Эволюция рыб
Эта статья включает список общих ссылок , но в ней отсутствуют достаточные соответствующие встроенные цитаты . ( февраль 2024 г. ) |

Эволюция рыб началась около 530 миллионов лет назад во время кембрийского взрыва . Именно в это время у ранних хордовых развились череп и позвоночный столб , что привело к появлению первых черепных животных и позвоночных животных . Первые рыбные линии принадлежат Agnatha , или бесчелюстным рыбам. Ранние примеры включают Haikouichthys . В позднем кембрии впервые появились угревидные бесчелюстные рыбы, называемые конодонтами , и небольшие, в основном панцирные рыбы, известные как остракодермы . Большинство бесчелюстных рыб сейчас вымерло; но современные миноги могут напоминать древнюю рыбу без челюстей. Миноги принадлежат к отряду Cyclostomata , в который входят современные миксины , и эта группа, возможно, рано отделилась от других бесчелюстных.
Самые ранние челюстные позвоночные , вероятно, появились в конце ордовика . Впервые они представлены в летописи окаменелостей двумя силура группами рыб: панцирными рыбами, известными как плакодермы , которые произошли от остракодерм; и акантодии (или колючие акулы). Челюстные рыбы, сохранившиеся до наших дней, также появились в позднем силуре: Chondrichthyes (или хрящевые рыбы) и Osteichthyes (или костистые рыбы). Костные рыбы разделились на две отдельные группы: Actinopterygii (или лучепёрые рыбы) и Sarcopterygii (в которую входят лопастёрые рыбы).
В девонский период произошло резкое увеличение разнообразия рыб, особенно среди остракодерм и плакодерм, а также лопастных рыб и ранних акул. Это привело к тому, что девон стал известен как век рыб . Именно от лопастных рыб произошли четвероногие — четвероногие позвоночные, представленные сегодня земноводными , рептилиями , млекопитающими и птицами . Переходные четвероногие впервые появились в раннем девоне, а к позднему девону появились первые четвероногие. Разнообразие челюстных позвоночных может указывать на эволюционное преимущество челюстного рта ; но неясно, является ли преимущество шарнирной челюсти большей силой укуса, улучшением дыхания или комбинацией факторов.
Рыбы, как и многие другие организмы, сильно пострадали от вымираний на протяжении всей естественной истории. Самые ранние из них — ордовикско-силурийское вымирание — привели к исчезновению многих видов. Позднедевонское вымирание привело к вымиранию к концу девона остракодерм и плакодерм, а также других рыб. Колючие акулы вымерли во время пермско-триасового вымирания ; конодонты вымерли во время триасово-юрского вымирания . Мел -палеогеновое вымирание и нынешнее голоценовое вымирание также повлияли на разнообразие рыб и рыбные запасы .
Обзор [ править ]

- Рыба:
- бесчелюстная рыба ( Агната )
- хрящевая рыба ( Chondrichthyes )
- лучепёрая рыба ( Actinopterygii )
- лопастоперая рыба ( Sarcopterygii )
- Четвероногие:
Рыба, возможно, произошла от животного, похожего на коралловую асцидию ( оболочник ) , чьи личинки во многом напоминают ранних рыб. Первопредки рыб, возможно, сохраняли личиночную форму до взрослой жизни , как это делают некоторые асцидии сегодня, хотя этот путь не может быть доказан.
Позвоночные животные , другими словами, первые рыбы , возникли около 530 миллионов лет назад во время кембрийского взрыва , который привел к увеличению разнообразия животных. [3]

