, как португальский военный корабль , до морских глубин. Они играют важную роль в экосистемах океана и являются одними из самых многочисленных студенистых хищников. [10]


 Желатиновый зоопланктон, такой как этот наркомедузан, может быть ключевым хищником в глубоких пелагических пищевых сетях.
Желатиновый зоопланктон, такой как этот наркомедузан, может быть ключевым хищником в глубоких пелагических пищевых сетях. Солмиссус проглатывает сальповую цепь
Солмиссус проглатывает сальповую цепь


 Шлемовая медуза питается кальмаром с крючком
Шлемовая медуза питается кальмаром с крючком Трахимедуза с большим красным мизидом в кишечнике.
Трахимедуза с большим красным мизидом в кишечнике.


Желе насос
[ редактировать ]
Биологические океанические процессы, в первую очередь производство углерода в эвфотической зоне , погружение и реминерализация , управляют глобальным биологическим углеродным насосом мягких тканей. [12] Опускающиеся и латерально переносимые частицы углерода питают донные экосистемы на окраинах континентов и в глубоком море. [13] [14] Морской зоопланктон играет важную роль в качестве инженера экосистем в прибрежных и открытых океанских экосистемах, поскольку он служит связующим звеном между первичной продукцией, более высокими трофическими уровнями и глубоководными сообществами. [15] [14] [16] В частности, студенистый зоопланктон (Cnidaria, Ctenophora и Chordata, а именно Thaliacea) являются универсальными членами планктонных сообществ, питающихся фитопланктоном и питающихся другим зоопланктоном и ихтиопланктоном. Они также могут быстро размножаться в течение нескольких дней, а при благоприятных условиях окружающей среды некоторые виды образуют густое цветение, простирающееся на многие квадратные километры. [17] Это цветение имеет негативные экологические и социально-экономические последствия, сокращая количество промысловых видов рыбы. [18] ограничение переноса углерода на другие трофические уровни, [19] усиление микробной реминерализации и тем самым снижение концентрации кислорода близко к бескислородному уровню. [20] [11]
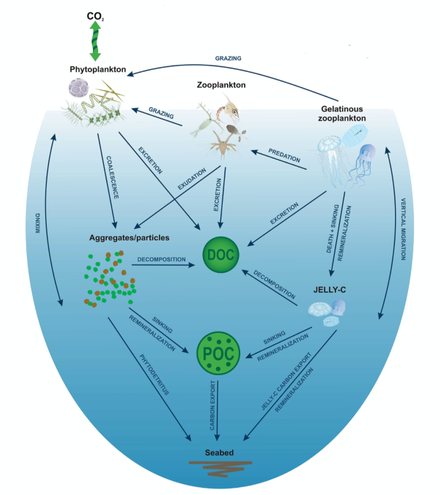
Желе углерода
[ редактировать ]Глобальная биомасса студенистого зоопланктона (иногда называемого желе-С ) в верхних 200 м океана составляет 0,038 Пг С. [21] Расчеты для мезозоопланктона (от 200 мкм до 2 см) предполагают около 0,20 Пг C. [22] Короткая продолжительность жизни большинства студенистого зоопланктона – от недель до 2–12 месяцев. [23] [24] предполагает темпы производства биомассы выше 0,038 Пг C в год. −1 , в зависимости от предполагаемых показателей смертности, которые во многих случаях являются видоспецифичными. Это намного меньше, чем мировое первичное производство (50 Пг C в год). −1 ), [25] что соответствует оценкам экспорта, близким к 6 Пг C в год. −1 ниже 100 м, [26] [27] в зависимости от используемого метода. Во всем мире численность и распределение студенистого зоопланктона во многом зависят от температуры и растворенного кислорода, а также от первичной продукции в качестве источника углерода. [21] Однако студенистый зоопланктон справляется с широким спектром условий окружающей среды, что указывает на способность адаптироваться и занимать наиболее доступные экологические ниши в водной массе. Что касается регионов Лонгхерста ( биогеографических провинций, разделяющих пелагическую среду, [28] [29] наибольшая плотность студенистого зоопланктона наблюдается в прибрежных водах течения Гумбольдта , шельфа северо-северо-востока США, шельфов Шотландии и Ньюфаундленда, Бенгельского течения , Восточно-Китайского и Желтого морей, за которыми следуют полярные районы Восточно-Берингова и Охотского морей, Южного океана, вкл. водоемы, такие как Черное море и Средиземное море, а также западно-тихоокеанские воды Японского моря и течения Куросио . [30] [31] [21] О больших количествах желеобразной углеродной биомассы, зарегистрированной в прибрежных районах открытых шельфов и полузамкнутых морей Северной Америки, Европы и Восточной Азии, свидетельствуют данные о выбросах на берег. [32] [11]
Экспорт углерода
