Речная экосистема

Речные экосистемы представляют собой текущие воды, истощающие ландшафт, и включают в себя биотические (живые) взаимодействия между растениями, животными и микроорганизмами, а также абиотические (неживые) физические и химические взаимодействия многих его частей. [1] [2] Речные экосистемы являются частью более крупных сетей водоразделов или водосборов, где меньшие истоки впадают в ручьи среднего размера, которые постепенно впадают в более крупные речные сети. Основные зоны речных экосистем определяются уклоном русла реки или скоростью течения. Быстро движущаяся турбулентная вода обычно содержит более высокие концентрации растворенного кислорода , что поддерживает большее биоразнообразие, чем медленно движущаяся вода в бассейнах. Эти различия лежат в основе разделения рек на горные и равнинные .
Пищевая база ручьев в прибрежных лесах в основном обеспечивается деревьями, но более широкие ручьи и те, у которых нет полога, получают большую часть своей пищевой базы из водорослей. Проходные рыбы также являются важным источником питательных веществ. Экологические угрозы рекам включают потерю воды, плотины, химическое загрязнение и интродуцированные виды . [3] Плотина оказывает негативное воздействие, которое продолжается вниз по водоразделу. Наиболее важными негативными последствиями являются сокращение весенних паводков, наносящих ущерб водно-болотным угодьям, и задержка наносов, что приводит к исчезновению дельтовых водно-болотных угодий. [4]
Речные экосистемы являются яркими примерами лотичных экосистем. Лотик означает текущую воду, от латинского лотоса , что означает омываемый. Лотические воды варьируются от родников шириной всего в несколько сантиметров до крупных рек шириной в километры. [5] Большая часть этой статьи относится к лотичным экосистемам в целом, включая связанные с ними лотичные системы, такие как ручьи и источники . Лотические экосистемы можно противопоставить лентическим экосистемам , которые включают относительно спокойные наземные воды, такие как озера, пруды и водно-болотные угодья . Вместе эти две экосистемы образуют более общую область изучения пресноводной или водной экологии .
Следующие объединяющие характеристики делают экологию проточных вод уникальной среди водных местообитаний: течение однонаправлено, существует состояние непрерывных физических изменений, существует высокая степень пространственной и временной неоднородности во всех масштабах ( микросреды обитания ), изменчивость между Лотические системы довольно высоки, и биота приспособлена к жизни в условиях стока. [6]
Абиотические компоненты (неживые)
[ редактировать ]Неживые компоненты экосистемы называются абиотическими компонентами.Например, камень, воздух, почва и т. д.
Поток воды
[ редактировать ]

Однонаправленный поток воды является ключевым фактором в лотковых системах, влияющим на их экологию. Однако поток потока может быть непрерывным или прерывистым. Речной сток является результатом совокупного воздействия грунтовых вод, осадков и сухопутного стока. Поток воды может варьироваться в зависимости от системы: от проливных порогов до медленных заводей, которые почти похожи на постные системы. Скорость или скорость потока воды в столбе воды также может варьироваться внутри системы и подвержена хаотической турбулентности, хотя скорость воды имеет тенденцию быть самой высокой в средней части русла ручья (известной как тальвег ) . Эта турбулентность приводит к отклонениям потока от среднего вектора потока по склону, что характерно для вихревых токов. Вектор средней скорости потока основан на изменчивости трения о дно или стенки канала, извилистости , препятствиях и градиенте наклона. [5] количество воды, поступающей в систему из-за прямых осадков, талого снега Кроме того, на скорость потока может влиять и/или грунтовых вод. Количество воды в ручье измеряется как расход (объем в единицу времени). По мере того, как вода течет вниз по течению, ручьи и реки чаще всего увеличивают объем воды, поэтому при базовом расходе (т. е. при отсутствии ливневых стоков) более мелкие ручьи в верховьях имеют очень низкий расход, тогда как более крупные реки имеют гораздо более высокий расход. «Режим стока» реки или ручья включает в себя общие закономерности расхода в годовом или десятилетнем масштабе времени и может отражать сезонные изменения стока. [7] [8]
Хотя поток воды во многом определяется уклоном, текущие воды могут изменить общую форму или направление русла ручья — характеристика, также известная как геоморфология . Профиль речной водной толщи состоит из трех основных процессов: эрозии, переноса и отложения. Реки называют «желобами, по которым стекают руины континентов». [9] Реки постоянно размывают , транспортируют и откладывают субстрат, отложения и органический материал. Непрерывное движение воды и увлеченного материала создает множество сред обитания, включая перекаты , спуски и лужи . [10]
Свет
[ редактировать ]Свет важен для лотичных систем, поскольку он обеспечивает энергию, необходимую для стимулирования первичного производства посредством фотосинтеза , а также может служить убежищем для видов-жертв в тени, которую он отбрасывает. Количество света, которое получает система, может быть связано с комбинацией внутренних и внешних переменных потока. Например, территория вокруг небольшого ручья может быть затенена окружающими лесами или стенами долины. Более крупные речные системы, как правило, широкие, поэтому влияние внешних переменных сводится к минимуму, и солнце достигает поверхности. Однако эти реки также имеют тенденцию быть более бурными, и частицы в воде все больше ослабляют свет по мере увеличения глубины. [10] Сезонные и суточные факторы также могут играть роль в доступности света, поскольку угол падения, то есть угол, под которым свет падает на воду, может привести к потере света из-за отражения. Известный как закон Бера , чем меньше угол, тем больше света отражается, а количество получаемой солнечной радиации логарифмически уменьшается с глубиной. [6] Дополнительные влияния на доступность света включают облачный покров, высоту и географическое положение. [11]
Температура
[ редактировать ]


Большинство лотических видов являются пойкилотермными животными , внутренняя температура которых зависит от окружающей среды, поэтому температура является для них ключевым абиотическим фактором. Воду можно нагревать или охлаждать за счет излучения на поверхности и проводимости к воздуху и окружающему субстрату или от него. Мелкие ручьи обычно хорошо перемешиваются и поддерживают относительно однородную температуру на территории. Однако в более глубоких и медленно движущихся водных системах может возникнуть сильная разница между температурой дна и поверхности. Системы родникового питания мало чем отличаются, поскольку родники обычно берут начало из источников подземных вод, температура которых часто очень близка к температуре окружающей среды. [6] Во многих системах наблюдаются сильные суточные колебания, а сезонные колебания наиболее выражены в арктических, пустынных и умеренных системах. [6] Степень затенения, климат и высота над уровнем моря также могут влиять на температуру лотковых систем. [5]
Химия
[ редактировать ]Химический состав воды в речных экосистемах варьируется в зависимости от того, какие растворенные растворенные вещества и газы присутствуют в водной толще ручья. В частности, речная вода может включать, помимо самой воды, [ нужна ссылка ]
- растворенные неорганические вещества и основные ионы (кальций, натрий, магний, калий, бикарбонат, сульфид, хлорид)
- растворенные неорганические питательные вещества (азот, фосфор, кремнезем)
- взвешенные и растворенные органические вещества
- газы (азот, закись азота, углекислый газ, кислород)
- следы металлов и загрязняющих веществ
Растворенные ионы и питательные вещества
[ редактировать ]Растворенные в потоке растворенные вещества можно считать либо реакционноспособными , либо консервативными . Реакционноспособные растворенные вещества легко биологически усваиваются автотрофной и гетеротрофной биотой водотока; примеры могут включать неорганические виды азота, такие как нитрат или аммоний, некоторые формы фосфора (например, растворимый химически активный фосфор) и кремнезем. Другие растворенные вещества можно считать консервативными, что указывает на то, что растворенное вещество не усваивается и не используется биологически; хлорид часто считают консервативным растворенным веществом. Консервативные растворенные вещества часто используются в качестве гидрологических индикаторов движения и переноса воды. Как реактивный, так и консервативный химический состав воды в водосборе в первую очередь определяется геологией водосбора или водосборной площади. На химический состав речной воды также могут влиять осадки и добавление загрязняющих веществ из антропогенных источников. [5] [10] Больших различий в химическом составе обычно не существует в небольших лоточных системах из-за высокой скорости смешивания. Однако в более крупных речных системах концентрации большинства питательных веществ, растворенных солей и pH снижаются по мере увеличения расстояния от истока реки. [6]
Растворенные газы
[ редактировать ]Что касается растворенных газов, кислород , вероятно, является наиболее важным химическим компонентом лотичных систем, поскольку он необходим всем аэробным организмам для выживания. Он попадает в воду в основном за счет диффузии на границе раздела вода-воздух. Растворимость кислорода в воде снижается с увеличением pH и температуры воды. Быстрые, турбулентные потоки подвергают воздействию воздуха большую часть поверхности воды и, как правило, имеют низкие температуры и, следовательно, больше кислорода, чем медленные заводи. [6] Кислород является побочным продуктом фотосинтеза, поэтому системы с высоким содержанием водных водорослей и растений также могут иметь высокие концентрации кислорода в течение дня. Эти уровни могут значительно снизиться ночью, когда первичные продуценты переключаются на дыхание. Кислород может быть лимитирующим, если циркуляция между поверхностными и более глубокими слоями плохая, если активность лотосных животных очень высока или происходит большое количество органического распада. [10]
Взвешенное вещество
[ редактировать ]Реки также могут переносить взвешенные неорганические и органические вещества. Эти материалы могут включать осадок [12] или органическое вещество земного происхождения, попадающее в русло ручья. [13] Часто органическое вещество перерабатывается в ручье посредством механического фрагментирования, потребления и выпаса беспозвоночными, а также микробного разложения. [14] В листьях и древесных остатках можно распознать крупнодисперсное органическое вещество (CPOM) в дисперсное органическое вещество (POM), вплоть до мелкодисперсного органического вещества. Древесные и недревесные растения имеют разную скорость разрушения в потоке: листовые растения или части растений (например, лепестки цветов) разрушаются быстрее, чем древесные бревна или ветки. [15]
Субстрат
[ редактировать ]Неорганический субстрат лотичных систем состоит из геологического материала, присутствующего в водосборе, который размывается, транспортируется, сортируется и откладывается течением. Неорганические субстраты классифицируются по размеру по шкале Вентворта , которая варьируется от валунов до гальки, гравия, песка и ила. [6] Обычно размер частиц субстрата уменьшается вниз по течению, при этом появляются более крупные валуны и камни в более горных районах и песчаное дно в равнинных реках. Это связано с тем, что более высокие уклоны горных рек способствуют более быстрому течению, перемещая более мелкие материалы субстрата дальше вниз по течению для осаждения. [10] Субстрат также может быть органическим и может включать мелкие частицы, осенние листья, крупный древесный мусор, такой как затопленные бревна, мох и околоводные растения. [5] Отложение субстрата не обязательно является постоянным явлением, поскольку во время наводнений оно может претерпевать значительные изменения. [10]
Биотические компоненты (живые)
[ редактировать ]Живые компоненты экосистемы называются биотическими компонентами. В ручьях обитают многочисленные типы биотических организмов, в том числе бактерии, первичные продуценты, насекомые и другие беспозвоночные, а также рыбы и другие позвоночные.