Первыми предками рыб или животных, которые, вероятно, были тесно связаны с рыбами, были Haikouichthys и Myllokunmingia . [6] [3] Все эти два рода появились около 530 млн лет назад . В отличие от другой фауны, доминировавшей в кембрии, эти группы имели базовое строение тела позвоночных : хорду , рудиментарные позвонки и четко выраженные голову и хвост. [7] У всех этих ранних позвоночных не было челюстей в обычном смысле слова, и они питались фильтратором вблизи морского дна. [8]
За ними последовали бесспорные ископаемые позвоночные в виде тяжелобронированных рыб, обнаруженные в породах ордовика (500–430 млн лет назад ).
Первые челюстные позвоночные появились в позднем ордовике и стали обычным явлением в девоне , часто известном как «Эра рыб». [9] Две группы костистых рыб , Actinopterygii и Sarcopterygii , эволюционировали и стали обычным явлением. [10] В девонском периоде вымерли практически все бесчелюстные рыбы, за исключением миног и миксин, а также плакодерми , группы панцирных рыб, доминировавших на протяжении большей части позднего силура , и появились первые лабиринтодонты , промежуточные между рыбами и амфибиями . [10]
Заселение новых ниш привело к разнообразию строения тела, а иногда и к увеличению размеров. Девонский . период (395–345 млн лет назад) принес таких гигантов, как плакодерма Dunkleosteus , которая могла вырасти до семи метров в длину, и первых рыб, дышащих воздухом, которые могли оставаться на суше в течение длительного периода времени Среди этой последней группы были предковые амфибии .
Рептилии произошли от лабиринтодонтов в последующем каменноугольном периоде. Анапсидные синапсидные и стали амниоты были распространены в позднем палеозое , а диапсиды доминировать в мезозое . В море костистая рыба стала доминировать.
Более поздние излучения, например, у рыб в силурийском и девонском периодах, включали меньшее количество таксонов, в основном с очень похожим строением тела. Первыми животными, вышедшими на сушу, были членистоногие . Некоторые рыбы имели легкие и сильные костистые плавники и могли также ползать по суше.
Бесчелюстная рыба [ править ]


Бесчелюстные рыбы относятся к Agnatha типа Chordata , надклассу подтипа Vertebrata . Агната означает «безчелюстная, без челюстей» (с древнегреческого ). [11] Сюда не входят все позвоночные животные с челюстями, известные как челюстноротые . Хотя бесчелюстные рыбы и являются второстепенным элементом современной морской фауны , они занимали видное место среди ранних рыб раннего палеозоя . два типа раннекембрийских животных, которые, по-видимому, имели плавники, мускулатуру позвоночных известны Из раннекембрийских сланцев Маотяньшань в Китае и жабры : Haikouichthys и Myllokunmingia . Жанвье предварительно назначил их Агнате. Третьим возможным бесчелюстным видом из того же региона является Haikouella .
Многие ордовикские , силурийские и девонские бесчелюстные были бронированы тяжелыми, костными и часто искусно выполненными пластинами, полученными из минерализованных чешуек. Первые панцирные бесчелюстные — остракодермы , предшественники костистых рыб и, следовательно, четвероногих (включая человека) — известны с среднего ордовика , а к позднему силуру бесчелюстные достигли высшей точки своей эволюции. Большинство остракодерм, таких как телодонты , остеостраканы и галеаспиды , были более тесно связаны с челюстноротыми, чем с выжившими бесчелюстными, известными как круглоротые. Круглоротые, по-видимому, отделились от других бесчелюстных до появления дентина и кости, которые присутствуют у многих ископаемых бесчелюстных, включая конодонтов . [12] Численность Agnathans в девоне сократилась и так и не восстановилась.
Бесчелюстные в целом парафилетичны . [13] потому что большинство вымерших бесчелюстных принадлежат к стволовой группе челюстноротых рыб, которые произошли от них. [14] [15] Молекулярные данные, как по рРНК [16] и из мтДНК [17] решительно поддерживает теорию о том, что живые бесчелюстные, известные как круглоротые , монофилетичны . [18] В филогенетической таксономии отношения между животными обычно не делятся на ранги, а иллюстрируются в виде вложенного «генеалогического древа», известного как кладограмма . Филогенетическим группам даны определения, основанные на их отношениях друг с другом, а не исключительно на физических признаках, таких как наличие позвоночника. Эта модель вложения часто сочетается с традиционной таксономией в практике, известной как эволюционная таксономия .

Кладограмма веб-проекта « бесчелюстных рыб основана на исследованиях Филиппа Жанвье и других для Древо жизни» . [20] (†=группа вымерла)
| Бесчелюстная рыба |
| |||||||||||||||||||||||||||||||||||||||
† Конодонты [ править ]

Конодонты напоминали примитивных бесчелюстных угрей. Они появились 520 млн лет назад и исчезли 200 млн лет назад. [21] Первоначально они были известны только по зубообразным микрофоссилиям, называемым конодонтовыми элементами . Эти «зубы» интерпретировались по-разному: как фильтрующие устройства или как «захватывающий и дробящий массив». [22] Конодонты имели длину от сантиметра до 40 см промиссума . Их большие глаза располагались сбоку, что делает маловероятной роль хищника. Сохранившаяся мускулатура намекает на то, что некоторые конодонты ( по крайней мере , Промиссум ) были эффективными крейсерами, но неспособными развивать максимальную скорость. [22] В 2012 году исследователи отнесли конодонтов к типу Хордовые на основании плавников с плавниковыми лучами, шевронных мышц и хорды . [23] Некоторые исследователи видят в них позвоночных, внешне похожих на современных миксин и миног . [24] хотя филогенетический анализ предполагает, что они более производны , чем любая из этих групп. [25]
† Остракодермы [ править ]