[ редактировать ]Большие количества желеобразного углерода быстро переносятся и реминерализуются на морском дне в прибрежных районах, включая эстуарии, лагуны и сублиторальные/литоральные зоны. [15] полки и откосы, [33] [34] [35] глубоководье. [36] и даже целые континентальные окраины, например, в Средиземном море. [37] Перенос углерода в желе начинается, когда студенистый зоопланктон умирает на заданной «глубине смерти» (глубине выхода), продолжается, когда биомасса опускается через толщу воды, и прекращается, когда биомасса реминерализируется во время погружения или достигает морского дна, а затем разлагается. Желеобразный углерод по сути представляет собой перенос «уже вывезенных» частиц (ниже смешанной поздней, эвфотической или мезопелагической зоны), возникших в первичной продукции, поскольку студенистый зоопланктон «переупаковывает» и интегрирует этот углерод в свои тела, а после гибели переносит его в недра океана. При погружении в толщу воды желеобразный углерод частично или полностью реминерализируется в виде растворенного органического/неорганического углерода и питательных веществ ( DOC , DIC , DON, DOP, DIN и DIP). [38] [39] [40] а любые остатки далее подвергаются микробному разложению или уничтожаются макрофауной и мегафауной , оказавшись на морском дне. [41] [42] Несмотря на высокую лабильность желе-С, [43] [39] удивительно большое количество биомассы попадает на морское дно ниже 1000 м. Во время погружения биохимический состав желе-C меняется за счет изменения соотношения C:N:P, как наблюдалось в экспериментальных исследованиях. [20] [44] [45] Тем не менее, реалистичные оценки переноса желе-C в глобальном масштабе остаются в зачаточном состоянии, что не позволяет количественно оценить вклад биологического углеродного насоса в мягкие ткани. [11]

Экспорт углерода из океана обычно оценивается по потоку тонущих частиц, которые либо улавливаются в отстойники, либо попадают в отстойники. [47] или количественно измерено на основе видеосъемки, [48] и впоследствии смоделировано с использованием скорости погружения. [49] Биогеохимические модели [50] [51] [52] обычно параметризуются с использованием данных по твердым органическим веществам (например, морской снег размером 0,5–1000 мкм и фекальные шарики), которые были получены в результате лабораторных экспериментов. [53] или на основе данных осадочных ловушек. [50] Эти модели не включают желе-C (кроме личинок, [54] [55] не только потому, что этот механизм переноса углерода считается временным/эпизодическим и обычно не наблюдается, а потоки массы слишком велики, чтобы их можно было собрать отстойниками, [27] но также и потому, что модели направлены на упрощение биотических компартментов для облегчения расчетов. Более того, отложения желе-C, как правило, не накапливаются на морском дне в течение длительного времени, например, фитодетрит (Beaulieu, 2002), быстро потребляясь демерсальными и бентосными организмами. [41] или разлагается микробами. [42] Скорость погружения желе-C зависит от размера, диаметра, биообъема, геометрии организма. [56] плотность, [57] и коэффициенты сопротивления. [58] В 2013 году Лебрато и др. определили среднюю скорость погружения желе-С с использованием образцов Cnidaria, Ctenophora и Thaliacea, которая колебалась от 800 до 1500 м/день (сальпы: 800–1200 м/день; сцифозы: 1000–1100 мд-1; гребневики: 1200–1500 м/день-1; пиросомы: 1300 м/день-1); [59] Моделирование модели Jelly-C показывает, что, независимо от таксонов, более высокие широты являются более эффективными коридорами для переноса желе-C на морское дно из-за более низких показателей реминерализации. [60] В субтропических и умеренных регионах значительное разложение происходит в толще воды на глубине более 1500 м, за исключением случаев, когда желе-C начинает опускаться ниже термоклина. В мелководных прибрежных регионах время является ограничивающим фактором, который предотвращает реминерализацию при погружении и приводит к накоплению разлагающегося желе-C из различных таксонов на морском дне. Это говорит о том, что студенистый зоопланктон переносит большую часть биомассы и углерода в глубины океана, увеличивая прибрежные потоки углерода через DOC и DIC, подпитывая микробные и мегафауны/макрофауны, питающиеся мусорщиками. Однако отсутствие спутниковых измерений желе-C (таких как первичное производство ) [61] а ограниченное количество глобальных наборов данных о биомассе зоопланктона затрудняет количественную оценку глобального производства желе-C и эффективности его переноса в глубины океана. [11]