Биопленка
[ редактировать ]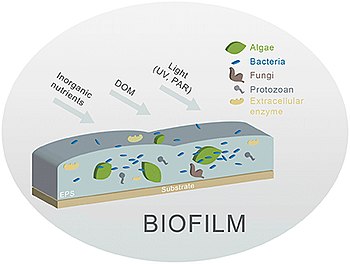
Биопленка представляет собой сочетание водорослей (диатомовых водорослей и т. д.), грибов, бактерий и других мелких микроорганизмов , которые существуют в пленке вдоль русла реки или бентоса . [17] Сами комплексы биопленок сложны. [18] и усложнить русло реки.
Различные компоненты биопленки (основными компонентами являются водоросли и бактерии) встроены в экзополисахаридный матрикс (ЭПС), являются чистыми рецепторами неорганических и органических элементов и остаются подверженными влиянию различных факторов окружающей среды. [16]


Биопленки являются одной из основных биологических промежуточных фаз в речных экосистемах и, вероятно, наиболее важными в перемежающихся реках , где значение водной толщи снижается в течение длительных периодов низкой активности гидрологического цикла . [16] Биопленки можно понимать как микробные консорциумы автотрофов (ЭПС ) и гетеротрофов , сосуществующие в матрице гидратированных внеклеточных полимерных веществ . Эти два основных биологических компонента представляют собой в основном водоросли и цианобактерии, с одной стороны, и бактерии и грибы , с другой. [16] Микро- и мейофауна также населяет биопленку, питаясь организмами и органическими частицами и способствуя ее эволюции и распространению. [20] Таким образом, биопленки образуют высокоактивный биологический консорциум, готовый использовать органические и неорганические материалы из водной фазы, а также готовые использовать световые или химические источники энергии. ЭПС иммобилизует клетки и удерживает их в непосредственной близости, обеспечивая интенсивные взаимодействия, включая межклеточную связь и образование синергических консорциумов. [21] ЭПС способен удерживать внеклеточные ферменты и, следовательно, позволяет использовать материалы из окружающей среды и превращать эти материалы в растворенные питательные вещества для использования водорослями и бактериями. В то же время EPS способствует защите клеток от высыхания, а также от других опасностей (например, биоцидов , УФ-излучения и т. д.) из внешнего мира. [16] С другой стороны, упаковка и защитный слой EPS ограничивают диффузию газов и питательных веществ, особенно для клеток, находящихся далеко от поверхности биопленки, и это ограничивает их выживаемость и создает сильные градиенты внутри биопленки. Как физическая структура биопленки, так и пластичность организмов, живущих в ней, обеспечивают и поддерживают их выживание в суровых условиях окружающей среды или при меняющихся условиях окружающей среды. [16]

Микроорганизмы
[ редактировать ]Бактерии присутствуют в больших количествах в лотковых водах. Свободноживущие формы связаны с разложением органического материала, биопленок на поверхности камней и растительности, между частицами, составляющими субстрат, и взвешенными в толще воды . Другие формы также связаны с кишечником лотосных организмов в качестве паразитов или в комменсальных отношениях. [6] Бактерии играют большую роль в переработке энергии (см. ниже ). [5]
Диатомеи являются одной из основных доминирующих групп перифитных водорослей в лотичных системах и широко используются в качестве эффективных индикаторов качества воды, поскольку они быстро реагируют на изменения окружающей среды, особенно на органическое загрязнение и эвтрофикацию, с широким спектром толерантности к различным условиям: от олиготрофного к эвтрофному. [19] [23] [24]
Первичные производители
[ редактировать ]Водоросли, состоящие из фитопланктона и перифитона , являются наиболее важными источниками первичной продукции в большинстве ручьев и рек. [6] Фитопланктон свободно плавает в толще воды и поэтому не может поддерживать популяцию в быстро текущих потоках. Однако они могут образовывать значительные популяции в медленно текущих реках и заводях. [5] Перифитон обычно представляет собой нитчатые и пучковые водоросли, которые могут прикрепляться к объектам, чтобы их не смыло быстрыми течениями. В местах, где скорость течения незначительна или отсутствует, перифитон может образовывать студенистый незакрепленный плавающий мат. [10]


Растения демонстрируют ограниченную адаптацию к быстрому течению и наиболее успешны при пониженном течении. Более примитивные растения, такие как мхи и печеночники, прикрепляются к твердым предметам. Обычно это происходит в более холодных верховьях, где в основном каменистый субстрат предлагает места для прикрепления. Некоторые растения свободно плавают на поверхности воды плотными ковриками, например ряска или водяной гиацинт . Другие имеют корни и могут быть классифицированы как подводные или надводные. Укоренившиеся растения обычно встречаются на участках с ослабленным течением, где встречаются мелкозернистые почвы. [11] [10] Эти укорененные растения гибкие, с удлиненными листьями, которые оказывают минимальное сопротивление току. [1]
Жизнь в проточной воде может быть полезна для растений и водорослей, поскольку течение обычно хорошо аэрируется и обеспечивает постоянный приток питательных веществ. [10] Эти организмы ограничены потоком, светом, химией воды, субстратом и давлением выпаса. [6] Водоросли и растения важны для лотичных систем как источники энергии, для формирования микросред обитания, защищающих другую фауну от хищников и течения, а также как пищевой ресурс. [11]
Насекомые и другие беспозвоночные
[ редактировать ]До 90% беспозвоночных в некоторых лотичных системах составляют насекомые . Эти виды демонстрируют огромное разнообразие, и их можно встретить почти в каждой доступной среде обитания, включая поверхность камней, глубоко под субстратом в гипорейной зоне , дрейфующие по течению и в поверхностной пленке. [ нужна ссылка ]
Насекомые выработали несколько стратегий проживания в разнообразных потоках лотичных систем. Некоторые избегают мест с сильным течением, населяя субстрат или защищенную сторону скал. У других плоские тела, чтобы уменьшить сопротивление, которое они испытывают при жизни в проточной воде. [25] Некоторые насекомые, такие как гигантский водяной клоп ( Belostomatidae ), избегают наводнений, покидая ручей, когда чувствуют дождь. [26] В дополнение к этому поведению и форме тела, насекомые имеют различные адаптации жизненного цикла , позволяющие им справляться с естественными физическими суровыми условиями речной среды. [27] Некоторые насекомые рассчитывают свои жизненные события в зависимости от того, когда происходят наводнения и засухи. Например, некоторые подёнки синхронизируются, когда они появляются летающими взрослыми особями, с тем, когда в ручьях Колорадо обычно происходит паводок из-за таяния снегов. Другие насекомые не имеют летной стадии и весь свой жизненный цикл проводят в реке.