Остракодермы ( букв. «ракушечные») — бесчелюстные рыбы палеозоя бронированные . Сегодня этот термин не часто встречается в классификациях, поскольку таксон является парафилетическим или полифилетическим и не имеет филогенетического значения. [26] Тем не менее, этот термин до сих пор неофициально используется для обозначения панцирных бесчелюстных рыб.
Панцирь остракодермы состоял из многоугольных пластин толщиной 3–5 мм, которые защищали голову и жабры, а затем перекрывались дальше по телу, как чешуя. Глаза были особенно защищены. Раньше хордовые использовали жабры как для дыхания, так и для питания, тогда как остракодермы использовали жабры только для дыхания . У них было до восьми отдельных жаберных глоточных мешков по бокам головы, которые были постоянно открыты и не имели защитной крышки . В отличие от беспозвоночных , которые используют ресничные движения для перемещения пищи, остракодермы использовали свою мускулистую глотку для создания всасывания, которое втягивало мелкую и медленно движущуюся добычу в рот .
Первыми обнаруженными ископаемыми рыбами были остракодермы. Швейцарский Шотландии анатом Луи Агассис получил несколько окаменелостей костистых панцирных рыб из в 1830-х годах. Ему было трудно их классифицировать, поскольку они не походили ни на одно живое существо. Сначала он сравнил их с современными панцирными рыбами, такими как сом и осетровые, но позже, поняв, что у них нет подвижных челюстей, в 1844 году отнес их к новой группе «остракодермы». [27]
Остракодермы существовали в двух основных группах: более примитивных гетеростраканах и цефаласпидах . Позже, около 420 миллионов лет назад, из одной из остракодерм произошли челюстные рыбы. После появления челюстных рыб большинство видов остракодерм подверглось упадку, а последние остракодермы вымерли в конце девона . [28]
Челюстная рыба [ править ]
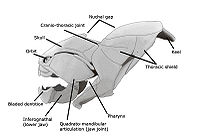

Челюсти позвоночных, вероятно, первоначально развились в силурийский период и появились у -плакодерм рыб , которые в дальнейшем диверсифицировались в девонском периоде . Считается, что двумя наиболее передними глоточными дугами стали сама челюсть и подъязычная дуга соответственно. Подъязычная система удерживает челюсть от черепной коробки, обеспечивая большую подвижность челюстей. которое уже долгое время считалось парафилетическим комплексом, ведущим к появлению более продвинутых челюстноротых, Открытие Entelognathus, предполагает, что плакодермы являются прямыми предками современных костистых рыб.
Как и у большинства позвоночных , челюсти рыб костные или хрящевые , расположены вертикально и состоят из верхней и нижней челюстей . Челюсть происходит от двух передних глоточных дуг, поддерживающих жабры, и обычно несет многочисленные зубы . Предполагается, что череп последнего общего предка современных челюстных позвоночных напоминал акулу. [29]
Считается, что первоначальные селективные преимущества челюстей были связаны не с питанием, а с повышением эффективности дыхания. Челюсти использовались в буккальном насосе (наблюдаемом у современных рыб и амфибий ), который перекачивает воду через жабры рыб или воздух в легкие у амфибий. С течением времени в процессе эволюции было выбрано более привычное (для человека) использование челюстей при кормлении, которое стало очень важной функцией у позвоночных. Многие костистые рыбы существенно модифицировали свои челюсти для всасывания и выпячивания челюстей , в результате чего появились очень сложные челюсти с десятками задействованных костей.
Челюстные позвоночные и челюстные рыбы произошли от более ранних бесчелюстных рыб. Кладограмма челюстных позвоночных является продолжением кладограммы из предыдущего раздела. (†=вымершие)
| Челюстные позвоночные |
| ||||||||||||||||||||||||||||||||||||||||||||||||
† Плакодермы [ править ]


