Холодное просачивание
| Морская среда обитания |
|---|
 |
| Прибрежные места обитания |
| Поверхность океана |
| Открытый океан |
| Морское дно |
Холодное просачивание (иногда называемое холодным жерлом ) — это область дна океана , где происходит просачивание жидкостей, богатых сероводородом , метаном и другими углеводородами , часто в виде бассейна с рассолом . Холод не означает, что температура просачивания ниже, чем температура окружающей морской воды; наоборот, его температура часто несколько выше. [1] «Холод» относится к очень теплым (не менее 60 ° C или 140 ° F) условиям гидротермальных источников . Холодные просачивания представляют собой биом, поддерживающий несколько эндемичных видов.
развивают уникальную топографию Холодные просачивания со временем , где реакции между метаном и морской водой создают карбонатные скальные образования и рифы . Эти реакции также могут зависеть от активности бактерий. Икаит , водный карбонат кальция, может быть связан с окислением метана на холодных выходах.
Типы
[ редактировать ]
Типы холодных выходов можно разделить в зависимости от глубины: мелкие холодные выходы и глубокие холодные выходы. [2] Холодные просачивания также можно выделить в деталях следующим образом:
- нефти /газа просачивание [2]
- просачивание газа: [2] метана просачивание
- гидрат газа просачивается [2]
- рассол просачивается [2] образуются в соляных лужах
- оспины [2]
- грязевые вулканы [2]
Формирование и экологическая преемственность
[ редактировать ]Холодные просачивания происходят над трещинами на морском дне, вызванными тектонической активностью. Нефть и метан «просачиваются» из этих трещин, рассеиваются осадками и выходят на территорию шириной в несколько сотен метров. [3]
Метан ( CH
4 ) является основным компонентом природного газа . [3] Но помимо того, что метан является важным источником энергии для людей, он также составляет основу экосистемы холодного просачивания . [3] холодного просачивания Биота на глубине ниже 200 м (660 футов) обычно демонстрирует гораздо большую систематическую специализацию и зависит от хемоавтотрофии , чем биота с глубин шельфа. [4] Осадки глубоководных высачиваний весьма неоднородны. [4] Они поддерживают различные геохимические и микробные процессы, которые отражаются в сложной мозаике местообитаний, населенных смесью специализированной ( гетеротрофной и симбионтной ) и фоновой фауны. [4]
Хемосинтетические сообщества
[ редактировать ]
Биологические исследования в холодных просачиваниях и гидротермальных жерлах были в основном сосредоточены на микробиологии и выдающихся макробеспозвоночных, питающихся за счет хемосинтезирующих микроорганизмов. [2] Гораздо меньше исследований проведено в отношении более мелкой донной фракции при размерах мейофауны (<1 мм). [2]
Упорядоченный переход состава сообщества от одного набора видов к другому называется экологической преемственностью . [3]
Первым типом организмов, воспользовавшимся преимуществами этого глубоководного источника энергии, являются бактерии . [3] Агрегируясь в бактериальные маты на холодных просачиваниях, эти бактерии метаболизируют метан и сероводород (еще один газ, который выходит из просачиваний) для получения энергии. [3] Этот процесс получения энергии из химических веществ известен как хемосинтез . [3]

На этом начальном этапе, когда метана относительно много, мидий . вблизи холодного просачивания также образуются плотные заросли [3] Эти мидии , состоящие в основном из видов рода Bathymodiolus , не потребляют пищу напрямую; [3] Вместо этого их питают симбиотические бактерии, которые также производят энергию из метана, подобно своим родственникам, образующим маты. [3] Хемосинтезирующие двустворчатые моллюски являются важными представителями фауны холодных просачиваний и представлены в этих условиях пятью семействами: Solemyidae , Lucinidae , Vesicomyidae , Thyasiridae и Mytilidae . [5]
