Гибодонтиформные
| Гибодонтиформные Временной диапазон: От позднего девона до позднего мела | |
|---|---|
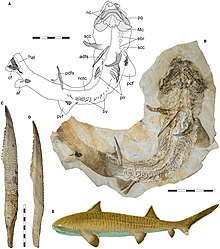 | |
| Ископаемое и восстановление жизни астераканта из поздней юры Европы | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Хондриктиес |
| Подкласс: | пластиножаберные |
| Инфракласс: | Евселахии |
| Заказ: | † Гибодонтиформы Паттерсон, 1966 год. |
| Семьи | |
Посмотреть текст | |
Hybodontiformes , обычно называемые гибодонтами , — вымершая группа акулоподобных хрящевых рыб (хондрихтианы), существовавшая с позднего девона до позднего мела . Гибодонты имеют близкое общее происхождение с современными акулами и скатами ( Neoselachii ) как часть клады Euselachii . Они отличаются от других хондрихтиев характерными шипами на плавниках и головными шипами, присутствующими на головах самцов. Экологически разнообразная группа, они были в изобилии в морской и пресноводной среде в течение позднего палеозоя и раннего мезозоя , но были редки в открытой морской среде к концу юрского периода , будучи в значительной степени заменены современными акулами, хотя они все еще были распространены в пресноводной среде. и маргинальные морские среды обитания. Они дожили до конца мелового периода , прежде чем вымерли.
Этимология
[ редактировать ]Термин «гибодонт» происходит от греческого слова ὕβος или ὑβός, означающего горб или горбатый, и ὀδούς, ὀδοντ, означающего зуб. Такое название было дано из-за их конических сжатых зубов.
Таксономическая история
[ редактировать ]Гибодонты были впервые описаны в девятнадцатом веке на основе изолированных ископаемых зубов ( Агассис , 1837). Гибодонты были впервые отделены от живых акул Зиттелем (1911). [1] Хотя исторически утверждалось, что они имеют тесную связь с современным отрядом акул Heterodontiformes , это было опровергнуто. [2] Hybodontiformes представляют собой группу пластиножаберных общую и сестринскую группу Neoselachii , в которую входят современные акулы и скаты . Hybodontiformes и Neoselachii сгруппированы вместе в кладе Euselachii, за исключением других пластиножаберных всей группы, таких как Xenacanthiformes . [3] Гибодонты разделены на ряд семейств, однако систематика гибодонтов более высокого уровня, особенно мезозойских таксонов, решена плохо. [4]
Описание
[ редактировать ]
Самые крупные гибодонты достигали длины 2–3 метра (6,6–9,8 футов). [4] в то время как некоторые другие гибодонты были намного меньше, их длина тела взрослой особи составляла около 25 сантиметров (0,82 фута). [5] Гибодонты имели в целом крепкое телосложение. Из-за того, что их хрящевые скелеты обычно распадаются после смерти, как и у других хондрихтиев , гибодонты обычно описываются и идентифицируются на основе окаменелостей зубов и плавников, которые с большей вероятностью сохранятся. [4] Редкие частичные или полные скелеты известны из районов исключительной сохранности . [6] [4] [7]