Плакодермы, класс Placodermi («пластинчатые») — вымершие панцирные доисторические рыбы, появившиеся около 430 млн лет назад в раннем и среднем силуре. В основном они были уничтожены во время вымирания в позднем девоне , 378 млн лет назад, хотя некоторые выжили и немного восстановили разнообразие в фаменскую эпоху, а затем полностью вымерли в конце девона, 360 млн лет назад; в конечном итоге они являются предками современных челюстноротых позвоночных. [30] [31] Их голова и грудь были покрыты массивными и часто орнаментированными панцирными пластинами. Остальная часть тела была чешуйчатой или обнаженной, в зависимости от вида. Броневой щит был шарнирным, головной доспех шарнирно крепился к грудному доспеху. Это позволяло плакодермам поднимать голову, в отличие от остракодерм. Плакодермы были первыми рыбами с челюстями; их челюсти, вероятно, произошли от первой из жаберных дуг. На диаграмме справа показаны возникновение и упадок отдельных линий плакодерм: Acanthothoraci , Rhenanida , Antiarchi , Petalichthyidae , Ptyctodontida и Arthrodira .
† Spiny sharks [ edit ]

Колючие акулы класса Acanthodii — вымершие рыбы, которые имеют общие черты как с костными, так и с хрящевыми рыбами, хотя в конечном итоге более тесно связаны с последними и являются их предками. Несмотря на то, что акантодийцы называются «колючими акулами», они произошли от акул, хотя и дали им начало. Они появились в море в начале силурийского периода, примерно за 50 миллионов лет до появления первых акул. В конце концов конкуренция со стороны костистых рыб оказалась слишком сильной, и колючие акулы вымерли в пермский период около 250 млн лет назад. По форме они напоминали акул, но эпидермис их был покрыт мелкими ромбовидными пластинками, наподобие чешуи голостеев ( сарков , боуфинов ).
Хрящевая рыба [ править ]

Хрящевые рыбы класса Chondrichthyes , состоящие из акул , скатов и химер , появились около 395 миллионов лет назад, в среднем девоне , эволюционировав от акантодий. Класс содержит подклассы Holocephali ( химеры ) и пластиножаберные ( акулы и скаты ). Радиация пластиножаберных на схеме справа разделена на следующие таксоны: Cladoselache , Eugeneodontiformes , Symmoriida , Xenacanthiformes , Ctenacanthiformes , Hybodontiformes , Galeomorphii , Squaliformes и Batoidea .
Костистая рыба [ править ]
Костные рыбы класса Osteichthyes характеризуются костным скелетом, а не хрящом . Они появились в позднем силуре , около 419 миллионов лет назад. Недавнее открытие Entelognathus убедительно свидетельствует о том, что костные рыбы (и, возможно, хрящевые рыбы, через акантодий) произошли от ранних плакодерм. [33] Подкласс Osteichthyes, лучепёрые рыбы ( Actinopterygii ), стал доминирующей группой рыб в постпалеозойском и современном мире, насчитывая около 30 000 живых видов. Группы костных (и хрящевых) рыб, появившиеся после девона, характеризовались устойчивыми улучшениями в добывании пищи и передвижении. [34]
Лопастёрая рыба [ править ]

Лопастеперые рыбы, рыбы, принадлежащие к классу Sarcopterygii , представляют собой в основном вымершие костистые рыбы, в основном характеризующиеся крепкими и короткими лопастными плавниками, содержащими прочный внутренний скелет, космоидную чешую и внутренние ноздри. Плавники мясистые, лопастные , парные, соединены с телом одной костью. [37] Плавники лопастных рыб отличаются от плавников всех остальных рыб тем, что каждый из них опирается на мясистый, лопастной, чешуйчатый стебель, отходящий от тела. Грудные сочленены так, что напоминают конечности четвероногих , и брюшные плавники предшественниками которых они были. Плавники превратились в ноги первых четвероногих наземных позвоночных, амфибий . У них также есть два спинных плавника с отдельными основаниями, в отличие от одного спинного плавника у лучепёрых рыб . Мозговая коробка лопастных рыб примитивно имеет шарнирную линию, но у четвероногих и двоякодышащих рыб она утрачена. Многие ранние лопастные рыбы имеют симметричный хвост. У всех лопастных рыб зубы покрыты настоящей эмалью .
Лопастёрые рыбы, такие как целаканты и двоякодышащие рыбы , были самой разнообразной группой костистых рыб в девоне. Таксономисты, придерживающиеся кладистического подхода, включают четвероногих в группу Sarcopterygii, а четвероногие, в свою очередь, включают все виды позвоночных с четырьмя конечностями. [38] Плавниковые конечности лопастных рыб, таких как целаканты, демонстрируют сильное сходство с ожидаемой предковой формой конечностей четвероногих. Кипастеперые рыбы, по-видимому, следовали двум разным линиям развития и соответственно разделены на два подкласса: Rhipidistia ( включая двоякодышащих рыб и Tetrapodomorpha , включающих четвероногих) и Actinistia (целаканты). Первые лопастноперые рыбы, обнаруженные в самых верхних силурийских периодах (ок. 418 млн лет назад), очень напоминали колючих акул , вымерших в конце палеозоя. В раннем и среднем девоне (416–385 млн лет назад), когда в морях доминировали хищные плакодермы , часть лопастных рыб проникла в пресноводные местообитания.

