Мегалодон
| Мегалодон Временной диапазон:
Ранний миоцен – ранний плиоцен , ок. | |
|---|---|

| |
| Модель челюстей мегалодона в Музее науки Теллуса | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Хондриктиес |
| Подкласс: | Elasmobranchii |
| Подразделение: | Селахиморфа |
| Заказ: | Ламнообразные |
| Семья: | † Отодонтиды |
| Род: | † Отодус |
| Разновидность: | † О. мегалодон
|
| Биномиальное имя | |
| † Отодус мегалодон | |
| Синонимы [ 3 ] [ 4 ] [ 5 ] [ 6 ] [ 7 ] | |
|
Список синонимов | |
Otodus megalodon ( / ˈ m ɛ ɡ əl ə d ɒ n / MEG -əl-ə-don ; означает «большой зуб»), широко известный как мегалодон , — вымерший вид гигантской макрельевой акулы , живший примерно от 23 до 3,6 миллионов человек. лет назад (Mya), от эпохи раннего миоцена до плиоцена . O. megalodon ранее считался членом семейства Lamnidae и близким родственником большой белой акулы ( Carcharodon carcharias ), но был переклассифицирован в вымершее семейство Otodontidae , которое отделилось от большой белой акулы в раннем меловом периоде .
Хотя мегалодон считается одним из самых крупных и могущественных хищников , когда-либо живших на свете, он известен только по фрагментарным останкам, а его внешний вид и максимальный размер неизвестны. Ученые расходятся во мнениях относительно того, напоминала ли бы она более коренастую версию большой белой акулы ( Carcharodon carcharias ), гигантскую акулу ( Cetorhinus maximus ) или песчаную тигровую акулу ( Carcharias taurus ). Самая последняя оценка с наименьшим диапазоном ошибок предполагает, что максимальная длина составляет до 20,3 метра (67 футов), хотя модальная длина оценивается в 10,5 метра (34 фута). Их зубы были толстыми и крепкими, предназначенными для хватания добычи и разрушения костей, а большие челюсти могли оказывать силу укуса от 108 500 до 182 200 ньютонов (от 24 390 до 40 960 фунтов силы).
Мегалодон, вероятно, оказал большое влияние на структуру морских сообществ . Летопись окаменелостей указывает на то, что он имел космополитическое распространение . Вероятно, его целью была крупная добыча, такая как киты , тюлени и морские черепахи . Молодь обитала в теплых прибрежных водах и питалась рыбой и мелкими китами. В отличие от большой белой белухи, которая нападает на добычу с мягкой нижней стороны, мегалодон, вероятно, использовал свои сильные челюсти, чтобы прорваться через грудную полость и проткнуть сердце и легкие своей добычи.
Животное столкнулось с конкуренцией со стороны китообразных, питающихся китами , таких как Ливиатан и другие макрорапториальные кашалоты и, возможно, более мелкие предки косаток ( Orcinus ). Поскольку акула предпочитала более теплые воды, считается, что охлаждение океана, связанное с наступлением ледниковых периодов , в сочетании с понижением уровня моря и, как следствие, потерей подходящих мест нагула, также могло способствовать ее сокращению. Сокращение разнообразия усатых китов и сдвиг их распространения в сторону полярных регионов, возможно, привели к сокращению основного источника пищи мегалодона. Вымирание акул совпадает с тенденцией к гигантизму среди усатых китов.
Классификация
Донаучная и ранняя история исследований


Зубы мегалодона раскапывали и использовали с древних времен. Они были ценным артефактом среди доколумбовых культур Америки из -за своих больших размеров и зазубренных лезвий, из которых они были преобразованы в наконечники снарядов , ножи, украшения и похоронные принадлежности. [ 8 ] [ 9 ] По крайней мере, некоторые из них, такие как панамские общества Ситио Конте , похоже, использовали их в первую очередь в церемониальных целях. [ 9 ] Горное дело [ 10 ] Зубы мегалодона у алгонкинских народов в Чесапикском заливе и их избирательная торговля с культурой Адены в Огайо произошли еще в 430 году до нашей эры . [ 8 ] Самый ранний письменный отчет о зубах мегалодона был сделан Плинием Старшим в 73 г. н.э. томе « Historia Naturalis» , который описал их как напоминающие окаменевшие человеческие языки , которые, как полагали римские фольклористы, падали с неба во время лунных затмений , и назвал их « glossopetrae» («камни языка»). "). [ 11 ] традиции XII века считалось, что предполагаемые языки Позднее в мальтийской принадлежали змеям, которых апостол Павел превратил в камень во время кораблекрушения наделил их противоядием . , и святой [ 12 ] Glossopetrae вновь появились по всей Европе в литературе конца 13-16 веков, и им приписывались более сверхъестественные свойства, способные излечивать более широкий спектр ядов . Использование зубов мегалодона для этой цели получило широкое распространение среди средневековой и ренессансной знати, которая превратила их в защитные амулеты и столовую посуду для якобы детоксикации отравленных жидкостей или тел, соприкоснувшихся с камнями. К 16 веку зубы непосредственно употреблялись в пищу в составе камней Гоа европейского производства . [ 11 ]


Истинная природа глоссопетр как акульих зубов некоторые придерживались, по крайней мере, с 1554 года, когда космограф Андре Теве описал это как слухи, хотя и не верил в это. Самый ранний научный аргумент в пользу этой точки зрения был выдвинут итальянским натуралистом Фабио Колонной , который в 1616 году опубликовал иллюстрацию зуба мальтийского мегалодона рядом с зубом большой белой акулы и отметил их поразительное сходство. Он утверждал, что первый и его подобия были не окаменевшими змеиными языками, а на самом деле зубами подобных акул, выброшенных на берег. Колонна поддержал этот тезис экспериментом по сжиганию образцов глоссопетр , в котором он обнаружил остатки углерода, которые он интерпретировал как доказательство органического происхождения. [ 12 ] Однако интерпретация камней как зубов акулы осталась широко непринятой. Отчасти это произошло из-за невозможности объяснить, почему некоторые из них находятся вдали от моря. [ 13 ] Аргумент о зубах акулы был снова поднят в академических кругах в конце 17 века английскими учёными Робертом Гуком , Джоном Рэем и датским натуралистом Нильсом Стинсеном (латинизированный Николас Стено ). [ 14 ] Аргумент Стинсена, в частности, наиболее известен как вывод из его вскрытия головы большой белой акулы, пойманной в 1666 году. В его отчете 1667 года были изображены гравюры с изображением головы акулы и зубов мегалодона, которые стали особенно знаковыми. Однако иллюстрированная голова на самом деле не была той головой, которую препарировал Стинсен, как и проиллюстрированные им ископаемые зубы. Обе гравюры были первоначально заказаны в 1590-х годах папским врачом Микеле Меркати , у которого также была голова большой белой лошади, для его книги «Металлотека» . Работа осталась неопубликованной во времена Стинсена из-за преждевременной смерти Меркати, и первый повторно использовал две иллюстрации по предложению Карло Роберто Дати , который считал, что изображение настоящей расчлененной акулы не подходит для читателей. [ 15 ] Стинсен также выдвинулся в качестве пионера стратиграфического объяснения того, как подобные камни появились дальше вглубь страны. Он заметил, что слои горных пород с зубами мегалодона содержат морские отложения, и выдвинул гипотезу, что эти слои связаны с периодом наводнения , которое позже было покрыто земными слоями и поднято геологической активностью. [ 13 ]

Швейцарский натуралист Луи Агассис дал мегалодону научное название в своей плодотворной работе 1833–1843 годов «Recherches sur les Poissons Foissons» («Исследование ископаемых рыб»). Он назвал его Carcharias megalodon на иллюстрации голотипа и дополнительных зубов 1835 года, родственной современной песчаной тигровой акуле . [ 1 ] [ 2 ] Видовое название представляет собой комбинацию древнегреческих odṓn слов μεγάλος ( megálos , что означает «большой») и ὀδών ( , что означает «зуб»). [ 16 ] [ 17 ] комбинированное значение «большой зуб». Агассис упомянул это название еще в 1832 году, но, поскольку образцы не упоминались, они не являются таксономически признанными видами использования . [ 2 ] Официальное описание вида было опубликовано в книге 1843 года, где Агассис изменил название на Carcharodon megalodon , поскольку его зубы были слишком большими для первого рода и больше напоминали большую белую акулу. [ 1 ] Он также ошибочно определил несколько зубов мегалодона как принадлежащие к дополнительным видам, в конечном итоге названным Carcharodon rectidens , Carcharodon subauriculatus , Carcharodon Productus и Carcharodon Polygurus . [ 1 ] [ 18 ] Поскольку Carcharodon megalodon впервые появился на иллюстрации 1835 года, остальные названия считаются младшими синонимами по принципу приоритета . [ 2 ] [ 18 ]
Эволюция

Хотя самые ранние останки мегалодона были зарегистрированы в позднем олигоцене , около 28 миллионов лет назад (млн лет назад), [ 19 ] [ 20 ] существуют разногласия относительно того, когда он появился: даты его появления варьируются от 16 млн лет назад. [ 21 ] Считалось, что мегалодон вымер примерно в конце плиоцена , около 2,6 млн лет назад; [ 21 ] [ 22 ] Заявления о зубах плейстоценового мегалодона возрастом менее 2,6 миллиона лет считаются ненадежными. [ 22 ] По оценкам 2019 года, дата вымирания переносится на более ранний период плиоцена, 3,6 млн лет назад. [ 23 ]
Мегалодон считается членом семейства Otodontidae, рода Otodus , в отличие от его предыдущей классификации в Lamnidae, рода Carcharodon . [ 21 ] [ 24 ] [ 22 ] [ 7 ] [ 25 ] Отнесение Мегалодона к Кархародону произошло из-за сходства зубов с большой белой акулой, однако большинство авторов считают, что это связано с конвергентной эволюцией . В этой модели большая белая акула более тесно связана с вымершим широкозубым мако ( Isurus hastalis ), чем с мегалодоном, о чем свидетельствует более похожее расположение зубов у этих двух акул; Зубы мегалодона имеют гораздо более тонкие зазубрины, чем зубы большой белой акулы. Большая белая акула более близка к акулам мако ( Isurus spp.), имея общего предка около 4 млн лет назад. [ 26 ] [ 27 ] Сторонники первой модели, согласно которой мегалодон и большая белая акула более тесно связаны, утверждают, что различия между их зубными рядами незначительны и неясны. [ 28 ] : 23–25

Род Carcharocles включает четыре вида: C. auriculatus , C. angustidens , C. chubutensis и C. megalodon . [ 29 ] : 30–31 Эволюция этой линии характеризуется увеличением зубцов, расширением коронки, развитием более треугольной формы и исчезновением боковых бугорков . [ 29 ] : 28–31 [ 30 ] Эволюция морфологии зубов отражает сдвиг в тактике хищничества от разрывающе-хватательного укуса к режущему, что, вероятно, отражает сдвиг в выборе добычи от рыб к китообразным. [ 31 ] Боковые бугорки были окончательно утеряны в результате постепенного процесса, который занял примерно 12 миллионов лет во время перехода от C. chubutensis к C. megalodon . [ 31 ] Род был предложен Д.С. Джорданом и Х. Ганнибалом в 1923 году как содержащий C. auriculatus . В 1980-х годах мегалодон был приписан к Кархароклу . [ 26 ] [ 29 ] : 30 род Procarcharodon Эдгаром Казье был выделен До этого, в 1960 году, французским ихтиологом , который включал этих четырех акул и считался отдельным от большой белой акулы. оно считается младшим синонимом Кархарокла С тех пор . [ 29 ] : 30 Род Palaeocarcharodon был выделен рядом с Procarcharodon, чтобы представить начало линии и, в модели, согласно которой мегалодон и большая белая акула тесно связаны, их последнего общего предка. считают, что это эволюционный тупик и не связанный с акулами Кархарокла . Авторы, отвергающие эту модель, [ 28 ] : 70