Гибодонты имеют зубы с выступающим бугорком, который выше боковых бугорков. [8] Зубы гибодонта часто сохраняются в виде неполных окаменелостей, поскольку основание зуба плохо прикреплено к коронке. [8] Гибодонты первоначально были разделены на две группы в зависимости от формы зубов. [9] У одной группы были зубы с заостренными бугорками без полости пульпы; это так называемые остеодонтные зубы. Другая группа имела другое расположение бугров и полость пульпы, такие зубы называются ортодонтными. [10] Например, у вида гибодонтов Heterophychodus steinmanni есть зубы-остеодонты с сосудистыми каналами дентина, расположенными вертикально параллельно друг другу, что также называется «трубчатым дентином». [11] Коронки этих зубов-остеодонтов покрыты одним слоем эмальоида . Зубы гибодонта выполняли различные функции в зависимости от вида, включая перемалывание, раздавливание ( дурофагия ), разрывание, сжимание и даже резание. [4]
Гибодонты характеризуются наличием двух спинных плавников, каждому из которых предшествует плавниковый шип. Морфология плавниковых шипов уникальна для каждого вида гибодонтов. Шипы плавников удлиненные и плавно изогнуты назад, при этом задняя часть позвоночника покрыта крючковатыми зубчиками, обычно расположенными в два параллельных ряда, идущих по длине позвоночника, иногда с гребнем между ними. Часть передней части шипов часто покрыта ребристым орнаментом, а у некоторых других гибодонтов эта область покрыта рядами мелких бугорков. Шипы минерализованы и состоят преимущественно из остеодентина , а орнамент состоит из эмали. [12] Подобные шипы плавников также встречаются у многих вымерших групп хондрихтианов, а также у некоторых современных акул, таких как Heterodontus и Squalids . [13] Самцы гибодонтов имели на голове одну или две пары головных шипов, что характерно для гибодонтов. [14] Эти шипы, хотя и имеют различное расположение, [10] всегда располагались позади глазницы, [15] и состояли из основания, разделенного на три доли, при этом основная часть шипа была изогнута назад, большинство экземпляров которых имели заусеницу возле вершины. [10] Эти шипы, как и шипы плавников, были минерализованы, их основание состояло из остеодентина. [16] при этом основная часть позвоночника была покрыта эмалью. Самцы гибодонтов обладали плавниковыми застежками , используемыми при спаривании, как у современных акул. [10] Гибодонты имели полностью гетероцеркальный хвостовой плавник, при этом верхняя лопасть плавника была намного больше нижней из-за входящего в нее шипа. [17] Подобно живым акулам и скатам, кожа гибодонтов была покрыта кожными зубчиками . [18] Гибодонты откладывали икру , подобную той, которую производят живые хрящевые рыбы. Большинство яиц гибодонтов отнесены к роду Palaeoxyris , который сужается к обоим концам, причем на одном конце есть усик, прикрепленный к субстрату, а средняя часть состоит как минимум из трех скрученных полос. [19]
 Зубы Durnovariaodus , члена семейства Hybodontidae.
Зубы Durnovariaodus , члена семейства Hybodontidae. Фрагмент челюсти строфода, специализированного дурофагового гибодонта.
Фрагмент челюсти строфода, специализированного дурофагового гибодонта. Челюсти астераканта , показывающие расположение зубов в челюстях. Зубы были предназначены для захвата
Челюсти астераканта , показывающие расположение зубов в челюстях. Зубы были предназначены для захвата Сохранившийся зубной ряд акродонтида Acrodus , у которого были низкие округлые зубы, используемые в дурофагии.
Сохранившийся зубной ряд акродонтида Acrodus , у которого были низкие округлые зубы, используемые в дурофагии. Зубы Planohybodus , гибодонтида, зубы которого были предназначены для разрывания.
Зубы Planohybodus , гибодонтида, зубы которого были предназначены для разрывания.