В раннем девоне (416-397 млн лет назад) лопастноперые рыбы разделились на две основные линии — целаканты и рипидистианы . Первые никогда не покидали океанов, и период их расцвета пришелся на поздний девон и каменноугольный период , с 385 по 299 млн лет назад, поскольку в эти периоды они были более распространены, чем в любой другой период фанерозоя; целаканты и сегодня обитают в океанах (род Latimeria ). Рипидистийцы, предки которых, вероятно, жили в эстуариях , мигрировали в пресноводные места обитания. Они, в свою очередь, разделились на две основные группы: двоякодышащие рыбы и тетраподоморфы . Наибольшее разнообразие двоякодышащих рыб наблюдалось в триасовый период; сегодня осталось менее десятка родов. У двоякодышащей рыбы в среднем девоне (397–385 млн лет назад) появились первые протолегкие и протоконечности, развив способность жить вне водной среды. Первые тетраподоморфы, в число которых входили гигантские ризодонты, имели ту же общую анатомию, что и двоякодышащие рыбы, которые были их ближайшими родственниками, но, по-видимому, они не покидали свою водную среду обитания до эпохи позднего девона (385–359 млн лет назад). из четвероногие (четвероногие позвоночные). Четвероногие — единственные тетраподоморфы, выжившие после девона. Лопастёрые рыбы существовали до конца палеозойской эры, понеся тяжелые потери во время пермско-триасового вымирания (251 млн лет назад).
Лучеперая рыба [ править ]

