Мегафауна

В зоологии греческого мегафауна (от неолатинского μέγας megas «большой» и фауна « животный мир») — крупные животные. Точное определение этого термина широко варьируется, хотя общий порог составляет примерно 45 килограммов (99 фунтов), а другие пороги составляют от 10 килограммов (22 фунта) или до 1000 килограммов (2200 фунтов). Большой размер тела обычно связан с другими характеристиками, такими как медленная скорость размножения, а у крупных травоядных - пониженная или незначительная смертность взрослых особей от гибели от хищников.
Виды мегафауны оказывают значительное воздействие на местную окружающую среду, включая подавление роста древесной растительности и, как следствие, снижение частоты лесных пожаров . Мегафауна также играет роль в регулировании и стабилизации численности более мелких животных.
В течение плейстоцена мегафауна была разнообразной по всему миру, при этом большинство континентальных экосистем демонстрировали такое же или большее видовое богатство мегафауны по сравнению с экосистемами современной Африки. Во время позднего плейстоцена , особенно примерно 50 000 лет назад, большинство крупных видов млекопитающих вымерло , в том числе 80% всех млекопитающих весом более 1 тонны (2200 фунтов), в то время как мелкие животные практически не пострадали. Это явное вымирание, связанное с размером, является беспрецедентным в геологической летописи. Большинство авторов считают людей и изменение климата вероятными причинами, хотя относительная важность каждого фактора была предметом серьезных споров.
История
[ редактировать ]Одним из первых упоминаний термина «мегафауна» является Альфреда Рассела Уоллеса работа «Географическое распространение животных » 1876 года . Он описал животных как «самые огромные, свирепые и странные формы». В конце 20-го и 21-го веков этот термин обычно относится к крупным животным. Существуют различия в пороговых значениях, используемых для определения мегафауны в целом или отдельных групп мегафауны. Во многих научных публикациях принят предложенный Полом С. Мартином порог в 45 килограммов (99 фунтов) для отнесения животных к мегафунам. Однако для пресноводных видов предпочтительным порогом является 30 кг (66 фунтов). Некоторые ученые определяют растительноядную наземную мегафауну как имеющую вес более 100 килограммов (220 фунтов), а наземную хищную мегафауну - как вес более 15 килограммов (33 фунта). Кроме того, Оуэн-Смит ввел термин «мегагербоядное» для описания травоядных животных весом более 1000 килограммов (2200 фунтов), который использовался другими исследователями. [1]
Среди современных животных термин «мегафауна» чаще всего используется для обозначения крупнейших современных наземных млекопитающих, в число которых входят (но не ограничиваются ими) слоны , жирафы , гиппопотамы , носороги и крупные быки . Из этих пяти категорий крупных травоядных только крупный рогатый скот в настоящее время встречается за пределами Африки и Южной Азии , но все остальные раньше были более широко распространены, а их ареалы и популяции постоянно сокращались и уменьшались с течением времени. Дикие лошади являются еще одним примером мегафауны, но их нынешние ареалы в основном ограничены Старым Светом , особенно Африкой и Азией. Виды мегафауны можно разделить на категории в зависимости от типа их питания: мегатравоядные (например, слоны ), мегахищные (например, львы ) и, реже, мегавсеядные (например, медведи ). [2] [3]
Экологическая стратегия
[ редактировать ]Животные мегафауны – в смысле крупнейших млекопитающих и птиц – обычно являются K -стратегами , с высокой продолжительностью жизни, медленными темпами роста популяции, низким уровнем смертности и (по крайней мере для самых крупных) небольшим количеством или отсутствием естественных хищников, способных убивать взрослых особей. [4] [1] Эти характеристики, хотя и не являются исключительными для такой мегафауны, делают их уязвимыми для чрезмерной эксплуатации человеком , отчасти из-за медленных темпов восстановления их популяции. [5] [6]
Эволюция больших размеров тела
[ редактировать ]Одно наблюдение, сделанное в отношении эволюции большего размера тела, заключается в том, что быстрые темпы увеличения, которые часто наблюдаются в течение относительно коротких интервалов времени, не являются устойчивыми в течение гораздо более длительных периодов времени. При изучении изменений массы тела млекопитающих с течением времени было обнаружено, что максимально возможное увеличение в заданном интервале времени масштабируется с увеличением длины интервала в степени 0,25. [7] Считается, что это отражает появление в ходе тенденции к увеличению максимального размера тела ряда анатомических, физиологических, экологических, генетических и других ограничений, которые необходимо преодолеть с помощью эволюционных инноваций, прежде чем дальнейшее увеличение размеров станет возможным. Поразительно более высокая скорость изменений была обнаружена при значительном уменьшении массы тела, что может быть связано с феноменом островной карликовости . При нормализации к длине поколения было обнаружено, что максимальная скорость уменьшения массы тела более чем в 30 раз превышает максимальную скорость увеличения массы тела при десятикратном изменении. [7]
У наземных млекопитающих
[ редактировать ]
После мел -палеогенового вымирания , которое уничтожило нептичьих динозавров около 66 млн лет назад (миллионов лет) назад, наземные млекопитающие претерпели почти экспоненциальное увеличение размеров тела, поскольку они диверсифицировались, чтобы занять экологические ниши, оставшиеся вакантными. [8] Начиная с нескольких кг до события, максимальный размер достиг ~ 50 кг несколько миллионов лет спустя и ~ 750 кг к концу палеоцена . Эта тенденция увеличения массы тела, по-видимому, выровнялась около 40 млн лет назад (в позднем эоцене ), что позволяет предположить, что физиологические или экологические ограничения были достигнуты после увеличения массы тела более чем на три порядка. [8] Однако, если рассматривать с точки зрения скорости увеличения размера за поколение, обнаруживается, что экспоненциальный рост продолжался до появления индрикотерия 30 млн лет назад. (Поскольку время генерации зависит от массы тела 0.259 , увеличение времени генерации с увеличением размера приводит к тому, что график зависимости массы бревна от времени изгибается вниз от линейной аппроксимации.) [7]
Мегатравоядные животные в конечном итоге достигли массы тела более 10 000 кг. Самые крупные из них, индрикотерии и хоботки , представляют собой ферментеры задней кишки , которые, как полагают, имеют преимущество перед ферментаторами передней кишки с точки зрения способности ускорять транзит по желудочно-кишечному тракту, чтобы приспособиться к очень большому объему потребляемой пищи. [9] темпов увеличения максимальной массы тела за поколение для разных клад Аналогичная тенденция проявляется при сравнении млекопитающих (с использованием темпов, усредненных по макроэволюционным временным масштабам). Среди наземных млекопитающих самые быстрые темпы увеличения массы тела 0.259 в зависимости от времени (в млн лет назад) наблюдались у непарнокопытных (наклон 2,1), за которыми следовали грызуны (1,2) и хоботные (1,1), [7] все они являются ферментаторами задней кишки. Скорость увеличения парнокопытных (0,74) составила примерно треть от темпа увеличения непарнокопытных. Этот показатель для хищных животных (0,65) был еще немного ниже, в то время как у приматов , возможно, из-за их древесных привычек, показатель был самым низким (0,39) среди исследованных групп млекопитающих. [7]
Наземные млекопитающие-хищники из нескольких эвтерий групп ( парнокопытные Эндрюсарх – ранее считавшиеся мезонихидами , оксиениды Саркастодон и хищные животные Амфикион и Арктодус ) все достигали максимального размера около 1000 кг. [8] (хищник Arctotherium и гиенодонтид Симбакубва , возможно, были несколько крупнее). Самый крупный из известных метатерийных хищников, Proborhyaena gigantea , по-видимому, достигал 600 кг, что также близко к этому пределу. [10] Аналогичный теоретический максимальный размер хищных млекопитающих был предсказан на основе скорости метаболизма млекопитающих, энергетических затрат на добычу добычи и максимального расчетного коэффициента скорости поглощения добычи. [11] Также было высказано предположение, что максимальный размер хищных млекопитающих ограничен нагрузкой, которую плечевая кость может выдержать при максимальной скорости бега. [10]
Анализ изменения максимального размера тела за последние 40 млн лет показывает, что снижение температуры и увеличение площади континентальной суши связаны с увеличением максимального размера тела. Первая корреляция соответствовала бы правилу Бергмана : [12] и может быть связано с терморегуляторным преимуществом большой массы тела в прохладном климате. [8] лучшая способность более крупных организмов справляться с сезонностью поставок пищи, [12] или другие факторы; [12] последнюю корреляцию можно объяснить ограничениями дальности действия и ресурсов. [8] Однако эти два параметра взаимосвязаны (из-за падения уровня моря, сопровождающего усиление оледенения), что затрудняет определение движущей силы тенденций максимального размера. [8]