Другая модель эволюции этого рода, также предложенная Казье в 1960 году, заключается в том, что прямым предком кархароклов является акула Otodus obliquus , жившая с палеоцена по миоценовые эпохи, от 60 до 13 млн лет назад. [ 27 ] [ 30 ] Род Otodus в конечном итоге произошел от Кретоламны , акулы мелового периода. [ 7 ] [ 32 ] В этой модели O. obliquus эволюционировал в O. aksuaticus , который эволюционировал в C. auriculatus , а затем в C. angustidens , а затем в C. chubutensis , а затем, наконец, в C. megalodon .
Другая модель эволюции кархароклов , предложенная в 2001 году палеонтологом Майклом Бентоном , заключается в том, что три других вида на самом деле представляют собой единый вид акул, который постепенно менялся с течением времени между палеоценом и плиоценом, превращая его в хроновид . [ 29 ] : 17 [ 20 ] [ 33 ] Некоторые авторы полагают, что C. auriculatus , C. angustidens и C. chubutensis следует отнести к одному виду рода Otodus , оставив C. megalodon единственным представителем Carcharocles . [ 20 ] [ 34 ]
Род Carcharocles может быть недействительным, а акула может фактически принадлежать к роду Otodus , что делает ее Otodus megalodon . Исследование палеогеновых акул, проведенное Анри Каппеттой в 1974 году, выделило подрод Megaselachus , классифицировав акулу как Otodus ( Megaselachus ) megalodon , наряду с O. (M.) chubutensis . [ 5 ] Обзор Chondrichthyes 2006 года возвел Megaselachus в род и классифицировал акул как Megaselachus megalodon и M. chubutensis . Открытие окаменелостей, отнесенных к роду Megalolamna, в 2016 году привело к переоценке Otodus , которая пришла к выводу, что он парафилетичен , то есть состоит из последнего общего предка , но не включает всех своих потомков. Включение акул Carcharocles в Otodus сделало бы его монофилетическим , а сестринской кладой будет Megalolamna . [ 7 ]
Кладограмма ниже представляет гипотетические отношения между мегалодоном и другими акулами, включая большую белую акулу. Модифицировано из Shimada et al. (2016), [ 7 ] Эрет и др. (2009), [ 27 ] и результаты Сиверссона и соавт. (2015). [ 32 ]
Биология
Появление

Одна из интерпретаций появления мегалодона заключалась в том, что это была крупная на вид акула, которая, возможно, имела телосложение, похожее на большую белую акулу. Челюсти могли быть тупее и шире, чем у большой белой, плавники также были схожей формы, хотя и толще из-за размера. Возможно, у него были маленькие, глубоко посаженные глаза, похожие на свиноглазые. [ 35 ]
Другая интерпретация состоит в том, что мегалодон имел сходство с китовой акулой ( Rhincodon typus ) или гигантской акулой ( Cetorhinus maximus ). Хвостовой плавник имел форму полумесяца, анальный плавник и второй спинной плавник были небольшими, а ) должен был присутствовать хвостовой киль по обе стороны от хвостового плавника (на хвостовом стебле . Такое строение характерно для других крупных водных животных, таких как киты, тунцы и другие акулы, для уменьшения сопротивления во время плавания. Форма головы может различаться у разных видов, поскольку большинство приспособлений, снижающих сопротивление, расположены ближе к хвостовой части животного. [ 29 ] : 35–36
Один связанный набор останков мегалодона был обнаружен с плакоидными чешуйками, максимальная ширина которых составляет от 0,3 до 0,8 мм (от 0,012 до 0,031 дюйма) и с широко расположенными килями. [ 36 ]
Размер


Из-за фрагментарных останков было сделано много противоречивых оценок размера мегалодона, поскольку их можно получить только по ископаемым зубам и позвонкам. [ 37 ] : 87 [ 38 ] Большая белая акула легла в основу реконструкции и оценки размеров, поскольку считается лучшим аналогом мегалодона. На основе сравнения зубов и позвонков мегалодона с зубами и позвонками большой белой было разработано несколько методов оценки общей длины. [ 35 ] [ 39 ] [ 40 ] [ 25 ]
Оценки размера мегалодона варьируются в зависимости от используемого метода: максимальная общая длина составляет 14,2–20,3 метра (47–67 футов). [ 35 ] [ 40 ] [ 25 ] Исследование 2015 года оценило среднюю общую длину тела в 10,5 метров (34 фута), рассчитанную на основе 544 зубов мегалодона, обнаруженных на протяжении геологического времени и в географическом пространстве, включая взрослых и молодых особей. [ 41 ] [ 24 ] Для сравнения, большие белые акулы обычно имеют длину около 6 метров (20 футов), при этом в некоторых спорных сообщениях говорится о более крупных размерах. [ 42 ] [ 43 ] [ 35 ] Китовая акула - самая крупная из ныне живущих рыб: сообщается, что одна крупная самка имела прекаудальную длину 15 метров (49 футов) и предполагаемую общую длину 18,8 метра (62 фута). [ 42 ] [ 44 ] Вполне возможно, что разные популяции мегалодонов по всему миру имели разные размеры тела и поведение из-за разного экологического давления. [ 24 ] Мегалодон считается самой крупной хищной акулой, которая когда-либо жила. [ 35 ]
«C. megalodon длиной около 16 метров весил бы около 48 метрических тонн (53 тонны). 17-метровый (56-футовый) C. megalodon весил бы около 59 метрических тонн (65 тонн), а 20,3-метровый (67 футов) монстр имел бы максимальную массу в 103 метрических тонны (114 тонн)». [ 45 ]
В своей книге 2015 года « История жизни в 25 окаменелостях: рассказы о бесстрашных охотниках за ископаемыми и чудесах эволюции » Дональд Протеро предложил оценки массы тела для разных особей разной длины путем экстраполяции от центров позвонков на основе размеров великих белый, [ 45 ] методология, также использованная в исследовании 2008 года, поддерживает оценку максимальной массы. [ 46 ]
В 2020 году Купер и его коллеги реконструировали 2D-модель мегалодона на основе размеров всех современных акул-ламнид и предположили, что мегалодон длиной 16 метров (52 фута) имел голову длиной 4,65 м (15,3 фута), длину 1,41 м. (4 фута 8 дюймов) в высоту, спинной плавник высотой 1,62 м (5 футов 4 дюйма), грудные плавники длиной 3,08 м (10 футов 1 дюйм) и хвостовой плавник высотой 3,85 м (12 футов 8 дюймов). [ 47 ] В 2022 году Купер и его коллеги также реконструировали 3D-модель на той же основе, что и исследование 2020 года, в результате чего оценка массы тела мегалодона длиной 16 метров (52 фута) составила 61,56 т (67,86 коротких тонн; 60,59 длинных тонн). выше предыдущих оценок); Для экстраполяции был использован образец позвоночника под названием IRSNB P 9893 (ранее IRSNB 3121), принадлежавший 46-летнему человеку из Бельгии . Особи такого размера потребовалось бы 98 175 ккал в день, что в 20 раз больше, чем требуется взрослой белой белке. [ 48 ]
Взрослый самец мегалодона мог иметь массу тела от 12,6 до 33,9 т (от 13,9 до 37,4 коротких тонн; от 12,4 до 33,4 длинных тонн), а взрослые самки могли иметь массу тела от 27,4 до 59,4 т (от 30,2 до 65,5 коротких тонн; от 27,0 до 58,5 длинных тонн). ), предполагая, что длина самцов может составлять от 10,5 до 14,3 метров (от 34 до 47 футов), а самок - от 13,3 до 17 метров (от 44 до 56 футов). [ 35 ]
Исследование 2015 года, связывающее размер акулы и типичную скорость плавания, показало, что мегалодон обычно плавал со скоростью 18 километров в час (11 миль в час) – при условии, что масса его тела обычно составляла 48 т (53 коротких тонны; 47 длинных тонн) – что согласуется с другие водные существа такого же размера, такие как финвал ( Balaenoptera physalus ), который обычно путешествует со скоростью от 14,5 до 21,5 км/ч (от 9,0 до 13,4 миль в час). [ 49 ] В 2022 году Купер и его коллеги преобразовали эти расчеты в относительную крейсерскую скорость (длину тела в секунду), в результате чего средняя абсолютная крейсерская скорость составила 5 километров в час (3,1 мили в час), а средняя относительная крейсерская скорость составила 0,09 длины тела в секунду для мегалодон длиной 16 метров (52 фута); авторы обнаружили, что их средняя абсолютная крейсерская скорость выше, чем у любых современных акул-ламнид, а их средняя относительная крейсерская скорость ниже, что соответствует предыдущим оценкам. [ 48 ]

Его большой размер мог быть обусловлен климатическими факторами и обилием крупной добычи, а также, возможно, на него повлияла эволюция региональной эндотермии (мезотермии), которая увеличила бы скорость его метаболизма и скорость плавания. Считалось, что отодонтидные акулы были эктотермными животными , поэтому на этом основании мегалодон был бы экзотермным. Однако самые крупные современные эктотермные акулы, такие как китовая акула, являются фильтраторами, а ламниды являются региональными эндотермами, что предполагает некоторую метаболическую корреляцию с хищническим образом жизни. Эти соображения, а также данные по изотопам кислорода в зубах и необходимость более высоких скоростей плавания у макрохищников эндотермической добычи, чем позволяет эктотермия, предполагают, что отодонтиды, включая мегалодона, вероятно, были региональными эндотермами. [ 50 ]
В 2020 году Шимада и его коллеги предположили, что большой размер вызван внутриутробным каннибализмом , когда более крупный плод поедает меньший плод, в результате чего плод становится все больше и больше, что требует от матери достижения еще большего размера, а также потребности в калориях, что способствовало бы увеличению размера плода. эндотермия. Самцам пришлось бы не отставать от самки по размеру, чтобы эффективно совокупляться (что, вероятно, включало в себя захват самки с помощью застежек , как у современных хрящевых рыб). [ 51 ]
Самая высокая оценка
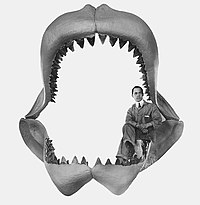
Первая попытка реконструировать челюсть мегалодона была предпринята Бэшфордом Дином в 1909 году и выставлена в Американском музее естественной истории . Судя по размерам этой реконструкции челюсти, было высказано предположение, что длина мегалодона могла достигать 30 метров (98 футов). Дин переоценил размер хрящей на обеих челюстях, из-за чего они оказались слишком высокими. [ 52 ] [ 53 ]

Джон Э. Рэндалл В 1973 году ихтиолог использовал высоту эмали (вертикальное расстояние лезвия от основания эмалевой части зуба до его кончика) для измерения длины акулы, получив максимальную длину около 13 метров (43 фута). [ 54 ] Однако высота зубной эмали не обязательно увеличивается пропорционально общей длине животного. [ 28 ] : 99
В 1994 году морские биологи Патрик Дж. Шембри и Стивен Папсон высказали мнение, что общая длина O. megalodon могла достигать примерно 24–25 метров (от 79 до 82 футов). [ 55 ] [ 56 ]
В 1996 году исследователи акул Майкл Д. Готфрид, Леонард Компаньо и С. Кертис Боуман предложили линейную зависимость между общей длиной большой белой акулы и высотой самого большого верхнего переднего зуба. Предлагаемое соотношение: общая длина в метрах = - (0,096) × [максимальная высота UA ( мм )]-(0,22). [ 57 ] [ 35 ] Используя это уравнение регрессии высоты зуба, авторы оценили общую длину в 15,9 метра (52 фута) на основе высоты зуба 16,8 сантиметра (6,6 дюйма), что авторы считали консервативной максимальной оценкой. Они также сравнили соотношение высоты зубов и общей длины крупных самок больших белых особей с самым большим зубом мегалодона. Самка большой белой самки длиной 6 метров (20 футов), которую авторы считали самой большой «достаточно заслуживающей доверия» общей длиной, по оценкам составила 16,8 метра (55 футов). Однако, основываясь на сообщениях о самой крупной самке большой белой белухи (7,1 метра (23 фута)), максимальная оценка составила 20,2 метра (66 футов). [ 35 ]
В 2002 году исследователь акул Клиффорд Джеремия предположил, что общая длина пропорциональна ширине корня верхнего переднего зуба. Он утверждал, что на каждый 1 сантиметр (0,39 дюйма) ширины корня приходится примерно 1,4 метра (4,6 фута) длины акулы. Иеремия отметил, что периметр челюсти акулы прямо пропорционален ее общей длине, причем инструментом для оценки периметра челюсти является ширина корней самых крупных зубов. Самый большой зуб, принадлежавший Иеремии, имел ширину корня около 12 сантиметров (4,7 дюйма), что давало общую длину 16,5 метра (54 фута). [ 29 ] : 88
В 2002 году палеонтолог Кеншу Шимада из Университета ДеПола после проведения анатомического анализа нескольких образцов предложил линейную зависимость между высотой коронки зуба и общей длиной, что позволило использовать зуб любого размера. Шимада заявил, что ранее предложенные методы были основаны на менее надежной оценке зубной гомологии мегалодона и большой белой акулы, и что скорость роста между коронкой и корнем не является изометрической , что он учел в своей модели. Используя эту модель, общая длина верхнего переднего зуба Готфрида и его коллег составила 15 метров (49 футов). [ 58 ] Среди нескольких экземпляров, найденных в формации Гатун в Панаме, другие исследователи использовали один верхний боковой зуб, чтобы с помощью этого метода получить оценку общей длины в 17,9 метра (59 футов). [ 34 ] [ 59 ]
В 2019 году Шимада вновь обратился к размеру мегалодона и не рекомендовал использовать для оценок непередние зубы, отметив, что точное положение изолированных непередних зубов определить сложно. Шимада предоставил оценку максимальной общей длины, используя самые большие передние зубы, имеющиеся в музеях. Зуб с самой высокой высотой коронки, известной Шимаде, NSM PV-19896, имел общую длину 14,2 метра (47 футов). Зуб с самой высокой общей высотой, FMNH PF 11306, имел высоту 16,8 см (6,6 дюйма). Однако Шимада повторно измерил зуб и обнаружил, что его размер на самом деле составляет 16,2 сантиметра (6,4 дюйма). Использование уравнения регрессии зубов общей высоты, предложенного Готфридом и его коллегами, дало оценку 15,3 метра (50 футов). [ 25 ] [ 40 ]
В 2021 году Виктор Дж. Перес, Ронни М. Ледер и Тедди Бадо предложили метод оценки общей длины мегалодона по сумме ширин коронок зубов. Используя более полные зубные ряды мегалодона, они реконструировали зубную формулу, а затем сравнили их с ныне живущими акулами. Исследователи отметили, что уравнения высоты кроны Симады 2002 года дают сильно различающиеся результаты для разных зубов, принадлежащих одной и той же акуле (диапазон погрешности ± 9 метров (30 футов)), что ставит под сомнение некоторые выводы предыдущих исследований с использованием этого метода. Используя самый большой зуб, доступный авторам, GHC 6, с шириной коронки 13,3 сантиметра (5,2 дюйма), они оценили максимальную длину тела примерно в 20 метров (66 футов) с диапазоном ошибок примерно ± 3,5 метра ( 11 футов). [ 40 ] Эта оценка максимальной длины также была поддержана Купером и его коллегами в 2022 году. [ 48 ]
Есть отдельные сообщения о зубах большего размера, чем те, что найдены в музейных коллекциях. [ 25 ] Гордон Хаббелл из Гейнсвилля, Флорида , обладает верхним передним зубом мегалодона, максимальная высота которого составляет 18,4 сантиметра (7,25 дюйма), одним из крупнейших известных образцов зубов акулы. [ 60 ] Кроме того, реконструкция челюсти мегалодона размером 2,7 на 3,4 метра (9 на 11 футов), разработанная охотником за ископаемыми Вито Бертуччи, содержит зуб, максимальная высота которого, как сообщается, превышает 18 сантиметров (7 дюймов). [ 61 ]
Зубы и сила прикуса