Эта микробная деятельность производит карбонат кальция , который откладывается на морском дне и образует слой породы. [3] В течение периода, продолжающегося до нескольких десятилетий, эти скальные образования привлекают сибоглинид трубчатых червей , которые селятся и растут вместе с мидиями. [3] Как и мидии, трубчатые черви для выживания полагаются на хемосинтезирующие бактерии (в данном случае на те, которым нужен сероводород вместо метана). [3] Верный любым симбиотическим отношениям, трубчатый червь также обеспечивает свои бактерии, присваивая сероводород из окружающей среды. [3] Сульфид не только поступает из воды, но и добывается из осадка через обширную «корневую» систему, которую трубчатый червь устанавливает в твердом карбонатном субстрате. [3] Куст трубчатого червя может содержать сотни отдельных червей, которые могут расти на высоте метра и более над осадком. [3]
Холодные просачивания не могут длиться бесконечно. Поскольку скорость утечки газа медленно снижается, короткоживущие, голодные по метану мидии (или, точнее, их голодные по метану бактериальные симбионты) начинают вымирать. [3] На этом этапе трубчатые черви становятся доминирующим организмом в сообществе сипа. [3] Пока в отложениях присутствует некоторое количество сульфида, трубчатые черви, добывающие сульфиды, могут сохраняться. [3] По оценкам, особи одного вида трубчатых червей Lamellibrachia luymesi живут в таких условиях более 250 лет. [3]
 |  |  |
Бентосный фильтр
[ редактировать ]Организмы, обитающие на холодных выходах, оказывают большое влияние на углеродный цикл и климат. Хемосинтезирующие организмы, в частности метаногенные (потребляющие метан) организмы, препятствуют выбросу метана, просачивающегося из-под морского дна, в воду наверху. Поскольку метан является мощным парниковым газом, выбросы метана могут вызвать глобальное потепление, когда резервуары газогидратов дестабилизируются. [7] Потребление метана аэробными и анаэробными организмами морского дна называется «бентическим фильтром». [8] Первая часть этого фильтра — анаэробные бактерии и археи под морским дном, которые потребляют метан посредством анаэробного окисления метана (АОМ). [8] Если поток метана, протекающего через осадок, слишком велик и максимальное количество метана потребляют анаэробные бактерии и археи, то избыток метана потребляется свободно плавающими или симбиотическими аэробными бактериями над осадком на морском дне. Симбиотические бактерии были обнаружены у таких организмов, как трубчатые черви и моллюски, живущие в холодных источниках; эти организмы снабжают кислород аэробными бактериями, поскольку бактерии обеспечивают энергию, которую они получают от потребления метана. Понимание того, насколько эффективен бентосный фильтр, может помочь предсказать, сколько метана покидает морское дно при холодных просачиваниях и попадает в толщу воды и, в конечном итоге, в атмосферу. Исследования показали, что 50-90% метана потребляется на холодных выходах с бактериальными матами. В районах с залежами моллюсков выделяется менее 15% метана. [7] Эффективность определяется рядом факторов. Бентический слой более эффективен при низком потоке метана, а эффективность снижается по мере увеличения потока метана или скорости потока. [8] Потребность в кислороде у экосистем холодного просачивания намного выше, чем у других донных экосистем, поэтому, если в придонных водах недостаточно кислорода, то эффективность аэробных микробов по удалению метана снижается. [7] Бентосный фильтр не может воздействовать на метан, который не проходит через осадок. Метан может обойти донный фильтр, если он всплывает на поверхность или проходит через трещины и трещины в отложениях. [7] Эти организмы являются единственным биологическим поглотителем метана в океане. [8]
Сравнение с другими сообществами
[ редактировать ]