У морских млекопитающих
[ редактировать ]
С тех пор как четвероногие (сначала рептилии , а затем млекопитающие ) вернулись в море в поздней перми , они доминировали в верхней части диапазона размеров морского тела из-за более эффективного поглощения кислорода, возможного с помощью легких. [13] [14] Считается, что предками китообразных были полуводные пакицетиды размером не крупнее собак, жившие около 53 миллионов лет назад. [15] К 40 млн лет назад китообразные достигли длины 20 м и более у Basilosaurus , удлиненного змеевидного кита, который во многих отношениях отличался от современных китов и не был их предком. После этого эволюция крупных размеров тела у китообразных, по-видимому, временно остановилась, а затем пошла вспять, хотя имеющиеся летописи окаменелостей ограничены. Однако в период с 31 млн лет назад (в олигоцене ) по настоящее время у китообразных наблюдался значительно более быстрый устойчивый рост массы тела (скорость увеличения массы тела 0.259 в 3,2 раза на миллион лет), чем достигается любой группой наземных млекопитающих. [7] Эта тенденция привела к появлению самого крупного животного всех времен — современного синего кита . Возможны несколько причин более быстрой эволюции крупных размеров тела у китообразных. Меньшие биомеханические ограничения на увеличение размеров тела могут быть связаны с подвешиванием в воде, а не с стоянием против силы тяжести, а также с плавательными движениями, в отличие от передвижения по суше . Кроме того, большая теплоемкость и теплопроводность воды по сравнению с воздухом может увеличить терморегулирующее преимущество большого размера тела у морских эндотермистов , хотя и снижается отдача. [7]
У зубатых китов максимальный размер тела, по-видимому, ограничен наличием пищи. Больший размер, как у кашалотов и клюворылых китов , облегчает более глубокое ныряние для доступа к относительно легко поймаемой крупной добыче головоногих моллюсков в менее конкурентной среде. По сравнению с зубатыми китообразными, эффективность фильтрационного питания усатых китов более благоприятно возрастает с увеличением размера, когда планктонная пища плотная, что делает больший размер более выгодным. Техника кормления рорквалов с выпадом оказывается кормление более энергоэффективной, чем балаеновых китов таранное ; последний метод используется с менее плотным и пятнистым планктоном. [16] Тенденция к похолоданию в новейшей истории Земли, возможно, привела к появлению новых мест с высокой численностью планктона посредством ветровых апвеллингов , способствуя эволюции гигантских китов. [16]
Китообразные — не единственные морские млекопитающие, достигающие огромных размеров. [17] Крупнейшими млекопитающими- хищниками всех времен являются морские ластоногие , самым крупным из которых является южный морской слон , который может достигать более 6 м (20 футов) в длину и весить до 5000 кг (11 000 фунтов). Среди других крупных ластоногих - северный морской слон весом 4000 кг (8800 фунтов), морж весом 2000 кг (4400 фунтов) и сивуч весом 1135 кг (2502 фунта). [18] [19] Сирены — еще одна группа морских млекопитающих, которые адаптировались к полностью водной жизни примерно в то же время , что и китообразные. Сирены тесно связаны со слонами. Самой крупной сиреной была стеллерова морская корова , которая достигала 10 м (33 фута) в длину и весила от 8 000 до 10 000 кг (от 18 000 до 22 000 фунтов) и была истреблена в 18 веке. [20]
У нелетающих птиц
[ редактировать ]
Из-за небольшого первоначального размера всех млекопитающих после вымирания нептичьих динозавров, немлекопитающие позвоночные имели примерно десять миллионов лет окна возможностей (во время палеоцена) для эволюции гигантизма без особой конкуренции. [21] В течение этого периода ниши высших хищников часто были заняты рептилиями, такими как наземные крокодилы (например, Pristihampsus ), крупные змеи (например, Титанобоа ) или ящерицы-вараниды , или нелетающие птицы. [8] (например, Paleopsilopterus в Южной Америке). Это также период, когда нелетающие травоядные гасторнитиды- в Северном полушарии появились мегафауны, а нелетающие палеогнаты развились до крупных размеров на Гондваны суше и в Европе . Гасторнитиды и, по крайней мере, одна линия нелетающих птиц-палеогнатов произошли в Европе, причем обе линии доминировали в нишах крупных травоядных животных, в то время как млекопитающие оставались ниже 45 кг (в отличие от других территорий, таких как Северная Америка и Азия , где произошла более ранняя эволюция более крупных млекопитающих) и были Крупнейшие европейские четвероногие палеоцена . [22]
Нелетающие палеогнаты, называемые бескилевыми , традиционно рассматривались как представители линии, отдельной от их мелких летающих родственников, неотропных тинамусов . Однако недавние генетические исследования показали, что тинамус хорошо гнездится внутри бескилевых деревьев и является сестринской группой вымерших моа Новой Зеландии. [21] [23] [24] Точно так же было обнаружено, что маленькие киви из Новой Зеландии являются сестринской группой вымерших слоновых птиц Мадагаскара. [21] Эти данные показывают, что нелетание и гигантизм возникали независимо несколько раз среди бескилевых в ходе параллельной эволюции . [25]
Хищные нелетающие птицы мегафауны нередко могли конкурировать с млекопитающими в раннем кайнозое . Однако позже в кайнозое они были вытеснены высокоразвитыми хищниками и вымерли. В Северной Америке баторнитиды Paracrax и Bathornis были высшими хищниками, но вымерли к раннему миоцену . В Южной Америке родственные форусрациды разделяли доминирующие хищные ниши с метатерийными спарассодонтами на протяжении большей части кайнозоя, но их численность сократилась и в конечном итоге вымерла после того, как эвтерийные хищники прибыли из Северной Америки (в рамках Великого американского обмена ) во время плиоцена . Напротив, до настоящего времени сохранились крупные травоядные нелетающие бескилевые. [25]
Однако ни одна из нелетающих птиц кайнозоя, в том числе хищный Brontornis , возможно, всеядный Dromornis stirtoni [25] или травоядный Aepyornis , когда-либо вырастал до массы, значительно превышающей 500 кг, и поэтому никогда не достигал размеров крупнейших хищных млекопитающих, не говоря уже о размерах крупнейших травоядных млекопитающих. Было высказано предположение, что увеличение толщины яичной скорлупы птиц пропорционально массе яйца с увеличением размера яйца устанавливает верхний предел размера птиц. [26] [примечание 1] Самый крупный вид Dromornis , D. stirtoni , возможно, вымер после того, как достиг максимальной птичьей массы тела, а затем был вытеснен сумчатыми дипротодонтами , которые в ходе эволюции достигли размеров в несколько раз больше. [29]
У гигантских черепах
[ редактировать ]Гигантские черепахи были важными компонентами позднекайнозойской мегафауны , присутствуя на каждом неполярном континенте до прибытия гоминидов . [30] [31] Самой крупной известной наземной черепахой была Megalochelys atlas , животное, которое, вероятно, весило около 1000 кг (2200 фунтов). [32]
Некоторые более ранние водные Testudines, например морской архелон мелового периода. [33] и пресноводные Stupendemys миоцена были значительно крупнее и весили более 2000 кг (4400 фунтов). [34]
Массовые вымирания мегафауны
[ редактировать ]Сроки и возможные причины
[ редактировать ]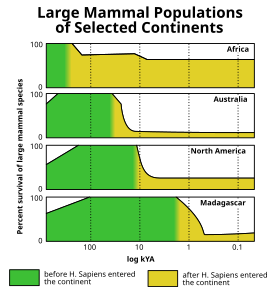
Многочисленные вымирания произошли во второй половине последнего ледникового периода , когда вымерло большинство крупных млекопитающих в Северной и Южной Америке , Австралии, Новой Гвинее и северной Евразии , включая более 80% всех наземных животных с массой тела более 1000 кг (2200 фунтов). ). Мелкие животные и другие организмы, такие как растения, в целом не пострадали от вымирания, что является беспрецедентным явлением в предыдущих вымираниях за последние 30 миллионов лет. [35]
Различные теории объясняют волну вымирания человеческой охотой , изменением климата , болезнями , инопланетным воздействием , конкуренцией со стороны других животных или другими причинами. Однако это вымирание ближе к концу плейстоцена было лишь одним из серии импульсов вымирания мегафауны, которые произошли в течение последних 50 000 лет на большей части поверхности Земли, в Африке и Южной Азии (где местная мегафауна имела шанс развиться). наряду с современными людьми), подвергаясь сравнительно меньшему воздействию. Последние районы действительно страдали от постепенного истощения мегафауны, особенно медленно движущихся видов (класс уязвимой мегафауны, представленной гигантскими черепахами ), в течение последних нескольких миллионов лет. [36] [37]
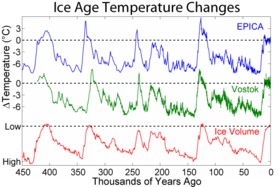