Экология
[ редактировать ]
Окаменелости гибодонта встречаются в средах отложения от морских до речных (речные отложения). [20] Считается, что многие гибодонты были эвригалинными и могли переносить широкий диапазон солености. [21] Гибодонты населяли пресноводную среду с самого начала своей эволюционной истории, начиная с каменноугольного периода. [22] Судя по изотопному анализу, некоторые виды гибодонтов, вероятно, постоянно обитали в пресноводной среде. [23] [24] в то время как другие, возможно, мигрировали между морской и пресноводной средой. [25] Предполагается, что один род гибодонтов, Onychoselache из нижнего карбона Шотландии, был способен к амфибийному передвижению, подобно современным оректолобиформным акулам, таким как бамбуковые и эполетные акулы, благодаря хорошо развитым грудным плавникам. [16] Было высказано предположение, что самцы гибодонтов использовали свои головные шипы для захвата самок во время спаривания. [26] Сохранившиеся ящики с яйцами гибодонтов, отнесенных к Palaeoxyris, указывают на то, что по крайней мере некоторые гибодонты откладывали яйца в пресноводной и солоноватой среде, при этом яйца прикреплялись к растительности с помощью усиков. Откладка икры в пресной воде не известна ни у одной ныне живущей хрящевой рыбы. [27] [19] [28] Предполагается, что по крайней мере некоторые гибодонты использовали определенные участки в качестве питомников, например, в триасовых озерных отложениях мадыгенской свиты Кыргызстана, где яйца лончидиона, как предполагается, откладывались на берегу озера или в верховьях реки, где молодые особи вылуплялись и повзрослели, прежде чем мигрировать глубже в озеро во взрослом возрасте. [27]

Считается, что некоторые гибодонты, такие как Hybodus, были активными хищниками, способными питаться быстро движущейся добычей. [2] с сохранившимся содержимым желудка экземпляра Hybodus hauffianus , что указывает на то, что они питались белемнитами . [29] Гибодонты имеют самые разнообразные формы зубов. Такое разнообразие предполагает, что они использовали несколько источников пищи. [8] Считается, что некоторые гибодонты, у которых были более широкие и плоские зубы, специализировались на раздавливании или измельчении добычи с твердым панцирем ( дурофагия ). [20] Часто в одной экосистеме сосуществовали несколько видов гибодонтов с разными предпочтениями в отношении добычи. [30] [11]
Эволюционная история
[ редактировать ]
Самые ранние останки гибодонта относятся к позднему девону ( фамену ) Ирана, принадлежат к роду Roongodus . [31] а также останки, приписанные к Лиссоду того же возраста из Бельгии. [32] Хотя первые окаменелости гибодонтов относятся к позднему девонскому периоду, они, вероятно, произошли от неоселахий (современных акул) в раннем девоне. [33] Каменноугольные гибодонты включают как дурофаговые, так и недурофаговые формы, при этом в пермский период доминировали дурофаговые формы. [32] К пермскому периоду гибодонты имели глобальное распространение. [32] [34] [35] Пермско -триасовое вымирание оказало лишь ограниченное влияние на разнообразие гибодонтов. [36] Максимальное разнообразие гибодонтов наблюдается в триасе. В триасе и ранней юре гибодонтиформы были доминирующими пластиножаберными как в морской, так и в неморской среде. [20] Сдвиг гибодонтов наблюдался в средней юре, переходе между совершенно разными комплексами, наблюдавшимися в триасе-ранней юре и поздней юре-мелу. [20] По мере дальнейшей диверсификации неоселахий (группа современных акул) в позднеюрском периоде гибодонтиформы стали менее распространены в условиях открытого моря, но оставались разнообразными в речных и ограниченных условиях в течение мелового периода. [20] К концу мелового периода количество гибодонтов сократилось до нескольких видов. [37] включая членов Лончидиона [38] и Меристодоноидес . [39] Последние гибодонты исчезли, по-видимому, внезапно, в рамках мел-палеогенового вымирания . [37]
Семьи и роды
[ редактировать ]Систематика гибодонтов считается малоизученной. [4] поэтому представленную классификацию не следует воспринимать как авторитетную.
- Lonchidiidae Герман, 1977 г. [40]
- Дистобатиды
- Акродонтиды
- Гибодонтиды
- Неуверенное сиденье
- Роды форм
- Palaeoxyris (род, используемый для яйцевых капсул гибодонтов)
Ссылки
[ редактировать ]- ^ Zittel, K. von, 1911, Grunzuege der Palaontologie, 2 ed. II. Abt. Vertebrata, vii + 598 pp. R. OldenburgVerlag, Muchen, Berlin.
- ^ Перейти обратно: а б Мэйси, Дж. Г., 2012, Что такое «пластожаберные»? Влияние палеонтологии на понимание филогении и эволюции пластиножаберных: Журнал биологии рыб, т. 80, вып. 5, с. 918-951.
- ^ Коутс, Майкл И.; Титджен, Кристен (март 2017 г.). «Нейрокраниум нижнекаменноугольной акулы Tristychius arcuatus (Agassiz, )» . Труды Королевского общества Эдинбурга по наукам о Земле и окружающей среде . 108 (1): 19–35. Бибкод : 2017EESTR.108...19C . дои : 10.1017/S1755691018000130 . ISSN 1755-6910 . S2CID 135297534 .