Холодные просачивания и гидротермальные жерла глубоких океанов — это сообщества, которые не полагаются на фотосинтез для производства продуктов питания и энергии. [2] Эти системы в значительной степени управляются хемосинтетической энергией. [2] Обе системы имеют общие характеристики, такие как наличие восстановленных химических соединений ( H 2 S и гидрокарбонатов ), локальная гипоксия или даже аноксия , высокая численность и метаболическая активность бактериальных популяций, а также производство автохтонного органического материала хемоавтотрофными бактериями. [2] Как гидротермальные источники, так и холодные просачивания демонстрируют сильно повышенный уровень биомассы многоклеточных животных в сочетании с низким местным разнообразием. [2] Это объясняется наличием плотных скоплений основных видов и эпизоотических животных, обитающих внутри этих скоплений. [2] Сравнения на уровне сообществ показывают, что макрофауна жерловых источников, просачиваний и органических осадков очень различна с точки зрения состава на уровне семейств, хотя они имеют много общих доминирующих таксонов среди высокосульфидных местообитаний. [4]
Однако гидротермальные источники и холодные просачивания также во многом различаются. По сравнению с более стабильными холодными выходами, жерла характеризуются локально высокими температурами, сильными колебаниями температуры, pH, концентраций сульфидов и кислорода, часто отсутствием отложений, относительно молодым возрастом и часто непредсказуемыми условиями, такими как рост и убыль. жерловых жидкостей или извержений вулканов. [2] В отличие от гидротермальных источников, которые представляют собой изменчивую и эфемерную среду, холодные просачивания излучают воду с медленной и стабильной скоростью. Вероятно, из-за более низких температур и стабильности многие организмы холодного просачивания живут гораздо дольше, чем те, что обитают в гидротермальных жерлах.
Конец сообщества холодного просачивания
[ редактировать ]путь для Наконец, когда холодные выходы становятся неактивными, трубчатые черви также начинают исчезать, освобождая кораллам поселения на теперь обнаженном карбонатном субстрате. [3] Кораллы не зависят от углеводородов, просачивающихся с морского дна. [3] Исследования Lophelia pertusa показывают, что они получают питание в основном с поверхности океана. [3] Хемосинтез играет лишь очень небольшую роль в их заселении и росте. [3] Хотя глубоководные кораллы, похоже, не являются организмами, основанными на хемосинтезе, хемосинтетические организмы, которые предшествовали им, обеспечивают существование кораллов. [3] Эта гипотеза о возникновении глубоководных коралловых рифов называется гидравлической теорией. [9] [10]
Распределение
[ редактировать ]Холодные выходы были обнаружены в 1983 году Чарльзом Поллом и его коллегами на Флоридском откосе в Мексиканском заливе на глубине 3200 метров (10 500 футов). [11] С тех пор просачивания были обнаружены во многих других частях мирового океана. Большинство из них были сгруппированы в пять биогеографических провинций: Мексиканский залив, Атлантический океан, Средиземноморье, Восточная часть Тихого океана и Западная часть Тихого океана. [12] но известны и холодные просачивания из-под шельфового ледника в Антарктиде , [13] Северный Ледовитый океан , Северное море , Скагеррак , Каттегат , Калифорнийский залив , Красное море , Индийский океан , у берегов южной Австралии и во внутренней части Каспийского моря . [14] На северо-западе Тихого океана холодный выход под названием «Оазис Пифии» . в 2015 году был обнаружен [15] С недавним открытием метанового просачивания в Южном океане , [16] Холодные просачивания теперь известны во всех крупных океанах. [4] Холодные просачивания распространены вдоль окраин континентов в районах высокой первичной продуктивности и тектонической активности, где деформация и уплотнение земной коры приводят к выбросам богатой метаном жидкости. [4] Холодные просачивания распределены неоднородно и чаще всего встречаются вблизи окраин океана, от литоральных до хадальных глубин. [4] В Чили холодные просачивания известны из приливной зоны. [17] в Каттегате выходы метана известны как «пузырящиеся рифы» и обычно находятся на глубине 0–30 м (0–100 футов). [18] а у северной Калифорнии их можно найти на глубине 35–55 м (115–180 футов). [14] Большинство холодных выходов расположены значительно глубже, далеко за пределами досягаемости обычного подводного плавания , а самое глубокое известное сообщество выходов находится в Японском желобе на глубине 7326 м (24035 футов). [19]