За пределами материковой части Афро-Евразии эти вымирания мегафауны следовали весьма своеобразной модели суши за территорией, которая во многом аналогична распространению людей в ранее необитаемые регионы мира и не показывает никакой общей корреляции с климатической историей (которую можно визуализировать с графиками климатических маркеров последних геологических периодов времени, таких как морские изотопы кислорода или уровни углекислого газа в атмосфере ). [38] [39] Австралия [40] и близлежащие острова (например, Флорес [41] ) были поражены первыми около 46 000 лет назад, за ними последовала Тасмания около 41 000 лет назад (после образования сухопутного моста в Австралию около 43 000 лет назад). [42] [43] [44] Роль людей в вымирании мегафауны Австралии и Новой Гвинеи оспаривается: многочисленные исследования показывают сокращение числа видов до прибытия людей на континент и отсутствие каких-либо свидетельств хищничества человека; [45] [46] [47] [48] Вместо этого в качестве причины их снижения упоминалось влияние изменения климата. [49] [45] Точно так же Япония потеряла большую часть своей мегафауны, по-видимому, около 30 000 лет назад. [50] Северная Америка 13 000 лет назад. [примечание 2] и Южная Америка примерно 500 лет спустя, [52] [53] Кипр 10 000 лет назад, [54] [55] Антильские острова 6000 лет назад, [56] [57] Новая Каледония [58] и близлежащие острова [59] 3000 лет назад, Мадагаскар 2000 лет назад, [60] Новая Зеландия 700 лет назад. [61] Маскаренцы , 400 лет назад [62] и Командорские острова 250 лет назад. [63] Почти все изолированные острова мира могут служить примером вымирания, произошедшего вскоре после прибытия людей , хотя на большинстве этих островов, таких как Гавайские острова , никогда не было наземной мегафауны, поэтому их вымершая фауна была меньше, но все же демонстрировала островной гигантизм. . [38] [39]
Анализ сроков вымирания и истребления голарктической мегафауны за последние 56 000 лет выявил тенденцию таких событий группироваться в межстадиальные периоды , периоды резкого потепления, но только тогда, когда также присутствовали люди. Люди, возможно, препятствовали процессам миграции и реколонизации, которые в противном случае позволили бы видам мегафауны адаптироваться к изменению климата. [64] По крайней мере, в некоторых областях интерстадиалы были периодами роста населения. [65]
Анализ грибковых спор Sporormiella (которые происходят в основном из навоза мегагербоядных) в кернах болотных отложений, охватывающих последние 130 000 лет из кратера Линча в Квинсленде , Австралия, показал, что мегафауна этого региона практически исчезла около 41 000 лет назад. когда изменения климата были минимальными; это изменение сопровождалось увеличением использования древесного угля, а затем последовал переход от тропических лесов к огнестойкой склерофилловой растительности. Хронология изменений с высоким разрешением подтверждает гипотезу о том, что человеческая охота сама по себе уничтожила мегафауну и что последующее изменение флоры, скорее всего, было следствием исчезновения браузеров и увеличения количества пожаров. [66] [67] [68] [69] Рост пожаров отстал от исчезновения мегафауны примерно на столетие и, скорее всего, стал результатом накопления топлива после прекращения просмотра. В течение следующих нескольких столетий трава увеличилась; Склерофилльная растительность увеличилась с опозданием еще на столетие, а склерофилловый лес развился примерно еще через тысячу лет. [68] В течение двух периодов изменения климата, около 120 000 и 75 000 лет назад, на этом месте также увеличилась склерофилловая растительность в ответ на переход к более прохладным и засушливым условиям; ни один из этих эпизодов не оказал существенного влияния на численность мегафауны. [68] Аналогичные выводы относительно виновности людей-охотников в исчезновении плейстоценовой мегафауны были сделаны на основе хронологий высокого разрешения, полученных посредством анализа большой коллекции фрагментов яичной скорлупы нелетающей австралийской птицы Genyornis newtoni . [70] [71] [69] на основе анализа спор грибка Sporormiella из озера на востоке Северной Америки. [72] [73] и в результате изучения отложений навоза наземного ленивца Шаста, оставшегося более чем в полудюжине пещер на юго-западе Америки. [74] [75]
Продолжающаяся человеческая охота и нарушение окружающей среды привели к дополнительному вымиранию мегафауны в недавнем прошлом и создали серьезную опасность дальнейшего вымирания в ближайшем будущем (см. примеры ниже). Прямое убийство людьми, в первую очередь ради мяса или других частей тела, является наиболее значимым фактором современного упадка мегафауны. [76] [77]
Ряд других массовых вымираний произошел ранее в геологической истории Земли, в ходе которых также вымерла часть или вся мегафауна того времени. Известно, что во время мел-палеогенового вымирания были уничтожены нептичьи динозавры и большинство других гигантских рептилий. Однако более ранние массовые вымирания были более глобальными и не столь избирательными для мегафауны; т. е. многие виды других типов, включая растения, морских беспозвоночных [78] и планктон тоже вымерли. Таким образом, более ранние события должны были быть вызваны более общими типами нарушений биосферы . [79]
Последствия истощения мегафауны
[ редактировать ]Истощение растительноядной мегафауны приводит к увеличению роста древесной растительности, [80] и, как следствие, увеличение частоты лесных пожаров . [81] Мегафауна может помочь подавить рост инвазивных растений. [82] Крупные травоядные и плотоядные животные могут подавлять численность более мелких животных, что приводит к увеличению их популяции при удалении мегафауны. [80]
Влияние на транспорт питательных веществ
[ редактировать ]Мегафауна играет значительную роль в латеральном переносе минеральных питательных веществ в экосистеме, имея тенденцию перемещать их из областей с высокой численностью в места с меньшей численностью. Они делают это за счет своего перемещения между моментом потребления питательного вещества и моментом высвобождения его путем элиминации (или, в гораздо меньшей степени, путем разложения после смерти). [83] в Южной Америке такая латеральная диффузия сократилась более чем на 98% после вымирания мегафауны, произошедшего примерно 12 500 лет назад. в бассейне Амазонки По оценкам, [84] [85] Учитывая, что доступность фосфора, как полагают, ограничивает продуктивность на большей части региона, считается, что уменьшение его переноса из западной части бассейна и из пойм рек (обе из которых получают свое питание за счет поднятия Анд ) в другие районы существенно повлияли на экологию региона, и последствия, возможно, еще не достигли своего предела. [85] Считается, что в море китообразные и ластоногие, питающиеся на глубине, переносят азот с глубокой воды на мелководье, повышая продуктивность океана и противодействуя активности зоопланктона , который имеет тенденцию делать противоположное. [86]
Влияние на выбросы метана
[ редактировать ]Большие популяции мегатравоядных животных могут внести значительный вклад в концентрацию в атмосфере метана , который является важным парниковым газом . Современные жвачные травоядные животные производят метан как побочный продукт ферментации передней кишки при пищеварении и выделяют его через отрыжку или метеоризм. Сегодня около 20% годовых выбросов метана приходится на выбросы метана в результате животноводства. в мезозойскую По оценкам, эпоху зауроподы могли ежегодно выбрасывать в атмосферу 520 миллионов тонн метана. [87] способствуя более теплому климату того времени (до 10 ° C теплее, чем в настоящее время). [87] [88] Столь большие выбросы обусловлены огромной предполагаемой биомассой зауроподов, а также тем, что производство метана отдельными травоядными считается почти пропорциональным их массе. [87]
Недавние исследования показали, что исчезновение мегафауны травоядных могло привести к сокращению количества метана в атмосфере . Эта гипотеза является относительно новой. [89] В одном исследовании изучались выбросы метана бизонами , населявшими Великие равнины Северной Америки до контакта с европейскими поселенцами. По оценкам исследования, уничтожение бизонов привело к сокращению объёмов добычи на 2,2 миллиона тонн в год. [90] Другое исследование изучало изменение концентрации метана в атмосфере в конце эпохи плейстоцена после вымирания мегафауны в Америке. После того, как первые люди мигрировали в Америку около 13 000 лет назад , их охота и другие связанные с этим экологические воздействия привели к исчезновению там многих видов мегафауны. Расчеты показывают, что это вымирание снизило производство метана примерно на 9,6 миллиона тонн в год. Это говорит о том, что отсутствие мегафаунистических выбросов метана могло способствовать резкому похолоданию климата в начале позднего дриаса . [89] Уменьшение содержания метана в атмосфере, произошедшее в то время, как зафиксировано в ледяных кернах , было в 2–4 раза быстрее, чем любое другое уменьшение за последние полмиллиона лет, что позволяет предположить, что действовал необычный механизм. [89]
Галерея
[ редактировать ]Вымершая плейстоценовая мегафауна
[ редактировать ] Моа ( на фото Динорнис )
Моа ( на фото Динорнис )
 « Мегалания » ( Varanus priscus ),
« Мегалания » ( Varanus priscus ), Американские львы ( Panthera atrox )
Американские львы ( Panthera atrox )
 Субфоссильный лемур Archaeoindris
Субфоссильный лемур Archaeoindris
 Восстановление Macrauchenia , представителя вымершего отряда копытных Litopterna размером с верблюда.