Наиболее распространенными окаменелостями мегалодона являются его зубы. Диагностические характеристики включают треугольную форму, прочную структуру, большой размер, мелкие зубцы, отсутствие боковых зубцов и видимую V-образную шейку (там, где корень встречается с коронкой ). [ 28 ] : 55 [ 34 ] Зуб встречался с челюстью под крутым углом, как у большой белой акулы. Зуб был закреплен волокнами соединительной ткани , а шероховатость основания, возможно, добавляла ему механической прочности . [ 62 ] Язычная сторона зуба, обращенная к языку, была выпуклой; а губная сторона, другая сторона зуба, была слегка выпуклой или плоской. Передние зубы были почти перпендикулярны челюсти и симметричны, тогда как задние зубы были наклонными и асимметричными. [ 63 ]
Зубы мегалодона могут иметь наклонную высоту (длину по диагонали) более 180 миллиметров (7,1 дюйма) и являются самыми крупными из всех известных видов акул. [ 29 ] : 33 подразумевая, что это была самая крупная из всех крупных хищных акул. [ 35 ] В 1989 году почти полный набор зубов мегалодона был обнаружен в Сайтаме, Япония . Еще один почти полный связанный зубной ряд мегалодона был раскопан в формациях Йорктаун в Соединенных Штатах и послужил основой для реконструкции челюсти мегалодона в Национальном музее естественной истории (USNM). На основе этих открытий в 1996 году была создана искусственная зубная формула мегалодона. [ 28 ] : 55 [ 64 ]
Зубная формула мегалодона: 2.1.7.4 3.0.8.4 . Как видно из формулы, у мегалодона в челюстях было четыре типа зубов: передние, промежуточные, боковые и задние. Технически промежуточный зуб мегалодона выглядит как верхний передний и называется «А3», потому что он довольно симметричен и не направлен мезиально (сторона зуба обращена к средней линии челюстей, где встречаются левая и правая челюсти). Мегалодон имел очень крепкий зубной ряд. [ 28 ] : 20–21 и имел более 250 зубов в челюстях, занимающих 5 рядов. [ 29 ] : iv Вполне возможно, что у крупных особей мегалодона были челюсти размером примерно 2 метра (6,6 футов) в поперечнике. [ 29 ] : 129 Зубы также были зазубренными , что повышало эффективность разрезания плоти или костей. [ 26 ] [ 29 ] : 1 Акула, возможно, была в состоянии открыть пасть под углом 75°, хотя реконструкция USNM приближается к углу в 100°. [ 35 ]

В 2008 году группа ученых под руководством С. Роу провела эксперимент по определению силы укуса большой белой акулы, используя экземпляр длиной 2,5 метра (8,2 фута), а затем изометрически масштабировала результаты для ее максимального размера и силы укуса. консервативный минимум и максимум массы тела мегалодона. Они оценили силу укуса последней от 108 514 до 182 201 ньютона (от 24 395 до 40 960 фунтов силы) при заднем укусе по сравнению с силой укуса 18 216 ньютонов (4095 фунтов силы) для самой крупной подтвержденной большой белой акулы и 7 495 ньютонов (1685 фунтов силы). для плакодермы рыбы Dunkleosteus . Кроме того, Роу и его коллеги отметили, что акулы трясутся в стороны во время кормления, увеличивая генерируемую силу, что, вероятно, привело бы к тому, что общая сила, испытываемая добычей, оказалась бы выше расчетной. [ 46 ] [ 65 ]
В 2021 году Антонио Балель и Умберто Феррон использовали моделирование методом конечных элементов , чтобы изучить распределение напряжений трех типов зубов мегалодона и близкородственных видов мегазубых при воздействии передних и боковых сил, последняя из которых возникает, когда акула трясется. его голова разрывает плоть. В результате моделирования были выявлены более высокие уровни стресса в зубах мегалодона под действием боковых силовых нагрузок по сравнению с его видами-предшественниками, такими как O. obliquus и O. angusteidens, когда размер зубов был исключен как фактор. Это говорит о том, что зубы мегалодона имели иное функциональное значение, чем предполагалось ранее, что бросает вызов предыдущим интерпретациям о том, что морфология зубов мегалодона была в первую очередь обусловлена изменением рациона питания в сторону морских млекопитающих. Вместо этого авторы предположили, что это был побочный продукт увеличения размеров тела, вызванный гетерохронным отбором . [ 66 ]
Внутренняя анатомия

Мегалодон представлен в летописи окаменелостей зубами, центрами позвонков и копролитами . [ 35 ] [ 67 ] Как и у всех акул, скелет мегалодона состоял из хрящей, а не из костей ; следовательно, большинство ископаемых образцов плохо сохранились. [ 68 ] Чтобы поддерживать крупный зубной ряд, челюсти мегалодона должны были быть более массивными, толстыми и более сильно развитыми, чем челюсти большой белой особи, у которой сравнительно изящный зубной ряд. Его хондрокраниум , хрящевой череп, имел бы более массивный и крепкий вид, чем у большой белой. Его плавники были пропорциональны его большему размеру. [ 35 ]
несколько ископаемых позвонков Было найдено . Наиболее примечательным примером является частично сохранившийся позвоночный столб единственного экземпляра, раскопанный в бассейне Антверпена , Бельгия, в 1926 году. Он состоит из 150 центров позвонков , размеры которых варьируются от 55 миллиметров (2,2 дюйма) до 155 миллиметров (6 дюймов). в диаметре. Позвонки акулы, возможно, стали намного больше, и изучение экземпляра показало, что у него было больше позвонков, чем у экземпляров любой известной акулы, возможно, более 200 центров; только великий белый приблизился к нему. [ 35 ] Еще один частично сохранившийся позвоночный столб мегалодона был раскопан в формации Грам в Дании в 1983 году и включает 20 позвонков, диаметр которых варьируется от 100 миллиметров (4 дюйма) до 230 миллиметров (9 дюймов). [ 62 ]

Копролитовые останки мегалодона имеют спиралевидную форму, что указывает на то, что у акулы, возможно, был спиральный клапан в форме штопора , часть нижнего кишечника , похожая на современные ламнообразные акулы. Останки миоценового копролита были обнаружены в округе Бофорт, Южная Каролина , один из них имел размер 14 см (5,5 дюйма). [ 67 ]
Готфрид и коллеги реконструировали весь скелет мегалодона, который позже был выставлен на обозрение в Морском музее Калверта в США и Южноафриканском музее Изико . [ 35 ] [ 30 ] Эта реконструкция имеет длину 11,3 метра (37 футов) и представляет собой взрослого мужчину. [ 35 ] : 61 основан на онтогенетических изменениях, которые испытывает большая белая акула в течение своей жизни. [ 35 ] : 65
Палеобиология
Ареал и среда обитания
Мегалодон имел космополитическое распространение ; [ 21 ] [ 59 ] его окаменелости были раскопаны во многих частях мира, включая Европу, Африку, Америку и Австралию. [ 28 ] : 67 [ 69 ] Чаще всего это происходило в субтропических и умеренных широтах. [ 21 ] [ 28 ] : 78 Он был обнаружен на широте до 55 ° с.ш .; предполагаемый диапазон допустимых температур составлял 1–24 ° C (34–75 ° F). [ примечание 1 ] Возможно, он обладал способностью переносить такие низкие температуры благодаря мезотермии, физиологической способности крупных акул поддерживать более высокую температуру тела, чем окружающая вода, путем сохранения метаболического тепла . [ 21 ]
Мегалодон населял широкий спектр морской среды (т.е. мелководье в прибрежных водах, районы прибрежного апвеллинга , болотистые прибрежные лагуны , песчаное побережье и глубоководные районы) и вел переходный образ жизни. Взрослые мегалодоны не были многочисленны на мелководье и в основном населяли морские районы. Мегалодон, возможно, перемещался между прибрежными и океаническими водами, особенно на разных стадиях своего жизненного цикла. [ 29 ] : 33 [ 71 ]
Ископаемые останки демонстрируют тенденцию к тому, что экземпляры в Южном полушарии в среднем крупнее, чем в Северном, со средней длиной 11,6 и 9,6 метров (38 и 31 фут) соответственно; а также больше в Тихом океане, чем в Атлантике, со средней длиной 10,9 и 9,5 метров (36 и 31 фут) соответственно. Они не предполагают какой-либо тенденции изменения размера тела с абсолютной широтой или изменения размера с течением времени (хотя считается, что линия Кархароклов в целом демонстрирует тенденцию к увеличению размера с течением времени). Общая модальная длина оценивается в 10,5 метров (34 фута), при этом распределение длины смещено в сторону более крупных особей, что предполагает экологическое или конкурентное преимущество более крупного размера тела. [ 24 ]
Местонахождение окаменелостей
Мегалодон имел глобальное распространение, и окаменелости акулы были найдены во многих местах по всему миру, граничащих со всеми океанами неогена . [ 72 ]
| Эпоха | Формирование | Состояние | Континент |
|---|

Отношения добычи

Хотя акулы, как правило, приспособлены к питанию, большие размеры мегалодона, его способность плавать на высокой скорости и мощные челюсти в сочетании с впечатляющим кормовым аппаратом сделали его высшим хищником , способным поедать широкий спектр животных. Otodus megalodon был, вероятно, одним из самых могущественных хищников, когда-либо существовавших. [ 48 ] Исследование, посвященное изотопам кальция вымерших и современных пластиножаберных акул и скатов, показало, что мегалодон питался на более высоком трофическом уровне, чем современная ему большая белая акула («выше» в пищевой цепи ). [ 77 ]
Ископаемые данные указывают на то, что мегалодон охотился на многие виды китообразных, такие как дельфины, небольшие киты, цетотерии , сквалодонтиды (акулозубые дельфины), кашалоты , гренландские киты и рорквалы . [ 52 ] [ 78 ] [ 79 ] Помимо этого, они также преследовали тюленей, сирен и морских черепах. [ 71 ] Акула была оппортунистом и рыбоядной , а также охотилась на более мелкую рыбу и других акул. [ 52 ] На многих костях китов были обнаружены глубокие порезы, скорее всего, оставленные их зубами. [ 28 ] : 75 Различные раскопки обнаружили зубы мегалодона, лежащие рядом с пережеванными останками китов. [ 28 ] : 75 [ 30 ] а иногда и в непосредственной связи с ними. [ 80 ]
Экология питания мегалодона, по-видимому, менялась в зависимости от возраста и места обитания, как и у современной большой белой акулы . Вполне вероятно, что взрослая популяция мегалодонов у берегов Перу нацеливалась в первую очередь на китов длиной от 2,5 до 7 метров (от 8,2 до 23 футов) и другую добычу, меньшую, чем она сама, а не на крупных китов того же размера, что и они сами. [ 78 ] Между тем, диета молодых особей, вероятно, состояла в основном из рыбы. [ 34 ] [ 81 ]
Соревнование