Помимо существующих сегодня холодных просачиваний, в нескольких частях мира были обнаружены ископаемые остатки древних систем просачиваний. Некоторые из них расположены далеко в глубине страны, в местах, ранее покрытых доисторическими океанами . [14] [20]
В Мексиканском заливе
[ редактировать ]
Открытия
[ редактировать ]Хемосинтезирующие сообщества Мексиканского залива интенсивно изучаются с 1990-х годов, и сообщества, впервые обнаруженные на верхнем склоне, вероятно, являются наиболее изученными сообществами просачивания в мире. История открытия этих замечательных животных вообще началась с 1980-х годов. Каждое крупное открытие было неожиданным — от первых гидротермальных жерл в любой точке мира до первых сообществ холодных просачиваний в Мексиканском заливе . [21]
Сообщества были обнаружены в восточной части Мексиканского залива в 1983 году с помощью подводного аппарата с экипажем DSV Alvin , во время круиза по исследованию дна Флоридского откоса в районах «холодного» просачивания рассола, где неожиданно были обнаружены трубчатые черви и мидии (Paull et al., 1984). [21] Две группы случайно обнаружили хемосинтетические сообщества в центральной части Мексиканского залива одновременно в ноябре 1984 года. Во время исследований Техасского университета A&M по определению воздействия просачивания нефти на экологию бентоса (до этого исследования считалось, что все последствия просачивания нефти пагубны), донные тралы неожиданно обнаружили обширные коллекции хемосинтезирующих организмов, включая трубчатых червей и моллюсков (Kennicutt et al., 1985). В то же время компания LGL Ecoological Research Associates проводила исследовательский рейс в рамках многолетнего исследования континентальных склонов северной части Мексиканского залива (Gallaway et al., 1988). Фотография дна (обработанная на борту судна) привела к получению четких изображений хемосинтетических сообществ везикомидных моллюсков по совпадению, так же, как и первое открытие с помощью фотосаней в Тихом океане в 1977 году. Фотографии во время того же круиза LGL / MMS также зафиксировали сообщества трубчатых червей. in situ в центральной части Мексиканского залива впервые (обработано только после круиза; Boland, 1986) до первоначальных исследований с помощью подводных лодок и описаний из первых рук. Буш Хилл (англ.
27 ° 47'02 "с.ш. 91 ° 30'31" з.д. / 27,78389 ° с.ш. 91,50861 ° з.д. ) в 1986 году (Росман и др., 1987a; Макдональд и др., 1989b). Объект был выбран из-за зон акустического «затухания» или отсутствия структуры субстрата, вызванного просачиванием углеводородов. Это было определено с помощью акустической системы пингера во время того же круиза на НИС « Эдвин Линк » (старый, всего 113 футов (34 м)), на котором использовался один из подводных аппаратов Johnson Sea Link . Для этого участка характерны плотные скопления трубчатых червей и мидий, а также обнаженные карбонатные обнажения с многочисленными горгонарий и лофелий колониями кораллов . Буш-Хилл стал одним из наиболее тщательно изученных мест хемосинтеза в мире. [21] WikiMiniAtlas
WikiMiniAtlasРаспределение
[ редактировать ]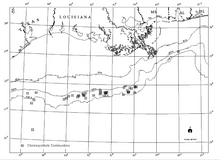
Существует четкая связь между известными месторождениями углеводородов на большой глубине на склоне залива и хемосинтетическими сообществами, просачиванием углеводородов и аутигенными минералами, включая карбонаты на морском дне (Sassen et al., 1993a и b). В то время как резервуары углеводородов представляют собой обширные территории в нескольких километрах под заливом, хемосинтетические сообщества встречаются в изолированных районах с тонкими слоями отложений толщиной всего несколько метров. [21]