Мониторинг
[ редактировать ]Из-за его хрупкой структуры получение изображений студенистого зоопланктона требует помощи компьютерного зрения . Автоматическое распознавание зоопланктона в образцах отложений возможно за счет использования таких технологий, как регуляризация Тихонова , машины опорных векторов и генетическое программирование . [62]

 Темно-красная медуза — гидрозойное животное, обитающее в Северном Ледовитом океане на глубине ниже 1000 м (3300 футов). [63]
Темно-красная медуза — гидрозойное животное, обитающее в Северном Ледовитом океане на глубине ниже 1000 м (3300 футов). [63] Сальпа . , еще один пример студенистых оболочников, часто встречается в виде колониальной цепочки
Сальпа . , еще один пример студенистых оболочников, часто встречается в виде колониальной цепочки


См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Перейти обратно: а б с д Хейс, Грэм С .; Дойл, Томас К.; Хоутон, Джонатан Д.Р. (2018). «Смена парадигмы трофического значения медуз?» . Тенденции в экологии и эволюции . 33 (11): 874–884. дои : 10.1016/j.tree.2018.09.001 . ПМИД 30245075 . S2CID 52336522 .
- ^ Лалли, К.М. и Парсонс, Т.Р. (2001) Биологическая океанография . Баттерворт-Хайнеманн.
- ^ Джонсен, С. (2000) Прозрачные животные. Scientific American 282 : 62–71.
- ^ Нувиан, К. (2007) Глубина . Издательство Чикагского университета.
- ^ Хамнер, В.М.; Мадин, LP; Олдридж, Алабама; Гилмер, RW; Хамнер, П.П. (1975). «Подводные наблюдения за студенистым зоопланктоном: проблемы отбора проб, биология питания и поведение1» . Лимнология и океанография . 20 (6): 907–917. Бибкод : 1975LimOc..20..907H . дои : 10.4319/lo.1975.20.6.0907 . ISSN 1939-5590 .
- ^ ХЭДДОК, SDH (2004) Золотой век гелаты: прошлые и будущие исследования планктонных гребневиков и книдарий. Гидробиология 530/531 : 549–556.
- ^ Перейти обратно: а б Гамильтон, Г. (2016) «Тайная жизнь медуз: медузы, долгое время считавшиеся второстепенными игроками в экологии океана, на самом деле являются важной частью морской пищевой сети» . Природа , 531 (7595): 432–435. дои : 10.1038/531432a
- ^ Кардона, Л., Де Кеведо, И.А., Боррелл, А. и Агилар, А. (2012) «Массовое потребление студенистого планктона высшими хищниками Средиземноморья». ПЛОС ОДИН , 7 (3): e31329. дои : 10.1371/journal.pone.0031329
- ^ Перейти обратно: а б Чой, Калифорния, Хэддок, С.Х. и Робисон, Б.Х. (2017) «Структура глубокой пелагической пищевой сети, выявленная наблюдениями за питанием in situ ». Труды Королевского общества B: Биологические науки , 284 (1868): 2017–2116. дои : 10.1098/rspb.2017.2116 .
 Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 . - ^ Манро, Катриона; Зиберт, Стефан; Сапата, Фелипе; Хоуисон, Марк; Дамиан-Серрано, Алехандро; Черч, Сэмюэл Х.; Гетц, Фрейя Э.; Пью, Филип Р.; Хэддок, Стивен HD; Данн, Кейси В. (2018). «Улучшение филогенетического разрешения сифонофоры (Cnidaria) с последствиями для эволюции признаков» . Молекулярная филогенетика и эволюция . 127 : 823–833. дои : 10.1016/j.ympev.2018.06.030 . ПМК 6064665 . ПМИД 29940256 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Перейти обратно: а б с д и ж Лебрато, Марио; Пахлоу, Маркус; Фрост, Джессика Р.; Кютер, Мари; Хесус Мендес, Педро; Молинеро, Хуан-Карлос; Ошлис, Андреас (2019). «Затопление студенистой биомассы зоопланктона повышает эффективность глубокого переноса углерода во всем мире» . Глобальные биогеохимические циклы . 33 (12): 1764–1783. Бибкод : 2019GBioC..33.1764L . дои : 10.1029/2019GB006265 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Бюсселер, КО; Ламборг, Швейцария; Бойд, П.В.; Лам, ПиДжей; Трулл, ТВ; Бидигаре, Р.Р.; Бишоп, JKB; Кашотти, КЛ; Дехайрс, Ф.; Элскенс, М.; Хонда, М.; Карл, DM; Сигел, Д.А.; Серебро, МВт; Стейнберг, ДК; Вальдес, Дж.; Ван Мой, Б .; Уилсон, С. (2007). «Возвращение к потоку углерода через сумеречную зону океана». Наука . 316 (5824): 567–570. Бибкод : 2007Sci...316..567B . дои : 10.1126/science.1137959 . ПМИД 17463282 . S2CID 8423647 .