Восстановление Macrauchenia , представителя вымершего отряда копытных Litopterna размером с верблюда.









Другая вымершая кайнозойская мегафауна
[ редактировать ]
 Азиатский индрикотерий и родственник носорога Paraceratherium был одним из крупнейших наземных млекопитающих. [91]
Азиатский индрикотерий и родственник носорога Paraceratherium был одним из крупнейших наземных млекопитающих. [91] Реконструированные челюсти мегалодона ( Otodus megalodon )
Реконструированные челюсти мегалодона ( Otodus megalodon )








Сохранившийся
[ редактировать ] Большая Рея
Большая Рея горилла Восточная
горилла Восточная

 Черный носорог
Черный носорог В отличие от шерстистых носорогов и мамонтов , овцебыки чудом пережили четвертичное вымирание . [92]
В отличие от шерстистых носорогов и мамонтов , овцебыки чудом пережили четвертичное вымирание . [92]
 Кашалот
Кашалот  Косатка
Косатка  Южный казуар
Южный казуар Обыкновенный страус
Обыкновенный страус Морской крокодил
Морской крокодил Дракон Комодо
Дракон Комодо Зеленая анаконда
Зеленая анаконда Китайская гигантская саламандра
Китайская гигантская саламандра Гигантская солнечная рыба
Гигантская солнечная рыба Нильский окунь
Нильский окунь Большая белая акула , крупнейшая хищная рыба и один из крупнейших видов хищных акул, встречается по всему миру. [93]
Большая белая акула , крупнейшая хищная рыба и один из крупнейших видов хищных акул, встречается по всему миру. [93] Манта
Манта  Осмотр 9-метрового гигантского кальмара
Осмотр 9-метрового гигантского кальмара




















См. также
[ редактировать ]- Австралийская мегафауна
- Правило Бергмана
- Харизматическая мегафауна
- Правило Коупа
- Глубоководный гигантизм
- Островной гигантизм
- Крупнейшие организмы
- Крупнейшие доисторические животные
- Список самых тяжелых наземных млекопитающих
- Список крупнейших млекопитающих
- Список мегафауны, обнаруженной в наше время
- Мегафауна (мифология)
- Мегафаунальный волк
- Мегафлора
- Мегахерб
- Вымирания плейстоцена Нового Света
- Плейстоценовая мегафауна
- Четвертичное вымирание
Примечания
[ редактировать ]- ^ Размер нонавианских динозавров не был таким же ограничен, поскольку у них была другая взаимосвязь между массой тела и размером яиц, чем у птиц. Яйца 400-килограммового эпиорниса были крупнее, чем почти у всех динозавров. [27] [28]
- ^ Анализ показывает, что в результате этого события более или менее одновременно вымерли 35 родов млекопитающих Северной Америки. [51]
Ссылки
[ редактировать ]- ^ Jump up to: а б Гриндер М, Санчес-Сапата Х.А., Доназар Х.А., Ревилла Е., Мартин-Лопес Б., Гутьеррес-Канос С., Гетц В.М., Моралес-Рейес З., Филдс-Арсес А., Краудер Л.Б., Галетти М., Гонсалес-Суарес М., Хе Ф. , Джордан П., Льюисон Р. (11 марта 2020 г.). «Переосмысление мегафауны» . Труды Королевского общества B: Биологические науки . 287 (1922): 20192643 doi : 10.1098/rspb.2019.2643 . hdl : 2263/79439 . ISSN 0962-8452 .
- ^ Малхи Ю., Даути С.Э., Галетти М., Смит Ф.А., Свеннинг Дж.К., Терборг Дж.В. (26 января 2016 г.). «Мегафауна и экосистема функционируют от плейстоцена до антропоцена» . Труды Национальной академии наук . 113 (4): 838–846. Бибкод : 2016PNAS..113..838M . дои : 10.1073/pnas.1502540113 . ISSN 0027-8424 . ПМЦ 4743772 . ПМИД 26811442 .
- ^ МакКленачан Л., Купер А.Б., Далви Н.К. (20 июня 2016 г.). «Переосмысление риска исчезновения морской и наземной мегафауны, вызванного торговлей» . Современная биология . 26 (12): 1640–1646. Бибкод : 2016CBio...26.1640M . дои : 10.1016/j.cub.2016.05.026 . ISSN 1879-0445 . ПМИД 27291051 .
- ^ Джонсон CN (07 ноября 2002 г.). «Детерминанты исчезновения видов млекопитающих во время вымирания мегафауны в позднечетвертичное время: история жизни и экология, но не размер тела» . Труды Лондонского королевского общества. Серия Б: Биологические науки . 269 (1506): 2221–2227. дои : 10.1098/рспб.2002.2130 . ISSN 0962-8452 . ПМК 1691151 . ПМИД 12427315 .
- ^ Барноский А.Д. (01.10.2004). «Оценка причин позднеплейстоценовых вымираний на континентах». Наука . 306 (5693): 70–75. Бибкод : 2004Sci...306...70B . CiteSeerX 10.1.1.574.332 . дои : 10.1126/science.1101476 . ПМИД 15459379 . S2CID 36156087 .
- ^ Брук Б.В., Джонсон CN (2006). «Выборочная охота на молодь как причина незаметного чрезмерного истребления мегафауны австралийского плейстоцена». Алчеринга: Австралазийский журнал палеонтологии . 30 (суп1): 39–48. Бибкод : 2006Алч...30С..39Б . дои : 10.1080/03115510609506854 . S2CID 84205755 .
- ^ Jump up to: а б с д и ж г Эванс А.Р., Джонс Д., Бойер А.Г., Браун Дж.Х., Коста Д.П., Эрнест С.К., Фицджеральд Э.М., Фортелиус М., Гиттлман Дж.Л., Гамильтон М.Дж., Хардинг Л.Е., Линтулааксо К., Лайонс С.К., Оки Дж.Г., Сааринен Дж.Дж., Сибли Р.М., Смит Ф.А. , Стивенс П.Р., Теодор Дж.М., Уэн, доктор медицинских наук (30 января 2012 г.). «Максимальная скорость эволюции млекопитающих» . ПНАС . 109 (11): 4187–4190. Бибкод : 2012PNAS..109.4187E . дои : 10.1073/pnas.1120774109 . ПМК 3306709 . ПМИД 22308461 .
- ^ Jump up to: а б с д и ж г Смит Ф.А., Бойер А.Г., Браун Дж.Х., Коста Д.П., Даян Т., Эрнест С.К., Эванс А.Р., Фортелиус М., Гиттлман Дж.Л., Гамильтон М.Дж., Хардинг Л.Е., Линтулааксо К., Лайонс С.К., Маккейн С., Оки Дж.Г., Сааринен Дж.Дж., Сибли Р.М. , Стивенс П.Р., Теодор Дж., Уэн, доктор медицинских наук (26 ноября 2010 г.). «Эволюция максимального размера тела наземных млекопитающих». Наука . 330 (6008): 1216–1219. Бибкод : 2010Sci...330.1216S . CiteSeerX 10.1.1.383.8581 . дои : 10.1126/science.1194830 . ПМИД 21109666 . S2CID 17272200 .