Мегалодон столкнулся с высококонкурентной средой . [ 82 ] Его положение на вершине пищевой цепи [ 83 ] вероятно, оказали существенное влияние на структурирование морских сообществ. [ 82 ] [ 84 ] Ископаемые свидетельства указывают на корреляцию между мегалодоном и появлением и диверсификацией китообразных и других морских млекопитающих. [ 28 ] : 78 [ 82 ] Молодые особи мегалодона предпочитали места обитания, где было много мелких китообразных, а взрослые особи мегалодона предпочитали места обитания, где было много крупных китообразных. Такие предпочтения могли возникнуть вскоре после их появления в олигоцене. [ 28 ] : 74–75
Мегалодон был современником зубатых китов , питающихся китами (особенно макрорапториальных кашалотов и сквалодонтид ), которые также, вероятно, были одними из главных хищников той эпохи и составляли конкуренцию. [ 82 ] Некоторые из них достигали гигантских размеров, например, Ливиатан , рост которого оценивался от 13,5 до 17,5 метров (от 44 до 57 футов). Окаменелые зубы неопределенного вида таких физетероидов из шахты Ли-Крик, Северная Каролина, указывают на то, что максимальная длина тела составляла от 8 до 10 м (от 26 до 33 футов), а максимальная продолжительность жизни составляла около 25 лет. Это сильно отличается от современных косаток аналогичного размера, которые живут до 65 лет, что позволяет предположить, что в отличие от последних, которые являются высшими хищниками, эти физетероиды подвергались нападению со стороны более крупных видов, таких как мегалодон или ливятан . [ 85 ] К позднему миоцену , около 11 млн лет назад, численность и разнообразие макрорапториалов значительно сократилось. Другие виды, возможно, заполнили эту нишу в плиоцене. [ 82 ] [ 86 ] такие как ископаемая косатка Orcinus citoniensis, которая, возможно, была стайным хищником и нацеливалась на добычу, крупнее его самого, [ 30 ] [ 87 ] [ 88 ] [ 89 ] но этот вывод оспаривается, [ 23 ] и, вероятно, это был хищник-универсал, а не специалист по морским млекопитающим. [ 90 ]
Мегалодон, возможно, стал причиной конкурентного исключения современных ему белых акул , поскольку летописи окаменелостей показывают, что другие виды акул избегали регионов, в которых они обитали, и в основном держались более холодных вод того времени. [ 91 ] [ 28 ] : 77 В районах, где их ареалы, казалось, перекрывались, например, в плиоценовой Нижней Калифорнии , вполне возможно, что мегалодон и большая белая акула занимали эту территорию в разное время года, преследуя разную мигрирующую добычу. [ 28 ] : 77 [ 92 ] Мегалодон, вероятно, также имел склонность к каннибализму , как и современные акулы. [ 93 ]
Стратегии кормления

Акулы часто используют сложные охотничьи стратегии, чтобы привлечь крупных животных. Стратегии охоты на большую белую акулу могут быть похожи на то, как мегалодон охотился на свою крупную добычу. [ 94 ] Следы укусов мегалодона на окаменелостях кита позволяют предположить, что он использовал иные стратегии охоты на крупную добычу, чем большая белая акула. [ 52 ]
Один конкретный экземпляр — останки неописанного миоценового усатого кита длиной 9 метров (30 футов) — предоставил первую возможность количественно проанализировать его атакующее поведение. В отличие от больших белых особей, которые нацелены на нижнюю часть живота своей добычи, мегалодон, вероятно, нацелился на сердце и легкие, а их толстые зубы приспособлены для прокусывания твердых костей, о чем свидетельствуют следы укусов, нанесенные на грудную клетку и другие твердые костные участки на останках китов. [ 52 ] Более того, схемы нападения могут различаться для добычи разного размера. Ископаемые останки некоторых мелких китообразных, например цетотериев, позволяют предположить, что их с большой силой протаранили снизу, прежде чем убить и съесть, на основании компрессионных переломов . [ 94 ]
Есть также свидетельства того, что для нападения на хищных кашалотов существовала возможная отдельная стратегия охоты; Зуб, принадлежащий неопределенному физетероиду длиной 4 м (13 футов), очень напоминающему зуб Acrophyseter, обнаруженный на фосфатной шахте Нутриен Аврора в Северной Каролине, позволяет предположить, что мегалодон или O. chubutensis мог целиться в голову кашалота, чтобы нанести смертельный укус, в результате которого на зубе остались характерные следы укуса. Хотя нельзя исключать возможность падальщика, расположение следов укусов больше соответствует нападениям хищников, чем кормлению падальщиками, поскольку челюсть не является особенно питательной областью, на которой акула может питаться или сосредоточиваться на ней. Тот факт, что следы укусов были обнаружены на корнях зубов, также позволяет предположить, что акула сломала челюсть кита во время укуса, что позволяет предположить, что укус был чрезвычайно сильным. Окаменелость также примечательна тем, что является первым известным примером антагонистического взаимодействия между кашалотом и зубатой акулой, зафиксированным в летописи окаменелостей. [ 95 ]
В плиоцене появились более крупные китообразные. [ 96 ] Мегалодон, по-видимому, еще больше усовершенствовал свою стратегию охоты, чтобы справиться с этими крупными китами. На многочисленных окаменевших костях ласт и хвостовых позвонках крупных китов плиоцена были обнаружены следы укусов мегалодона, что позволяет предположить, что мегалодон обездвиживал большого кита перед тем, как убить его и питаться им. [ 46 ] [ 52 ]
Рост и размножение

В 2010 году Эрет подсчитал, что скорость роста мегалодона почти в два раза выше, чем у нынешней большой белой акулы. Он также подсчитал, что замедление или прекращение соматического роста мегалодона происходит примерно в 25-летнем возрасте, что позволяет предположить, что этот вид имел чрезвычайно запоздалую половую зрелость. [ 97 ] В 2021 году Шимада и его коллеги рассчитали скорость роста особи ростом примерно 9,2 м (30 футов) на основе образца колонны бельгийских позвоночных, который предположительно содержит годовые кольца роста на трех позвонках. По их оценкам, человек умер в возрасте 46 лет, со скоростью роста 16 см (6,3 дюйма) в год и длиной 2 м (6 футов 7 дюймов) при рождении. Для особи ростом 15 м (49 футов), которую они считали максимально достижимым размером, это будет соответствовать продолжительности жизни от 88 до 100 лет. [ 98 ] Однако в 2022 году Купер и его коллеги оценили длину этого 46-летнего человека почти в 16 м (52 фута) на основе 3D-реконструкции, в результате которой длина всего позвоночного столба составила 11,1 м (36 футов); исследователи утверждали, что такая разница в размерах произошла из-за того, что Шимада и его коллеги экстраполировали его размер только на основе центров позвонков. [ 48 ]
Мегалодон, как и современные акулы, использовал нагульные территории для рождения детенышей, особенно в тепловодной прибрежной среде с большим количеством пищи и защитой от хищников. [ 34 ] Питомники были обнаружены в формации Гатун в Панаме, формации Калверт в Мэриленде, Банко де Консепсьон на Канарских островах , [ 99 ] и формация Bone Valley во Флориде. Учитывая, что все современные ламнообразные акулы рождают живых детенышей, считается, что это справедливо и в отношении мегалодона. [ 100 ] Младенческие мегалодоны в наименьшем случае были около 3,5 метров (11 футов). [ 35 ] : 61 а детеныши были уязвимы для нападения других видов акул, таких как большая акула-молот ( Sphyrna mokarran ) и кривозубая акула ( Hemipristis serra ). [ 34 ] Их диетические предпочтения демонстрируют онтогенетический сдвиг : [ 35 ] : 65 Молодые мегалодоны обычно охотятся на рыбу. [ 34 ] морские черепахи, [ 71 ] дюгони , [ 29 ] : 129 и мелкие китообразные; зрелый мегалодон переселился в прибрежные районы и поедал крупных китообразных. [ 28 ] : 74–75
Исключительный случай в летописи окаменелостей позволяет предположить, что молодые мегалодоны могли время от времени нападать на гораздо более крупных баленоптеридных китов. Три следа зубов, по-видимому, оставленные плиоценовой акулой длиной от 4 до 7 метров (от 13 до 23 футов), были обнаружены на ребре предка синего или горбатого кита, что свидетельствует о последующем заживлении, которое предположительно было нанесено ювенильный мегалодон. [ 101 ] [ 102 ]
Вымирание
Изменение климата
За время существования мегалодона на Земле произошел ряд изменений, которые повлияли на морскую жизнь. Тенденция к похолоданию, начавшаяся в олигоцене 35 млн лет назад, в конечном итоге привела к оледенению на полюсах. Геологические события изменили течения и осадки; среди них было закрытие Центральноамериканского морского пути и изменения в океане Тетис , способствовавшие охлаждению океанов. Остановка Гольфстрима не позволила воде, богатой питательными веществами, достичь основных морских экосистем, что могло отрицательно повлиять на источники пищи. Самое большое колебание уровня моря в кайнозойскую эру произошло в плио-плейстоцене , примерно от 5 миллионов до 12 тысяч лет назад, из-за расширения ледников на полюсах, что отрицательно повлияло на прибрежную среду и, возможно, способствовало ее исчезновению. наряду с таковыми некоторых других видов морской мегафауны. [ 103 ] Эти океанографические изменения, в частности падение уровня моря, возможно, ограничили многие подходящие мелководные места нагула мегалодонов с теплой водой, препятствуя размножению. [ 104 ] Питомники имеют решающее значение для выживания многих видов акул, отчасти потому, что они защищают молодь от хищников. [ 105 ] [ 34 ]
Поскольку его ареал, очевидно, не распространялся на более холодные воды, мегалодон, возможно, не был способен сохранять значительное количество метаболического тепла, поэтому его ареал ограничивался сокращением более теплых вод. [ 104 ] [ 79 ] [ 106 ] Ископаемые свидетельства подтверждают отсутствие мегалодона в регионах по всему миру, где температура воды значительно снизилась во время плиоцена. [ 28 ] : 77 Однако анализ распространения мегалодона во времени позволяет предположить, что изменение температуры не сыграло прямой роли в его вымирании. Его распространение в миоцене и плиоцене не коррелировало с тенденциями потепления и похолодания; хотя численность и распространение в плиоцене снизились, мегалодон действительно продемонстрировал способность населять более холодные широты. Он был обнаружен в местах со средней температурой от 12 до 27 ° C (от 54 до 81 ° F) и общим диапазоном от 1 до 33 ° C (от 34 до 91 ° F), что указывает на то, что глобальная протяженность подходящей среды обитания не должны были сильно зависеть от произошедших изменений температуры. [ 21 ] Это согласуется с доказательствами того, что это была мезотерма . [ 50 ]
Изменение экосистемы

