Водное дыхание

Водное дыхание — это процесс , при котором водный организм обменивает дыхательные газы с водой , получая кислород из кислорода, растворенного в воде, и выделяя углекислый газ в воду и некоторые другие продукты обмена веществ.
Одноклеточные и простые мелкие организмы
[ редактировать ]У очень мелких животных, растений и бактерий простая диффузия газообразных метаболитов достаточна для дыхательной функции, и не обнаружено никаких специальных приспособлений, способствующих дыханию. Пассивная диффузия или активный транспорт также являются достаточными механизмами для многих крупных водных животных, таких как многие черви , медузы , губки , мшанки и подобные организмы. В таких случаях специфических органов дыхания или органелл не обнаруживается.
Высшие растения
[ редактировать ]Хотя высшие растения обычно используют углекислый газ и выделяют кислород во время фотосинтеза, они также дышат, и, особенно в темноте, многие растения выделяют углекислый газ и нуждаются в кислороде для поддержания нормальных функций. У полностью погруженных в воду высших растений специальные структуры, такие как устьица на поверхности листьев, контролируют газообмен. У многих видов эти структуры можно контролировать и открывать или закрывать в зависимости от условий окружающей среды. В условиях высокой интенсивности освещения и относительно высоких концентраций карбонат-ионов кислород может вырабатываться в количествах, достаточных для образования газовых пузырьков на поверхности листьев, и может вызывать перенасыщение кислородом в окружающем водоеме.
Животные
[ редактировать ]Все животные, практикующие истинно водное дыхание, являются пойкилотермными . Все водные гомойотермные животные и птицы, включая китообразных и пингвинов, дышат воздухом, несмотря на полностью водный образ жизни.
Иглокожие
[ редактировать ]У иглокожих есть специализированная водная сосудистая система , которая выполняет ряд функций, включая обеспечение гидравлической энергии для трубчатых ножек , а также служит для доставки насыщенной кислородом морской воды в организм и вывода сточных вод обратно. У многих родов вода поступает через мадрепорит , ситообразную структуру на верхней поверхности, но может также проникать через реснички в ножках трубки или через специальные решетчатые органеллы. [1]
Моллюски
[ редактировать ]У моллюсков обычно есть жабры, которые позволяют осуществлять обмен дыхательных газов из водной среды в кровеносную систему. У этих животных есть сердце, которое перекачивает кровь, содержащую гемоцианин в качестве молекулы, захватывающей кислород. Дыхательная система брюхоногих моллюсков может включать как жабры, так и легкие.
Членистоногие
[ редактировать ]Водные членистоногие обычно обладают жабрами той или иной формы, в которых газообмен происходит путем диффузии через экзоскелет . Другие могут дышать атмосферным воздухом, оставаясь под водой, через дыхательные трубки или захваченные пузырьки воздуха, хотя некоторые водные насекомые могут оставаться под водой неопределенно долго и дышать с помощью пластрона . У ряда насекомых есть водная ювенильная фаза и взрослая фаза на суше. теряются приспособления к жизни в воде В этих случаях при окончательном шелушении . Ряд отрядов насекомых, таких как подёнки , ручейники и веснянки, имеют водные ювенильные стадии, в то время как некоторые отряды, такие как чешуекрылые, имеют лишь несколько экземпляров, например, китайская моль . Очень немногие паукообразные переняли водный образ жизни, включая паука-колокольчика . Во всех случаях кислород поступает из воздуха, захваченного волосками. [2] вокруг тела животного.
Рыба
[ редактировать ]Большинство рыб обменивают газы с помощью жабр по обе стороны глотки ( горла), образуя спланхнокраниум , часть скелета, где хрящ черепа сходится с хрящом глотки и связанными с ним частями. [3] Жабры представляют собой ткани, состоящие из нитевидных структур, называемых нитями . Эти нити выполняют множество функций и участвуют в переносе ионов и воды, а также в обмене кислорода, углекислого газа, кислоты и аммиака. [4] Каждая нить содержит капиллярную сеть, обеспечивающую большую площадь поверхности для обмена газами и ионами. Рыбы обменивают газы, втягивая через рот богатую кислородом воду и перекачивая ее через жабры. У таких видов, как акула и другие акулы и скаты, дыхальце , которое закачивает воду в жабры, когда животное не движется. в верхней части головы имеется [5] У некоторых рыб капиллярная кровь течет в направлении, противоположном воде, вызывая противоток . Мышцы по бокам глотки выталкивают обедненную кислородом воду через жаберные отверстия. У костистых рыб перекачиванию бедной кислородом воды способствует кость, окружающая жабры, называемая жаберной крышкой . [6]
Земноводные
[ редактировать ]легкие служат и кожа Органами дыхания у амфибий . Кожа этих животных хорошо васкуляризирована и влажная, влажность поддерживается за счет секреции слизи специализированными клетками. Хотя легкие имеют первостепенное значение для контроля дыхания, уникальные свойства кожного дыхания дополняют быстрый газообмен, когда земноводные погружаются в воду, богатую кислородом. [7]
Водные рептилии, птицы и млекопитающие
[ редактировать ]Все водные амниоты ( рептилии , птицы и млекопитающие ) имеют толстые и непроницаемые оболочки , которые препятствуют кожному дыханию , и поэтому для дыхания воздухом они полагаются исключительно на легкие. Находясь под водой, животное, по сути, задерживает дыхание и вынуждено регулярно возвращаться на поверхность, чтобы вдохнуть новый воздух. Следовательно, все амниотные животные, даже те, которые проводят больше времени в воде, чем на улице, могут утонуть , если не могут достичь поверхности, чтобы дышать.