- ^ Клаусс М., Фрей Р., Кифер Б., Лехнер-Долл М., Лелейн В., Полстер К., Росснер Г.Е., Штрайх У.Дж. (24 апреля 2003 г.). «Максимально достижимый размер тела травоядных млекопитающих: морфофизиологические ограничения передней кишки и адаптации ферментеров задней кишки» (PDF) . Экология . 136 (1): 14–27. Бибкод : 2003Oecol.136...14C . дои : 10.1007/s00442-003-1254-z . ПМИД 12712314 . S2CID 206989975 . Архивировано из оригинала (PDF) 8 июня 2019 г. Проверено 13 июля 2019 г.
- ^ Jump up to: а б Соркин Б. (10 апреля 2008 г.). «Биомеханическое ограничение массы тела у наземных хищников-млекопитающих». Летайя . 41 (4): 333–347. Бибкод : 2008Лета..41..333С . дои : 10.1111/j.1502-3931.2007.00091.x .
- ^ Карбон С., Учитель А., Роуклифф Дж. М. (16 января 2007 г.). «Издержки плотоядства» . ПЛОС Биология . 5 (2, e22): 363–368. doi : 10.1371/journal.pbio.0050022 . ПМК 1769424 . ПМИД 17227145 .
- ^ Jump up to: а б с Эштон К.Г., Трейси, MC, де Кейрос, А. (октябрь 2000 г.). «Действительно ли правило Бергмана для млекопитающих?». Американский натуралист . 156 (4): 390–415. дои : 10.1086/303400 . JSTOR 10.1086/303400 . ПМИД 29592141 . S2CID 205983729 .
- ^ Уэбб Дж. (19 февраля 2015 г.). «Эволюция «отдает предпочтение более крупным морским существам» » . Новости Би-би-си . Би-би-си . Архивировано из оригинала 22 февраля 2015 г. Проверено 22 февраля 2015 г.
- ^ Хейм Н.А., Ноуп М.Л., Шаал Е.К., Ван С.К., Пейн Дж.Л. (20 февраля 2015 г.). «Правило Коупа в эволюции морских животных» . Наука . 347 (6224): 867–870. Бибкод : 2015Sci...347..867H . дои : 10.1126/science.1260065 . ПМИД 25700517 . Архивировано из оригинала 5 июля 2019 г. Проверено 13 июля 2019 г.
- ^ Тьювиссен Дж.Г., Баджпай С. (1 января 2001 г.). «Происхождение китов как образец макроэволюции» . Бионаука . 51 (12): 1037–1049. doi : 10.1641/0006-3568(2001)051[1037:WOAAPC]2.0.CO;2 . ISSN 0006-3568 .
- ^ Jump up to: а б Голдбоген Дж.А., Кейд Д.Э., Вишневска Д.М., Потвин Дж., Сегре П.С., Савока М.С., Хазен Е.Л., Чапанский М.Ф., Кахане-Раппорт С.Р., ДеРуитер С.Л., Геро С., Тоннесен П., Гоф В.Т., Хансон М.Б., Холт М.М., Дженсен Ф.Х., Саймон М., Симперт А.К., Арранц П., Джонстон Д.В., Новачек Д.П., Паркс С.Е., Виссер Ф., Фридлендер А.С., Тайак П.Л., Мэдсен П.Т., Пенсон Н.Д. (2019). «Почему киты большие, но не больше: физиологические факторы и экологические ограничения в эпоху океанских гигантов». Наука . 366 (6471): 1367–1372. Бибкод : 2019Sci...366.1367G . дои : 10.1126/science.aax9044 . hdl : 10023/19285 . ПМИД 31831666 . S2CID 209339266 .
- ^ Бейкер Дж., Мид А., Пейджел М., Вендитти С. (21 апреля 2015 г.). «Адаптивная эволюция к увеличению размеров млекопитающих» . Труды Национальной академии наук . 112 (16): 5093–5098. Бибкод : 2015PNAS..112.5093B . дои : 10.1073/pnas.1419823112 . ISSN 0027-8424 . ПМЦ 4413265 . ПМИД 25848031 .
- ^ Черчилль М., Клеменц М.Т., Коно Н. (19 декабря 2014 г.). «Правило Коупа и эволюция размеров тела у Pinnipedimorpha (Mammalia: Carnivora)». Эволюция . 69 (1): 201–215. дои : 10.1111/evo.12560 . ISSN 0014-3820 . ПМИД 25355195 .
- ^ Хейли, член парламента, Deutsch CJ, Boeuf BJ (апрель 1991 г.). «Метод оценки массы крупных ластоногих». Наука о морских млекопитающих . 7 (2): 157–164. Бибкод : 1991MMamS...7..157H . дои : 10.1111/j.1748-7692.1991.tb00562.x . ISSN 0824-0469 .
- ^ Гольдбоген Дж. А. (17 апреля 2018 г.). «Физиологические ограничения на размер тела морских млекопитающих» . Труды Национальной академии наук . 115 (16): 3995–3997. Бибкод : 2018PNAS..115.3995G . дои : 10.1073/pnas.1804077115 . ISSN 0027-8424 . ПМЦ 5910879 . ПМИД 29618615 .
- ^ Jump up to: а б с Митчелл К.Дж., Лламас Б., Субриер Дж., Роуленс Нью-Джерси, Уорти Т.Х., Вуд Дж., Ли М.С., Купер А. (23 мая 2014 г.). «Древняя ДНК показывает, что птицы-слоны и киви являются родственными таксонами, и проясняет эволюцию бескилевых птиц» (PDF) . Наука . 344 (6186): 898–900. Бибкод : 2014Sci...344..898M . дои : 10.1126/science.1251981 . hdl : 2328/35953 . ПМИД 24855267 . S2CID 206555952 . Архивировано (PDF) из оригинала 15 марта 2023 г. Проверено 24 сентября 2019 г.
- ^ Баффетаут Э., Ангст Д. (ноябрь 2014 г.). «Стратиграфическое распространение крупных нелетающих птиц в палеогене Европы и его палеобиологические и палеогеографические последствия». Обзоры наук о Земле . 138 : 394–408. Бибкод : 2014ESRv..138..394B . doi : 10.1016/j.earscirev.2014.07.001 .
- ^ Филлипс М.Дж., Гибб Г.К., Кримп Е.А., Пенни Д. (январь 2010 г.). «Тинамус и моа собираются вместе: анализ последовательности митохондриального генома показывает независимые потери способности летать среди бескилевых» . Систематическая биология . 59 (1): 90–107. дои : 10.1093/sysbio/syp079 . ПМИД 20525622 .
- ^ Бейкер А.Дж., Хадрат О., Макферсон Дж.Д., Клотье А. (2014). «Геномная поддержка клады Моа-Тинаму и адаптивная морфологическая конвергенция нелетающих бескилевых» . Молекулярная биология и эволюция . 31 (7): 1686–1696. дои : 10.1093/molbev/msu153 . ПМИД 24825849 .
- ^ Jump up to: а б с Мюррей П.Ф., Викерс-Рич П. (2004). Великолепные михирунги: колоссальные нелетающие птицы австралийской эпохи грез . Издательство Университета Индианы. стр. 51, 314. ISBN. 978-0-253-34282-9 . Проверено 7 января 2012 г.
- ^ Там же (2004). п. 212 . Издательство Университета Индианы. ISBN 978-0-253-34282-9 .
- ^ Кеннет Карпентер (1999). Яйца, гнезда и детеныши динозавров: взгляд на размножение динозавров . Издательство Университета Индианы . п. 100 . ISBN 978-0-253-33497-8 . OCLC 42009424 . Проверено 6 мая 2013 г.
- ^ Джексон Ф.Д., Варриккио DJ, Джексон Р.А., Вила Б., Кьяппе Л.М. (2008). «Сравнение проводимости водяного пара в яйце титанозавра из верхнего мела Аргентины и яйце Megaloolithus siruguei из Испании». Палеобиология . 34 (2): 229–246. doi : 10.1666/0094-8373(2008)034[0229:COWVCI]2.0.CO;2 . ISSN 0094-8373 . S2CID 85880201 .
- ^ Там же (2004). п. 277 . Издательство Университета Индианы. ISBN 978-0-253-34282-9 .