- ^ Коппельманн Р. и Фрост Дж. (2008). «Экологическая роль зоопланктона в сумеречных и темных зонах океана». В: Л. П. Мертенс (ред.), Тенденции биологических океанографических исследований , стр. 67–130. Нью-Йорк: Издательство Nova Science.
- ^ Перейти обратно: а б Робинсон, Кэрол; Стейнберг, Дебора К.; Андерсон, Томас Р.; Аристеги, Хавьер; Карлсон, Крейг А.; Фрост, Джессика Р.; Гильоне, Жан-Франсуа; Эрнандес-Леон, Сантьяго; Джексон, Джордж А.; Коппельманн, Рольф; Кегинер, Бернар; Рагено, Оливье; Рассулзадеган, Ферейдун; Робисон, Брюс Х.; Тамбурини, Кристиан; Танака, Цунео; Вишнер, Карен Ф.; Чжан, Цзин (2010). «Экология и биогеохимия мезопелагической зоны – синтез». Глубоководные исследования, часть II: Актуальные исследования в океанографии . 57 (16): 1504–1518. Бибкод : 2010DSRII..57.1504R . дои : 10.1016/j.dsr2.2010.02.018 .
- ^ Перейти обратно: а б Лебрато, Марио; Питт, Кайли А.; Свитман, Эндрю К.; Джонс, Дэниел О.Б.; Картес, Джоан Э.; Ошлис, Андреас; Кондон, Роберт Х.; Молинеро, Хуан Карлос; Адлер, Летиция; Гайяр, Кристиан; Льорис, Доминго; Биллетт, Дэвид С.М. (2012). «Исторические и недавние наблюдения о желе-падении: обзор, который определит будущие направления исследований». Гидробиология . 690 : 227–245. дои : 10.1007/s10750-012-1046-8 . S2CID 15428213 .
- ^ Робисон, Британская Колумбия; Райзенбихлер, КР; Шерлок, RE (2005). «Гигантские дома-личинки: быстрый перенос углерода на глубоководное дно». Наука . 308 (5728): 1609–1611. Бибкод : 2005Sci...308.1609R . дои : 10.1126/science.1109104 . ПМИД 15947183 . S2CID 730130 .
- ^ Кондон, Р.Х.; Дуарте, CM; Питт, Калифорния; Робинсон, КЛ; Лукас, Швейцария; Сазерленд, КР; Мианзан, HW; Богеберг, М.; Перселл, Дж. Э.; Декер, МБ; Уе, С.-и.; Мадин, LP; Бродер, Р.Д.; Хэддок, SHD; Малей, А.; Пэрри, Грузия; Эриксен, Э.; Хиноны, Дж.; Ача, М.; Харви, М.; Артур, Дж. М.; Грэм, WM (2013). «Периодическое цветение медуз является следствием глобальных колебаний» . Труды Национальной академии наук . 110 (3): 1000–1005. Бибкод : 2013PNAS..110.1000C . дои : 10.1073/pnas.1210920110 . ПМК 3549082 . ПМИД 23277544 .
- ^ Поли, Дэниел; Грэм, Уильям; Либралато, Симона; Мориссетт, Лайн; Дэн Паломарес, ML (2009). «Медузы в экосистемах, онлайн-базах данных и моделях экосистем». Гидробиология . 616 : 67–85. дои : 10.1007/s10750-008-9583-x . S2CID 12415790 .
- ^ Кондон, Р.Х.; Стейнберг, ДК; Дель Джорджио, Пенсильвания; Бувье, TC; Бронк, округ Колумбия; Грэм, WM; Даклоу, HW (2011). «Цветение медуз приводит к крупному микробному дыхательному стоку углерода в морских системах» . Труды Национальной академии наук . 108 (25): 10225–10230. Бибкод : 2011PNAS..10810225C . дои : 10.1073/pnas.1015782108 . ПМК 3121804 . ПМИД 21646531 .