Как и большинство основных потребителей, лотические беспозвоночные часто во многом полагаются на течение, которое приносит им пищу и кислород. [28] Беспозвоночные играют важную роль как в качестве потребителей, так и в качестве добычи в лотичных системах. [ нужна ссылка ]
К обычным отрядам насекомых, встречающимся в речных экосистемах, относятся Ephemeroptera (также известный как подёнка ), Trichoptera (также известный как ручейник ), Plecoptera (также известный как веснянка ) , Diptera (также известный как настоящая муха ), некоторые виды жесткокрылых и (также известных как жук ), Odonata (группа, в которую входят ) некоторых стрекозы и стрекозы видов полужесткокрылых (также известных как настоящие клопы). [ нужна ссылка ]
Дополнительные таксоны беспозвоночных, характерные для проточных вод, включают моллюсков, таких как улитки , блюда , моллюски , мидии , а также ракообразных, таких как раки , амфиподы и крабы . [10]
Рыбы и другие позвоночные
[ редактировать ]

Рыбы, вероятно, самые известные обитатели лотичных систем. Способность вида рыб жить в проточной воде зависит от скорости, с которой он может плавать, и от продолжительности сохранения этой скорости. Эта способность может сильно различаться у разных видов и зависит от среды обитания, в которой они могут выжить. Непрерывное плавание затрачивает огромное количество энергии, поэтому на полном течении рыбы проводят лишь короткие периоды времени. Вместо этого особи остаются близко ко дну или на берегу, за препятствиями и защищенными от течения, плывя по течению только для того, чтобы накормиться или сменить местонахождение. [1] Некоторые виды приспособились жить только на дне системы, никогда не выходя в открытый поток воды. Эти рыбы сплюснуты в дорсо-вентральном направлении , чтобы уменьшить сопротивление потоку, и часто имеют глаза на макушке, чтобы наблюдать за тем, что происходит над ними. У некоторых также есть сенсорные стволы, расположенные под головой, чтобы помочь в тестировании субстрата. [11]
Лотические системы обычно соединяются друг с другом, образуя путь к океану (родник → ручей → река → океан), и у многих рыб жизненный цикл требует этапов как в пресной, так и в соленой воде. Лосось , например, является анадромным видом, который рождается в пресной воде, но проводит большую часть своей взрослой жизни в океане, возвращаясь в пресную воду только для нереста. Угри — это катадромные виды, которые во взрослом состоянии живут в пресной воде, но мигрируют в океан для нереста. [6]
Другие таксоны позвоночных, населяющие лотичные системы, включают амфибий , таких как саламандры , рептилии (например, змеи, черепахи, крокодилы и аллигаторы), различные виды птиц и млекопитающих (например, выдры , бобры , бегемоты и речные дельфины ). За исключением нескольких видов, эти позвоночные не привязаны к воде, как рыбы, и проводят часть времени в наземных средах обитания. [6] Многие виды рыб важны как потребители и как добыча для упомянутых выше крупных позвоночных. [ нужна ссылка ]
Динамика трофического уровня
[ редактировать ]Концепция трофических уровней используется в пищевых сетях для визуализации способа передачи энергии из одной части экосистемы в другую. [29] Трофическим уровням могут быть присвоены числа, определяющие, насколько далеко организм находится в пищевой цепи .
- Уровень первый: продуценты , растительноподобные организмы, которые производят себе пищу с помощью солнечной радиации, включая водоросли , фитопланктон , мхи и лишайники .
- Второй уровень: консументы , животные организмы, которые получают энергию от поедания производителей, таких как зоопланктон , мелкая рыба и ракообразные .
- Уровень третий: Разрушители , организмы, которые расщепляют мертвые вещества потребителей и производителей и возвращают питательные вещества обратно в систему. Примером являются бактерии и грибы .
Все энергетические транзакции внутри экосистемы происходят от единственного внешнего источника энергии — Солнца. [29] Часть этой солнечной радиации используется производителями (растениями) для превращения неорганических веществ в органические вещества, которые могут использоваться в пищу потребителями (животными). [29] Растения выделяют часть этой энергии обратно в экосистему посредством катаболического процесса. Затем животные потребляют потенциальную энергию, выделяемую производителями. За этой системой следует смерть организма-потребителя, который затем возвращает питательные вещества обратно в экосистему. Это обеспечивает дальнейший рост растений, и цикл продолжается. Разбивка циклов на уровни облегчает экологам понимание экологической преемственности при наблюдении за передачей энергии внутри системы. [29]
Воздействие сверху вниз и снизу вверх
[ редактировать ]
Общая проблема динамики трофического уровня заключается в том, как регулируются ресурсы и производство. [30] Использование и взаимодействие между ресурсами оказывают большое влияние на структуру пищевых сетей в целом. Температура играет роль во взаимодействиях пищевых сетей, включая силы сверху вниз и снизу вверх внутри экологических сообществ. Регулирование снизу вверх в пищевой сети происходит, когда ресурс, доступный в основании или нижней части пищевой сети, увеличивает продуктивность, которая затем поднимается по цепочке и влияет на доступность биомассы для более высоких трофических организмов. [30] Правила «сверху вниз» применяются, когда популяция хищников увеличивается. Это ограничивает доступную популяцию добычи, что ограничивает доступность энергии для более низких трофических уровней в пищевой цепи. Многие биотические и абиотические факторы могут влиять на взаимодействия сверху вниз и снизу вверх. [31]
Трофический каскад
[ редактировать ]Другим примером взаимодействия пищевых сетей являются трофические каскады . Понимание трофических каскадов позволило экологам лучше понять структуру и динамику пищевых сетей внутри экосистемы. [31] Феномен трофических каскадов позволяет ключевым хищникам структурировать всю пищевую сеть с точки зрения того, как они взаимодействуют со своей добычей. Трофические каскады могут вызвать радикальные изменения в потоке энергии внутри пищевой сети. [31] Например, когда верхний или краеугольный хищник поедает организмы, расположенные ниже него в пищевой сети, плотность и поведение добычи изменятся. Это, в свою очередь, влияет на численность организмов, потребляемых дальше по цепочке, что приводит к каскадному снижению трофических уровней. Однако эмпирические данные показывают, что трофические каскады гораздо более распространены в наземных пищевых сетях, чем в водных. [31]
Пищевая цепочка
[ редактировать ]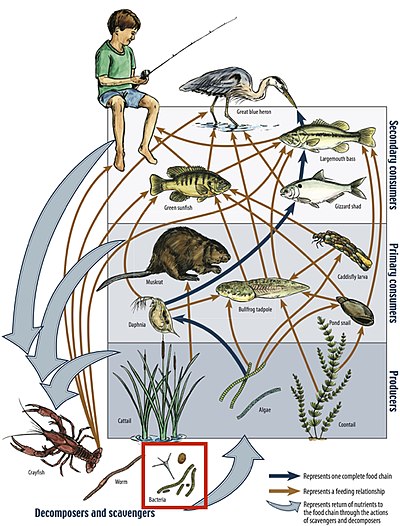
Пищевая цепь — это линейная система звеньев, которая является частью пищевой сети и представляет собой порядок, в котором организмы потребляются от одного трофического уровня к другому. Каждое звено пищевой цепи связано с трофическим уровнем экосистемы. Пронумерованные шаги, которые необходимо сделать, чтобы первоначальный источник энергии, начиная снизу и достигнув вершины пищевой сети, называется длиной пищевой цепи. [33] Хотя длина пищевой цепи может колебаться, водные экосистемы начинаются с первичных производителей, которые потребляются первичными потребителями, которые потребляются вторичными потребителями, а те, в свою очередь, могут потребляться третичными потребителями и так далее, пока не будет достигнута вершина пищевой цепи. достиг. [ нужна ссылка ]
Первичные производители
[ редактировать ]Первичные производители начинают каждую пищевую цепочку. Производство энергии и питательных веществ происходит от солнца посредством фотосинтеза . Водоросли вносят большой вклад в производство энергии и питательных веществ в основании пищевой цепи вместе с наземным мусором , который попадает в ручей или реку . [34] Производство органических соединений, таких как углерод, — это то, что передается вверх по пищевой цепи. Первичные продуценты потребляются травоядными беспозвоночными , которые выступают в роли первичных потребителей . На продуктивность этих продуцентов и функцию экосистемы в целом влияет организм, стоящий над ним в пищевой цепи. [35]
Первичные потребители
[ редактировать ]Первичными консументами являются беспозвоночные и макробеспозвоночные, питающиеся первичными продуцентами. Они играют важную роль в инициировании передачи энергии с базового трофического уровня на следующий. Это регуляторные организмы, которые облегчают и контролируют скорость круговорота питательных веществ и смешивания водных и наземных растительных материалов. [36] Они также транспортируют и сохраняют некоторые питательные вещества и материалы. [36] Существует множество различных функциональных групп этих беспозвоночных, в том числе травоядные, организмы, питающиеся биопленкой водорослей , которая скапливается на затопленных объектах, измельчители, которые питаются крупными листьями и детритом и помогают расщеплять крупный материал. Также фильтраторы , макробеспозвоночные, которые полагаются на поток ручья для доставки мелких твердых частиц органического вещества (FPOM), взвешенных в толще воды , и собиратели, которые питаются FPOM, обнаруженным на дне реки или ручья. [36]
Вторичные потребители
[ редактировать ]Вторичные потребители в речной экосистеме являются хищниками первичных потребителей. Сюда относятся в основном насекомоядные рыбы. [37] Потребление беспозвоночными насекомыми и макробеспозвоночными является еще одним шагом потока энергии вверх по пищевой цепи. В зависимости от своей численности эти хищные потребители могут формировать экосистему тем, как они влияют на трофические уровни ниже них. Когда рыба имеет высокую численность и поедает много беспозвоночных, биомасса водорослей и первичная продукция в ручье больше, а когда вторичные консументы отсутствуют, биомасса водорослей может уменьшаться из-за высокой численности первичных консументов. [37] Энергия и питательные вещества, которые начинаются с первичных производителей, продолжают продвигаться вверх по пищевой цепи и, в зависимости от экосистемы, могут закончиться у этих хищных рыб.