- ^ Перейти обратно: а б с д и ж Штумпф, Себастьян; Лопес-Ромеро, Фавиэль А.; Киндлиманн, Рене; Лакомба, Фредерик; Пол, Буркхард; Кривет, Юрген (август 2021 г.). Кэвин, Лайонел (ред.). «Уникальный гибодонтиформный скелет дает новое представление о мезозойской жизни хондрихтиев» . Статьи по палеонтологии . 7 (3): 1479–1505. Бибкод : 2021PPal....7.1479S . дои : 10.1002/spp2.1350 . ISSN 2056-2799 . S2CID 234204226 .
- ^ Вулло, Ромен; Неродо, Дидье; Депре, Эрик (октябрь 2013 г.). «Останки позвоночных из сеноманского (позднемелового) растительного Lagerstätte из Пюи-Пюи (Приморская Шаранта, Франция)» . Меловые исследования . 45 : 314–320. Бибкод : 2013CrRes..45..314V . дои : 10.1016/j.cretres.2013.06.002 .
- ^ Лейн, Дж. А., и Мэйси, Дж. Г., 2009, Грудная анатомия Tribodus limae (Elasmobranchii: Hybodontiformes) из нижнего мела северо-востока Бразилии: Журнал палеонтологии позвоночных, т. 29, вып. 1, с. 25-38.
- ^ Ван, Новая Зеландия; Чжан, X.; Чжу, М.; Чжао, В.-Ж. (май 2009 г.). «Новый сочлененный гибодонтоид из поздней перми северо-западного Китая» . Акта Зоология . 90 : 159–170. дои : 10.1111/j.1463-6395.2008.00382.x .
- ^ Перейти обратно: а б с Кут М.Б., Куни Г., Тинтори А. и Твитчетт Р.Дж., 2013, Новая разнообразная фауна акул из вордской (средней перми) формации Хуфф во внутренней области Хауши-Хукф, Султанат Оман: Палеонтология, v .56, нет. 2, с. 303-343.
- ^ Агассис, Л., 1833-1844, Исследования ископаемых ядов. Невшатель, 5 рейсов. 1420 стр. с доп.
- ^ Перейти обратно: а б с д Мэйси, Дж. Г., 1982, Анатомия и взаимоотношения мезозойских гибодонтных акул : Американский музей Novitates, т. 2724.
- ^ Перейти обратно: а б Куни Г., Сутитхорн В., Баффето Э. и Филипп М., 2003, Гибодонтные акулы из мезозойской группы Корат в Таиланде: Журнал Университета Махасаракхама, т. 22.
- ^ Мэйси, Дж. Г., 1978, Рост и форма шипов гибодонтных акул : Палеонтология, т. 21, вып. 3, с. 657-666.
- ^ Мэйси, Джон Г. (12 марта 2009 г.). «Комплекс позвоночник-щетка у симморииформных акул (Chondrichthyes; Symmoriiformes) с комментариями по модульности спинного плавника» . Журнал палеонтологии позвоночных . 29 (1): 14–24. Бибкод : 2009JVPal..29...14M . дои : 10.1671/039.029.0130 . ISSN 0272-4634 . S2CID 86408226 .
- ^ Рис, Ян (март 2002 г.). «Акулья фауна и среда отложения самых ранних меловых глин Витабека в Эриксдале, южная Швеция» . Труды Королевского общества Эдинбурга по наукам о Земле и окружающей среде . 93 (1): 59–71. дои : 10.1017/S0263593300000328 . ISSN 1473-7116 . S2CID 130984933 .
- ^ Штумпф, Себастьян; Гравирует, Стив; Андервуд, Чарли Дж.; Кривет, Юрген (11 мая 2021 г.). «Durnonovariaodus maiseyi gen. et sp. nov., новый гибодонтиформный акулоподобный хондрихтиан из верхнеюрской глиняной формации Киммеридж в Англии» . ПерДж . 9 : е11362. дои : 10.7717/peerj.11362 . ISSN 2167-8359 . ПМК 8121075 . ПМИД 34026354 .