- ^ Перейти обратно: а б Фрост, Джессика Р.; Джейкоби, Чарльз А.; Фрейзер, Томас К .; Циммерман, Эндрю Р. (2012). «Пульсовые нарушения из-за бактериального разложения Chrysaora quinquecirrha (Scyphozoa: Pelagiidae)». Гидробиология . 690 : 247–256. дои : 10.1007/s10750-012-1042-z . S2CID 17113316 .
- ^ Перейти обратно: а б с Лукас, Кэти Х.; Джонс, Дэниел О.Б.; Холлихед, Кэтрин Дж.; Кондон, Роберт Х.; Дуарте, Карлос М.; Грэм, Уильям М.; Робинсон, Келли Л.; Питт, Кайли А.; Шильдхауэр, Марк; Регетц, Джим (2014). «Биомасса студенистого зоопланктона в мировом океане: географические изменения и экологические факторы» (PDF) . Глобальная экология и биогеография . 23 (7): 701–714. дои : 10.1111/geb.12169 .
- ^ Мориарти, Р.; О'Брайен, ТД (2013). «Распределение биомассы мезозоопланктона в Мировом океане» . Данные науки о системе Земли . 5 (1): 45–55. Бибкод : 2013ESSD....5...45M . дои : 10.5194/essd-5-45-2013 .
- ^ Чех, Джанья; Гонсалес, Хорхе; Пачеко, Альдо С.; Риаскос, Хосе М. (2015). «Неуловимый жизненный цикл сцифозных медуз – новый взгляд на метагенез» . Научные отчеты . 5 : 12037. Бибкод : 2015NatSR...512037C . дои : 10.1038/srep12037 . ПМЦ 4495463 . ПМИД 26153534 .
- ^ Раскофф, Кевин А.; Соммер, Фрейя А.; Хамнер, Уильям М.; Кросс, Катрина М. (2003). «Методы сбора и культивирования студенистого зоопланктона» . Биологический бюллетень . 204 (1): 68–80. дои : 10.2307/1543497 . JSTOR 1543497 . ПМИД 12588746 . S2CID 22389317 .
- ^ Поле, CB; Беренфельд, МЮ; Рандерсон, Джей Ти; Фальковски, П. (1998). «Первичная продукция биосферы: интеграция наземных и океанических компонентов» . Наука . 281 (5374): 237–240. Бибкод : 1998Sci...281..237F . дои : 10.1126/science.281.5374.237 . ПМИД 9657713 .
- ^ Мур, Дж. Кейт; Дони, Скотт С .; Линдси, Кейт (2004). «Динамика экосистемы верхних слоев океана и круговорот железа в глобальной трехмерной модели». Глобальные биогеохимические циклы . 18 (4): н/д. Бибкод : 2004GBioC..18.4028M . дои : 10.1029/2004GB002220 . hdl : 1912/3396 . S2CID 3575218 .
- ^ Перейти обратно: а б Сигел, Д.А.; Бюсселер, КО; Дони, Южная Каролина ; Сайли, Сан-Франциско; Беренфельд, МЮ; Бойд, PW (2014). «Глобальная оценка экспорта углерода из океана путем объединения спутниковых наблюдений и моделей пищевой сети». Глобальные биогеохимические циклы . 28 (3): 181–196. Бибкод : 2014GBioC..28..181S . дои : 10.1002/2013GB004743 . hdl : 1912/6668 . S2CID 43799803 .
- ^ Лонгхерст, Алан (1995). «Сезонные циклы пелагического производства и потребления». Прогресс в океанографии . 36 (2): 77–167. Бибкод : 1995Proce..36...77L . дои : 10.1016/0079-6611(95)00015-1 .
- ^ Лонгхерст, Арканзас (1998). Экологическая география моря , стр. 398. Академическое издательство.
- ^ Бротц, Лукас; Чунг, Уильям В.Л.; Кляйснер, Кристин; Пахомов, Евгений; Поли, Дэниел (2012). «Увеличение популяции медуз: тенденции в крупных морских экосистемах» . Гидробиология . 690 : 3–20. дои : 10.1007/s10750-012-1039-7 . S2CID 12375717 .