Северный склон Мексиканского залива включает стратиграфический разрез толщиной более 10 км (6 миль) и подвергся сильному влиянию движения соли . Мезозойские нефтематеринские породы от верхней юры до верхнего мела генерируют нефть на большинстве месторождений на склонах Персидского залива (Sassen et al., 1993a и b). Миграционные каналы доставляют свежие углеводородные материалы по вертикали на расстояние 6–8 км (4–5 миль) к поверхности. Поверхностные проявления миграции углеводородов называются просачиваниями. Геологические данные показывают, что просачивание углеводородов и рассола сохраняется в пространственно дискретных областях в течение тысяч лет. [21]
Временной масштаб миграции нефти и газа из систем-источников составляет миллионы лет (Сассен, 1997). Просачивание из источников углеводородов через разломы к поверхности, как правило, распространяется через вышележащие отложения, обнажения карбонатов и отложения гидратов , поэтому соответствующие сообщества выходов углеводородов, как правило, крупнее (шириной несколько сотен метров), чем хемосинтетические сообщества, обнаруженные вокруг гидротермальных источников. восточной части Тихого океана (Макдональд, 1992). [21] Существуют большие различия в концентрациях углеводородов на участках просачивания. Робертс (2001) представил спектр реакций, которые следует ожидать в различных условиях скорости потока, варьирующихся от очень медленной просачивания до быстрой вентиляции. [21] [22] Участки с очень медленной просачиванием не поддерживают сложные хемосинтетические сообщества; скорее, они обычно поддерживают только простые микробные маты ( Beggiatoa sp.). [21]
В верхней части склона твердые субстраты, образовавшиеся в результате осаждения карбонатов, могут содержать сообщества нехемосинтезирующих животных, включая множество сидячих книдарий , таких как кораллы и морские анемоны . На конце спектра с быстрым потоком псевдоожиженный осадок обычно сопровождает углеводороды и пластовые флюиды, достигающие морского дна. В результате возникают грязевые вулканы и грязевые потоки . Где-то между этими двумя конечными членами существуют условия, которые поддерживают густонаселенные и разнообразные сообщества хемосинтезирующих организмов (микробные маты, сибоглинидные трубчатые черви, батимодиолиновые мидии, люцинидные и везикомидные моллюски и ассоциированные организмы). Эти области часто связаны с поверхностными или приповерхностными залежами газогидратов . У них также есть локализованные участки литифицированного морского дна, обычно аутигенные карбонаты, но иногда и более экзотические минералы, такие как барит . присутствуют [21]
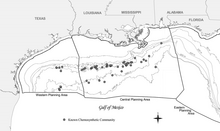
Широко распространенный характер хемосинтетических сообществ Мексиканского залива был впервые задокументирован в ходе контрактных исследований, проведенных Группой геологических и экологических исследований (GERG) Техасского университета A&M для Комитета морских операторов (Brooks et al., 1986). [21] Это исследование остается наиболее распространенным и всеобъемлющим, хотя с тех пор было зарегистрировано множество дополнительных сообществ. [21] Промышленная разведка энергетических запасов в Мексиканском заливе также выявила многочисленные новые сообщества на самых разных глубинах, включая самое глубокое из известных месторождений в центральной части Мексиканского залива в блоке 818 каньона Аламинос на глубине 2750 м (9022 фута). . [21] Появление хемосинтезирующих организмов, зависящих от просачивания углеводородов, было зарегистрировано на глубине от 290 м (951 фут) (Roberts et al., 1990) до 2744 м (9003 фута). [21] Этот диапазон глубин конкретно помещает хемосинтетические сообщества в глубоководный район Мексиканского залива, который определяется как глубина воды более 305 м (1000 футов). [21]
Хемосинтетические сообщества не встречаются на континентальном шельфе , хотя в летописи окаменелостей они появляются на глубине менее 200 м (656 футов). [21] Одна из теорий, объясняющая это, заключается в том, что давление хищников существенно менялось в течение рассматриваемого периода времени (Каллендер и Пауэлл, 1999). [21] [23] В настоящее время известно, что более 50 сообществ существуют в 43 блоках Внешнего континентального шельфа (OCS). [21] Хотя систематическое исследование для выявления всех хемосинтетических сообществ в Мексиканском заливе не проводилось, есть данные, указывающие на то, что может существовать гораздо больше таких сообществ. [21] Пределы глубины открытий, вероятно, отражают пределы разведки (отсутствие подводных аппаратов, способных погружаться на глубину более 1000 м (3281 фут)). [21]
