Модели распределения ниши
Механистические модели распределения ниш — это биологические модели, используемые для объяснения распределения относительной численности видов . Эти модели распределения ниш описывают, как виды распределяют пул ресурсов в многомерном пространстве, определяя распределение численности особей между видами. Относительная численность видов обычно выражается в виде графика Уиттекера или графика ранговой численности, где виды ранжируются по количеству особей по оси X, а по оси Y - логарифм относительной численности каждого вида. Относительную численность можно измерить как относительное количество особей внутри вида или относительную биомассу особей внутри вида.
История
[ редактировать ]Модели распределения ниш были разработаны потому, что экологи искали биологические объяснения относительного распределения численности видов . Макартур (1957, 1961), [1] [2] был одним из первых, кто выразил недовольство чисто статистическими моделями , представив вместо этого три модели механистического распределения ниш. Макартур считал, что экологические ниши в ресурсном пуле можно разбить, как палку, причем каждый кусок палки представляет собой нишу, занятую в сообществе. При участии Сугихары (1980), [3] Токеши (1990, 1993, 1996) [4] [5] [6] расширил модель сломанной палки, создав примерно семь моделей механистического распределения ниш. Эти механистические модели служат полезной отправной точкой для описания видового состава сообществ.
Описание
[ редактировать ]
Модель распределения ниш может использоваться в ситуациях, когда один пул ресурсов последовательно или одновременно разбивается на более мелкие ниши в результате колонизации видов или видообразования (пояснение об использовании ресурсов: виды внутри гильдии используют одни и те же ресурсы, в то время как виды внутри сообщества не могут ).
Эти модели описывают, как виды, использующие один и тот же пул ресурсов (например, гильдия (экология) ), делят свою нишу. Ресурсный пул разрушается либо последовательно, либо одновременно, и две составляющие процесса фрагментации ниши включают в себя выбор выбранного фрагмента и размер полученного фрагмента (рис. 2).
Модели распределения ниш использовались в основной литературе для объяснения и описания изменений в распределении относительной численности разнообразных таксонов, включая пресноводных насекомых, рыб, жуков-мохообразных, паразитов перепончатокрылых, скоплений планктона и солончаковой травы.
Предположения
[ редактировать ]
Механистические модели, описывающие эти графики, работают в предположении, что графики ранговой численности основаны на строгой оценке численности особей внутри вида и что эти меры отражают фактическое распределение численности видов. Более того, независимо от того, используется ли количество особей в качестве меры численности или биомасса особей, эти модели предполагают, что эта величина прямо пропорциональна размеру ниши, занимаемой организмом. Одно из предположений состоит в том, что численность, измеряемая как количество особей, может иметь меньшие отклонения, чем те, которые используют биомассу . Таким образом, некоторые исследования, использующие изобилие в качестве показателя распределения ниш, могут однородность сообщества переоценивать . Это происходит потому, что не существует четкого различия между размером тела, численностью (экологией) и использованием ресурсов . Часто в исследованиях не удается включить размерную структуру или оценки биомассы в показатели фактической численности, и эти меры могут создать более высокую дисперсию моделей распределения ниш, чем численность, измеряемая строго как количество особей. [7] [8]
Механистические модели распределения ниш Токеши
[ редактировать ]Ниже описаны семь механистических моделей, описывающих распределение ниш. Модели представлены в порядке возрастания равномерности: от наименее четной модели «Преимущество доминирования» до наиболее равномерной модели «Распад доминирования» и «Фракция Макартура».
Преимущество доминирования
[ редактировать ]Эта модель описывает ситуацию, когда после первоначальной колонизации (или видообразования) каждый новый вид занимает более 50% наименьшей оставшейся ниши. В модели распределения ниш с упреждением доминирования виды колонизируют случайную часть от 50 до 100% наименьшей оставшейся ниши, что делает эту модель стохастической по своей природе. Близкородственная модель, Геометрическая серия, [9] представляет собой детерминированную версию модели упреждения доминирования, в которой процент оставшегося пространства ниши, которое занимает новый вид (k), всегда один и тот же. Фактически, модели упреждения доминирования и геометрические ряды концептуально схожи и будут давать одинаковое распределение относительной численности, когда доля заполненной меньшей ниши всегда равна 0,75. Модель упреждения доминирования лучше всего подходит для распределения относительной численности некоторых сообществ речных рыб в Техасе , включая некоторые таксономические группы и конкретные функциональные группы. [10]
Геометрический (k=0,75)


Случайный ассортимент
[ редактировать ]В модели случайного ассортимента пул ресурсов случайным образом делится между одновременно или последовательно колонизирующими видами. Эта закономерность может возникнуть из-за того, что показатель численности не зависит от количества ниш, занимаемых видом, или потому, что временные изменения в численности вида или ширине ниши вызывают разрыв в распределении ниш с течением времени, и, таким образом, виды, по-видимому, не имеют связи между степенью занятости вида. и их ниша. Токеши (1993) [5] объяснил, что эта модель во многом похожа на нейтральную теорию биоразнообразия Касвелла, главным образом потому, что виды, по-видимому, действуют независимо друг от друга.