- ^ Перейти обратно: а б Коутс, Майкл И.; Гесс, Роберт В. (2007). «Новая реконструкция Onychoselache Traquairi, комментарии к ранним хондрихтианским грудным поясам и филогении гибодонтиформ» . Палеонтология . 50 (6): 1421–1446. Бибкод : 2007Palgy..50.1421C . дои : 10.1111/j.1475-4983.2007.00719.x . S2CID 140556654 .
- ^ Ким, Сунь Х.; Симада, Кенсю; Ригсби, Синтия К. (март 2013 г.). «Анатомия и эволюция гетероцеркального хвоста пластинчатообразных акул: ХВОСТОВЫЕ ПЛАВНИКИ ПЛАСТИКОВЫХ АКУЛ» . Анатомическая запись . 296 (3): 433–442. дои : 10.1002/ar.22647 . ПМИД 23381874 . S2CID 205411092 .
- ^ Мэйси, Джон Г.; Дентон, Джон СС (02 сентября 2016 г.). «Формирование дермального рисунка зубцов у меловой гибодонтной акулы Tribodus limae (Euselachii, Hybodontiformes) и его значение для эволюции паттерна хондрихтиевого кожного скелета» . Журнал палеонтологии позвоночных . 36 (5): e1179200. Бибкод : 2016JVPal..36E9200M . дои : 10.1080/02724634.2016.1179200 . ISSN 0272-4634 . S2CID 88591830 .
- ^ Перейти обратно: а б Фишер, Ян; Аксмит, Брайан Дж.; Эш, Сидни Р. (01 марта 2010 г.). «Первое однозначное напоминание о яичной капсуле гибодонтной акулы Palaeoxyris в мезозое Северной Америки» . Новогодний ежегодник геологии и палеонтологии - Трактаты . 255 (3): 327–344. дои : 10.1127/0077-7749/2009/0028 . ISSN 0077-7749 .
- ^ Перейти обратно: а б с д и Рис, Ян, и Андервуд, СиДжей, 2008, Гибодонтные акулы английского бата и келловея (средняя юра): Палеонтология, т. 51, вып. 1, с. 117-147.
- ^ Бэтчелор, Тревор Джон (декабрь 2013 г.). «Новый вид Vectiselachos (Chondrichthyes, Selachii) из раннего мела южной Англии» . Труды Ассоциации геологов . 124 (6): 967–972. Бибкод : 2013ПрГА..124..967Б . дои : 10.1016/j.pgeola.2013.05.001 .
- ^ Вэнь, Вэнь; Кривет, Юрген; Чжан, Циюэ; Бентон, Майкл Дж.; Даффин, Кристофер Дж.; Хуан, Цзинъюань; Чжоу, Чангюн; Ху, Шисюэ; Ма, Чжисинь (30 июня 2022 г.). «Останки гибодонтиформных акул (Chondrichthyes, Elasmobranchii) из нижнего триаса провинции Юньнань, Китай, с комментариями о разнообразии гибодонтиформ в PTB» . Журнал палеонтологии позвоночных . 42 (1). Бибкод : 2022JVPal..42E8712W . дои : 10.1080/02724634.2022.2108712 . ISSN 0272-4634 . S2CID 252136919 .
- ^ Фишер, Ян; Шнайдер, Йорг В.; Фойгт, Силке; Иоахимски, Майкл М.; Тихомирова, Марион; Тюткен, Томас; Гетце, Йенс; Бернер, Ульрих (29 марта 2013 г.). «Изотопы кислорода и стронция из зубов ископаемых акул: экологические и экологические последствия для европейских бассейнов позднего палеозоя» . Химическая геология . 342 : 44–62. Бибкод : 2013ЧГео.342...44Ф . doi : 10.1016/j.chemgeo.2013.01.022 . ISSN 0009-2541 .