Макдональд и др. (1993 и 1996) проанализировали изображения дистанционного зондирования из космоса, которые показывают наличие нефтяных пятен в северо-центральной части Мексиканского залива. [21] [24] [25] Результаты подтвердили обширное естественное просачивание нефти в Мексиканском заливе, особенно на глубине более 1000 м (3281 фут). [21] Всего было зарегистрировано 58 дополнительных потенциальных мест, где источники на морском дне могли образовывать постоянные нефтяные пятна (MacDonald et al., 1996). [21] Расчетные скорости фильтрации варьировались от 4 баррелей в сутки (0,64 м3). 3 /сут) до 70 баррелей в сутки (11 м 3 /сут) по сравнению с менее чем 0,1 барр./сутки (0,016 м 3 /d) для судовых сбросов (оба нормированы на расстояние 1000 миль). 2 (640 000 акр)). [21] Эти данные значительно увеличивают площадь, на которой можно ожидать появления хемосинтетических сообществ, зависящих от просачивания углеводородов. [21]
Самые плотные скопления хемосинтезирующих организмов были обнаружены на глубине около 500 м (1640 футов) и глубже. [21] Самое известное из этих сообществ было названо исследователями, которые впервые его описали, Буш-Хиллом (MacDonald et al., 1989b). [21] Это удивительно большое и плотное сообщество хемосинтетических трубчатых червей и мидий на месте естественной утечки нефти и газа над соляным диапиром в блоке 185 Грин-Каньон. Место просачивания представляет собой небольшой холм, возвышающийся примерно на 40 м (131 фут) над уровнем моря. окружающее морское дно на глубине около 580 м (1903 фута). [21]
Стабильность
[ редактировать ]По мнению Сассена (1997), роль гидратов в хемосинтетических сообществах сильно недооценена. [21] Биологическое изменение замороженных газогидратов было впервые обнаружено в ходе исследования MMS под названием «Стабильность и изменения в хемосинтетических сообществах Мексиканского залива». [26] Предполагается (MacDonald, 1998b), что динамика гидратных изменений может играть важную роль в качестве механизма регуляции выделения углеводородных газов, подпитывающих биогеохимические процессы, а также может играть существенную роль в стабильности сообщества. Зарегистрированные колебания температуры придонной воды на несколько градусов в некоторых районах, таких как участок Буш-Хилл (4–5 °C на глубине 500 м (1640 футов)), как полагают, приводят к диссоциации гидратов, что приводит к увеличению концентрации газа. потоки (MacDonald et al., 1994). не так разрушительна, как вулканизм в местах жерл срединно-океанических хребтов Хотя динамика образования и движения мелких гидратов , она явно повлияет на сидячих животных , которые составляют часть барьера просачивания. Существует вероятность катастрофического события, когда целый слой мелководных гидратов может оторваться от дна и существенно повлиять на местные сообщества хемосинтетической фауны. [21] На больших глубинах (>1000 м, >3281 фут) температура придонной воды холоднее (приблизительно на 3 °C) и испытывает меньшие колебания. Образование более устойчивых и, возможно, более глубоких гидратов влияет на приток легких углеводородных газов к поверхности осадков, что влияет на морфологию поверхности и характеристики хемосинтетических сообществ. В сложных сообществах, таких как Буш-Хилл, нефть кажется менее важной, чем считалось ранее (MacDonald, 1998b). [21]