Анатомическая структура легких у рептилий менее сложна, чем у млекопитающих, поскольку у рептилий отсутствует очень обширное бронхиальное дерево , характерное для легких млекопитающих. Газообмен у рептилий еще происходит в альвеолах у рептилий нет , но диафрагмы , поэтому вентиляция происходит за счет изменения объема полости тела, которое , контролируется сокращением межреберных мышц у всех рептилий, кроме черепах . У черепах сокращение определенных пар боковых мышц управляет вдохом или выдохом . [8]
Ныряющие и пелагические морские птицы дышат воздухом с помощью легких, как рептилии и млекопитающие, но легкие птиц представляют собой довольно жесткие структуры, которые не так эластично расширяются и сжимаются. Вместо этого структуры, которые действуют как сильфоны и вентилируют легкие, представляют собой бессосудистые воздушные мешки , которые распределены по большей части тела птиц. [9] и перемещать воздух в одном направлении через парабронхи , где газообмен . происходит [10] [11] Хотя легкие птиц меньше, чем у млекопитающих сопоставимого размера, воздушные мешки составляют 15% от общего объема тела, тогда как у млекопитающих альвеолы ( выполняющие функцию мехов) составляют только 7% от общего объема тела. [12] Как и у их собратьев-рептилий, у птиц также отсутствует диафрагма, и поэтому они полагаются на межреберные и брюшные мышцы , чтобы изменить объем всей торакоабдоминальной полости. Активной фазой дыхания у птиц является выдох, требующий сокращения дыхательных мышц . [11] при этом расслабление этих мышц вызывает вдох.
Жабры
[ редактировать ]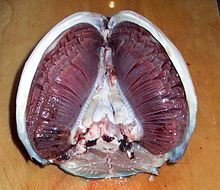
У многих водных животных для дыхания появились жабры, специально приспособленные к их функциям. У рыб, например, есть:
- Большая площадь поверхности позволяет как можно большему количеству кислорода проникать в жабры, поскольку большая часть газа контактирует с мембраной.
- Хорошее кровоснабжение для поддержания необходимого градиента концентрации
- Тонкая мембрана, обеспечивающая короткий путь диффузии.
- Каждая жаберная дуга имеет два ряда (полубранши) жаберных лепестков .
- Каждая жаберная нить имеет множество пластинок.
У остихтиев жабры содержат по 4 жаберные дуги с каждой стороны головы, по две с каждой стороны у хондрихтиев или по семь жаберных корзинок с каждой стороны головы рыбы у миног .У рыб длинный костный покров жабр (жаберная крышка) может использоваться для выталкивания воды. Некоторые рыбы качают воду с помощью жаберной крышки. другие методы, например, таранная вентиляция Без жаберной крышки необходимы . Эту систему используют некоторые виды акул . Когда они плавают, вода течет в рот и через жабры. Поскольку эти акулы используют эту технику, им приходится продолжать плавать, чтобы дышать.
Костные рыбы используют противоток, чтобы максимизировать потребление кислорода, который может диффундировать через жабры. Противоток возникает, когда дезоксигенированная кровь движется через жабры в одном направлении, а насыщенная кислородом вода движется через жабры в противоположном направлении. Этот механизм поддерживает градиент концентрации , тем самым повышая эффективность процесса дыхания и не позволяя уровням кислорода достичь равновесия . У хрящевых рыб нет системы противотока, поскольку у них отсутствуют кости, необходимые для открытия жабр, которые есть у костистых рыб.
Контроль дыхания
[ редактировать ]У рыб нейроны, расположенные в стволе мозга отвечают за генез дыхательного ритма . [13] Положение этих нейронов несколько отличается от центров респираторного генеза млекопитающих, но они расположены в одном и том же отделе мозга, что вызвало споры о гомологии дыхательных центров у водных и наземных видов. И при дыхании водных, и наземных животных точные механизмы, с помощью которых нейроны могут генерировать этот непроизвольный ритм, до сих пор полностью не изучены (см. Непроизвольный контроль дыхания ).