Наибольшего разнообразия морские млекопитающие достигли в миоцене. [ 28 ] : 71 например, у усатых китов, у которых более 20 признанных миоценовых родов по сравнению с только шестью существующими родами. [ 108 ] Такое разнообразие представляло идеальные условия для существования такого суперхищника, как мегалодон. [ 28 ] : 75 К концу миоцена многие виды усатых китов вымерли; [ 82 ] выжившие виды могли быть более быстрыми пловцами и, следовательно, более неуловимой добычей. [ 29 ] : 46 Кроме того, после закрытия Центральноамериканского морского пути разнообразие и численность тропических китов уменьшились. [ 106 ] Вымирание мегалодона коррелирует с упадком многих мелких линий усатых китов, и вполне возможно, что он полностью зависел от них как от источника пищи. [ 78 ] Кроме того, было обнаружено, что вымирание морской мегафауны во время плиоцена привело к уничтожению 36% всех крупных морских видов, включая 55% морских млекопитающих, 35% морских птиц, 9% акул и 43% морских черепах. Вымирание было избирательным для эндотермных и мезотермных животных по сравнению с пойкилотермными животными , что предполагает уменьшение запасов пищи. [ 103 ] и, таким образом, согласуется с тем, что мегалодон является мезотермичным. [ 50 ] Мегалодон, возможно, был слишком большим, чтобы поддерживать себя за счет сокращающихся морских пищевых ресурсов. [ 104 ] Охлаждение океанов в плиоцене могло ограничить доступ мегалодона к полярным регионам, лишив его мигрировавших туда крупных китов. [ 106 ]
Конкуренция со стороны крупных зубатых китов, таких как макрохищные кашалоты, появившиеся в миоцене, и представители рода Orcinus (т. е. Orcinus citoniensis ) в плиоцене. [ 82 ] [ 86 ] предполагается, что это способствовало упадку и исчезновению мегалодона. [ 21 ] [ 29 ] : 46–47 [ 104 ] [ 109 ] Но это предположение оспаривается: [ 23 ] Orcininae появились в среднем плиоцене, когда O. citoniensis был зарегистрирован из Плиоцен Италии, [ 86 ] [ 110 ] и подобные формы, зарегистрированные в плиоцене Англии и Южной Африки. [ 86 ] что указывает на способность этих дельфинов справляться со все более распространенными температурами холодной воды в высоких широтах. [ 86 ] В некоторых исследованиях предполагалось, что эти дельфины были макрофагами. [ 21 ] но при ближайшем рассмотрении оказывается, что эти дельфины не являются макрофагами и вместо этого питаются мелкой рыбой. [ 110 ] С другой стороны, о гигантских макрохищных кашалотах, таких как ливьятан -подобные формы, в последний раз сообщалось из Австралии и Южной Африки около 5 миллионов лет назад. [ 111 ] [ 112 ] [ 113 ] Другие, такие как Hoplocetus и Scaldicetus, также занимали нишу, аналогичную нише современных косаток , но последняя из этих форм исчезла в плиоцене. [ 114 ] [ 110 ] Представители рода Orcinus стали крупными и макрофагами в плейстоцене . [ 110 ]
Палеонтолог Роберт Бессенекер и его коллеги перепроверили летопись окаменелостей мегалодона на наличие ошибок радиоуглеродного датирования и пришли к выводу, что он исчез около 3,5 миллионов лет назад. [ 23 ] Бессенекер и его коллеги также предполагают, что мегалодон пострадал от фрагментации ареала из-за климатических изменений. [ 23 ] и конкуренция с белыми акулами могла способствовать его упадку и исчезновению. [ 23 ] Предполагается, что конкуренция с белыми акулами является фактором и в других исследованиях. [ 115 ] [ 21 ] [ 109 ] но эта гипотеза требует дальнейшей проверки. [ 116 ] Считается, что множественные совокупные факторы окружающей среды и экологии, включая изменение климата и температурные ограничения, сокращение популяций добычи и конкуренцию за ресурсы с белыми акулами, способствовали сокращению и исчезновению мегалодона. [ 109 ]
Вымирание мегалодона подготовило почву для дальнейших изменений в морских сообществах. Средний размер тела усатых китов значительно увеличился после его исчезновения, хотя, возможно, из-за других, связанных с климатом, причин. [ 117 ] И наоборот, увеличение размера усатых китов могло способствовать исчезновению мегалодона, поскольку они, возможно, предпочитали охотиться на китов меньшего размера; Следы укусов на крупных видах китов могли остаться от акул-падальщиков. Мегалодон, возможно, просто вымер с более мелкими видами китов, такими как Piscobalaena nana . [ 107 ] Вымирание мегалодона оказало положительное влияние на других хищников того времени, таких как большая белая акула, в некоторых случаях распространившись на регионы, где мегалодон исчез. [ 21 ] [ 115 ] [ 118 ]
В популярной культуре