- ^ Клюг, Стефани; Тюткен, Томас; Крылья, Оливер; Пфретчнер, Ганс-Ульрих; Мартин, Томас (1 сентября 2010 г.). «Позднеюрский комплекс пресноводных акул (Chondrichthyes, Hybodontiformes) из южной части Джунгарского бассейна, Синьцзян, Северо-Западный Китай» . Палеобиоразнообразие и палеосреда . 90 (3): 241–257. Бибкод : 2010ПдПе...90..241К . дои : 10.1007/s12549-010-0032-2 . ISSN 1867-1608 . S2CID 129236098 .
- ^ Лейцингер, Л.; Кочиш, Л.; Бийон-Брюя, Ж.-П.; Спеццаферри, С.; Веннеманн, Т. (07 декабря 2015 г.). «Исследование стабильных изотопов новой хондрихтийской фауны (кимеридж, Поррентрю, швейцарская Юра): необычный изотопный состав гибодонтовой акулы <i>Asteracanthus</i> под влиянием пресной воды» . Биогеонауки . 12 (23): 6945–6954. Бибкод : 2015BGeo...12.6945L . дои : 10.5194/bg-12-6945-2015 . hdl : 11336/80921 . ISSN 1726-4189 . S2CID 129044300 .
- ^ Турмине-Юэль, Пернель; Уилкс, Ричард; Брокхерст, Дэвид; Остин, Питер А.; Даффин, Кристофер Дж.; Бентон, Майкл Дж. (декабрь 2019 г.). «Микропозвоночные из глиняной формации Уодхерст (нижний мел) кирпичного завода Эшдаун, Восточный Суссекс, Великобритания» . Труды Ассоциации геологов . 130 (6): 752–769. Бибкод : 2019ПрГА..130..752Т . дои : 10.1016/j.pgeola.2019.08.003 . S2CID 202904040 .
- ^ Перейти обратно: а б Фишер Я.Н., Фойгт С., Шнайдер Дж.В., Бухвиц М. и Фойгт С., 2011, Селахская пресноводная фауна из триаса Кыргызстана и ее значение для мезозойских питомников акул: Журнал палеонтологии позвоночных, т. 31, нет. 5, с. 937-953.
- ^ Крюгер, Эшли; Слейтер, Сэм; Вайда, Виви (3 июля 2021 г.). «3D-изображения яиц акул (Paleoxyris) из Швеции с новым пониманием экологии акул ранней юры» . ГФФ . 143 (2–3): 229–247. Бибкод : 2021GFF...143..229K . дои : 10.1080/11035897.2021.1907442 . ISSN 1103-5897 .
- ^ Клюг, Кристиан; Швайгерт, Гюнтер; Хоффманн, Рене; Вайс, Роберт; Де Баетс, Кеннет (декабрь 2021 г.). «Ископаемые остатки падают как источники палеоэкологических данных: «пабулит», состоящий из ракообразных, белемнитов и позвоночных, из раннеюрских сланцев Посидония» . Швейцарский журнал палеонтологии . 140 (1): 10. Бибкод : 2021SwJP..140...10K . дои : 10.1186/s13358-021-00225-z . ISSN 1664-2376 . ПМЦ 8549986 . ПМИД 34721282 .
- ^ Каппетта, Х., Баффетаут, Э., Куни, Г., и Сутитхорн, В., 2006, Новый комплекс пластиножаберных из нижнего мела Таиланда, палеонтология, т. 49, вып. 3, с. 547-555.
- ^ Хайрапетян, В. и Гинтер, М. 2009. Фаменские останки хондрихтиана из разреза Чахрисе, центральный Иран. Acta Geologica Polonica, 59, 173–200.
- ^ Перейти обратно: а б с Ходнетт, Дж. П., Эллиот, Д. К. и Олсон, Т. Дж. 2013. Новый базальный гибодонт (Chondrichthyes, Hybodontiformes) из среднепермской (роудской) формации Кайбаб в северной Аризоне. Бюллетень Музея естественной истории и науки Нью-Мексико, 60:103–108.
- ^ Коутс, М.И., и Гесс, Р.В., 2007, Новая реконструкция Onychoselache Traquairi, комментарии к ранним грудным поясам хондрихтиев и филогении гибодонтиформ: Палеонтология, т. 50, вып. 6, с. 1421-1446.