- ^ Кондон, Роберт Х.; Грэм, Уильям М.; Дуарте, Карлос М.; Питт, Кайли А.; Лукас, Кэти Х.; Хэддок, Стивен HD; Сазерленд, Келли Р.; Робинсон, Келли Л.; Доусон, Майкл Н.; Декер, Мэри Бет; Миллс, Клаудия Э.; Перселл, Дженнифер Э.; Малей, Аленка; Мианзан, Гермес; Уе, Син-Ичи; Гельчих, Стефан; Мадин, Лоуренс П. (2012). «Под вопросом рост студенистого зоопланктона в Мировом океане» . Бионаука . 62 (2): 160–169. дои : 10.1525/bio.2012.62.2.9 . S2CID 2721425 .
- ^ Домашняя страница JellyWatch jellywatch.org . По состоянию на 17 апреля 2022 г.
- ^ Биллетт, DSM; Бетт, Би Джей; Джейкобс, CL; Роуз, ИП; Уигэм, Б.Д. (2006). «Массовое осаждение медуз в глубоком Аравийском море» . Лимнология и океанография . 51 (5): 2077–2083. Бибкод : 2006LimOc..51.2077B . дои : 10.4319/lo.2006.51.5.2077 .
- ^ Лебрато, М.; Джонс, год рождения (2009). «Массовое осаждение туш Pyrosoma atlanticum у Берега Слоновой Кости (Западная Африка)» . Лимнология и океанография . 54 (4): 1197–1209. Бибкод : 2009LimOc..54.1197L . дои : 10.4319/lo.2009.54.4.1197 .
- ^ Свитман, Эндрю К.; Чепмен, Аннелиза (2011). «Первые наблюдения желе-падений на морском дне глубоководного фьорда». Глубоководные исследования. Часть I: Статьи океанографических исследований . 58 (12): 1206–1211. Бибкод : 2011DSRI...58.1206S . дои : 10.1016/j.dsr.2011.08.006 .
- ^ Смит, К.Л. младший; Шерман, AD; Хаффард, CL; МакГилл, PR; Хентхорн, Р.; фон Тун, С.; Рул, штат Ха; Кару, М.; Оман, доктор медицины (2014). «Крупный экспорт сальпового цветения из верхних слоев океана и реакция донных сообществ в абиссальной северо-восточной части Тихого океана: решение по дням и неделям» . Лимнология и океанография . 59 (3): 745–757. Бибкод : 2014LimOc..59..745S . дои : 10.4319/lo.2014.59.3.0745 .
- ^ Лебрато, Марио; Молинеро, Хуан-Карлос; Картес, Джоан Э.; Льорис, Доминго; Мелен, Фредерик; Бени-Касаделла, Лайя (2013). «Тонущий желеобразный углерод раскрывает потенциальную изменчивость окружающей среды вдоль континентальной окраины» . ПЛОС ОДИН . 8 (12): е82070. Бибкод : 2013PLoSO...882070L . дои : 10.1371/journal.pone.0082070 . ПМЦ 3867349 . ПМИД 24367499 .
- ^ Челский, Ариэлла; Питт, Кайли А.; Валлийский, Дэвид Т. (2015). «Биогеохимические последствия разложения цветения медуз в меняющемся климате». Устьевые, прибрежные и шельфовые науки . 154 : 77–83. Бибкод : 2015ECSS..154...77C . дои : 10.1016/j.ecss.2014.12.022 . hdl : 10072/101243 .
- ^ Перейти обратно: а б Свитман, Эндрю К.; Челский, Ариэлла; Питт, Кайли А.; Андраде, Гектор; Ван Овелен, Дик; Рено, Поль Э. (2016). «Разложение медуз на морском дне быстро меняет биогеохимический цикл и поток углерода через донные пищевые сети» . Лимнология и океанография . 61 (4): 1449–1461. Бибкод : 2016LimOc..61.1449S . дои : 10.1002/lno.10310 .
- ^ Уэст, Элизабет Джейн; Уэлш, Дэвид Томас; Питт, Кайли Энн (2009). «Влияние разлагающихся медуз на потребность отложений в кислороде и динамику питательных веществ». Гидробиология . 616 : 151–160. дои : 10.1007/s10750-008-9586-7 . S2CID 46695384 .
- ^ Перейти обратно: а б Свитман, Эндрю К.; Смит, Крейг Р.; Дейл, Трина; Джонс, Дэниел О.Б. (2014). «Быстрое очищение туш медуз показывает важность студенистого материала для глубоководных пищевых сетей» . Труды Королевского общества B: Биологические науки . 281 (1796). дои : 10.1098/rspb.2014.2210 . ПМЦ 4213659 . ПМИД 25320167 .