Мегалодон изображался во многих художественных произведениях, включая фильмы и романы, и продолжает оставаться популярной темой художественной литературы, связанной с морскими чудовищами . [ 119 ] Сообщения о предположительно свежих зубах мегалодона, таких как те, которые были найдены на корабле HMS Challenger в 1873 году и которым зоолог Владимир Чернецкий в 1959 году датирован возрастом от 11 000 до 24 000 лет, помогли популяризировать среди криптозоологов заявления о недавнем выживании мегалодона . [ 120 ] Эти утверждения были дискредитированы и, вероятно, представляют собой зубы, которые хорошо сохранились благодаря толстому осадку в диоксида марганца минеральной корке и поэтому имели более низкую скорость разложения и сохранили белый цвет во время окаменения . Цвет ископаемых зубов мегалодона может варьироваться от почти белого до темно-коричневого, серого и синего, а некоторые ископаемые зубы могли быть переотложены в более молодой слой . Утверждения о том, что мегалодон мог оставаться неуловимым на глубине, подобно большеротой акуле , которая была обнаружена в 1976 году, маловероятны, поскольку акула жила в теплых прибрежных водах и, вероятно, не могла выжить в холодной и бедной питательными веществами глубоководной морской среде . [ 121 ] [ 122 ]
Современная художественная литература о мегалодоне, дожившем до наших дней, была впервые представлена в романе Мег: Роман о глубоком ужасе» « Стива Альтена 1997 года и его последующих продолжениях. Впоследствии Мегалодон начал появляться в фильмах, таких как фильм « Атака акулы 3: Мегалодон» 2002 года , а затем «Мег» , фильм 2018 года, основанный на книге 1997 года, который собрал в прокате более 500 миллионов долларов. [ 120 ] [ 123 ]
Animal Planet « Псевдодокументальный фильм Русалки: найденное тело» включал в себя встречу 1,6 млн лет назад стаи русалок и мегалодона. [ 124 ] Позже, в августе 2013 года, канал Discovery открыл свой ежегодный сериал «Неделя акул» еще одним телевизионным фильмом «Мегалодон: Акула-монстр жива» . [ 125 ] спорная документация о существе, которая представила предполагаемые доказательства того, что мегалодоны все еще живы. Эта программа подверглась критике за то, что она полностью вымышленная и неадекватно раскрывает свою вымышленную природу; например, все предполагаемые изображенные ученые были оплачиваемыми актерами, и в самом документальном фильме не было раскрыто, что он был вымышленным. По опросу Discovery, 73% зрителей документального фильма считают, что мегалодон не вымер. В 2014 году «Дискавери» повторно выпустил в эфир «Жизнь акулы-монстра» вместе с новой часовой программой « Мегалодон: Новые доказательства » и дополнительной художественной программой под названием «Акула тьмы: Гнев подводной лодки» , что вызвало дальнейшую негативную реакцию со стороны средств массовой информации и научное сообщество. [ 52 ] [ 126 ] [ 127 ] [ 128 ] Несмотря на критику со стороны ученых, «Мегалодон: Жизнь акулы-монстра» имел огромный успех в рейтингах, собрав 4,8 миллиона зрителей, что является наибольшим показателем для любого эпизода «Недели акул» на тот момент. [ 129 ]
Зубы мегалодона — окаменелость штата Северная Каролина . [ 130 ]
См. также
Примечания
- ^ Карбонизированный биоапатит из зуба мегалодона (местонахождение неизвестного источника), возраст которого составляет 5,75 ± 0,9 млн лет назад, был проанализирован на содержание изотопных соотношений кислорода ( 18 Т/ 16 О) и углерод ( 13 С/ 12 C), используя методику термометра с карбонатными изотопами, чтобы получить оценку температуры окружающей среды в среде, окружающей этого человека, в 19 ± 4 ° C. [ 70 ]
Ссылки
- ^ Jump up to: а б с д Агассис, Л. (1843). Исследования ископаемых рыб (на французском языке). Полет. 3. дои : 10.5962/bhl.title.4275 .
- ^ Jump up to: а б с д Бриньон, А. (2021). «Исторические и номенклатурные заметки о зубах некоторых большезубых акул (Elasmobranchii, Otodontidae) кайнозоя Нью-Джерси (США)» . Rivista Italiana di Paleontologia e Stratigrafia . 127 (3): 595–625. дои : 10.13130/2039-4942/16440 .
- ^ « Отод ( Megaselachus ) мегалодон (Агассис, 1837)» . SharkReferences.com . Проверено 24 октября 2017 г.
- ^ Истман, ЧР (1904). Геологическая служба Мэриленда . Том. 2. Балтимор, Мэриленд: Университет Джонса Хопкинса. п. 82.
- ^ Jump up to: а б Каппетта, Х. (1987). «Мезозойские и кайнозойские пластиножаберные». Справочник по палеоихтиологии . Том. 3Б. Мюнхен, Германия: Фридрих Пфайль. ISBN 978-3-89937-046-1 . ОСЛК 829906016 .
- ^ Хэй, ОП (1901). «Библиография и каталог ископаемых позвоночных Северной Америки» . Бюллетень Геологического общества США (179): 308.
- ^ Jump up to: а б с д и Шимада, К.; Чендлер, RE; Лам, ОЛТ; Танака, Т.; Уорд, диджей (2016). «Новая неуловимая акула-отодонтид (Lamniformes: Otodontidae) из нижнего миоцена и комментарии к таксономии родов отодонтид, включая кладу «мегатубатых»». Историческая биология . 29 (5): 1–11. дои : 10.1080/08912963.2016.1236795 . S2CID 89080495 .
- ^ Jump up to: а б Лоури, Д.; Годфри, SJ; Эшельман, Р. (2011). «Комплексная геология, палеонтология и археология: использование коренными американцами зубов ископаемых акул в районе Чесапикского залива» . Археология восточной части Северной Америки . 9 : 93–108. JSTOR 23265116 .
- ^ Jump up to: а б Фаррелл, AD (2021). Износ и функциональный анализ преконтактных наборов зубов акул из Флориды (MS). Государственный университет Флориды.
- ^ Джейкобс, Дж. «Национальный исторический парк культуры Хоупвелл «Акулы зубы» HOCU - 2832 и 4222» . Служба национальных парков .
- ^ Jump up to: а б Даффин, CJ (2017). «Зубы ископаемых акул как алексифармы». У Филипа Векслера (ред.). Токсикология Средних веков и Возрождения . стр. 125–133. дои : 10.1016/B978-0-12-809554-6.00012-3 . ISBN 9780128095546 .
- ^ Jump up to: а б Форли, М.; Геррини, А. (2022). «Quaestio de Fossilibus: Glossopetres, Snake Tongues и Ceraunids». История окаменелостей на протяжении веков . стр. 41–83. дои : 10.1007/978-3-031-04687-2_6 . ISBN 978-3-031-04686-5 .
- ^ Jump up to: а б Брессан, Д. (14 января 2016 г.). «Как рассечение головы акулы раскрыло истинную природу окаменелостей» . Форбс .
- ^ «Николас Стено» . Музей палеонтологии Калифорнийского университета .
- ^ Дэвидсон, JP (2000). «Историческая точка зрения: рыбные сказки: приписывание первой иллюстрации зуба ископаемой акулы Ричарду Верстегану (1605 г.) и Николасу Стено (1667 г.)» . Труды Академии естественных наук Филадельфии . 150 : 329–344. JSTOR 4065077 .
- ^ μεγάλος . Лидделл, Генри Джордж ; Скотт, Роберт ; Греко-английский лексикон в проекте «Персей»
- ^ ὀδών . Лидделл, Генри Джордж ; Скотт, Роберт ; Греко-английский лексикон в проекте «Персей»
- ^ Jump up to: а б Вудворд, А.С. (1899 г.). Каталог ископаемых рыб Британского музея (Естественная история), Часть I. Содержащие пластиножаберные . Британский музей (естественная история). стр. 415–420. дои : 10.5962/bhl.title.61854 .
- ^ Ябе, Х.; Гото, М.; Канеко, Н. (2004). «Возраст Carcharocles megalodon (Lamniformes: Otodontidae): обзор стратиграфических записей». Палеонтологическое общество Японии . 75 : 7–15.
- ^ Jump up to: а б с Готфрид, доктор медицины; Фордайс, RE (2001). «Связанный экземпляр Carcharodon angustidens (Chondrichthyes, Lamnidae) из позднего олигоцена Новой Зеландии с комментариями о Carcharodon взаимоотношениях » . Журнал палеонтологии позвоночных . 21 (4): 730–739. doi : 10.1671/0272-4634(2001)021[0730:AASOCA]2.0.CO;2 . S2CID 86092645 . Архивировано из оригинала 17 декабря 2019 года . Проверено 6 ноября 2017 г.
- ^ Jump up to: а б с д и ж г час я дж к л Пимьенто, К.; Макфадден, Би Джей; Клементс, CF; Варела, С.; Харамильо, К.; Велес-Хуарбе, Дж.; Силлиман, БР (2016). «Географические закономерности распространения Carcharocles megalodon с течением времени открывают подсказки о механизмах вымирания». Журнал биогеографии . 43 (8): 1645–1655. Бибкод : 2016JBiog..43.1645P . дои : 10.1111/jbi.12754 . S2CID 55776834 .
- ^ Jump up to: а б с Пимьенто, К.; Клементс, CF (2014). «Когда вымер Carcharocles megalodon ? Новый анализ ископаемых летописей» . ПЛОС ОДИН . 9 (10): e111086. Бибкод : 2014PLoSO...9k1086P . дои : 10.1371/journal.pone.0111086 . ПМК 4206505 . ПМИД 25338197 .
- ^ Jump up to: а б с д и ж Бессенекер, RW; Эрет, диджей; Лонг, диджей; Черчилль, М.; Мартин, Э.; Бессенекер, SJ (2019). «Раннеплиоценовое вымирание мегазубой акулы Otodus megalodon : вид из восточной части северной части Тихого океана» . ПерДж . 7 : е6088. дои : 10.7717/peerj.6088 . ПМК 6377595 . ПМИД 30783558 .
- ^ Jump up to: а б с д Пимьенто, К.; Балк, Массачусетс (2015). «Тенденции размеров тела вымершей гигантской акулы Carcharocles megalodon : глубокий взгляд на морских хищников» . Палеобиология . 41 (3): 479–490. Бибкод : 2015Pbio...41..479P . дои : 10.1017/pab.2015.16 . ПМЦ 4541548 . ПМИД 26321775 .
- ^ Jump up to: а б с д и Симада, Кенсю (2019). «Возвращение к размеру большезубой акулы Otodus megalodon (Lamniformes: Otodontidae)». Историческая биология . 33 (7): 1–8. дои : 10.1080/08912963.2019.1666840 . ISSN 0891-2963 . S2CID 208570844 .
- ^ Jump up to: а б с д Нюберг, КГ; Чампальо, CN; Рэй Г.А. (2006). «Прослеживание происхождения большой белой акулы Carcharodon carcharias с использованием морфометрического анализа ископаемых зубов». Журнал палеонтологии позвоночных . 26 (4): 806–814. doi : 10.1671/0272-4634(2006)26[806:TTAOTG]2.0.CO;2 . S2CID 53640614 .
- ^ Jump up to: а б с д Эрет диджей; Хаббелл Г.; Макфадден Би Джей (2009). «Исключительная сохранность белой акулы Кархародона из раннего плиоцена Перу». Журнал палеонтологии позвоночных . 29 (1): 1–13. дои : 10.1671/039.029.0113 . JSTOR 20491064 . S2CID 129585445 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р Климли, Питер; Эйнли, Дэвид (1996). «Эволюция» . Большие белые акулы: биология Carcharodon carcharias . Сан-Диего, Калифорния: Academic Press. ISBN 978-0-12-415031-7 . OCLC 212425118 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот Ренц, Марк (2002). Мегалодон: Охота на охотника . Лихай-Эйкерс, Флорида: PaleoPress. стр. 100-1 1–159. ISBN 978-0-9719477-0-2 . OCLC 52125833 .
- ^ Jump up to: а б с д и Андрес, Лутц (2002). « C. megalodon — Большезубая акула, Кархародон против Кархарокла » . сайт ископаемых.com . Проверено 16 января 2008 г.
- ^ Jump up to: а б Перес, виджей; Годфри, SJ; Кент, BW; Уимс, RE; Нэнс, младший (2019). «Переход между Carcharocles chubutensis и Carcharocles megalodon (Otodontidae, Chondrichthyes): потеря бокового бугорка с течением времени» . Журнал палеонтологии позвоночных . 38 (6): e1546732. дои : 10.1080/02724634.2018.1546732 .
- ^ Jump up to: а б Сиверссон, М.; Линдгрен, Дж.; Ньюбри, МГ; Седерстрем, П.; Кук, ТД (2015). «Сеноман-кампанские (поздний мел) среднепалеоширотные акулы типа Cretalamna Appendiculata » (PDF) . Acta Palaeontologica Polonica . 60 (2): 339–384. дои : 10.4202/app.2012.0137 . S2CID 58906204 .
- ^ Бентон, MJ; Пирсон, ПН (2001). «Видообразование в летописи окаменелостей». Тенденции экологии и эволюции . 16 (7): 405–411. дои : 10.1016/s0169-5347(01)02149-8 . ПМИД 11403874 .
- ^ Jump up to: а б с д и ж г час Пимьенто, Каталина; Эрет, Дана Дж.; Макфадден, Брюс Дж.; Хаббелл, Гордон (2010). Степанова, Анна (ред.). «Древний питомник вымершей гигантской акулы Мегалодона из миоцена Панамы» . ПЛОС ОДИН . 5 (5): е10552. Бибкод : 2010PLoSO...510552P . дои : 10.1371/journal.pone.0010552 . ПМК 2866656 . ПМИД 20479893 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р Готфрид, доктор медицины; Компаньо, LJV; Боуман, Южная Каролина (1996). «Размер и анатомия скелета гигантской большезубой акулы Carcharodon megalodon ». В Климли; Эйнли (ред.). Большие белые акулы: биология Carcharodon carcharias . Сан-Диего, Калифорния: Academic Press. стр. 55–89. ISBN 978-0124150317 .
- ^ Симада, Кенсю; Ямаока, Юта; Курихара, Юкито; Такакува, Юджи; Майш, Гарри М.; Беккер, Мартин А.; Игл, Роберт А.; Гриффитс, Майкл Л. (23 июня 2023 г.). «Мозаичный кальцинированный хрящ и плакоидная чешуя неогеновой большезубой акулы Otodus megalodon (Lamniformes: Otodontidae) позволяют по-новому взглянуть на ее биологию и эволюцию региональной эндотермии и гигантизма в кладе отодонтид» . Историческая биология . 36 (7): 1259–1273. дои : 10.1080/08912963.2023.2211597 . ISSN 0891-2963 . S2CID 259597157 .
- ^ Ренц, Марк (2002). Мегалодон: Охота на охотника . Лихай-Эйкерс, Флорида: PaleoPress. стр. 100-1 1–159. ISBN 978-0-9719477-0-2 . OCLC 52125833 .