Рыбы с лучевыми плавниками, класс Actinopterygii , отличаются от рыб с лопастными плавниками тем, что их плавники состоят из перепонок кожи, поддерживаемых шипами («лучами»), сделанными из кости или рога. Существуют и другие различия в структурах дыхания и кровообращения. Скелет лучепёрых рыб обычно состоит из настоящей кости, но это не относится к осетрам и веслоносам . [41]
Лучеперые рыбы — доминирующая группа позвоночных, в которую входит половина всех известных видов позвоночных. Они обитают в морских глубинах, прибрежных бухтах, пресноводных реках и озерах и являются основным источником пищи для человека. [41]
Хронология [ править ]
См. также [ править ]
- Сравнительная анатомия
- Эволюция парных плавников
- Ихтиолит
- Конвергентная эволюция рыб
- Список ископаемых мест
- Списки доисторических рыб
- Список лет палеонтологии
- Старый красный песчаник
- Пародии на символ ихтис
- Доисторическая жизнь
- Ходячая рыба - рыба с особенностями четвероногих.
- Палеонтология позвоночных
Ссылки [ править ]
- ^ Бентон, MJ (2005) Палеонтология позвоночных. Архивировано 9 июня 2020 г. в Wayback Machine John Wiley, 3-е издание, стр. 14. ISBN 9781405144490 .
- ^ Ромер 1970 .
- ^ Jump up to: Перейти обратно: а б Докинз, Ричард (2004). Рассказ предка: паломничество к заре жизни . Бостон: Компания Houghton Mifflin. п. 357. ИСБН 978-0-618-00583-3 .
- ^ Гевин, В. (2005). «Функциональная геномика расширяет биологический сюжет» . ПЛОС Биология . 3 (6): е219. дои : 10.1371/journal.pbio.0030219 . ПМЦ 1149496 . ПМИД 15941356 .
- ↑ Геном ланцетника (амфиоксуса) и происхождение позвоночных. Архивировано 4 марта 2016 г. в Wayback Machine Ars Technica , 19 июня 2008 г.
- ^ Шу, ДГ.; Луо, ХЛ.; Конвей Моррис, С.; Чжан, XL.; Ху, SX.; Чен, Л.; и др. (4 ноября 1999 г.). «Позвоночные животные нижнего кембрия из южного Китая». Природа . 402 (6757): 42–46. Бибкод : 1999Natur.402...42S . дои : 10.1038/46965 . S2CID 4402854 .
- ^ Ваггонер, Бен. «Позвоночные животные: летопись окаменелостей» . УКМП. Архивировано из оригинала 29 июня 2011 года . Проверено 15 июля 2011 г.
- ^ Хейнс, Тим; Чемберс, Пол (2005). Полное руководство по доисторической жизни . Книги Светлячка.
- ^ Британская энциклопедия 1954 , стр. 107.
- ^ Jump up to: Перейти обратно: а б Берг 2004 , стр. 599.
- ^ «агнатан» . Оксфордский словарь английского языка (онлайн-изд.). Издательство Оксфордского университета . (Требуется подписка или членство участвующей организации .)
- ^ Бейкер, Клэр В.Х. (2008). «Эволюция и развитие клеток нервного гребня позвоночных». Текущее мнение в области генетики и развития . 18 (6): 536–543. дои : 10.1016/j.где.2008.11.006 . ПМИД 19121930 .
- ^ Пурнелл, Массачусетс (2001). Дерек Э.Г. Бриггс и Питер Р. Кроутер (ред.). Палеобиология II . Оксфорд : Издательство Блэквелл . п. 401. ИСБН 978-0-632-05149-6 .
- ^ Чжао Вэнь-Цзинь; Чжу Минь (2007). «Диверсификация и фаунистический сдвиг силуро-девонских позвоночных Китая» . Геологический журнал . 42 (3–4): 351–369. Бибкод : 2007GeolJ..42..351W . дои : 10.1002/gj.1072 . S2CID 84943412 . Архивировано из оригинала 5 января 2013 года.
- ^ Сансом, Роберт С. (2009). «Филогения, классификация и полярность признаков Osteostraci (Vertebrata)» . Журнал систематической палеонтологии . 7 (1): 95–115. Бибкод : 2009JSPal...7...95S . дои : 10.1017/S1477201908002551 . S2CID 85924210 . Архивировано из оригинала 19 октября 2012 года . Проверено 16 января 2013 г.
- ^ Маллат, Дж.; Дж. Салливан. 1998. (1998). «Последовательности рибосомальной ДНК 28S и 18S поддерживают монофилию миног и миксин» . Молекулярная биология и эволюция . 15 (12): 1706–1718. doi : 10.1093/oxfordjournals.molbev.a025897 . ПМИД 9866205 .
{{cite journal}}: CS1 maint: числовые имена: список авторов ( ссылка ) - ^ ДеЛарбр Кристиан; Галлут Кирилл; Барриэль Вероник; Жанвье Филипп; Гашелен Габриэль (2002). «Полная митохондриальная ДНК миксины Eptatretus burgeri: сравнительный анализ последовательностей митохондриальной ДНК убедительно подтверждает монофилию круглоротых». Молекулярная филогенетика и эволюция . 22 (2): 184–192. Бибкод : 2002МОЛПЭ..22..184Д . дои : 10.1006/mpev.2001.1045 . ПМИД 11820840 .