- ^ Перейти обратно: а б Тинта, Тинкара; Коговшек, Тяша; Турок, Валентина; Шиганова Тамара А.; Микаэлян Александр Сергеевич; Малей, Аленка (2016). «Микробная трансформация органического вещества медуз влияет на круговорот азота в толще морской воды на примере Черного моря». Журнал экспериментальной морской биологии и экологии . 475 : 19–30. дои : 10.1016/j.jembe.2015.10.018 .
- ^ Атес, Рон М.Л. (2017). «Бентосные падальщики и хищники медуз, материал для обзора» . Исследования планктона и бентоса . 12 : 71–77. дои : 10.3800/pbr.12.71 .
- ^ Семпере, Р.; Йоро, Южная Каролина; Ван Вамбеке, Ф.; Шарьер, Б. (2000). «Микробное разложение крупных органических частиц в северо-западной части Средиземного моря: экспериментальный подход» . Серия «Прогресс в области морской экологии» . 198 : 61–72. Бибкод : 2000MEPS..198...61S . дои : 10.3354/meps198061 .
- ^ Тительман, Дж.; Риман, Л.; Сёрнес, Т.А.; Нильсен, Т.; Грикспур, П.; Бомштедт, У. (2006). «Оборот мертвых медуз: стимуляция и замедление микробной активности» . Серия «Прогресс в области морской экологии» . 325 : 43–58. Бибкод : 2006MEPS..325...43T . дои : 10.3354/meps325043 .
- ^ Талиацеи и углеродный цикл
- ^ Аспер, Вернон Л. (1987). «Измерение потока и скорости опускания агрегатов морского снега». Глубоководные исследования. Часть A. Статьи океанографических исследований . 34 (1): 1–17. Бибкод : 1987DSRA...34....1A . дои : 10.1016/0198-0149(87)90117-8 .
- ^ Джексон и др., 1997.
- ^ Мартин, Джон Х.; Кнауэр, Джордж А.; Карл, Дэвид М.; Броенков, Уильям В. (1987). «VERTEX: Круговорот углерода в северо-восточной части Тихого океана». Глубоководные исследования. Часть A. Статьи океанографических исследований . 34 (2): 267–285. Бибкод : 1987DSRA...34..267M . дои : 10.1016/0198-0149(87)90086-0 .
- ^ Перейти обратно: а б Гелен, М.; Бопп, Л.; Эмприн, Н.; Омон, О.; Хайнце, К.; Рагено, О. (2006). «Согласование продуктивности поверхностного океана, потоков экспорта и состава отложений в глобальной биогеохимической модели океана» . Биогеонауки . 3 (4): 521–537. Бибкод : 2006BGeo....3..521G . дои : 10.5194/bg-3-521-2006 .
- ^ Ильина, Татьяна; Шесть, Катарина Д.; Зегшнейдер, Иоахим; Майер-Реймер, Эрнст; Ли, Хунмэй; Нуньес-Рибони, Исмаэль (2013). «Модель биогеохимии глобального океана HAMOCC: архитектура и производительность модели как компонента модели системы MPI-Земля в различных экспериментальных реализациях CMIP5» . Журнал достижений в моделировании систем Земли . 5 (2): 287–315. Бибкод : 2013JAMES...5..287I . дои : 10.1029/2012MS000178 .
- ^ Лауфкоттер, К.; Фогт, М.; Грубер, Н.; Айта-Ногучи, М.; Омон, О.; Бопп, Л.; Буйтенхейс, Э.; Дони, Южная Каролина; Данн, Дж.; Хашиока, Т.; Хаук, Дж.; Хирата, Т.; Джон, Дж.; Ле Кере, К.; Лима, ID; Накано, Х.; Сефериан Р.; Тоттерделл, И.; Вичи, М.; Фёлькер, К. (2015). «Драйверы и неопределенности будущей глобальной морской первичной продукции в моделях морских экосистем» . Биогеонауки . 12 (23): 6955–6984. Бибкод : 2015BGeo...12.6955L . дои : 10.5194/bg-12-6955-2015 .
- ^ Плауг, Хелле ; Иверсен, Мортен Хвитфельдт; Фишер, Герхард (2008). «Балласт, скорость погружения и кажущаяся диффузия в морском снеге и фекальных гранулах зоопланктона: влияние на круговорот субстрата прикрепленными бактериями» . Лимнология и океанография . 53 (5): 1878–1886. Бибкод : 2008LimOc..53.1878P . дои : 10.4319/lo.2008.53.5.1878 .