- ^ Пикук, Брэндон Р.; Бронсон, Эллисон В.; Отоо, Бенджамин К.А.; Сидор, Кристиан А. (ноябрь 2021 г.). «Фауна пресноводных рыб из двух пермских рифтовых долин Замбии, новое дополнение к ихтиофауне южной Пангеи» . Журнал африканских наук о Земле . 183 : 104325. Бибкод : 2021JAfES.18304325P . doi : 10.1016/j.jafrearsci.2021.104325 .
- ^ Сьоне, Альберто Луис; Гуирик-Кавалли, Соледад; Меннуччи, Хорхе Аугусто; Кабрера, Даниэль Альфредо; Фрейе, Рубен Уго (2010). «Останки первых тел позвоночных из перми Аргентины (Elasmobranchii и Actinopterygii)» . Труды Ассоциации геологов . 121 (3): 301–312. Бибкод : 2010ПрГА..121..301С . дои : 10.1016/j.pgeola.2010.04.003 .
- ^ Вэнь, Вэнь; Чжан, Циюэ; Бентон, Майкл Дж.; Кривет, Юрген; Ху, Шисюэ; Хуан, Цзиньюань; Чжоу, Чангюн; Цуй, Синьдун; Ма, Чжисинь; Мин, Сяо (февраль 2023 г.). «Первое появление зубов гибодонтид в биоте Луопинга (средний триас, аниз), что подчеркивает восстановление морской экосистемы после массового вымирания в конце перми» . Палеогеография, Палеоклиматология, Палеоэкология . 617 : 111471. doi : 10.1016/j.palaeo.2023.111471 .
- ^ Перейти обратно: а б Кривет, Юрген; Бентон, Майкл Дж. (ноябрь 2004 г.). «Неоселахское разнообразие (Chondrichthyes, Elasmobranchii) на границе мела и третичного периода» . Палеогеография, Палеоклиматология, Палеоэкология . 214 (3): 181–194. дои : 10.1016/j.palaeo.2004.02.049 .
- ^ Гейтс, Терри А.; Горскак, Эрик; Маковицкий, Питер Дж. (май 2019 г.). «Новые акулы и другие хондрихтии из позднего маастрихта (позднего мела) Северной Америки» . Журнал палеонтологии . 93 (3): 512–530. Бибкод : 2019JPal...93..512G . дои : 10.1017/jpa.2018.92 . ISSN 0022-3360 .
- ^ Штумпф, Себастьян; Мэн, Стефан; Кривет, Юрген (26 января 2022 г.). «Особенности разнообразия позднеюрских хондрихтиев: новые данные на основе исторически собранного комплекса гиподонтиформных зубов из Польши» . Разнообразие . 14 (2): 85. дои : 10.3390/d14020085 . ISSN 1424-2818 .
- ^ Мансанарес, Эстер; Пла, Кристина; Мартинес-Перес, Карлос; Феррон, Умберто; Ботелла, Эктор (2 января 2017 г.). «Lonchidion derenzii, sp. nov., новая акула-лонхидиид (Chondrichthyes, Hybodontiforms) из верхнего триаса Испании, с замечаниями по эмалоиду лонхидиид» . Журнал палеонтологии позвоночных . 37 (1): e1253585. Бибкод : 2017JVPal..37E3585M . дои : 10.1080/02724634.2017.1253585 . HDL : 10550/85565 . ISSN 0272-4634 . S2CID 132467956 .

- Гибодонтиформные
- Отряды доисторических хрящевых рыб
- Первые появления в каменноугольном периоде
- Пенсильванские таксономические порядки
- Приуральские таксономические отряды
- Гваделупские таксономические порядки
- Лопингианские таксономические порядки
- Таксономические порядки раннего триаса
- Таксономические порядки среднего триаса
- Таксономические порядки позднего триаса
- Раннеюрские таксономические порядки
- Среднеюрские таксономические отряды
- Позднеюрские таксономические порядки
- Раннемеловые таксономические порядки
- Таксономические порядки позднего мела