- ^ Жанвье, П. 2010. «МикроРНК возрождают старые взгляды на дивергенцию и эволюцию бесчелюстных позвоночных». Труды Национальной академии наук (США) 107:19137-19138. [1] Архивировано 24 сентября 2015 г. в Wayback Machine « Хотя я был одним из первых сторонников парафилии позвоночных, я впечатлен доказательствами, предоставленными Хаймбергом и др., И готов признать, что круглоротые на самом деле монофилетичны. В результате они мало что могут рассказать нам о заре эволюции позвоночных, за исключением того, что интуиция зоологов XIX века была верна, полагая, что эти странные позвоночные (особенно миксины) сильно выродились и утратили многие признаки с течением времени. время. "
- ^ Jump up to: Перейти обратно: а б Бентон, MJ (2005) Палеонтология позвоночных, Блэквелл, 3-е издание, рис. 3.25 на странице 73.
- ^ Жанвье, Филипп (1997) Позвоночные. Животные с позвоночником. Архивировано 12 марта 2013 г. в Wayback Machine . Версия от 1 января 1997 г. в веб-проекте «Древо жизни». Архивировано 15 мая 2011 г. в Wayback Machine.
- ^ Де Ренци М., Будоров К., Судар М. (1996). «Вымирание конодонтов – по дискретным элементам – на границе триаса и юры» . Cuadernos de Geologia Ibérica . 20 : 347–364. Архивировано из оригинала 6 августа 2016 года . Проверено 20 января 2013 г.
- ^ Jump up to: Перейти обратно: а б Габботт, SE; Р. Дж. Олдридж; Дж. Н. Терон (1995). «Гигантский конодонт с сохранившейся мышечной тканью из верхнего ордовика Южной Африки». Природа . 374 (6525): 800–803. Бибкод : 1995Natur.374..800G . дои : 10.1038/374800a0 . S2CID 4342260 .
- ^ Бриггс, Д. (май 1992 г.). «Конодонты: основная вымершая группа, добавленная к позвоночным». Наука . 256 (5061): 1285–1286. Бибкод : 1992Sci...256.1285B . дои : 10.1126/science.1598571 . ПМИД 1598571 .
- ^ Милсом, Клэр; Ригби, Сью (2004). «Позвоночные». Окаменелости с первого взгляда . Виктория, Австралия: Blackwell Publishing. п. 88. ИСБН 978-0-632-06047-4 .
- ^ Донохью, PCJ; Фори, Польша; Олдридж, Р.Дж. (2000). «Сродство конодонтов и филогения хордовых» . Биологические обзоры . 75 (2): 191–251. дои : 10.1111/j.1469-185X.1999.tb00045.x . ПМИД 10881388 . S2CID 22803015 . Архивировано из оригинала 13 апреля 2020 года . Проверено 7 апреля 2008 г.
- ^ Бентон 2005 , с. 44 .
- ^ Мэйси, Джон Г. (1996). Открытие ископаемых рыб (иллюстрированное ред.). Нью-Йорк: Генри Холт и компания. п. 37 . ISBN 9780805043662 .
- ↑ Конструкция челюсти позвоночных заблокирована досрочно. Архивировано 8 сентября 2012 г. в Wayback Machine.
- ^ Дэвис, С.; Финарелли, Дж.; Коутс, М. (2012). «Акантоды и акулоподобные состояния у последнего общего предка современных челюстноротых» . Природа . 486 (7402): 247–250. Бибкод : 2012Natur.486..247D . дои : 10.1038/nature11080 . ПМИД 22699617 . S2CID 4304310 . Архивировано из оригинала 25 марта 2019 года . Проверено 25 марта 2019 г.
- ^ Барфорд, Элиот (25 сентября 2013 г.). «На лице древней рыбы видны корни современной челюсти» . Природа . дои : 10.1038/nature.2013.13823 . S2CID 87470088 . Архивировано из оригинала 31 октября 2013 года . Проверено 26 сентября 2013 г.
- ^ Мередит Смит, Мойя; Кларк, Бретт; Гуже, Даниэль; Йохансон, Зерина (17 августа 2017 г.). «Эволюционное происхождение зубов у челюстных позвоночных: противоречивые данные по зубным пластинкам акантоторацида («Плакодерми»)» . Палеонтология . 60 (6): 829–836. Бибкод : 2017Palgy..60..829M . дои : 10.1111/пала.12318 . hdl : 10141/622339 . ISSN 0031-0239 .
- ^ Бентон, MJ (2005) Палеонтология позвоночных, Блэквелл, 3-е издание, рис. 7.13 на странице 185.
- ^ Чжу, Мин; Сяобо Ю; Пер Эрик Альберг; Брайан Чу; Цзин Лу; Туо Цяо; Цинмин Цюй; Вэньцзинь Чжао; Ляньтао Цзя; Хеннинг Блом; Юань Чжу (2013). «Силурийская плакодерма с остихтианоподобными маргинальными костями челюсти». Природа . 502 (7470): 188–193. Бибкод : 2013Natur.502..188Z . дои : 10.1038/nature12617 . ПМИД 24067611 . S2CID 4462506 .
- ^ Хелфман и др. 2009 , с. 198.