Сложность пищевой сети
[ редактировать ]Разнообразие , продуктивность , видовое богатство , состав и стабильность – все это взаимосвязано серией цепочек обратной связи. Сообщества могут иметь ряд сложных, прямых и/или косвенных ответов на серьезные изменения в биоразнообразии . [35] Пищевые сети могут включать в себя широкий спектр переменных. Три основные переменные, на которые экологи обращают внимание в отношении экосистем, включают видовое богатство, продуктивность биомассы и стабильность /устойчивость к изменениям. [35] Когда вид добавляется или удаляется из экосистемы, это оказывает влияние на оставшуюся пищевую сеть, интенсивность этого эффекта связана со связностью видов и устойчивостью пищевой сети. [38] Когда в речную экосистему добавляется новый вид, интенсивность эффекта связана с устойчивостью или устойчивостью к изменениям существующей пищевой сети. [38] Когда вид удаляется из речной экосистемы, интенсивность эффекта связана с его связью с пищевой сетью. [38] Инвазивные виды могут быть удалены практически без эффекта, но если удалить важных и местных первичных производителей, добычу или хищную рыбу, это может привести к отрицательному трофическому каскаду . [38] Одним из весьма изменчивых компонентов речных экосистем является обеспечение пищей ( биомасса первичных производителей ). [39] Предложение продовольствия или тип производителей постоянно меняются в зависимости от сезона и среды обитания в речной экосистеме. [39] Еще одним весьма изменчивым компонентом речных экосистем является поступление питательных веществ из водно-болотных угодий и наземного детрита . [39] Изменчивость поставок продовольствия и питательных веществ важна для преемственности , устойчивости и связанности организмов речной экосистемы. [39]
Трофические отношения
[ редактировать ]Энергозатраты
[ редактировать ]
Источники энергии могут быть автохтонными и аллохтонными.
- Автохтонные (от латинского «авто» = «собственный») источники энергии — это источники энергии, полученные изнутри лотической системы. во время фотосинтеза Например, первичные продуценты образуют органические соединения углерода из углекислого газа и неорганического вещества. Производимая ими энергия важна для сообщество, поскольку оно может быть переведено на более высокие трофические уровни посредством потребления. Кроме того, высокие темпы первичной продукции могут привести к попаданию растворенных органических веществ (РОВ). в воды [10] Другая форма автохтонной энергии возникает в результате разложения мертвых организмов и фекалий, образующихся внутри лотической системы. В этом случае бактерии разлагают детрит или крупнодисперсный органический материал (CPOM; кусочки >1 мм) на мелкодисперсное органическое вещество (FPOM; кусочки <1 мм), а затем далее на неорганические соединения, необходимые для фотосинтеза. [5] [10] [40] Более подробно этот процесс обсуждается ниже.

- Аллохтонные источники энергии — это те, которые получены извне лотической системы, то есть из земной среды. Листья, ветки, плоды и т. д. являются типичными формами наземного СПОМ, попавшими в воду в результате прямого падения опада или бокового удара листьев. [6] Кроме того, материалы наземного животного происхождения, такие как фекалии или туши, которые были добавлены в систему, являются примерами аллохтонного CPOM. CPOM подвергается специфическому процессу деградации. Аллан приводит пример листа, упавшего в ручей. [5] Во-первых, растворимые химические вещества растворяются и вымываются из листа при его насыщении водой. Это увеличивает нагрузку DOM в системе. Затем микробы, такие как бактерии и грибы, колонизируют лист, размягчая его по мере прорастания в него мицелия гриба. На состав микробного сообщества влияет вид дерева, с которого сбрасываются листья (Rubbo and Kiesecker, 2004). Эта комбинация бактерий, грибов и листьев является источником пищи для измельчения беспозвоночных . [41] которые после потребления оставляют только ФПОМ. Эти мелкие частицы могут снова колонизироваться микробами или служить источником пищи для животных, потребляющих FPOM. Органические вещества также могут попасть в систему лотков уже на стадии FPOM ветром, поверхностным стоком , береговой эрозией или грунтовыми водами . Аналогичным образом, РОВ может попасть в почву через капли дождя или поверхностные потоки. [6]
Беспозвоночные
[ редактировать ]Беспозвоночные могут быть организованы во множество кормовых гильдий в лотичных системах. Некоторые виды являются измельчителями, которые используют большие и мощные части рта для питания недревесным CPOM и связанными с ним микроорганизмами. Другие являются суспензионными питателями , которые используют свои щетинки , фильтрующие аппараты, сети или даже выделения для сбора FPOM и микробов из воды. Эти виды могут быть пассивными коллекторами, использующими естественный поток системы, или они могут генерировать собственный ток для забора воды, а также FPOM в Аллане. [5] Члены гильдии собирателей-сборщиков активно ищут FPOM под камнями и в других местах, где поток реки достаточно ослаб, чтобы позволить осаждение. [10] Пасущиеся беспозвоночные используют приспособления для соскабливания, царапания и просмотра, чтобы питаться перифитоном и детритом . Наконец, некоторые семейства являются хищниками, захватывая и поедая добычу животных. И количество видов, и численность особей внутри каждой гильдии во многом зависят от наличия пищи. Таким образом, эти значения могут варьироваться как в зависимости от сезона, так и в зависимости от системы. [5]
Рыба
[ редактировать ]Рыбу также можно поместить в кормовые гильдии . Планктофаги собирают планктон из толщи воды . Травоядные - детритофаги - виды , питающиеся доном , поедающие без разбора как перифитон , так и детрит . Поверхностные питатели и питатели толщи воды захватывают поверхностную добычу (в основном наземных и вылетающих насекомых) и дрейфуют ( донные беспозвоночные, плывущие вниз по течению). Донные беспозвоночные питаются в основном неполовозрелыми насекомыми, но могут поедать и других донных беспозвоночных. Высшие хищники поедают рыбу и/или крупных беспозвоночных. Всеядные животные поедают разнообразную добычу. Они могут быть цветочными , фаунистическими и/или детритными по своей природе. Наконец, паразиты живут за счет видов-хозяев, обычно других рыб. [5] Рыбы гибко выполняют свои кормовые функции, ловя разную добычу в зависимости от сезона и стадии развития. Таким образом, за свою жизнь они могут состоять в нескольких кормовых гильдиях. Число видов в каждой гильдии может сильно различаться в зависимости от системы: в реках с умеренной теплой водой больше всего донных питающихся беспозвоночных, а в тропических системах имеется большое количество питателей детрита из-за высоких темпов поступления аллохтонных организмов. [10]
Модели и разнообразие сообщества
[ редактировать ]

Местное видовое богатство
[ редактировать ]В крупных реках сравнительно больше видов, чем в мелких ручьях. Многие связывают эту закономерность с увеличением площади и объема более крупных систем, а также с увеличением разнообразия среды обитания. Однако в некоторых системах наблюдается плохое соответствие между размером системы и видовым богатством . сочетание таких факторов, как исторические темпы видообразования и вымирания , тип субстрата , наличие микросреды обитания , химический состав воды, температура и нарушения, такие как наводнения. В этих случаях представляется важным [6]
Разделение ресурсов
[ редактировать ]Хотя было выдвинуто множество альтернативных теорий о способности товарищей по гильдии сосуществовать (см. Morin 1999), разделение ресурсов было хорошо документировано в лотических системах как средство уменьшения конкуренции. Три основных типа разделения ресурсов включают сегрегацию среды обитания, питания и временную сегрегацию. [6]
Было обнаружено, что сегрегация среды обитания является наиболее распространенным типом разделения ресурсов в природных системах (Schoener, 1974). В лотичных системах микросреды обитания обеспечивают такой уровень физической сложности, который позволяет поддерживать разнообразие организмов (Винцин и Хоукнис, 1998). Разделение видов по предпочтениям субстрата хорошо документировано для беспозвоночных. Уорд (1992) смог разделить обитателей субстрата на шесть широких групп, включая тех, кто обитает в грубом субстрате, гравии, песке, грязи, древесных остатках, а также тех, кто связан с растениями, демонстрируя один слой сегрегации. В меньших масштабах дальнейшее разделение среды обитания может происходить на одном субстрате или вокруг него, например куске гравия. Некоторые беспозвоночные предпочитают участки с высоким потоком на открытой поверхности гравия, другие обитают в расщелинах между одним куском гравия и другим, а третьи живут на дне этого куска гравия. [6]
Диетическая сегрегация является вторым по распространенности типом разделения ресурсов. [6] Высокая степень морфологической специализации или поведенческих различий позволяет организмам использовать определенные ресурсы. Например , размер сетей, построенных некоторыми видами беспозвоночных, питающихся взвешенными веществами , может фильтровать из воды частицы FPOM разного размера (Edington et al. 1984). Точно так же члены гильдии пастухов могут специализироваться на сборе водорослей или детрита в зависимости от морфологии их скребковых аппаратов. Кроме того, некоторые виды, похоже, отдают предпочтение определенным видам водорослей. [6]
Временная сегрегация — менее распространенная форма разделения ресурсов, но, тем не менее, это наблюдаемое явление. [6] Обычно это объясняет сосуществование, связывая его с различиями в моделях жизненного цикла и сроками максимального роста среди товарищей по гильдии. Тропические рыбы на Борнео , например, перешли к более короткой продолжительности жизни в ответ на сокращение экологической ниши , которое ощущается по мере увеличения уровня видового богатства в их экосистеме (Watson and Balon 1984).
Постоянство и преемственность
[ редактировать ]В долговременных масштабах наблюдается тенденция к тому, чтобы видовой состав в нетронутых системах оставался в стабильном состоянии. [42] Это было обнаружено как для беспозвоночных, так и для видов рыб. [6] Однако в более коротких временных масштабах изменчивость стока и необычный характер осадков снижают стабильность среды обитания и все это может привести к снижению уровня устойчивости. Способность поддерживать такую устойчивость в течение длительного времени связана со способностью лотических систем относительно быстро возвращаться к исходной конфигурации сообщества после нарушения (Townsend et al. 1987). Это один из примеров временной сукцессии — изменений в сообществе, специфичных для конкретного участка, включающих изменения в видовом составе с течением времени. открывается новая среда обитания Другая форма временной преемственности может возникнуть, когда для колонизации . В этих случаях может утвердиться совершенно новое сообщество, хорошо адаптированное к условиям этой новой территории. [6]
Концепция речного континуума
[ редактировать ]


Концепция речного континуума (RCC) представляла собой попытку создать единую структуру для описания функции лотичных экосистем умеренного пояса от истоков до более крупных рек и связать ключевые характеристики с изменениями в биотическом сообществе (Vannote et al. 1980). [43] Физической основой RCC является размер и расположение вдоль уклона от небольшого ручья, который в конечном итоге переходит в большую реку. Порядок потоков (см. характеристики потоков ) используется как физическая мера положения вдоль RCC.