- ^ Хансен Д.М., Донлан, СиДжей, Гриффитс, СиДжей, Кэмпбелл, К.Дж. (апрель 2010 г.). «Экологическая история и скрытый природоохранный потенциал: большие и гигантские черепахи как модель для замены таксонов» (PDF) . Экография . 33 (2): 272–284. Бибкод : 2010Экогр..33..272H . дои : 10.1111/j.1600-0587.2010.06305.x . Архивировано из оригинала (PDF) 24 июля 2011 года . Проверено 26 февраля 2011 г.
- ^ Сьоне А.Л., Тонни, Е.П., Сойбельзон, Л. (2003). «Разорванный зигзаг: позднекайнозойское вымирание крупных млекопитающих и черепах в Южной Америке» . Преподобный Муз. Аргентино Сиенс. Нат . Новая серия. 5 (1): 1–19. дои : 10.22179/REVMACN.5.26 . ISSN 1514-5158 .
- ^ Гордон И.Дж., Принс Х.Х., Мэллон Дж., Пук Л.Д., Миранда Э.Б., Старлинг-Манн С., ван дер Вал Р., Мур Б., Фоли В. (2019), Гордон И.Дж., Принс Х.Х. (ред.), «Экология просмотра и Выпас других таксонов позвоночных», «Экология просмотра и выпаса II» , Cham: Springer International Publishing, стр. 339–404, doi : 10.1007/978-3-030-25865-8_15 , ISBN 978-3-030-25865-8
- ^ Джаффе А.Л., Слейтер Дж.Дж., Альфаро М.Э. (23 августа 2011 г.). «Эволюция островного гигантизма и изменение размеров тела черепах и черепах» . Письма по биологии . 7 (4): 558–561. дои : 10.1098/rsbl.2010.1084 . ISSN 1744-9561 . ПМК 3130210 . ПМИД 21270022 .
- ^ Кадена Э.А., Линк А, Кук С.Б., Стройк Л.К., Ванегас А.Ф., Таллман М. (декабрь 2021 г.). «Новые сведения об анатомии и онтогенезе крупнейших вымерших пресноводных черепах» . Гелион . 7 (12): e08591. Бибкод : 2021Heliy...708591C . doi : 10.1016/j.heliyon.2021.e08591 . ISSN 2405-8440 . ПМЦ 8717240 . ПМИД 35005268 .
- ^ Свеннинг Дж.К., Лемуан Р.Т., Бергман Дж., Буитенверф Р., Ле Ру Э., Лундгрен Э., Мунги Н., Педерсен Р.О. (2024). «Позднечетвертичное вымирание мегафауны: закономерности, причины, экологические последствия и последствия для управления экосистемами в антропоцене» . Кембриджские призмы: вымирание . 2 . дои : 10.1017/доб.2024.4 . ISSN 2755-0958 .
- ^ Корлетт РТ (2006). «Вымирание мегафауны в тропической Азии» (PDF) . Тропинет . 17 (3): 1–3. Архивировано (PDF) из оригинала 4 марта 2016 г. Проверено 4 октября 2010 г.
- ^ Эдмидс Б. «Мегафауна — первые жертвы антропогенного вымирания» . megafauna.com . (книга, опубликованная в Интернете, с предисловием Пола С. Мартина ). Архивировано из оригинала 25 декабря 2014 г. Проверено 13 февраля 2020 г.
- ^ Jump up to: а б Мартин П.С. (2005). «Глава 6. Смертельная синкопа» . Сумерки мамонтов: вымирание ледникового периода и возрождение Америки . Издательство Калифорнийского университета . стр. 118–128. ISBN 978-0-520-23141-2 . OCLC 58055404 . Архивировано из оригинала 27 марта 2024 г. Проверено 11 ноября 2014 г.
- ^ Jump up to: а б Берни Д.А., Фланнери Т.Ф. (июль 2005 г.). «Пятьдесят тысячелетий катастрофических вымираний после контакта с человеком» (PDF) . Тенденции в экологии и эволюции . 20 (7): 395–401. дои : 10.1016/j.tree.2005.04.022 . ПМИД 16701402 . Архивировано из оригинала (PDF) 10 июня 2010 г. Проверено 11 ноября 2014 г.
- ^ Робертс Р.Г., Фланнери Т.Ф. , Эйлифф Л.К., Йошида Х., Олли Дж.М., Придо Дж.Дж., Ласлетт Г.М., Бэйнс А., Смит М.А., Джонс Р., Смит Б.Л. (08.06.2001). «Новый век последней австралийской мегафауны: вымирание по всему континенту около 46 000 лет назад» (PDF) . Наука . 292 (5523): 1888–1892. Бибкод : 2001Sci...292.1888R . дои : 10.1126/science.1060264 . ПМИД 11397939 . S2CID 45643228 . Архивировано (PDF) из оригинала 10 февраля 2019 г. Проверено 26 августа 2011 г.
- ^ Каллауэй Э (21 сентября 2016 г.). «В пещере хоббитов найдены человеческие останки». Природа . дои : 10.1038/nature.2016.20656 . S2CID 89272546 .
- ^ Даймонд Джей (13 августа 2008 г.). «Палеонтология: Последний гигантский кенгуру». Природа . 454 (7206): 835–836. Бибкод : 2008Natur.454..835D . дои : 10.1038/454835a . ПМИД 18704074 . S2CID 36583693 .
- ^ Терни К.С., Фланнери Т.Ф. , Робертс Р.Г., Рид С., Файфилд Л.К., Хайэм Т.Ф., Джейкобс З., Кемп Н., Колхун Э.А., Калин Р.М., Огл Н. (21 августа 2008 г.). «Поздно сохранившаяся мегафауна в Тасмании, Австралия, указывает на причастность человека к их исчезновению» . ПНАС . 105 (34): 12150–12153. Бибкод : 2008PNAS..10512150T . дои : 10.1073/pnas.0801360105 . ПМК 2527880 . ПМИД 18719103 .
- ^ Робертс Р., Джейкобс З. (октябрь 2008 г.). «Затерянные гиганты Тасмании» (PDF) . Австралазийская наука . 29 (9): 14–17. Архивировано из оригинала (PDF) 27 сентября 2011 г. Проверено 26 августа 2011 г.
- ^ Jump up to: а б Филд Дж., Роу С., Труман С.Н., Гарви Дж., Вятт-Спратт С. (8 февраля 2013 г.). «В поисках археологического следа вымирания австралийской мегафауны» . Четвертичный интернационал . Заселение последних новых миров: первая колонизация Сахула и Америки. 285 : 76–88. Бибкод : 2013QuInt.285...76F . дои : 10.1016/j.quaint.2011.04.013 . ISSN 1040-6182 . Архивировано из оригинала 18 декабря 2012 г.
- ^ Додсон Дж., Филд Дж.Х. (2018). «Что означает появление спор Sporormiella (Preussia) в австралийских окаменелостях?» . Журнал четвертичной науки . 33 (4): 380–392. Бибкод : 2018JQS....33..380D . дои : 10.1002/jqs.3020 . ISSN 1099-1417 . S2CID 133737405 . Архивировано из оригинала 14 февраля 2022 г.
- ^ Роу С., Филд Дж.Х., Арчер М., Грейсон Д.К., Прайс Дж.Дж., Луис Дж., Фейт Дж.Т., Уэбб Дж.Е., Дэвидсон И., Муни С.Д. (3 сентября 2013 г.). «Ответ Бруку и др.: Нет эмпирических доказательств чрезмерного истребления человеком мегафауны в Сахуле» . Труды Национальной академии наук . 110 (36): Е3369. Бибкод : 2013PNAS..110E3369W . дои : 10.1073/pnas.1310440110 . ISSN 0027-8424 . ПМК 3767508 . ПМИД 24137797 .
- ^ Дортч Дж., Каппер М., Грюн Р., Харпли Б., Ли К., Филд Дж. (01.08.2016). «Время и причина массовой гибели мегафауны в болоте Лансфилд, юго-восток Австралии» . Четвертичные научные обзоры . 145 : 161–182. Бибкод : 2016QSRv..145..161D . doi : 10.1016/j.quascirev.2016.05.042 . ISSN 0277-3791 . Архивировано из оригинала 27 марта 2024 г.
- ^ Роу С., Филд Дж.Х., Арчер М., Грейсон Д.К., Прайс Дж.Дж., Луис Дж., Фейт Дж.Т., Уэбб Дж.Е., Дэвидсон И., Муни С.Д. (28 мая 2013 г.). «Изменение климата лежит в основе дебатов по поводу исчезновения мегафауны в Сахуле (плейстоценовая Австралия-Новая Гвинея)» . Труды Национальной академии наук . 110 (22): 8777–8781. Бибкод : 2013PNAS..110.8777W . дои : 10.1073/pnas.1302698110 . ISSN 0027-8424 . ПМК 3670326 . ПМИД 23650401 .