- ^ Ломбард, Фабьен; Кьёрбо, Томас (2010). «Морской снег из аппендикулярных домов: характеристики оседания в зависимости от возраста». Глубоководные исследования. Часть I: Статьи океанографических исследований . 57 (10): 1304–1313. Бибкод : 2010DSRI...57.1304L . дои : 10.1016/j.dsr.2010.06.008 .
- ^ Ломбард, Ф.; Лежандр, Л.; Пишераль, М.; Шиандра, А.; Горский, Г. (2010). «Прогнозирование экологических ниш и экспорта углерода аппендикуляристами с использованием новой многовидовой экофизиологической модели» . Серия «Прогресс в области морской экологии» . 398 : 109–125. Бибкод : 2010MEPS..398..109L . дои : 10.3354/meps08273 .
- ^ Уолсби, AE; Ксиполита, Анастасия (1977). «Сопротивление формы хитановых волокон, прикрепленных к клеткам Thalassiosira fluviatilis Hustedt » . Британский психологический журнал . 12 (3): 215–223. дои : 10.1080/00071617700650231 .
- ^ Ямамото, Джун; Хиросе, Миюки; Отани, Тецуя; Сугимото, Каташи; Хирасе, Казуэ; Симамото, Нобуо; Шимура, Цуёси; Хонда, Нацуми; Фухимори, Ясудзуми; Мукаи, Тору (2008). «Перенос органического вещества на морское дно падалью гигантской медузы Nemopilema nomurai в Японском море». Морская биология . 153 (3): 311–317. дои : 10.1007/s00227-007-0807-9 . S2CID 89540757 .
- ^ Макдоннелл, Эндрю, член парламента; Бюсселер, Кен О. (2010). «Изменчивость средней скорости погружения морских частиц» . Лимнология и океанография . 55 (5): 2085–2096. Бибкод : 2010LimOc..55.2085M . дои : 10.4319/lo.2010.55.5.2085 .
- ^ Лебрато, Марио; Мендес, Педро де Хесус; Стейнберг, Дебора К.; Картес, Джоан Э.; Джонс, Бетан М.; Бирса, Лаура М.; Бенавидес, Роберто; Ошлис, Андреас (2013). «Скорость погружения желеобразной биомассы свидетельствует о механизме быстрого экспорта углерода» (PDF) . Лимнология и океанография . 58 (3): 1113–1122. Бибкод : 2013LimOc..58.1113L . дои : 10.4319/lo.2013.58.3.1113 . S2CID 3661129 .
- ^ Лебрато, Марио; Пахлоу, Маркус; Ошлис, Андреас; Питт, Кайли А.; Джонс, Дэниел О.Б.; Молинеро, Хуан Карлос; Кондон, Роберт Х. (2011). «Глубинное ослабление экспорта органических веществ, связанное с падением желе». Лимнология и океанография . 56 (5): 1917–1928. Бибкод : 2011LimOc..56.1917L . дои : 10.4319/lo.2011.56.5.1917 . hdl : 10072/43275 . S2CID 3693276 .
- ^ Беренфельд, Майкл Дж.; Фальковски, Пол Г. (1997). «Скорость фотосинтеза определяется на основе концентрации хлорофилла, полученной со спутников». Лимнология и океанография . 42 (1): 1–20. Бибкод : 1997LimOc..42....1B . дои : 10.4319/lo.1997.42.1.0001 . S2CID 15857675 .
- ^ Лоренцо Корньяти; Симоне Марини; Лука Маццеи; Эннио Оттавиани; Стефано Алиани; Алессандра Конверси; Анналиса Гриффа (2016). «Взгляд внутрь океана: на пути к автономной системе визуализации для мониторинга студенистого зоопланктона» . Датчики . 12 (16): 2124. Бибкод : 2016Senso..16.2124C . дои : 10.3390/s16122124 . ISSN 1424-8220 . OCLC 8148647236 . ПМК 5191104 . ПМИД 27983638 .
- ^ Раскофф К. и Р. Хопкрофт (2010). Кроссота норвежская. Разнообразие Северного Ледовитого океана. По состоянию на 25 августа 2020 г.
Внешние ссылки
[ редактировать ]- Plankton Chronicles Короткометражные документальные фильмы и фотографии
- Ocean Explorer: студенистый зоопланктон из Северного Ледовитого океана
- Медузы и другой студенистый зоопланктон. Архивировано 13 августа 2010 г. в Wayback Machine.
- ПЛАНКТОННАЯ СЕТЬ: информация обо всех видах планктона, включая студенистый зоопланктон.
- Глубоководный студенистый зоопланктон из The Deep (Нувиан, 2007)