По данным RCC, низкоупорядоченные участки представляют собой небольшие затененные ручьи, где аллохтонные примеси CPOM являются необходимым ресурсом для потребителей. По мере расширения реки на участках среднего порядка энергозатраты должны меняться. Достаточное количество солнечного света должно достигать дна в этих системах, чтобы поддерживать значительное производство перифитона. Кроме того, ожидается, что биологическая переработка CPOM (крупнодисперсных органических веществ – более 1 мм) на участках выше по течению приведет к переносу больших количеств FPOM (мелкодисперсных органических веществ – менее 1 мм) в эти экосистемы нижнего течения. По мере увеличения размера реки растения должны становиться более многочисленными на берегах реки, особенно в равнинных реках, где откладываются более мелкие отложения, которые способствуют укоренению. В основных каналах, вероятно, слишком сильное течение и мутность, а также недостаток субстрата для поддержания растений или перифитона. Фитопланктон должен производить здесь единственные автохтонные ресурсы, но скорость фотосинтеза будет ограничена из-за мутности и перемешивания. Таким образом, ожидается, что аллохтонные ресурсы станут основным источником энергии для крупных рек. Этот FPOM будет поступать как с участков, расположенных выше по течению, в результате процесса разложения, так и через боковые поступления из пойм рек.
Биота должна измениться вместе с этим изменением энергии от истоков до устья этих систем. А именно, шредеры должны процветать в системах низкого порядка, а травоядные — в системах среднего порядка. Микробное разложение должно играть наибольшую роль в производстве энергии на участках низкого порядка и крупных реках, в то время как фотосинтез, в дополнение к деградированным аллохтонным поступлениям из верхнего течения, будет иметь важное значение в системах среднего порядка. Поскольку участки среднего порядка теоретически будут получать наибольшее разнообразие энергетических затрат, можно ожидать, что они будут содержать наибольшее биологическое разнообразие (Vannote et al. 1980). [5] [6]
Насколько хорошо RCC на самом деле отражает закономерности в природных системах, неясно, и его общность может стать помехой при применении к разнообразным и конкретным ситуациям. [5] Наиболее заметные критические замечания в адрес RCC заключаются в следующем: 1. Он фокусируется в основном на макробеспозвоночных , игнорируя тот факт, что разнообразие планктона и рыб является самым высоким у высших отрядов; 2. Это в значительной степени зависит от того факта, что участки с низким уровнем упорядоченности имеют высокие поступления CPOM, даже несмотря на то, что у многих ручьев отсутствуют прибрежные места обитания; 3. Он основан на древних системах, которые сегодня редко существуют; и 4. Оно сосредоточено на функционировании рек умеренного пояса. Несмотря на свои недостатки, RCC остается полезной идеей для описания того, как закономерности экологических функций в лотической системе могут меняться от истока до устья. [5]
Такие возмущения, как перегруженность плотинами или природные явления, такие как затопление берегов, не включены в модель RCC. [44] С тех пор различные исследователи расширили модель, чтобы учесть такие нарушения. Например, Дж. В. Уорд и Дж. А. Стэнфорд в 1983 году разработали Концепцию последовательного разрыва, которая учитывает влияние геоморфологических нарушений, таких как заторы и интегрированные притоки. Те же авторы в 1993 г. представили концепцию «Гипорейного коридора», в которой были соединены вертикальная (по глубине) и латеральная (от берега к берегу) структурная сложность реки. [45] Концепция пульсации паводка , разработанная У. Дж. Джанк в 1989 г. и далее модифицированная П. Б. Бэйли в 1990 г. и К. Токнером в 2000 г., учитывает большое количество питательных веществ и органического материала, попадающего в реку из отложений окружающих затопленных территорий. земля. [44]
Человеческое воздействие
[ редактировать ]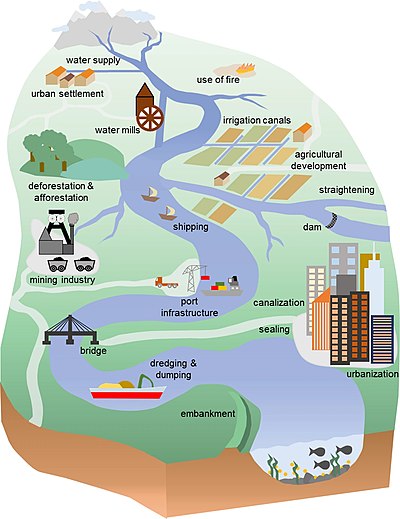
Люди проявляют геоморфическую силу, которая теперь может соперничать с силой естественной Земли. [47] [48] Период доминирования человека был назван антропоценом , и было предложено несколько дат его начала. Многие исследователи подчеркивали драматические изменения, связанные с промышленной революцией в Европе примерно после 1750 г. н.э. (нашей эры) и Великим ускорением развития технологий примерно в 1950 г. н.э. [49] [50] [51] [52] [53]
Однако заметный след человека в окружающей среде насчитывает тысячи лет. [54] [55] [56] [57] а акцент на недавних изменениях сводит к минимуму огромные преобразования ландшафта, вызванные людьми в древности. [58] Важные ранние последствия деятельности человека со значительными последствиями для окружающей среды включают вымирание мегафауны между 14 000 и 10 500 кал. лет назад; [59] одомашнивание растений и животных ближе к началу голоцена , 11 700 кал. лет назад; методы ведения сельского хозяйства и вырубка лесов на уровне 10 000–5 000 кал. лет назад; и широко распространенное образование антропогенных почв около 2000 кал. лет назад. [52] [60] [61] [62] [63] Ключевые свидетельства ранней антропогенной деятельности закодированы в ранних речных последовательностях . [64] [65] , которые усилились антропогенные воздействия за последние столетия и привели к современному всемирному речному кризису. [66] [67] [53]
Загрязнение
[ редактировать ]Загрязнение рек может включать, помимо прочего: увеличение вывоза наносов, избыток питательных веществ из удобрений или городских стоков, [68] канализационные и септические вводы, [69] пластиковое загрязнение , [70] наночастицы, фармацевтические препараты и средства личной гигиены, [71] синтетические химикаты, [72] дорожная соль, [73] неорганические загрязнители (например, тяжелые металлы) и даже тепло в результате тепловых загрязнений. [74] Последствия загрязнения часто зависят от контекста и материала, но могут ухудшить функционирование экосистемы , ограничить экосистемные услуги , уменьшить биоразнообразие рек и повлиять на здоровье человека. [75]
Источники загрязняющих веществ в лотичных системах трудно контролировать, поскольку они могут образовываться, часто в небольших количествах, на очень обширной территории и попадать в систему во многих местах по ее длине. Хотя прямое загрязнение лотковых систем значительно сократилось в Соединенных Штатах в соответствии с правительственным Законом о чистой воде , загрязнения из рассеянных неточечных источников остаются серьезной проблемой. [10] Сельскохозяйственные поля часто доставляют большое количество отложений, питательных веществ и химикатов в близлежащие ручьи и реки. Городские и жилые районы также могут усугубить это загрязнение, когда загрязняющие вещества накапливаются на непроницаемых поверхностях, таких как дороги и парковки, которые затем стекают в систему. Повышенные концентрации питательных веществ, особенно азота и фосфора, которые являются ключевыми компонентами удобрений, могут увеличить рост перифитона, что может быть особенно опасно в медленно текущих потоках. [10] Другой загрязнитель, кислотные дожди , образуется из диоксида серы и закиси азота, выбрасываемых заводами и электростанциями. Эти вещества легко растворяются в атмосферной влаге и попадают в лотковые системы с осадками. Это может снизить pH этих участков, затрагивая все трофические уровни от водорослей до позвоночных. [11] Среднее видовое богатство и общее количество видов в системе уменьшаются с уменьшением pH. [6]
Модификация потока
[ редактировать ]
Изменение стока может произойти в результате строительства плотин , регулирования и отбора воды, модификации русла, разрушения поймы реки и прилегающих прибрежных зон. [76]
Плотины изменяют поток, температуру и режим отложений в лотковых системах. [6] Кроме того, многие реки перекрыты плотинами в нескольких местах, что усиливает воздействие. Плотины могут повысить прозрачность и уменьшить изменчивость речного стока, что, в свою очередь, приводит к увеличению численности перифитона . У беспозвоночных, находящихся непосредственно под плотиной, может наблюдаться сокращение видового богатства из-за общего снижения гетерогенности среды обитания. [10] Кроме того, температурные изменения могут повлиять на развитие насекомых: аномально высокие зимние температуры скрывают признаки нарушения диапаузы яиц , а слишком низкие летние температуры оставляют слишком мало приемлемых дней для завершения роста. [5] Наконец, плотины фрагментируют речные системы, изолируя ранее непрерывные популяции и предотвращая миграцию анадромных и катадромных видов. [10]
Инвазивные виды
[ редактировать ]Инвазивные виды попали в лотковые системы как в результате целенаправленных действий (например, зарыбление дичи и пищевых видов), так и непреднамеренных событий (например, путешествие автостопом на лодках или куликов-рыболовов). Эти организмы могут влиять на местных жителей посредством конкуренции за добычу или среду обитания, хищничества, изменения среды обитания, гибридизации или внедрения вредных болезней и паразитов. [6] После того, как эти виды укоренятся, их будет трудно контролировать или искоренить, особенно из-за взаимосвязанности лотичных систем. Инвазивные виды могут быть особенно вредными в районах, где биота находится под угрозой исчезновения , таких как мидии на юго-востоке США, или в тех, где есть локализованные эндемичные виды, например, лотичные системы к западу от Скалистых гор, где многие виды развивались изолированно.