- ^ Нортон С.Дж., Кондо, Ю., Оно, А., Чжан, Ю., Диаб, MC (23 мая 2009 г.). «Природа вымирания мегафауны во время перехода MIS 3–2 в Японии». Четвертичный интернационал . 211 (1–2): 113–122. Бибкод : 2010QuInt.211..113N . дои : 10.1016/j.quaint.2009.05.002 .
- ^ Фейт Дж.Т., Суровелл Т.А. (8 декабря 2009 г.). «Синхронное вымирание млекопитающих плейстоцена Северной Америки» . Труды Национальной академии наук . 106 (49): 20641–20645. Бибкод : 2009PNAS..10620641F . дои : 10.1073/pnas.0908153106 . ПМК 2791611 . ПМИД 19934040 .
- ^ Хейнс Дж. (2009). «Введение в том» . В Хейнсе Дж. (ред.). Вымирание американской мегафауны в конце плейстоцена . Палеобиология и палеоантропология позвоночных. Спрингер . стр. 1–20. дои : 10.1007/978-1-4020-8793-6_1 . ISBN 978-1-4020-8792-9 . [ постоянная мертвая ссылка ]
- ^ Фидель С. (2009). «Внезапные смерти: хронология терминального вымирания мегафауны плейстоцена». В Хейнсе Дж. (ред.). Вымирание американской мегафауны в конце плейстоцена . Палеобиология и палеоантропология позвоночных. Спрингер . стр. 21–37. дои : 10.1007/978-1-4020-8793-6_2 . ISBN 978-1-4020-8792-9 .
- ^ Симмонс АХ (1999). Вымирание фауны в островном обществе: охотники на карликовых бегемотов на Кипре . Междисциплинарный вклад в археологию. Издательство Kluwer Academic/Plenum . п. 382. дои : 10.1007/b109876 . ISBN 978-0-306-46088-3 . OCLC 41712246 . Архивировано из оригинала 27 марта 2024 г. Проверено 7 мая 2016 г.
- ^ Симмонс А.Х., Мандель, Р.Д. (декабрь 2007 г.). «Не такой уж новый свет: ответ Аммерману и Ноллеру». Мировая археология . 39 (4): 475–482. дои : 10.1080/00438240701676169 . JSTOR 40026143 . S2CID 161791746 .
- ^ Стедман Д.В. , Мартин П.С. , Макфи Р.Д., Джулл А.Дж., Макдональд Х.Г., Вудс Калифорния, Итурральде-Винент М., Ходжинс Г.В. (16 августа 2005 г.). «Асинхронное вымирание позднечетвертичных ленивцев на континентах и островах» . Учеб. Натл. акад. наук. США . 102 (33): 11763–11768. Бибкод : 2005PNAS..10211763S . дои : 10.1073/pnas.0502777102 . ПМК 1187974 . ПМИД 16085711 .
- ^ Кук С.Б., Давалос Л.М., Михайлив А.М., Терви С.Т., Апхэм Н.С. (2017). «Антропогенное вымирание доминирует над голоценовым сокращением численности млекопитающих Вест-Индии». Ежегодный обзор экологии, эволюции и систематики . 48 (1): 301–327. doi : 10.1146/annurev-ecolsys-110316-022754 . S2CID 90558542 .
- ^ Андерсон А., Сэнд К., Петчи Ф., Уорти Т.Д. (2010). «Вымирание фауны и проживание людей в Новой Каледонии: первоначальные результаты и последствия новых исследований в пещерах Пиндай». Журнал тихоокеанской археологии . 1 (1): 89–109. hdl : 10289/5404 .
- ^ Уайт А.В., Уорти, Т.Х., Хокинс, С., Бедфорд, С., Сприггс, М. (16 августа 2010 г.). «Мегафаунальные рогатые черепахи-мейоланииды дожили до первых поселений человека в Вануату, юго-западная часть Тихого океана» . Учеб. Натл. акад. наук. США . 107 (35): 15512–15516. Бибкод : 2010PNAS..10715512W . дои : 10.1073/pnas.1005780107 . ПМЦ 2932593 . ПМИД 20713711 .
- ^ Берни Д.А., Берни, Л.П., Годфри, Л.Р., Юнгерс, В.Л., Гудман, С.М., Райт, Х.Т., Джулл. AJT (июль 2004 г.). «Хронология позднего доисторического Мадагаскара». Журнал эволюции человека . 47 (1–2): 25–63. дои : 10.1016/j.jhevol.2004.05.005 . ПМИД 15288523 .
- ^ Холдэуэй Р.Н., Джейкомб, К. (24 марта 2000 г.). «Быстрое вымирание моа (Aves: Dinornithiformes): модель, тест и последствия». Наука . 287 (5461): 2250–2254. Бибкод : 2000Sci...287.2250H . дои : 10.1126/science.287.5461.2250 . ПМИД 10731144 .
- ^ Джану А. (апрель 2005 г.). «Обнаружение отдельных костей дронта ( Raphus cucullatus (L.), Aves, Columbiformes) в пещерных убежищах Маврикия подчеркивает факт хищничества человека с комментарием о статусе семейства Raphidae Wetmore, 1930». Анналы палеонтологии . 91 (2): 167–180. Бибкод : 2005AnPal..91..167J . дои : 10.1016/j.annpal.2004.12.002 .
- ^ Андерсон П.К. (июль 1995 г.). «Конкуренция, хищничество, а также эволюция и вымирание стеллеровой морской коровы Hydrodamalis gigas » . Наука о морских млекопитающих . 11 (3): 391–394. Бибкод : 1995MMamS..11..391A . дои : 10.1111/j.1748-7692.1995.tb00294.x . Архивировано из оригинала 11 мая 2011 г. Проверено 30 августа 2011 г.
- ^ Купер А., Терни С., Хьюэн К.А., Брук Б.В., Макдональд Х.Г., Брэдшоу СиДжей (23 июля 2015 г.). «Резкое потепление привело к смене мегафауны в позднем плейстоцене Голарктики» . Наука . 349 (6248): 602–6. Бибкод : 2015Sci...349..602C . дои : 10.1126/science.aac4315 . ПМИД 26250679 . S2CID 31686497 .
- ^ Мюллер Калифорнийский университет, Просс Дж., Цедакис ПК, Гэмбл С., Коттофф Ю., Шмидл Г., Вульф С., Кристанис К. (февраль 2011 г.). «Роль климата в расселении современного человека в Европу». Четвертичные научные обзоры . 30 (3–4): 273–279. Бибкод : 2011QSRv...30..273M . doi : 10.1016/j.quascirev.2010.11.016 .
- ^ Бьелло Д. (22 марта 2012 г.). «Большое убийство, а не большое охлаждение, прикончили гигантских кенгуру» . Научные американские новости . Архивировано из оригинала 23 марта 2012 г. Проверено 25 марта 2012 г.
- ^ МакГлоун М (23 марта 2012 г.). «Охотники сделали это». Наука . 335 (6075): 1452–1453. Бибкод : 2012Sci...335.1452M . дои : 10.1126/science.1220176 . ПМИД 22442471 . S2CID 36914192 .
- ^ Jump up to: а б с Правило С, Брук, Б.В., Хаберле, С.Г., Терни, КСМ, Кершоу, АП (23 марта 2012 г.). «Последствия вымирания мегафауны: трансформация экосистемы в плейстоценовой Австралии». Наука . 335 (6075): 1483–1486. Бибкод : 2012Sci...335.1483R . дои : 10.1126/science.1214261 . ПМИД 22442481 . S2CID 26675232 .
- ^ Jump up to: а б Джонсон С.Н., Элрой Дж., Битон Н.Дж., Берд М.И., Брук Б.В., Купер А., Гиллеспи Р., Эррандо-Перес С., Джейкобс З., Миллер Г.Х., Придо Дж.Дж., Робертс Р.Г., Родригес-Рей М., Салтре Ф., Терни К.С., Брэдшоу CJ (10 февраля 2016 г.). «Что стало причиной исчезновения плейстоценовой мегафауны Сахула?» . Труды Королевского общества B: Биологические науки . 283 (1824): 20152399. doi : 10.1098/rspb.2015.2399 . ПМК 4760161 . ПМИД 26865301 .