Дыхательный ритм модулируется для адаптации к потреблению кислорода организмом. Как наблюдалось у млекопитающих, рыбы «дышат» быстрее и тяжелее, когда выполняют физические упражнения . Механизмы, с помощью которых происходят эти изменения, являются предметом дискуссий. [14] Эти точки зрения можно классифицировать следующим образом: либо основная часть дыхательных изменений заранее запрограммирована в мозге, что подразумевает, что нейроны центров локомоции мозга соединяются с дыхательными центрами в ожидании движений, либо что большая часть дыхательные изменения возникают в результате обнаружения сокращения мышц, и дыхание адаптируется в результате мышечного сокращения и потребления кислорода. Последняя точка зрения подразумевает, что мозг обладает некими механизмами обнаружения, которые запускают дыхательную реакцию при мышечном сокращении.
Многие теперь согласны с тем, что оба механизма, вероятно, присутствуют и дополняют друг друга или работают вместе с механизмом, который может обнаруживать изменения насыщения крови кислородом и/или углекислым газом.
См. также
[ редактировать ]- Анаэробное дыхание
- Клеточное дыхание
- Контроль дыхания
- Экосистемное дыхание
- Поддержание дыхания
- Дыхательная система
Примечания
[ редактировать ]- ^ Николс, Дэвид (1967). Иглокожие (третье изд.). Библиотека Университета Хатчинсона. п. 44.
- ^ «Получение кислорода» (PDF) . Британское экологическое общество . Великобритания. Апрель 2016.
- ^ «Введение в костную систему» . www.shsu.edu . Проверено 7 июня 2019 г.
- ^ Эванс, Дэвид Х. (18 июня 2010 г.). «Краткая история изучения осморегуляции рыб: центральная роль биологической лаборатории на горе Необитаемый остров» . Границы в физиологии . 1 : 13. doi : 10.3389/fphys.2010.00013 . ISSN 1664-042X . ПМК 3059943 . ПМИД 21423356 .
- ^ Вишницер, Саул (1967). Атлас и руководство по сравнительной анатомии . Соединенные Штаты Америки. п. 22. ISBN 0-7167-0691-1 .
{{cite book}}: CS1 maint: отсутствует местоположение издателя ( ссылка ) - ^ Киммел, Чарльз Б.; Агирре, Виндзор Э.; Ульманн, Бонни; Карри, Марк; Креско, Уильям А. (2008). «Аллометрические изменения сопровождают эволюцию оперкулярной формы у аляскинских трехиглых колюшек». Поведение . 145 (4/5): 669–691. дои : 10.1163/156853908792451395 . ISSN 0005-7959 . JSTOR 40295944 . S2CID 53466588 .
- ^ Готлиб, Г; Джексон, округ Колумбия (1976). «Важность легочной вентиляции для контроля дыхания у лягушки-быка». Am J Physiol . 230 (3): 608–13. дои : 10.1152/ajplegacy.1976.230.3.608 . ПМИД 4976 .
- ^ «рептилия-животное» . Проверено 8 сентября 2016 г.
- ^ Колдер, Уильям А. (1996). Размер, функция и история жизни . Минеола, Нью-Йорк: Публикации Courier Dove. п. 91. ИСБН 978-0-486-69191-6 .
- ^ Майна, Джон Н. (2005). Система легких воздушных мешков развития, строения и функций птиц; с 6 столами . Берлин: Шпрингер. стр. 3.2–3.3 «Легкие», «Система дыхательных путей (бронхиол)» 66–82. ISBN 978-3-540-25595-6 .
- ^ Перейти обратно: а б Краутвальд-Юнгханнс, Мария-Элизабет; и др. (2010). Диагностическая визуализация экзотических домашних животных: птиц, мелких млекопитающих, рептилий . Германия: Издательство Manson. ISBN 978-3-89993-049-8 .
- ^ Уиттоу, Дж. Кози (2000). Птичья физиология Стерки . Сан-Диего, Калифорния: Academic Press. стр. 233–241 . ISBN 978-0-12-747605-6 .
- ^ Рассел, Дэвид Ф. (1986). «Генерация дыхательного паттерна у взрослых миног (Lampetra fluviatilis): интернейроны и сброс всплесков». Журнал сравнительной физиологии А. 158 (1): 91–102. дои : 10.1007/BF00614523 . ПМИД 3723432 . S2CID 19436421 .
- ^ Уолдроп, Тони Г.; Гэри А. Ивамото; Филипп Хаузи (10 ноября 2005 г.). «Точка: Контрапункт: супраспинальные локомоторные центры вносят/не вносят существенного вклада в гиперпноэ при динамических упражнениях». Журнал прикладной физиологии . 100 (3): 1077–1083. doi : 10.1152/japplphysicalol.01528.2005 . ПМИД 16467394 .