Случайная дробь
[ редактировать ]Модель случайной фракции описывает процесс, в котором размер ниши выбирается случайным образом путем последовательной колонизации видов. Первоначальный вид выбирает случайную часть общей ниши, а последующие виды-колонизаторы также выбирают случайную часть общей ниши и делят ее случайным образом до тех пор, пока все виды не колонизируются. Токеши (1990) [4] обнаружили, что эта модель совместима с некоторыми эпифитными сообществами креветок-хиромонид, а в последнее время она использовалась для объяснения относительного распределения численности сообществ фитопланктона, растительности соляных лугов, некоторых сообществ насекомых отряда двукрылых, некоторых сообществ жужелиц, функциональных и таксономические группы речных рыб биорегионов Техаса и паразитоидов ихневмонид. Похожая модель была разработана Сугихарой в попытке дать биологическое объяснение логарифмически нормальному распределению Престона (1948). [11] Сугихара (1980) [3] Модель с фиксированным делением была похожа на модель случайных фракций, но случайность модели основана на треугольном распределении со средним значением 0,75, а не на нормальном распределении со средним значением 0,5, используемом в случайной дроби. Сугихара использовал треугольное распределение для рисования случайных величин, поскольку случайность некоторых природных популяций соответствует треугольному распределению со средним значением 0,75.
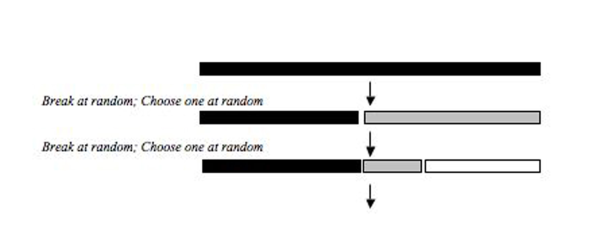
Мощность фракции
[ редактировать ]Эта модель может объяснить распределение относительной численности, когда вероятность колонизации существующей ниши в пуле ресурсов положительно связана с размером этой ниши (измеряемой как численность, биомасса и т. д.). Вероятность колонизации части ниши зависит от относительных размеров установленных ниш и масштабируется показателем степени k. k может принимать значение от 0 до 1, и если k>0, всегда существует несколько более высокая вероятность того, что большая ниша будет колонизирована. Эта модель считается более реалистичной с биологической точки зрения, поскольку можно представить множество случаев, когда ниша с большей долей ресурсов будет с большей вероятностью захвачена, поскольку в этой нише больше ресурсного пространства и, следовательно, больше возможностей для приобретения. Модель распределения ниш случайными дробями представляет собой крайнюю форму модели доли власти, где k = 0, а другой крайний вариант доли власти, когда k = 1, напоминает модель фракции Макартура, где вероятность колонизации прямо пропорциональна размеру ниши. [6] [12]

фракция Макартура
[ редактировать ]Эта модель требует, чтобы начальная ниша была разбита случайным образом, а последующие ниши выбирались с вероятностью, пропорциональной их размеру. В этой модели самая большая ниша всегда имеет большую вероятность быть разрушенной по сравнению с меньшими нишами в пуле ресурсов. Эта модель может привести к более равномерному распределению, при котором более крупные ниши с большей вероятностью будут разрушены, что будет способствовать сосуществованию между видами в нишах одинакового размера. Основой модели «Фракция Макартура» является «Сломанная палка», разработанная Макартуром (1957). Эти модели дают схожие результаты, но одно из основных концептуальных отличий заключается в том, что в модели «Сломанная палка» ниши заполняются одновременно, а не последовательно, как во «Фракции Макартура». Токеши (1993) [5] утверждает, что последовательное вторжение в ресурсный пул более реалистично с биологической точки зрения, чем одновременное разрушение нишевого пространства. Когда обилие рыб из всех биорегионов Техаса было объединено, распределение напоминало модель распределения ниш сломанной палки, что предполагает относительно равномерное распределение видов пресноводных рыб в Техасе. [10]

Распад доминирования
[ редактировать ]Эту модель можно рассматривать как обратную модель упреждения доминирования. Во-первых, первоначальный пул ресурсов колонизируется случайным образом, а оставшиеся, последующие колонизаторы всегда колонизируют самую большую нишу, независимо от того, колонизирована она уже или нет. Эта модель создает наиболее равномерное сообщество по сравнению с описанными выше моделями распределения ниш, поскольку самая большая ниша всегда разбивается на два меньших фрагмента, которые с большей вероятностью будут эквивалентны размеру меньшей ниши, которая не была разбита. Сообщества такого «уровня» выравненности, по-видимому, редки в природных системах. Однако одно из таких сообществ включает относительное распределение численности фильтраторов на одном участке реки Дунай в Австрии. [13]
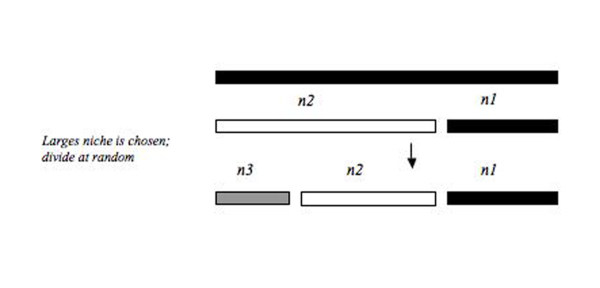
Композитный
[ редактировать ]Составная модель существует, когда комбинация моделей распределения ниш действует в разных частях пула ресурсов. Фесл (2002). [13] показывает, как комплексная модель может выглядеть при изучении пресноводных двукрылых , поскольку разные модели распределения ниш соответствуют разным функциональным группам данных. Другой пример, приведенный Хиггинсом и Штраусом (2008), моделирующий рыбные комплексы, показал, что рыбные сообщества из разных местообитаний и с разным видовым составом соответствуют разным моделям распределения ниш, таким образом, весь видовой комплекс представляет собой комбинацию моделей в разных регионах ареала вида. .