- ^ Портелл, Роджер; Хьюбелл, Гордон; Донован, Стивен; Грин, Джереми; Харпер, Дэвид ; Пикерилл, Рон (2008). «Миоценовые акулы в формациях Кендис и Гранд-Бэй на Карриаку, Гренадины, Малые Антильские острова» (PDF) . Карибский научный журнал . 44 (3): 279–286. дои : 10.18475/cjos.v44i3.a2 . S2CID 87154947 . Архивировано из оригинала (PDF) 20 июля 2011 года.
- ^ Пимьенто, Каталина; Эрет, Дана Дж.; Макфадден, Брюс Дж.; Хаббелл, Гордон (2010). Степанова, Анна (ред.). «Древний питомник вымершей гигантской акулы Мегалодона из миоцена Панамы» . ПЛОС ОДИН . 5 (5): е10552. Бибкод : 2010PLoSO...510552P . дои : 10.1371/journal.pone.0010552 . ПМК 2866656 . ПМИД 20479893 .
- ^ Jump up to: а б с д Перес, Виктор; Ледер, Ронни; Бадо, Тедди (2021). «Оценка длины тела неогеновых макрофагов пластинчатообразных акул (Carcharodon и Otodus), полученная на основе связанных с ними ископаемых зубных рядов» . Электронная палеонтология . 24 (1): 1–28. дои : 10.26879/1140 .
- ^ Пимьенто, К.; Макфадден, Би Джей; Клементс, CF; Варела, С.; Харамильо, К.; Велес-Хуарбе, Дж.; Силлиман, БР (2016). «Географические закономерности распространения Carcharocles megalodon с течением времени открывают подсказки о механизмах вымирания». Журнал биогеографии . 43 (8): 1645–1655. Бибкод : 2016JBiog..43.1645P . дои : 10.1111/jbi.12754 . S2CID 55776834 .
- ^ Jump up to: а б Макклейн, Крейг Р.; Балк, Меган А.; Бенфилд, Марк К.; Бранч, Тревор А.; Чен, Кэтрин; Косгроув, Джеймс; Дав, Алистер DM; Гаскинс, Линдси С.; Хелм, Ребекка Р.; Хохберг, Фредерик Г.; Ли, Фрэнк Б. (13 января 2015 г.). «Определение размеров океанских гигантов: закономерности внутривидовых изменений размеров морской мегафауны» . ПерДж . 3 : е715. дои : 10.7717/peerj.715 . ISSN 2167-8359 . ПМЦ 4304853 . ПМИД 25649000 .
- ^ И., Кастро, Хосе (2011). Акулы Северной Америки . Издательство Оксфордского университета, США. ISBN 978-0-19-978097-6 . OCLC 958576172 .
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Боррель, Асунсьон; Агилар, Алекс; Газо, Манель; Кумарран, РП; Кардона, Луис (1 декабря 2011 г.). «Профили стабильных изотопов китовой акулы (тип Rhincodon) предполагают сегрегацию и различия в рационе в зависимости от пола и размера» . Экологическая биология рыб . 92 (4): 559–567. Бибкод : 2011EnvBF..92..559B . дои : 10.1007/s10641-011-9879-y . ISSN 1573-5133 . S2CID 37683420 .
- ^ Jump up to: а б Протеро, Дональд Р. (25 августа 2015 г.). «09. Мегачелюсти: самая большая рыба. Кархарокл». История жизни в 25 окаменелостях: рассказы о бесстрашных охотниках за ископаемыми и чудесах эволюции . Нью-Йорк Чичестер, Западный Суссекс: Издательство Колумбийского университета. стр. 96–110. дои : 10.7312/prot17190-010 . ISBN 978-0-231-53942-5 .
- ^ Jump up to: а б с Роу, С.; Хубер, доктор медицинских наук; Лоури, М.; МакГенри, К.; Морено, К.; Клаузен, П.; Феррара, TL; Каннингем, Э.; Дин, Миннесота; Саммерс, AP (2008). «Трёхмерный компьютерный анализ механики челюстей белой акулы: насколько сильно может укусить большая белая акула?» (PDF) . Журнал зоологии . 276 (4): 336–342. дои : 10.1111/j.1469-7998.2008.00494.x .
- ^ Купер, Дж.А.; Пимьенто, К.; Феррон, ХГ; Бентон, MJ (2020). «Размеры тела вымершей гигантской акулы Otodus megalodon : 2D-реконструкция» . Научные отчеты . 10 (14596): 14596. Бибкод : 2020NatSR..1014596C . дои : 10.1038/s41598-020-71387-y . ПМЦ 7471939 . ПМИД 32883981 .
- ^ Jump up to: а б с д и Купер, Дж.А.; Хатчинсон-младший; Бернви, округ Колумбия; Клифф, Г.; Уилсон, Р.П.; Дикен, ML; Мензель, Дж.; Роу, С.; Пирло, Дж.; Пимьенто, К. (2022). «Вымершая акула Otodus megalodon была трансокеанским суперхищником: выводы из 3D-моделирования» . Достижения науки . 8 (33): eabm9424. Бибкод : 2022SciA....8M9424C . дои : 10.1126/sciadv.abm9424 . ISSN 2375-2548 . ПМЦ 9385135 . ПМИД 35977007 .
- ^ Джейкоби, DMP; Сириват, П.; Фриман, Р.; Карбоне, К. (2015). «Зависит ли увеличение скорости плавания акул от метаболизма?» . Письма по биологии . 12 (10): 20150781. doi : 10.1098/rsbl.2015.0781 . ПМЦ 4707698 . ПМИД 26631246 .
- ^ Jump up to: а б с Феррон, ХГ (2017). «Региональная эндотермия как триггер гигантизма у некоторых вымерших макрохищных акул» . ПЛОС ОДИН . 12 (9): e0185185. Бибкод : 2017PLoSO..1285185F . дои : 10.1371/journal.pone.0185185 . ПМК 5609766 . ПМИД 28938002 .
- ^ Шимада, К.; Беккер, Массачусетс; Гриффитс, МЛ (2020). «Длина тела, челюстей и зубных рядов пластинчатообразных акул-макрофагов, а также эволюция размеров тела у пластинообразных с особым упором на «запредельный» гигантизм большезубой акулы Otodus megalodon » . Историческая биология . 33 (11): 1–17. дои : 10.1080/08912963.2020.1812598 .
- ^ Jump up to: а б с д и ж г Протеро, ДР (2015). «Мега-Челюсти» . История жизни в 25 окаменелостях . Нью-Йорк, Нью-Йорк: Издательство Колумбийского университета . стр. 96–110. ISBN 978-0-231-17190-8 . ОСЛК 897505111 .
- ^ Хельфман, Г.; Берджесс, GH (2014). Акулы: Руководство по ответам для животных . Балтимор, Мэриленд: Издательство Университета Джонса Хопкинса . п. 19. ISBN 978-1-4214-1310-5 . OCLC 903293986 .
- ^ Рэндалл, Джон Э. (1973). «Размер большой белой акулы ( Кархародона )». Научный журнал . 181 (4095): 169–170. Бибкод : 1973Sci...181..169R . дои : 10.1126/science.181.4095.169 . ПМИД 17746627 . S2CID 36607712 .
- ^ Шембри, Патрик (1994). «Природное наследие Мальты» . Природное наследие. В : 105–124.
- ^ Папсон, Стивен (1992). «Авторское право: пересечь линию террора». Журнал американской культуры . 15 (4): 67–81. дои : 10.1111/j.1542-734X.1992.1504_67.x .
- ^ Готфрид, доктор медицины; Фордайс, RE (2001). «Связанный экземпляр Carcharodon angustidens (Chondrichthyes, Lamnidae) из позднего олигоцена Новой Зеландии с комментариями о Carcharodon взаимоотношениях » . Журнал палеонтологии позвоночных . 21 (4): 730–739. doi : 10.1671/0272-4634(2001)021[0730:AASOCA]2.0.CO;2 . S2CID 86092645 . Архивировано из оригинала 17 декабря 2019 года . Проверено 6 ноября 2017 г.
- ^ Симада, Кенсю (2002). «Взаимосвязь между размером зубов и общей длиной тела белой акулы Carcharodon carcharias (Lamniformes: Lamnidae)». Журнал исследований ископаемых . 35 (2): 28–33.
- ^ Jump up to: а б Пеппер, Кэтрин; Жерар Гонсалес-Бёрд; Дана Дж. Эрет; Остин Дж. В. Хенди; Брюс Дж. Макфадден; Чарльз Харамилло (2013). «Акулы и скаты (Chondrichthyes, Elasmobranchii) из позднемиоценовой формации Гатун в Панаме» (PDF ) Журнал палеонтологии . 87 (5): 755–774. Бибкод : 2013JPal...87..755P . дои : 10.1666/12-117 . S2CID 45662900 . Архивировано из оригинала (PDF) 29 октября.
- ^ Крейн, Б. (2017). «Доисторический убийца, похороненный в грязи» . Житель Нью-Йорка . Проверено 10 декабря 2017 г.
- ^ Мастейн, А. (2011). «Продается: самые большие в мире акульи челюсти» . ЖиваяНаука . Проверено 31 августа 2017 г.
- ^ Jump up to: а б Алмгрин, SE Bendix (15 ноября 1983 г.). « Carcharodon megalodon из верхнего миоцена Дании, с комментариями к эмалоиду пластиножаберных зубов: короноин» . Бюллетень Геологического общества Дании . 32 : 1–32. CiteSeerX 10.1.1.514.1782 . дои : 10.37570/bgsd-1983-32-01 . S2CID 53311833 . НАИД 10012345550 .
- ^ Реолид, М.; Молина, Дж. М. (2015). «Находка Carcharocles megalodon в восточном бассейне Гвадалквивира (верхний миоцен, Южная Испания)» . Геологические исследования . 71 (2): e032. дои : 10.3989/egeol.41828.342 .
- ^ Уено, Т.; Сакамото, О.; Секин, Х. (1989). «Описание почти полного набора зубов Carcharodon megalodon из слоя среднего миоцена в префектуре Сайтама, Япония». Бюллетень Музея естественной истории Сайтамы . 7 : 73–85.
- ^ Андерсон, PSL; Вестнит, М. (2009). «Биомеханическая модель кинематики питания Dunkleosteus terrelli (Arthrodira, Placodermi)». Палеобиология . 35 (2): 251–269. Бибкод : 2009Pbio...35..251A . дои : 10.1666/08011.1 . S2CID 86203770 .
- ^ Баллелл, А.; Феррон, ХГ (2021). «Биомеханические данные о зубном ряде большезубых акул (Lamniformes: Otodontidae)» . Научные отчеты 11 (1232): 1232. doi : 10.1038/s41598-020-80323-z . ПМЦ 7806677 . ПМИД 33441828 .
- ^ Jump up to: а б Стрингер, Г.Л.; Кинг, Л. (2012). «Позднеэоценовые акулий-копролиты из глины Язу на северо-востоке Луизианы» . Музей естественной истории и науки Нью-Мексико, Бюллетень . Позвоночные корполиты. 57 :301.
- ^ «Факты и информация об акуле-мегалодоне: подробности» . сайт ископаемых.com . Проверено 18 сентября 2017 г.
- ^ Фицджеральд, Эрих (2004). «Обзор местонахождений ископаемых третичных китообразных (млекопитающих) в Австралии» . Мемуары музея Виктории . 61 (2): 183–208. дои : 10.24199/j.mmv.2004.61.12 .
- ^ Леффлер, Н.; Фибиг, Дж.; Мульч, А.; Тюткен, Т.; Шмидт, Британская Колумбия; Байнаи, Д.; Конрад, AC; Вакер, У.; Бетчер, Мэн (2019). «Уточнение температурной зависимости кислорода и слипшегося изотопного состава структурно связанного карбоната в апатите». Geochimica et Cosmochimica Acta . 253 : 19–38. Бибкод : 2019GeCoA.253...19L . дои : 10.1016/j.gca.2019.03.002 . S2CID 107992832 .
- ^ Jump up to: а б с Агилера О.; Огилера ERD (2004). «Гигантозубые белые акулы и широкозубые мако (Lamnidae) из неогена Венесуэлы: их роль в Карибском бассейне, комплекс мелководных рыб» . Карибский научный журнал . 40 (3): 362–368.
- ^ Jump up to: а б « Кархароклес мегалодон » . Ископаемые работы . Вход в базу данных палеобиологии . Проверено 28 августа 2017 г.
- ^ «Молодой принц Джордж подарил 23-миллионному зубу вымершей акулы, найденной на Мальте» . Времена Мальты . 27 сентября 2020 г. Проверено 28 сентября 2020 г.
- ^ Юн, К. (2020). «Новый экземпляр Cosmopolitodus hastalis (Lamniformes, Lamnidae) из миоценовой Южной Кореи» . Зооразнообразие . 54 (5): 433–438. дои : 10.15407/zoo2020.05.433 . S2CID 229274996 .
- ^ Малышкина Татьяна П.; Уорд, Дэвид Дж.; Назаркин Михаил Владимирович; Нам, Ги-Су; Квон, Сын Хёк; Ли, Чон Хён; Ким, Тэ-Ван; Ким, До-Квон; Пэк, Ду Сон (2022). «Миоценовые пластиножаберные из формации Духо, Южная Корея». Историческая биология . 35 (9): 1726–1741. дои : 10.1080/08912963.2022.2110870 .
- ^ Чхве, Сын; Ли, Юнг-Нам (2017). «Обзор окаменелостей тел позвоночных с Корейского полуострова и перспективы» . Геонаучный журнал . 21 (6): 867–889. Бибкод : 2017GescJ..21..867C . дои : 10.1007/s12303-017-0040-6 . ISSN 1226-4806 . S2CID 133835817 .
- ^ Мартин, Дж. Э.; Такаил, Т.; Сильвен, А.; Кэтрин, Г.; Винсент, Б. (2015). «Изотопы кальция раскрывают трофическое положение современных и ископаемых пластиножаберных». Химическая геология . 415 : 118–125. Бибкод : 2015ЧГео.415..118М . doi : 10.1016/j.chemgeo.2015.09.011 .
- ^ Jump up to: а б с Колларета, А.; Ламберт, О.; Ландини, В.; Ди Сельма, К.; Малинверно, Э.; Варас-Малка, Р.; Урбина, М.; Бьянуччи, Г. (2017). «Гигантская вымершая акула Carcharocles megalodon нацелилась на мелкую добычу? Следы укусов на останках морских млекопитающих из позднего миоцена Перу». Палеогеография, Палеоклиматология, Палеоэкология . 469 : 84–91. Бибкод : 2017PPP...469...84C . дои : 10.1016/j.palaeo.2017.01.001 . hdl : 10281/151854 .
- ^ Jump up to: а б Морган, Гэри С. (1994). «Где гигантская белая акула?» (PDF) . Палеонтологические темы . 2 (3): 1–2. Архивировано из оригинала (PDF) 22 июля 2016 года.
- ^ Огилера, Оранжель А.; Гарсия, Луи; Коззуоль, Марио А. (2008). «Гигантозубые белые акулы и трофическое взаимодействие китообразных из плиоценовой карибской формации Парагуана». Палеонтологический журнал 82 (2): 204–208. Бибкод : 2008PalZ...82..204A . дои : 10.1007/BF02988410 . ISSN 0038-2353 . S2CID 84251638 .
- ^ Ландини, В.; Альтамирано-Сера, А.; Колларета, А.; Ди Сельма, К.; Урбина, М.; Бьянуччи, Г. (2017). «Комплекс пластиножаберных позднего миоцена из Серро-Колорадо (формация Писко, Перу)». Журнал южноамериканских наук о Земле . 73 : 168–190. Бибкод : 2017JSAES..73..168L . дои : 10.1016/j.jsames.2016.12.010 .
- ^ Jump up to: а б с д и ж г час Ламберт, О.; Бьянуччи, Г.; Пост, П.; де Мюзон, К.; Салас-Жисмонди, Р.; Урбина, М.; Реймер, Дж. (2010). «Гигантский укус нового хищного кашалота из миоценовой эпохи Перу». Природа . 466 (7302): 105–108. Бибкод : 2010Natur.466..105L . дои : 10.1038/nature09067 . ПМИД 20596020 . S2CID 4369352 .
- ^ Компаньо, Леонард СП (1989). «Альтернативные стили жизни хрящевых рыб во времени и пространстве». Экологическая биология рыб . 28 (1–4): 33–75. дои : 10.1007/BF00751027 . S2CID 22527888 .