- ^ Миллер Г.Х., Маги Дж.В., Джонсон Б.Дж., Фогель М.Л., Спунер Н.А., Маккалок М.Т., Эйлифф Л.К. (08 января 1999 г.). «Плейстоценовое вымирание Genyornis newtoni : воздействие человека на австралийскую мегафауну». Наука . 283 (5399): 205–208. дои : 10.1126/science.283.5399.205 . ПМИД 9880249 .
- ^ Миллер Дж., Маги Дж., Смит М., Спунер Н., Бейнс А., Леман С., Фогель М., Джонстон Х., Уильямс Д., Кларк П., Флориан С., Холст Р., ДеВогель С. (29 января 2016 г.). «Человеческое хищничество способствовало исчезновению австралийской мегафауны Genyornis newtoni ~47 тыс. лет назад» . Природные коммуникации . 7 : 10496. Бибкод : 2016NatCo...710496M . дои : 10.1038/ncomms10496 . ПМК 4740177 . ПМИД 26823193 .
- ^ Джонсон С. (20 ноября 2009 г.). «Упадок и падение мегафауны». Наука . 326 (5956): 1072–1073. Бибкод : 2009Sci...326.1072J . дои : 10.1126/science.1182770 . ПМИД 19965418 . S2CID 206523763 .
- ^ Джилл Дж.Л., Уильямс Дж.В., Джексон С.Т., Лайнингер К.Б., Робинсон Г.С. (20 ноября 2009 г.). «Коллапс плейстоценовой мегафауны, новые растительные сообщества и усиление пожарных режимов в Северной Америке» (PDF) . Наука . 326 (5956): 1100–1103. Бибкод : 2009Sci...326.1100G . дои : 10.1126/science.1179504 . ПМИД 19965426 . S2CID 206522597 . Архивировано (PDF) из оригинала 22 сентября 2017 г. Проверено 9 ноября 2018 г.
- ^ Фидал С (2009). «Внезапные смерти: хронология терминального вымирания мегафауны плейстоцена». В Хейнсе Дж. (ред.). Вымирание американской мегафауны в конце плейстоцена . Палеобиология и палеоантропология позвоночных. Спрингер . стр. 21–37. дои : 10.1007/978-1-4020-8793-6_2 . ISBN 978-1-4020-8792-9 .
- ^ Мартин П.С. (2005). «Глава 4. Наземные ленивцы в домашних условиях» . Сумерки мамонтов: вымирание ледникового периода и возрождение Америки . Издательство Калифорнийского университета . стр. 78–99. ISBN 978-0-520-23141-2 . OCLC 58055404 . Архивировано из оригинала 27 марта 2024 г. Проверено 11 ноября 2014 г.
- ^ Мильман О. (6 февраля 2019 г.). «Исследование показывает, что убийство крупных видов подталкивает их к вымиранию» . Хранитель . Архивировано из оригинала 7 февраля 2019 года . Проверено 13 февраля 2019 г.
- ^ Риппл У.Дж. и др. (2019). «Мы едим мировую мегафауну до полного исчезновения?» . Письма о сохранении . 12 (3): e12627. Бибкод : 2019ConL...12E2627R . дои : 10.1111/conl.12627 .
- ^ Элрой Дж. (12 августа 2008 г.). «Динамика возникновения и исчезновения морской летописи окаменелостей» . ПНАС . 105 Приложение 1 (Дополнение_1): 11536–11542. Бибкод : 2008PNAS..10511536A . дои : 10.1073/pnas.0802597105 . ПМК 2556405 . ПМИД 18695240 .
- ^ Д'Ондт С (1 декабря 2005 г.). «Последствия мелового/палеогенового массового вымирания для морских экосистем» . Ежегодный обзор экологии, эволюции и систематики . 36 (1): 295–317. doi : 10.1146/annurev.ecolsys.35.021103.105715 . ISSN 1543-592X .
- ^ Jump up to: а б Малхи Ю., Даути С.Э., Галетти М., Смит Ф.А., Свеннинг Дж.К., Терборг Дж.В. (26 января 2016 г.). «Мегафауна и экосистема функционируют от плейстоцена до антропоцена» . Труды Национальной академии наук . 113 (4): 838–846. дои : 10.1073/pnas.1502540113 . ISSN 0027-8424 . ПМЦ 4743772 . ПМИД 26811442 .
- ^ Джонсон С. (22 июля 2009 г.). «Экологические последствия позднечетвертичного вымирания мегафауны» . Труды Королевского общества B: Биологические науки . 276 (1667): 2509–2519. дои : 10.1098/рспб.2008.1921 . ISSN 0962-8452 . ПМЦ 2684593 . ПМИД 19324773 .
- ^ Мунги Н.А., Джала Ю.В., Куреши К., Ле Ру Э., Свеннинг Дж.К. (октябрь 2023 г.). «Мегагербоядные животные обеспечивают биотическую устойчивость к доминированию чужеродных растений» . Экология и эволюция природы . 7 (10): 1645–1653. дои : 10.1038/s41559-023-02181-y . ISSN 2397-334X .
- ^ Вольф А., Даути С.Э., Малхи Ю. (2013). «Боковая диффузия питательных веществ травоядными млекопитающими в наземных экосистемах» . ПЛОС ОДИН . 8 (8): е71352. Бибкод : 2013PLoSO...871352W . дои : 10.1371/journal.pone.0071352 . ПМЦ 3739793 . ПМИД 23951141 .
- ^ Маршалл М. (11 августа 2013 г.). «Экосистемы все еще ощущают боль древнего вымирания» . Новый учёный . Архивировано из оригинала 4 июля 2015 г. Проверено 12 августа 2013 г.
- ^ Jump up to: а б Даути CE, Вольф А, Малхи Ю (11 августа 2013 г.). «Наследие вымирания плейстоценовой мегафауны на доступность питательных веществ в Амазонии». Природа Геонауки . 6 (9): 761–764. Бибкод : 2013NatGe...6..761D . дои : 10.1038/ngeo1895 .
- ^ Роман Дж., Маккарти Дж. (2010). «Китовый насос: морские млекопитающие повышают первичную продуктивность в прибрежном бассейне» . ПЛОС ОДИН . 5 (10): е13255. Бибкод : 2010PLoSO...513255R . дои : 10.1371/journal.pone.0013255 . ПМЦ 2952594 . ПМИД 20949007 .
- ^ Jump up to: а б с Уилкинсон Д.М., Нисбет, Э.Г., Ракстон, Г.Д. (08 мая 2012 г.). «Мог ли метан, производимый динозаврами-зауроподами, способствовать потеплению мезозойского климата?» . Современная биология . 22 (9): Р292–Р293. Бибкод : 2012CBio...22.R292W . дои : 10.1016/j.cub.2012.03.042 . ПМИД 22575462 .
- ^ «Газы динозавров «согрели Землю» » . Новости природы Би-би-си . 07.05.2012. Архивировано из оригинала 1 декабря 2015 г. Проверено 8 мая 2012 г.
- ^ Jump up to: а б с Смит Ф.А., Эллиот, С.М., Лайонс, СК (23 мая 2010 г.). «Выбросы метана вымершей мегафауны». Природа Геонауки . 3 (6): 374–375. Бибкод : 2010NatGe...3..374S . дои : 10.1038/ngeo877 .
- ^ Келлихер Ф.М., Кларк, Х. (15 марта 2010 г.). «Выбросы метана от бизонов — историческая оценка поголовья Великих равнин Северной Америки». Сельскохозяйственная и лесная метеорология . 150 (3): 473–577. Бибкод : 2010AgFM..150..473K . дои : 10.1016/j.agrformet.2009.11.019 .
- ^ Цубамото Т (2012). «Оценка массы тела по таранной кости у млекопитающих». Acta Palaeontologica Polonica : 259–265. дои : 10.4202/app.2011.0067 . S2CID 54686160 .
- ^ Стюарт Эй Джей (ноябрь 1991 г.). «Вымирание млекопитающих в позднем плейстоцене Северной Евразии и Северной Америки». Биологические обзоры . 66 (4): 453–562. дои : 10.1111/j.1469-185X.1991.tb01149.x . ПМИД 1801948 . S2CID 41295526 .
- ^ Сэмпл, Ян (19 февраля 2010 г.). «Большая белая акула находится под большей угрозой исчезновения, чем тигр, — утверждает учёный» . Хранитель . Архивировано из оригинала 9 сентября 2013 года . Проверено 14 августа 2013 г.