См. также
[ редактировать ]- Betty's Brain - программное обеспечение, которое «узнает» о речных экосистемах.
- Концепция импульса наводнения
- Экосистема озера
- Реофил
- Прибрежная зона
- Концепция речного континуума
- Речная дренажная система
- РИВПАКС
- Хранители реки
- Горные и равнинные реки
Ссылки
[ редактировать ]- ^ Jump up to: а б с Анжелье, Э. 2003. Экология ручьев и рек. Science Publishers, Inc., Энфилд. Стр. 215.
- ^ «Концепции и связи биологии, шестое издание», Кэмпбелл, Нил А. (2009), страницы 2, 3 и G-9. Проверено 14 июня 2010 г.
- ^ Александр, Дэвид Э. (1 мая 1999 г.). Энциклопедия наук об окружающей среде . Спрингер . ISBN 0-412-74050-8 .
- ^ Кедди, Пол А. (2010). Экология водно-болотных угодий. Принципы и сохранение . Издательство Кембриджского университета. п. 497. ИСБН 978-0-521-51940-3 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п Аллан, JD 1995. Экология потоков: структура и функции проточных вод. Чепмен и Холл, Лондон. Стр. 388.
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С Гиллер, С. и Б. Мальмквист. 1998. Биология ручьев и рек. Издательство Оксфордского университета, Оксфорд. Стр. 296.
- ^ Пофф, Н. Лерой; Уорд, СП (октябрь 1989 г.). «Канадское научное издательство». Канадский журнал рыболовства и водных наук . 46 (10): 1805–1818. дои : 10.1139/f89-228 .
- ^ Пофф, Н. Лерой; Аллан, Дж. Дэвид; Бэйн, Марк Б.; Карр, Джеймс Р.; Престегаард, Карен Л.; Рихтер, Брайан Д.; Спаркс, Ричард Э.; Стромберг, Джули К. (декабрь 1997 г.). «Режим естественного течения» . Бионаука . 47 (11): 769–784. дои : 10.2307/1313099 . ISSN 0006-3568 . JSTOR 1313099 .
- ^ «Речные процессы в геоморфологии» . store.doverpublications.com . Проверено 26 ноября 2018 г.
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р Кушинг, К.Э. и Дж.Д. Аллан. 2001. Ручьи: их экология и жизнь. Академическое издательство, Сан-Диего. Стр. 366.
- ^ Jump up to: а б с д и Браун 1987 г.
- ^ Дин, Дэвид Дж.; Топпинг, Дэвид Дж.; Шмидт, Джон К.; Гриффитс, Рональд Э.; Сабол, Томас А. (январь 2016 г.). «Подача наносов в сравнении с местным гидравлическим контролем транспортировки и хранения наносов в реке с большими нагрузками наносов» . Журнал геофизических исследований: Поверхность Земли . 121 (1): 82–110. Бибкод : 2016JGRF..121...82D . дои : 10.1002/2015jf003436 . ISSN 2169-9003 .
- ^ Вебстер, младший; Мейер, Дж.Л. (1997). «Бюджеты потоков органических веществ: Введение». Журнал Североамериканского бентологического общества . 16 (1): 3–13. дои : 10.2307/1468223 . JSTOR 1468223 . S2CID 88167408 .
- ^ Гесснер, Марк О.; Шове, Эрик; Добсон, Майк (1999). «Перспектива распределения опавших листьев в ручьях | Запросить PDF-файл» . Ойкос . 85 (2): 377. дои : 10.2307/3546505 . JSTOR 3546505 . Проверено 27 ноября 2018 г.
- ^ Вебстер, младший; Бенфилд, EF (ноябрь 1986 г.). «Разрушение сосудистых растений в пресноводных экосистемах». Ежегодный обзор экологии и систематики . 17 (1): 567–594. doi : 10.1146/annurev.es.17.110186.003031 . ISSN 0066-4162 .
- ^ Jump up to: а б с д и ж Сабатер, Сержи; Тимонер, Ксиска; Боррего, Карлес; Акунья, Висенс (2016). «Реакция потоковых биопленок на прерывистость потока: от клеток к экосистемам» . Границы в науке об окружающей среде . 4 . дои : 10.3389/fenvs.2016.00014 . hdl : 10256/16946 . S2CID 18984955 .
 Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 . - ^ Баттин, Том Дж.; Бесемер, Катарина; Бенгтссон, Миа М.; Романи, Анна М.; Пакманн, Аарон И. (апрель 2016 г.). «Экология и биогеохимия ручьевых биопленок» (PDF) . Обзоры природы Микробиология . 14 (4): 251–263. дои : 10.1038/nrmicro.2016.15 . ISSN 1740-1526 . ПМИД 26972916 . S2CID 6573778 .
- ^ Бесемер, Катарина; Питер, Ханнес; Лог, Юрг Б; Лангенхедер, Силке; Линдстрем, Ева С; Транвик, Ларс Дж; Баттин, Том Дж (август 2012 г.). «Распутывание сборки сообществ ручьевых биопленок» . Журнал ISME . 6 (8): 1459–1468. Бибкод : 2012ISMEJ...6.1459B . дои : 10.1038/ismej.2011.205 . ISSN 1751-7362 . ПМК 3400417 . ПМИД 22237539 .
- ^ Jump up to: а б Генрих, Карла Гизельда; Паласиос-Пеньаранда, Марта Л.; Пенья-Саламанка, Энрике; Шуч, Марилия; Лобо, Эдуардо А. (2019). «Эпилитическая диатомовая флора в гидрографическом бассейне реки Кали, Колумбия» . Родригезия . 70 . дои : 10.1590/2175-7860201970041 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Мажди, Набиль; Миале, Бенуа; Бойер, Стефани; Таккс, Мишель; Лефлев, Жозефина; Булетро, Стефани; Тен-Хейдж, Лоик; Жюльен, Фредерик; Фернандес, Роберт; Буффан-Дюбау, Эвелин (2012). «Взаимосвязь между стабильностью эпилитной биопленки и связанной с ней мейофауной при двух типах нарушений наводнений» (PDF) . Наука о пресной воде . 31 : 38–50. дои : 10.1899/11-073.1 . S2CID 3473853 .
- ^ Флемминг, Ганс-Курт; Вингендер, Йост (2010). «Матрица биопленки». Обзоры природы Микробиология . 8 (9): 623–633. дои : 10.1038/nrmicro2415 . ПМИД 20676145 . S2CID 28850938 .
- ^ Лю, Ян; Цюй, Сяодун; Элзер, Джеймс Дж.; Пэн, Вэньци; Чжан, Мин; Рен, Зе; Чжан, Хайпин; Чжан, Юхан; Ян, Хуа (2019). «Влияние градиентов питательных веществ и стехиометрии на микробные сообщества в озере Эрхай и его входных потоках» . Вода . 11 (8): 1711. doi : 10.3390/w11081711 .
- ^ Лобо, Эдуардо А.; Генрих, Карла Гизельда; Шуч, Марилия; Ветцель, Карлос Эдуардо; Эктор, Люк (2016). «Диатомовые водоросли как биоиндикаторы в реках». Речные водоросли . стр. 245–271. дои : 10.1007/978-3-319-31984-1_11 . ISBN 978-3-319-31983-4 .
- ^ Саломони, ЮВ; Роча, О.; Германи, Г.; Лобо, Е.А. (2011). «Применение биологических показателей качества воды с использованием диатомовых водорослей в качестве биоиндикаторов в реке Граватаи, РС, Бразилия» . Бразильский биологический журнал . 71 (4): 949–959. дои : 10.1590/S1519-69842011000500015 .
- ^ Пофф, Н. Лерой; и др. (2006). «Ниши функциональных признаков лотосных насекомых Северной Америки: экологические применения на основе признаков в свете филогенетических взаимоотношений» . Журнал Североамериканского бентологического общества . 25 (4): 730–755. doi : 10.1899/0887-3593(2006)025[0730:FTNONA]2.0.CO;2 . S2CID 18882022 .
- ^ Литл, Дэвид А. (1999). «Использование сигналов о дождях Abedus herberti (Hemiptera: Belostomatidae): механизм предотвращения внезапных наводнений» (PDF) . Журнал поведения насекомых . 12 (1): 1–12. дои : 10.1023/А:1020940012775 . S2CID 25696846 . Проверено 26 ноября 2018 г.
- ^ Литл, Дэвид А.; Пофф, Н. Лерой (2004). «Адаптация к естественным режимам стока». Тенденции экологии и эволюции . 19 (2): 94–100. дои : 10.1016/j.tree.2003.10.002 . ПМИД 16701235 .
- ^ Камминс, Кеннет В.; Клуг, Майкл Дж. (1979). «Экология питания ручьевых беспозвоночных». Ежегодный обзор экологии и систематики . 10 : 147–172. doi : 10.1146/annurev.es.10.110179.001051 . JSTOR 2096788 .
- ^ Jump up to: а б с д Линдеман, Раймонд Л. (1942). «Трофо-динамический аспект экологии» . Экология . 23 (4): 399–417. Бибкод : 1942Экол...23..399Л . дои : 10.2307/1930126 . ISSN 1939-9170 . JSTOR 1930126 .