- ^ Ферретти, Франческо; Борис Ворм; Грегори Л. Бриттен; Майкл Р. Хейтхаус; Хайке К. Лотце1 (2010). «Схемы и экосистемные последствия сокращения численности акул в океане» (PDF) . Экологические письма . 13 (8): 1055–1071. Бибкод : 2010EcolL..13.1055F . дои : 10.1111/j.1461-0248.2010.01489.x . ПМИД 20528897 . Архивировано из оригинала (PDF) 6 июля 2011 года . Проверено 19 февраля 2011 г.
{{cite journal}}: CS1 maint: числовые имена: список авторов ( ссылка ) - ^ Гилберт, КНЦ; Ивани, ЖК ; Уэн, доктор медицины (2018). «Жить быстро и умирать молодым: история жизни и экология неогенового кашалота». Журнал палеонтологии позвоночных . 38 (2): e1439038. Бибкод : 2018JVPal..38E9038G . дои : 10.1080/02724634.2018.1439038 . S2CID 89750852 .
- ^ Jump up to: а б с д и Хейнинг, Джон; Дальхейм, Мэрилин (1988). «Орцинус косатка» (PDF) . Виды млекопитающих (304): 1–9. дои : 10.2307/3504225 . JSTOR 3504225 . S2CID 253914153 . Архивировано из оригинала (PDF) 5 декабря 2010 года.
- ^ Бьянуччи, Джованни; Уолтер, Ландини (2006). «Кашалот-убийца: новый базальный физетероид (Mammalia, Cetacea) из позднего миоцена Италии» . Зоологический журнал Линнеевского общества . 148 (1): 103–131. дои : 10.1111/j.1096-3642.2006.00228.x .
- ^ Линдберг, доктор медицинских наук; Пайенсон, Северная Дакота (2006). «Эволюционные закономерности китообразных: вылов добычи в глубинах времени» . Киты, китобойный промысел и экосистемы океана . Издательство Калифорнийского университета. п. 77. ИСБН 978-0-520-24884-7 .
- ^ Бессенекер, RW (2013). «Новый комплекс морских позвоночных из поздненеогеновой формации Пурисима в Центральной Калифорнии, часть II: ластоногие и китообразные» . Геодиверситас . 35 (4): 815–940. дои : 10.5252/g2013n4a5 . S2CID 85940452 .
- ^ Бьянуччи, Г. (1997). « Hemisyntrachelus cortesii (Cetacea, Delphinidae) из плиоценовых отложений карьера Кампоре (Сальсомаджори-Терме, Италия») . Бюллетень Итальянского палеонтологического общества . 36 (1): 75–83.
- ^ Антунес, Монтана; Легоинья, П.; Балбинг, А. (2015). «Мегалодон, акула мако и планктонные фораминиферы с континентального шельфа у берегов Португалии и их возраст» . Геология Акта . 13 : 181–190.
- ^ «Палеоэкология мегалодона и белой акулы» . Биология акул и скатов . Проверено 1 октября 2017 г.
- ^ Танке, Даррен; Карри, Филип (1998). «Кусание головы динозавров-тероподов: палеопатологические данные» (PDF) . Гайя (15): 167–184.
- ^ Jump up to: а б Годфри, SJ; Альтман, Дж. (2005). «Миоценовый позвонок китообразных с частично восстановленным коэффициентом сжатия, результат судорог или неудачного нападения гигантской белой акулы Carcharodon megalodon » (PDF) . Джефферсониана (16): 1–12. Архивировано из оригинала (PDF) 31 января 2014 года . Проверено 1 октября 2017 г.
- ^ Стивен Дж. Годфри; Джон Р. Нэнс; Норман Л. Райкер (2021). «Зуб кашалота, укушенный отодусом, из неогена восточного побережья США» (PDF) . Acta Palaeontologica Polonica . 66 (3): 599–603.
- ^ Демере, Томас А.; Берта, Анналиса; Макгоуэн, Майкл Р. (2005). «Таксономическая и эволюционная история ископаемых и современных балаеноптероидных усатых китов». Журнал эволюции млекопитающих . 12 (1/2): 99–143. дои : 10.1007/s10914-005-6944-3 . S2CID 90231 .
- ^ Эрет диджей (2010). «Глава 5 – Макроэволюция, возраст и рост большезубых акул (Lamniformes Otodonridae)». Палеобиология и таксономия вымерших ламнид и отодонтидных акул (Chondrichthyes, Elasmobranchii, Lamniformes) (PDF) . стр. 100–136.
- ^ С., Кеншу; Боннан, МФ; Беккер, Массачусетс; Гриффитс, МЛ (2021). «Онтогенетическая модель роста вымершей большезубой акулы Otodus megalodon – последствия для ее репродуктивной биологии, развития и продолжительности жизни» . Историческая биология . 33 (12): 3254–3259. Бибкод : 2021HBio...33.3254S . дои : 10.1080/08912963.2020.1861608 .
- ^ «Определите канарские окаменелости «мегалодона», самой большой акулы, которая когда-либо жила» [Определите канарские окаменелости «мегалодона», самой большой акулы, которая когда-либо жила] (на испанском языке). Europa Press Noticias SA. 2013 . Проверено 29 августа 2017 г.
- ^ Далви, Северная Каролина; Рейнольдс, доктор медицинских наук (1997). «Эволюционные переходы между яйцекладкой, живорождением и материнским вкладом у акул и скатов» . Труды Королевского общества B: Биологические науки . 264 (1386): 1309–1315. Бибкод : 1997RSPSB.264.1309D . дои : 10.1098/rspb.1997.0181 . ПМК 1688595 .
- ^ Годфри, Стивен (2004). «Экфора» (PDF) . Информационный бюллетень Клуба ископаемых Морского музея Калверта . 19 (1): 1–13. Архивировано из оригинала (PDF) 10 декабря 2010 года.
- ^ Каллал, Р.Дж.; Годфри, SJ; Ортнер, диджей (27 августа 2010 г.). «Реакция костей на ребре плиоценового китообразного указывает на кратковременное выживание после нападения хищников». Международный журнал остеоархеологии . 22 (3): 253–260. дои : 10.1002/oa.1199 .
- ^ Jump up to: а б Пимьенто, К.; Гриффин, JN; Клементс, CF; Сильвестро, Д.; Варела, С.; Уэн, доктор медицины; Харамилло, К. (2017). «Вымирание морской мегафауны плейстоцена и его влияние на функциональное разнообразие» . Экология и эволюция природы . 1 (8): 1100–1106. Бибкод : 2017NatEE...1.1100P . дои : 10.1038/s41559-017-0223-6 . ПМИД 29046566 . S2CID 3639394 .
- ^ Jump up to: а б с д «Вымирание Мегалодона» . Биология акул и скатов . Проверено 31 августа 2017 г.
- ^ Рейли, Майкл (29 сентября 2009 г.). «Доисторический акулий питомник породил гигантов» . Новости Дискавери. Архивировано из оригинала 10 марта 2012 года . Проверено 23 ноября 2013 г.
- ^ Jump up to: а б с Олмон, Уоррен Д.; Стивен Д. Эмсли; Дуглас С. Джонс; Гэри С. Морган (2006). «Океанографические изменения в позднем неогене вдоль западного побережья Флориды: доказательства и механизмы». Журнал геологии . 104 (2): 143–162. Бибкод : 1996JG....104..143A . дои : 10.1086/629811 . S2CID 128418299 .
- ^ Jump up to: а б Колларета, А.; Ламберт, О.; Ландини, В.; Бьянуччи, Г. (2017). «Гигантская вымершая акула Carcharocles megalodon нацелилась на мелкую добычу? Следы укусов на останках морских млекопитающих из позднего миоцена Перу». Палеогеография, Палеоклиматология, Палеоэкология . 469 : 84–91. Бибкод : 2017PPP...469...84C . дои : 10.1016/j.palaeo.2017.01.001 . hdl : 10281/151854 .
- ^ Дули AC; Николас К.Ф.; Ло ZX (2006). «Самый ранний известный представитель клады рорквалов – серых китов (Mammalia, Cetacea)». Журнал палеонтологии позвоночных . 24 (2): 453–463. дои : 10.1671/2401 . JSTOR 4524731 . S2CID 84970052 .
- ^ Jump up to: а б с МакКормак, Джереми; Гриффитс, Майкл Л.; Ким, Сора Л.; Симада, Кенсю; Карнес, Молли; Майш, Гарри; Педерзани, Сара; Бургон, Николя; Яуэн, Клервия; Беккер, Мартин А.; Йёнс, Нильс (31 мая 2022 г.). «Трофическое положение Otodus megalodon и больших белых акул во времени, выявленное с помощью изотопов цинка» . Природные коммуникации . 13 (1): 2980. Бибкод : 2022NatCo..13.2980M . дои : 10.1038/s41467-022-30528-9 . ISSN 2041-1723 . ПМЦ 9156768 . ПМИД 35641494 . S2CID 249235478 .
- ^ Jump up to: а б с д Цитрон, Сара; Гейслер, Джонатан Х.; Альберто, Колларета; Джованни, Бьянуччи (2022). «Систематика, филогения и пищевое поведение старейшей косатки: переоценка Orcinus citoniensis (Capellini, 1883) из плиоцена Тосканы (Италия)». Бюллетень Итальянского палеонтологического общества . 61 (2): 167–186. дои : 10.4435/BSPI.2022.13 .
- ^ «Огромный зуб обнаруживает доисторического Моби Дика в Мельбурне» . Австралазийский научный журнал . Проверено 24 апреля 2016 г.
- ^ «Подвинься, Моби Дик: познакомься с мега-китом Мельбурна» . Сидней Морнинг Геральд. 21 апреля 2016 г.
- ^ Говендер, Р. (2021). «Ископаемые китообразные раннего плиоцена из залива Хондеклип, Намакваленд, Южная Африка». Историческая биология . 33 (4): 574–593. Бибкод : 2021HBio...33..574G . дои : 10.1080/08912963.2019.1650273 . S2CID 202019648 .
- ^ Хампе, О. (2006). «Останки кашалота среднего / позднего миоцена (Odontoceti: Physeteridae) Северной Германии с исправленной классификацией Hoplocetinae» . Ископаемый рекорд . 9 (1): 61–86. Бибкод : 2006FossR...9...61H . дои : 10.1002/mmng.200600002 .
- ^ Jump up to: а б Антунес, Мигель Теллес; Бальбино, Аусенда Касерес (2010). «Большая белая акула Carcharodon carcharias (Linne, 1758) в плиоцене Португалии и ее раннее распространение в Восточной Атлантике» . Испанский журнал палеонтологии . 25 (1): 1–6.
- ^ Каст, Эмма Р.; Гриффитс, Майкл Л.; Ким, Сора. Л.; Рао, Цзысюань К.; Симада, Кенсу; Беккер, Мартин А.; Майш, Гарри М.; Игл, Роберт А.; Кларк, Челесия А.; Нойманн, Эллисон Н.; Карнес, Молли Э.; Людеке, Тина; Лейхлитер, Дженнифер Н.; Мартинес-Гарсия, Альфредо; Ахтар Аллия А.; Ван, Синчен Т.; Хауг, Джеральд Х.; Сигман, Дэниел М. (22 июня 2022 г.). «Кайнозойские большезубые акулы занимали чрезвычайно высокие трофические позиции» . Достижения науки . 8 (25): eabl6529. Бибкод : 2022SciA....8L6529K . дои : 10.1126/sciadv.abl6529 . ПМК 9217088 . ПМИД 35731884 .
- ^ Слейтер, Дж.Дж.; Гольдбоген, Дж.А.; Пайенсон, Северная Дакота (2017). «Независимая эволюция гигантизма усатых китов, связанная с динамикой океана плио-плейстоцена» . Труды Королевского общества B: Биологические науки . 284 (1855): 20170546. doi : 10.1098/rspb.2017.0546 . ПМЦ 5454272 . ПМИД 28539520 .
- ^ Сильвен, Аднет; АК Бальбино; МТ Антунес; Х. М. Марин-Феррер (2010). «Новый ископаемый зуб белой акулы ( Carcharodon carcharias ) из раннего плиоцена Испании. Значение для его палеоэкологии в Средиземноморье». Новогодний ежегодник геологии и палеонтологии . 256 (1): 7–16. дои : 10.1127/0077-7749/2009/0029 .
- ^ Вайншток, Дж. А. (2014). Эшгейтская энциклопедия литературных и кинематографических монстров . Фарнем, Великобритания: Рутледж . стр. 107–108. ISBN 978-1-4094-2562-5 . OCLC 874390267 .
- ^ Jump up to: а б Гимон, Эдвард (5 октября 2021 г.). «Мегалодон: Монстр новой мифологии» . Журнал М/К . 24 (5). дои : 10.5204/mcj.2793 . ISSN 1441-2616 . S2CID 241813307 .
- ^ Роеш, Б.С. (1998). «Критическая оценка предполагаемого современного существования Carcharocles megalodon » . Обзор криптозоологии . 3 (2): 14–24. Архивировано из оригинала 21 октября 2013 года . Проверено 11 ноября 2014 г.
- ^ «Мегалодон еще жив?» . Биология акул и скатов . Проверено 2 октября 2017 г.
- ^ «Мэг (2018)» . Касса Моджо . Проверено 22 декабря 2017 г.
- ^ Сид Беннетт (режиссер) (27 мая 2012 г.). Русалки: Тело найдено (Кинофильм). Планета животных .
- ^ «Неделя акул «Мегалодон: акула-монстр жива» пытается доказать существование доисторической акулы (ВИДЕО)» . Хафф Пост Грин. 5 августа 2013 года . Проверено 11 августа 2013 г.
- ^ Уинстон, Б.; Ванстон, Г.; Чи, В. (2017). «Прогулка по лесу» . Акт документирования: документальный фильм в XXI веке . Нью-Йорк, Нью-Йорк: Издательство Bloomsbury. ISBN 978-1-5013-0918-2 . OCLC 961183719 .
- ^ Фланагин, Дж. (2014). «Извините, фанаты. «Дискавери» перескочил с недели акул» . Нью-Йорк Таймс . Проверено 16 августа 2014 г.
- ^ Шиффман, Дэвид (15 августа 2014 г.). «Неделя акул снова лжет об акулах-монстрах-мегалодонах» . Журнал «Сланец» . Проверено 31 июля 2022 г.
- ^ О'Коннелл, Майки (5 августа 2013 г.). «Телерейтинги: Неделя акул достигла рекордного уровня из-за фальшивого документа «Мегалодон»» . Голливудский репортер . Проверено 31 июля 2022 г.
- ^ «Ископаемые, окаменелые зубы акулы-мегалодона | NCpedia» . ncpedia.org . Проверено 17 октября 2019 г.
Дальнейшее чтение
- Диксон, Калифорния; Грэм, Дж. Б. (ноябрь – декабрь 2004 г.). «Эволюция и последствия эндотермии у рыб» . Физиологическая и биохимическая зоология . 77 (6): 998–1018. дои : 10.1086/423743 . ПМИД 15674772 . S2CID 40104003 .
- Кент, Бреттон В. (1994). Ископаемые акулы региона Чесапикского залива . Колумбия, Мэриленд: Иган Рис и Бойер. ISBN 978-1-881620-01-3 . OCLC 918266672 .
Внешние ссылки
- Появление суперхищных акул
- Кархарокл : вымершая зубастая акула.
- Палеонтолог Марк Ренц показывает на YouTube один из крупнейших зубов мегалодона.
- Специальный выпуск «Недели акул» о мегалодоне с Пэт Маккарти и Джоном Бабиарзом на YouTube с комментариями о его исчезновении.
- свидетельства существования питомника акул возрастом 10 миллионов лет Ископаемые зубы мегалодона показывают на YouTube
- Взгляд эксперта: информация о мегалодоне на YouTube (с участием эксперта Даны Эрет)
- Пластинчатые акулы: 110 миллионов лет господства над океаном на YouTube (с участием эксперта Микаэля Сиверсона)
- Взлет и падение неогеновых гигантских акул на YouTube (с участием эксперта Бреттона Кента)
- Мур, Калли (19 декабря 2018 г.). «Почему мегалодон (определенно) вымер» . PBS Эоны . Архивировано из оригинала 14 ноября 2021 года — на YouTube .