Соответствие механистических моделей распределения ниш эмпирическим данным
[ редактировать ]Механистические модели распределения ниш предназначены для описания сообществ. Исследователи использовали эти модели разными способами для изучения временных и географических тенденций численности видов.
В течение многих лет подбор моделей распределения ниш проводился на глаз, а графики моделей сравнивались с эмпирическими данными. [5] Совсем недавно были разработаны статистические тесты соответствия моделей распределения ниш эмпирическим данным. [14] [15] Более поздний метод (Этьен и Олльф, 2005 г.) [15] использует байесовское моделирование моделей для проверки их соответствия эмпирическим данным. Первый метод, который до сих пор широко используется, моделирует ожидаемую относительную численность на основе нормального распределения каждой модели с учетом того же количества видов, что и эмпирические данные. Каждая модель моделируется несколько раз, и можно рассчитать среднее и стандартное отклонение, чтобы назначить доверительные интервалы для каждого распределения относительной численности. Достоверность каждого ранга можно проверить на основе эмпирических данных для каждой модели, чтобы определить ее соответствие. Доверительные интервалы рассчитываются следующим образом. [4] [12] Дополнительную информацию о моделировании моделей распределения ниш можно найти на веб-сайте [1] [ постоянная мертвая ссылка ] , который объясняет работу программы Power Niche. [14]

r = доверительный предел смоделированных данных σ = стандартное отклонение смоделированных данныхn = количество повторов эмпирической выборки
Ссылки
[ редактировать ]- ^ Макартур, Р.Х. (1957). Об относительном обилии видов птиц. Учеб. Натл. акад. наук. 43, 293-295.
- ^ Макартур, Р. Х. Макартур, JW (1961). О видовом разнообразии птиц. Экология. 42, 594–598.
- ^ Jump up to: а б Сугихара, Г. (1980). Минимальная структура сообщества: объяснение закономерностей обилия видов. Являюсь. Нат. 116. 770-787.
- ^ Jump up to: а б с Токеши, М. (1990). Распределение ниш или случайный ассортимент: новый взгляд на структуру обилия видов. Журнал экологии животных. 59, 1129–1146.
- ^ Jump up to: а б с д Токеши, М. (1993). Структура видового обилия и структура сообщества. Адв. Экол. Рез. 24, 112–186.
- ^ Jump up to: а б Токеши, М. (1996). Power Fraction: новое объяснение закономерностей обилия видов в богатых видами сообществах. Ойкос. 75, 543-550.
- ^ Гастон, К.В. Блэкберн, TM (2000). Макроэкология, Оксфорд, Великобритания: Blackwell Science.
- ^ Тапер, М.Л. Марке, Пенсильвания (1996). Как виды на самом деле делят ресурсы? Американский натуралист. 147, 1072–1086.
- ^ Мэй, РМ (1975). Закономерности видового обилия и разнообразия. В экологии и эволюции сообществ. 81-120, Кембридж, Массачусетс: Издательство Гарвардского университета.
- ^ Jump up to: а б Хиггинс, К.Л. Штраус, Р.Э. (2008). Моделирование сообществ речных рыб с помощью моделей распределения ниш: закономерности. процессы и масштабная зависимость. Труды Американского общества рыболовства. 137, 696-706.
- ^ Престион, FW (1948). Распространенность и редкость видов. Экология. 29, 254–253.
- ^ Jump up to: а б Магуррран, А.Е. (2004). Измерение биологического разнообразия. Оксфорд, Великобритания: Blackwell Science
- ^ Jump up to: а б Фесл, К. (2002). Модели нишево-ориентированного обилия видов: различные подходы и их применение к сообществам личинок хирономид (Diptera) в крупной реке. Дж. Аним. Экол. 71, 1085–1094.
- ^ Jump up to: а б Дрозд, П. Новотный, В. (2000). Ниша власти: модели разделения ниш для анализа сообщества. Версия 1. Руководство и программа опубликованы на сайте www.entu.cas/png/PowerNiche.
- ^ Jump up to: а б Этьен, Р. С. Олльф, Х. (2005). Сопоставление различных моделей структуры сообщества с данными о численности видов: сравнение байесовской модели. Экологические письма. 8, 493–504.