- ^ Аллен, Г.Р., С.Х. Миджли, М. Аллен. Полевое руководство по пресноводным рыбам Австралии. Ред. Ян Найт/Венди Балгин . Перт, Вашингтон: Музей Западной Австралии, 2002. стр. 54–55.
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Амемия, Коннектикут; Альфельди, Дж.; Ли, АП; Фан, С.; Филипп, Х.; МакКаллум, И.; и др. (2013). «Геном африканского целаканта дает представление об эволюции четвероногих» . Природа . 496 (7445): 311–316. Бибкод : 2013Natur.496..311A . дои : 10.1038/nature12027 . ПМЦ 3633110 . ПМИД 23598338 .
- ^ Клак, Дженнифер А. (2012). Завоевывая позиции: происхождение и эволюция четвероногих . Издательство Университета Индианы. ISBN 9780253356758 .
- ^ Нельсон 2006 .
- ^ Йохансон Зерина; Лонг Джон А.; Талант Джон А.; Жанвье Филипп; Уоррен Джеймс В. (2006). «Старейший целакант из раннего девона Австралии» . Письма по биологии . 2 (3): 443–46. дои : 10.1098/rsbl.2006.0470 . ПМК 1686207 . ПМИД 17148426 . Архивировано из оригинала 19 февраля 2013 года.
- ^ Фори, Питер Л. (1998). История рыб целакантов . Лондон: Чепмен и Холл.
- ^ Jump up to: Перейти обратно: а б Знакомство с Actinopterygii. Архивировано 17 февраля 2013 г. в Wayback Machine Палеонтологическом музее Калифорнийского университета.
Источники [ править ]
- Бентон, Майкл Дж. (2005). Палеонтология позвоночных .
- Берг, Линда Р.; Эльдра Перл Соломон; Дайана В. Мартин (2004). Биология . Cengage Обучение. ISBN 978-0-534-49276-2 .
- Британская энциклопедия (1954). Британская энциклопедия: новый обзор универсальных знаний Том. 17.
- Хельфман, Г.; Коллетт, Б.; Фейси, Д.; Боуэн, Б. (2009). Разнообразие рыб: биология, эволюция и экология (2-е изд.). Уайли-Блэквелл. ISBN 978-1-4051-2494-2 . Архивировано из оригинала 26 августа 2021 года . Проверено 26 января 2010 г.
- Нельсон, Джозеф С. (2006). Рыбы мира . Джон Уайли и сыновья . ISBN 978-0-471-25031-9 .
- Ромер, А.С. (1970). Тело позвоночного (4-е изд.). Лондон: ВБ Сондерс.
Дальнейшее чтение [ править ]
| Внешние видео | |
|---|---|
- Бентон, М.Дж. (1998) «Качество летописи окаменелостей позвоночных». Архивировано 25 августа 2012 г. в Wayback Machine. Страницы 269–303 в Доноване, С.К., и Пол, CRC (ред.), Адекватность летописи окаменелостей . Уайли. ISBN 9780471969884 .
- Клотье, Р. (2010). «Ископаемая летопись онтогенеза рыб: понимание закономерностей и процессов развития». Семинары по клеточной биологии и биологии развития . 21 (4): 400–413. дои : 10.1016/j.semcdb.2009.11.004 . ПМИД 19914384 .
- Жанвье, Филипп (1998) Ранние позвоночные , Оксфорд, Нью-Йорк: Издательство Оксфордского университета. ISBN 0-19-854047-7
- Лонг, Джон А. (1996) Возникновение рыб: 500 миллионов лет эволюции Издательство Университета Джона Хопкинса. ISBN 0-8018-5438-5
- Маккензи, диджей; Фаррелл, AP; и Браунер, К.Дж. (2011) Физиология рыб: Академическое издательство примитивных рыб. ISBN 9780080549521 .
- Мэйси, Дж. Г. (1996) Пристанище ископаемых рыб . ISBN 9780805043662 .
- Рядом, Ти Джей; Дорнбург, А.; Эйтан, Род-Айленд; Кек, БП; Смит, В.Л.; Кун, КЛ; Мур, Дж.А.; Цена, SA; Бербринк, штат Форт; Фридман, М. (2013). «Филогения и темпы диверсификации сверхизлучения колючих рыб» . Труды Национальной академии наук . 110 (31): 12738–12743. Бибкод : 2013PNAS..11012738N . дои : 10.1073/pnas.1304661110 . ПМК 3732986 . ПМИД 23858462 .
- Шубин, Нил (2009) Ваша внутренняя рыба: путешествие в 3,5-миллиардную историю человеческого тела . Винтажные книги. ISBN 9780307277459 .
- позвоночных Знакомство с палеонтологическим музеем Калифорнийского университета .
Внешние ссылки [ править ]
- Ископаемая рыба
- Происхождение рыбы
- Обзор эволюции - Карл Саган
- Происхождение позвоночных Марк В. Киршнер , iBioSeminars .
- 150 миллионов лет эволюции рыб в одной удобной фигурке Scientific American , 29 августа 2013 г.
- Музей Age Of Fishes. Архивировано 17 января 2020 года в Wayback Machine , Кановиндра. Это постоянная экспозиция одних из лучших из тысяч окаменелостей, датируемых девонским периодом, найденных поблизости.



| Базы данных органов управления : Национальные |
|---|