- ^ Jump up to: а б Хокман, Дэвид (2010). «Усиление тепла: температура влияет на относительную важность эффектов сверху вниз и снизу вверх» . Экология . 91 (10): 2819–2825. Бибкод : 2010Ecol...91.2819H . дои : 10.1890/10-0260.1 . ISSN 1939-9170 . ПМИД 21058543 .
- ^ Jump up to: а б с д Риппл, Уильям Дж.; Эстес, Джеймс А.; Шмитц, Освальд Дж.; Констант, Ванесса; Кейлор, Мэтью Дж.; Ленц, Адам; Мотли, Дженнифер Л.; Селф, Кэтрин Э.; Тейлор, Дэвид С.; Вольф, Кристофер (2016). «Что такое трофический каскад?» . Тенденции в экологии и эволюции . 31 (11): 842–849. дои : 10.1016/j.tree.2016.08.010 . ISSN 0169-5347 . ПМИД 27663836 .
- ^ Шульц, Кестин; Смит, Мария В.; Херфорт, Лиди; Саймон, Холли М. (2018). «Невидимый мир в реке» . Границы для молодых умов . 6 . дои : 10.3389/фрим.2018.00004 . S2CID 3344238 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Пост, Дэвид М. (2002). «Длинная и короткая длина пищевой цепи». Тенденции в экологии и эволюции . 17 (6): 269–277. дои : 10.1016/S0169-5347(02)02455-2 .
- ^ Полис, Гэри А.; Андерсон, Венди Б.; Холт, Роберт Д. (1997). «НА ПУТИ К ИНТЕГРАЦИИ ЛАНДШАФТА И ЭКОЛОГИИ ПИЩЕВОЙ СЕТИ: динамика пространственно субсидируемых пищевых сетей». Ежегодный обзор экологии и систематики . 28 : 289–316. doi : 10.1146/annurev.ecolsys.28.1.289 . HDL : 1808/8160 .
- ^ Jump up to: а б с Червь, Борис; Даффи, Дж. Эммет (2003). «Биоразнообразие, продуктивность и стабильность реальных пищевых сетей». Тенденции в экологии и эволюции . 18 (12): 628–632. дои : 10.1016/j.tree.2003.09.003 .
- ^ Jump up to: а б с Уоллес, Дж. Брюс; Вебстер, Джексон Р. (1996). «Роль макробеспозвоночных в функционировании речной экосистемы» . Ежегодный обзор энтомологии . 41 (1): 115–139. doi : 10.1146/annurev.en.41.010196.000555 . ISSN 0066-4170 . ПМИД 15012327 .
- ^ Jump up to: а б Власть, Мэн (1990). «Влияние рыбы на речные пищевые сети». Наука . 250 (4982): 811–814. Бибкод : 1990Sci...250..811P . дои : 10.1126/science.250.4982.811 . ПМИД 17759974 . S2CID 24780727 .
- ^ Jump up to: а б с д Данн, Дженнифер А.; Уильямс, Ричард Дж.; Мартинес, Нео Д. (2002). «Сетевая структура и потеря биоразнообразия в пищевых сетях: надежность увеличивается по мере связи» . Экологические письма . 5 (4): 558–567. Бибкод : 2002EcolL...5..558D . дои : 10.1046/j.1461-0248.2002.00354.x . ISSN 1461-0248 . S2CID 2114852 .
- ^ Jump up to: а б с д Янг, Мэтью; Хау, Эмили; О'Рир, Тиджей; Берридж, Кэтлин; Мойл, Питер (11 июля 2020 г.). «Топливо пищевой сети различается в зависимости от среды обитания и сезона в приливном пресноводном устье» . Эстуарии и побережья . 44 : 286–301. дои : 10.1007/s12237-020-00762-9 . ISSN 1559-2731 .
- ^ Танк, Дженнифер Л .; Рози-Маршалл, Эмма Дж.; Гриффитс, Натали А.; Энтрекин, Салли А.; Стивен, Миа Л. (март 2010 г.). «Обзор динамики и метаболизма аллохтонного органического вещества в ручьях». Журнал Североамериканского бентологического общества . 29 (1): 118–146. дои : 10.1899/08-170.1 . ISSN 0887-3593 . S2CID 54609464 .
- ^ Агентство по охране окружающей среды США. «Классификации макробеспозвоночных» . Проверено 3 июля 2012 г.
- ^ Хилдрю, AG и П.С. Гиллер. ручья 1994. Пятнистость, взаимодействие видов и нарушение бентоса . В водной экологии: масштабная структура и процесс. П. С. Гиллер, А. Г. Хилдрю и Д. Г. Рафаэлли (ред.). Блэквелл, Оксфорд. Стр. 21–62.
- ^ Ванноте, РЛ (1980). «Концепция речного континуума». Канадский журнал рыболовства и водных наук . 37 : 130–137. дои : 10.1139/f80-017 . S2CID 40373623 – через Canadian Science Publishing.
- ^ Jump up to: а б Джанк Дж. В., П. Б. Бэйли, Р. Э. Спаркс: «Концепция пульсации паводков в системах речных пойм». Канадские специальные публикации по рыболовству и водным наукам. 106. 1989.
- ^ Уорд Дж. В., Дж. А. Стэнфорд: Концепция последовательной неоднородности речных экосистем. Т. Д. Фонтейн, С. М. Бартелл: «Динамика лотических экосистем». Научные публикации, Анн-Арбор, штат Мичиган, 29–42. 1983.
- ^ Маас, Анна-Лиза; Шюттрампф, Хольгер; Лемкуль, Франк (16 октября 2021 г.). «Воздействие человека на речные системы в Европе с особым вниманием к сегодняшнему восстановлению рек» . Науки об окружающей среде Европы . 33 (1). ООО «Спрингер Сайенс энд Бизнес Медиа». дои : 10.1186/s12302-021-00561-4 . ISSN 2190-4707 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Гук, Роджер ЛеБ. (2000). «К истории человека как геоморфного агента». Геология . 28 (9): 843. Бибкод : 2000Geo....28..843L . doi : 10.1130/0091-7613(2000)28<843:OTHOHA>2.0.CO;2 . ISSN 0091-7613 .
- ^ Уилкинсон, Брюс Х. (2005). «Люди как геологические агенты: глубокая перспектива». Геология . 33 (3): 161. Бибкод : 2005Geo....33..161W . дои : 10.1130/G21108.1 .
- ^ Штеффен, Уилл; Крутцен, Пол Дж.; Макнил, Джон Р. (2007). «Антропоцен: люди теперь подавляют великие силы природы». Амбио: журнал о человеческой среде . 36 (8): 614–21. doi : 10.1579/0044-7447(2007)36[614:TAAHNO]2.0.CO;2 . hdl : 1885/29029 . ISSN 0044-7447 . ПМИД 18240674 . S2CID 16218015 .
- ^ Штеффен, Уилл; Бродгейт, Венди; Дойч, Лиза; Гаффни, Оуэн; Людвиг, Корнелия (2015). «Траектория антропоцена: великое ускорение». Обзор антропоцена . 2 (1): 81–98. Бибкод : 2015AntRv...2...81S . дои : 10.1177/2053019614564785 . hdl : 1885/66463 . S2CID 131524600 .
- ^ Уотерс, Китай; Заласевич Дж.; Саммерхейс, К.; Барноский, А.Д.; Пуарье, К.; Га Ушка, А.; Серрета, А.; Эджворт, М.; Эллис, ЕС; Эллис, М.; Жандель, К. ; Лейнфельдер, Р.; Макнил-младший; Рихтер, Д.Д.; Стеффен, В.; Сивицкий, Дж.; Видас, Д.; Вагрейх, М.; Уильямс, М.; Чжишен, А.; Гриневальд, Дж.; Одада, Э.; Орескес, Н. ; Вулф, AP (2016). «Антропоцен функционально и стратиграфически отличается от голоцена». Наука . 351 (6269): аад2622. дои : 10.1126/science.aad2622 . ПМИД 26744408 . S2CID 206642594 .
- ^ Jump up to: а б Крутцен, Пол Дж. (2002). «Геология человечества» . Природа . 415 (6867): 23. Бибкод : 2002Natur.415...23C . дои : 10.1038/415023а . ПМИД 11780095 . S2CID 9743349 .
- ^ Jump up to: а б Гиблинг, Мартин Р. (2018). «Речные системы и антропоцен: хронология человеческого влияния в позднем плейстоцене и голоцене» . Четвертичный период . 1 (3): 21. дои : 10,3390/quat1030021 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Кляйн Голдевейк, Кес; Бойзен, Артур; Ван Дрехт, Жерар; Де Вос, Мартина (2011). «Пространственно выраженная база данных HYDE 3.1 о глобальных изменениях в землепользовании, вызванных деятельностью человека, за последние 12 000 лет». Глобальная экология и биогеография . 20 : 73–86. дои : 10.1111/j.1466-8238.2010.00587.x .
- ^ Кляйн Голдевейк, Кес; Деккер, Стефан С.; Ван Занден, Ян Люитен (2017). «Оценки долгосрочного исторического землепользования на душу населения и последствия для исследований глобальных изменений» . Журнал науки о землепользовании . дои : 10.1080/1747423X.2017.1354938 . S2CID 133825138 .
- ^ Кляйн Голдевейк, Кес; Бойзен, Артур; Доулман, Джонатан; Стефест, Эльке (2017). «Оценки антропогенного землепользования в голоцене - HYDE 3.2» . Данные науки о системе Земли . 9 (2): 927–953. Бибкод : 2017ESSD....9..927K . дои : 10.5194/essd-9-927-2017 .
- ^ Эллис, Эрл К.; Фуллер, Дориан К.; Каплан, Джед О.; Латтерс, Уэйн Г. (2013). «Датирование антропоцена: к эмпирической глобальной истории человеческой трансформации земной биосферы» . Элемента: Наука об антропоцене . 1 : 000018. Бибкод : 2013EleSA...1.0018E . doi : 10.12952/journal.elementa.000018 .
- ^ Раддиман, Уильям Ф.; Эллис, Эрл К.; Каплан, Джед О.; Фуллер, Дориан К. (2015). «Определение эпохи, в которой мы живем». Наука . 348 (6230): 38–39. Бибкод : 2015Sci...348...38R . дои : 10.1126/science.aaa7297 . ПМИД 25838365 . S2CID 36804466 .
- ^ Брадже, Тодд Дж.; Эрландсон, Джон М. (2013). «Ускорение исчезновения животных и растений человеком: континуум позднего плейстоцена, голоцена и антропоцена». Антропоцен . 4 : 14–23. Бибкод : 2013Антр...4...14Б . дои : 10.1016/j.ancene.2013.08.003 .
- ^ Раддиман, Уильям Ф. (2003). «Антропогенная тепличная эра началась тысячи лет назад» . Климатические изменения . 61 (3): 261–293. doi : 10.1023/B:CLIM.0000004577.17928.fa . S2CID 2501894 .
- ^ Чертини, Джакомо; Скаленге, Риккардо (2011). «Антропогенные почвы — золотые шипы антропоцена». Голоцен . 21 (8): 1269–1274. Бибкод : 2011Holoc..21.1269C . дои : 10.1177/0959683611408454 . S2CID 128818837 .
- ^ Раддиман, Уильям; Ваврус, Стив; Куцбах, Джон; Он, Фэн (2014). «Удваивает ли доиндустриальное потепление общий объем антропогенного воздействия?». Обзор антропоцена . 1 (2): 147–153. Бибкод : 2014AntRv...1..147R . дои : 10.1177/2053019614529263 . S2CID 129727301 .
- ^ Эджворт, Мэтт; Деб Рихтер, Дэн; Уотерс, Колин; Хафф, Питер; Нил, Кэт; Прайс, Саймон Джеймс (2015). «Диахронное начало антропоцена: нижняя ограничивающая поверхность антропогенных отложений» (PDF) . Обзор антропоцена . 2 (1): 33–58. Бибкод : 2015AntRv...2...33E . дои : 10.1177/2053019614565394 . S2CID 131236197 .
- ^ Браун, Энтони; Томс, Филипп; Кэри, Крис; Роудс, Эдди (2013). «Геоморфология антропоцена: трансгрессивные во времени разрывы антропогенного аллювиации». Антропоцен . 1 :3–13. Бибкод : 2013Anthr...1....3B . дои : 10.1016/j.ancene.2013.06.002 .
- ^ Браун, Энтони Г.; Зуб, Стивен; Буллард, Джоанна Э.; Томас, Дэвид С.Г.; Чиверрелл, Ричард К.; Плятер, Эндрю Дж.; Мертон, Джулиан; Торндикрафт, Варил Р.; Таролли, Паоло; Роуз, Джеймс; Уэйнрайт, Джон; Даунс, Питер; Аалто, Рольф (2017). «Геоморфология антропоцена: возникновение, состояние и последствия» . Процессы на поверхности Земли и формы рельефа . 42 (1): 71–90. Бибкод : 2017ESPL...42...71B . дои : 10.1002/особ.3943 . hdl : 11577/3199318 . S2CID 55224176 .
- ^ Вонг, К.М., Уильямс, К.Э., Коллиер, У., Шелле, П. и Питток, Дж. (2007) 10 крупнейших рек мира, находящихся под угрозой, Всемирный фонд дикой природы .
- ^ Вёрёсмарти, CJ; Макинтайр, ПБ; Гесснер, Миссури; Даджен, Д.; Прусевич А.; Грин, П.; Глидден, С.; Банн, Швеция; Салливан, Калифорния; Лиерманн, К. Рейди; Дэвис, премьер-министр (2010). «Глобальные угрозы водной безопасности человека и биоразнообразию рек» . Природа . 467 (7315): 555–561. Бибкод : 2010Natur.467..555V . дои : 10.1038/nature09440 . ПМИД 20882010 . S2CID 4422681 .
- ^ Бьелло, Дэвид. «Сток удобрений переполняет ручьи и реки, создавая обширные «мертвые зоны» » . Научный американец . Проверено 27 ноября 2018 г.
- ^ Университет штата Мичиган. «Септики не защищают реки и озера от фекалий» . МГУ сегодня . Проверено 27 ноября 2018 г.
- ^ «Растущая обеспокоенность по поводу пластикового загрязнения рек и озер» . Морская исполнительная власть . Проверено 27 ноября 2018 г.
- ^ Колпин, Дана В.; Ферлонг, Эдвард Т.; Мейер, Майкл Т.; Турман, Э. Майкл; Заугг, Стивен Д.; Барбер, Ларри Б.; Бакстон, Герберт Т. (март 2002 г.). «Фармацевтические препараты, гормоны и другие органические загрязнители сточных вод в реках США, 1999–2000 годы: национальная разведка» . Экологические науки и технологии . 36 (6): 1202–1211. Бибкод : 2002EnST...36.1202K . дои : 10.1021/es011055j . ISSN 0013-936X . ПМИД 11944670 .
- ^ Бернхардт, Эмили С; Рози, Эмма Дж; Гесснер, Марк О (24 января 2017 г.). «Синтетические химикаты как агенты глобальных изменений». Границы в экологии и окружающей среде . 15 (2): 84–90. Бибкод : 2017FrEE...15...84B . дои : 10.1002/плата.1450 . ISSN 1540-9295 . S2CID 90620297 .
- ^ «Реки в США становятся более солеными – и это происходит не только из-за ухода за дорогами зимой» . Проверено 27 ноября 2018 г.
- ^ «Рейтинг рек, загрязненных жарой» . Природа . 538 (7626): 431. Октябрь 2016 г. doi : 10.1038/538431d . ISSN 0028-0836 . ПМИД 27786199 .
- ^ «Загрязнение воды: все, что вам нужно знать» . НРДЦ . Проверено 27 ноября 2018 г.
- ^ «Измененный расход воды (гидрология)» . НИВА . 6 июля 2009 г. Проверено 27 ноября 2018 г.
Дальнейшее чтение
[ редактировать ]- Браун, А.Л. 1987. Экология пресной воды. Образовательные книги Heinimann, Лондон. С. 163.
- Карлайл, DM и MD Вудсайд. 2013. Экологическое здоровье рек страны , Геологическая служба США . П. 6.
- Эдингтон, Дж. М., Эдингтон, Массачусетс, и Дж. А. Дорман. 1984. Распределение среды обитания среди личинок гидрофизхид малайзийского ручья. Энтомология 30: 123–129.
- Хайнс, HBN 1970. Экология проточных вод. Первоначально опубликовано в Торонто издательством University of Toronto Press, 555 стр.
- Морин, П.Дж. 1999. Экология сообщества. Блэквелл Сайенс, Оксфорд. С. 424.
- Власть, Мэн (1990). «Влияние рыбы на речные пищевые сети» (PDF) . Наука . 250 (4982): 811–814. Бибкод : 1990Sci...250..811P . дои : 10.1126/science.250.4982.811 . ПМИД 17759974 . S2CID 24780727 . Архивировано из оригинала (PDF) 24 мая 2013 г.
- Руббо, MJ; Кизекер, Дж. М. (2004). «Состав листовой подстилки и структура сообщества: перевод региональных изменений видов в местную динамику» . Экология . 85 (9): 2519–2525. Бибкод : 2004Ecol...85.2519R . дои : 10.1890/03-0653 .
- Шонер, Т.В. (1974). «Распределение ресурсов в экологических сообществах». Наука . 2 (4145): 369–404. Бибкод : 1974Sci...185...27S . дои : 10.1126/science.185.4145.27 . ПМИД 17779277 . S2CID 43846597 .
- Таунсенд, Чехия; Хилдрю, AG; Шофилд, К. (1987). «Сохранение сообществ речных беспозвоночных в зависимости от изменчивости окружающей среды». Экология животных . 56 (2): 597–613. Бибкод : 1987JAnEc..56..597T . дои : 10.2307/5071 . JSTOR 5071 .
- Ванноте, РЛ; Миншалл, ГВ; Камминс, КВт; Седелл, младший; Кушинг, CE (1980). «Концепция речного континуума». Канадский журнал рыболовства и водных наук . 37 : 130–137. дои : 10.1139/f80-017 . S2CID 40373623 .
- Винсон, MR; Хокинс, КП (1998). «Биоразнообразие речных насекомых: вариации в местном, бассейновом и региональном масштабах». Ежегодный обзор энтомологии . 43 : 271–293. дои : 10.1146/annurev.ento.43.1.271 . ПМИД 15012391 .
- Уорд, СП 1992. Экология водных насекомых: биология и среда обитания. Уайли, Нью-Йорк. С. 456.
- Уотсон, диджей; Балон, ЭК (1984). «Экоморфологический анализ таксоценов рыб в ручьях тропических лесов северного Борнео». Журнал биологии рыб . 25 (3): 371–384. Бибкод : 1984JFBio..25..371W . дои : 10.1111/j.1095-8649.1984.tb04885.x .
Внешние ссылки
[ редактировать ]

| Базы данных органов управления : Национальные |
|---|