Благодаря тафономическим исследованиям (комплексы отмерших раковин) и интерпретации состава высачивающихся комплексов из кернов Пауэлл и др. (1998) сообщили, что в целом сообщества сипов сохранялись в течение периодов 500–1000 лет и, вероятно, на протяжении всего плейстоцена. [ сомнительно – обсудить ] . Некоторые участки сохранили оптимальную среду обитания в течение геологических временных масштабов . Пауэлл сообщил о свидетельствах существования сообществ мидий и моллюсков на одних и тех же участках в течение 500–4000 лет. Пауэлл также обнаружил, что как видовой состав, так и трофическое распределение сообществ выходов углеводородов, как правило, довольно постоянны во времени, с временными изменениями только в количественной численности. Он обнаружил несколько случаев, когда тип сообщества менялся (например, от сообщества мидий к сообществу моллюсков) или полностью исчезал. Фаунистическая преемственность не наблюдалась. Удивительно, но когда произошло восстановление после прошлого разрушительного события, те же самые хемосинтетические виды вновь заняли это место. Свидетельств катастрофических захоронений было мало, но два случая были обнаружены в сообществах мидий в блоке 234 Грин-Каньон. Наиболее примечательным наблюдением, о котором сообщил Пауэлл (1995), была уникальность каждого участка хемосинтетического сообщества. [21]
Осадки аутигенных карбонатов и другие геологические события, несомненно, изменят структуру поверхностного просачивания в течение многих лет, хотя посредством прямых наблюдений не наблюдалось никаких изменений в распределении или составе хемосинтетической фауны на семи отдельных участках исследования (MacDonald et al., 1995). Несколько более длительный период (19 лет) можно отнести к Буш-Хиллу, первому сообществу в Центральной Мексиканском заливе, описанному in situ в 1986 году. Никаких массовых вымираний или крупномасштабных сдвигов в составе фауны не наблюдалось (с учетом за исключением коллекций для научных целей) за 19-летнюю историю исследований на этом объекте. [21]
Все хемосинтетические сообщества расположены в глубинах воды за пределами действия сильных штормов, включая ураганы, и не было бы никаких изменений этих сообществ, вызванных поверхностными штормами, включая ураганы . [21]
Биология
[ редактировать ]
Макдональд и др. (1990) описал четыре основных типа сообществ. Это сообщества, в которых доминируют вестиментиферовые трубчатые черви ( Lamellibrachia cf barhami и Escarpia spp.), митилиды (Seep Mytilid Ia, Ib, III и др.), пузырчатые моллюски ( Vesicomya cordata и Calyptogena ponderosa ), а также инфаунальные люциниды или тиазириды моллюски . ( Lucinoma sp. или Thyasira sp.). Бактериальные маты присутствуют на всех объектах, посещенных к настоящему времени. Эти группы фауны имеют тенденцию проявлять отличительные характеристики с точки зрения того, как они агрегируют, размера скоплений, геологических и химических свойств местообитаний, в которых они встречаются, и, в некоторой степени, гетеротрофной фауны, которая встречается с ними. Многие из видов, обитающих в этих сообществах холодных просачиваний в Мексиканском заливе, являются новыми для науки и остаются неописанными . [21]
Отдельные ламеллибрахидные трубчатые черви , более длинный из двух таксонов, встречающихся в просачиваниях, могут достигать длины 3 м (10 футов) и жить сотни лет (Fisher et al., 1997; Bergquist et al., 2000). Скорость роста , определенная для извлеченных маркированных трубчатых червей, была различной: от отсутствия роста у 13 особей, измеренных в течение одного года, до максимального роста 9,6 см/год (3,8 дюйма/год) у особи Lamellibrachia (MacDonald, 2002). Средняя скорость роста составила 2,19 см/год (0,86 дюйма/год) для Escarpia -подобных видов и 2,92 см/год (1,15 дюйма/год) для ламеллибрахид. Это более медленные темпы роста, чем у их родственников из гидротермальных жерл , но особи Lamellibrachia могут достигать длины в 2–3 раза больше, чем у крупнейших известных видов гидротермальных жерл. [21] Особи Lamellibrachia sp. Несколько раз собирались экземпляры высотой более 3 м (10 футов), что соответствует возрасту, вероятно, превышающему 400 лет (Fisher, 1995). Нерест вестиментиферовых трубчатых червей не носит сезонного характера, а пополнение носит эпизодический характер. [21]
Трубчатые черви бывают мужскими и женскими. Одно недавнее открытие указывает на то, что нерест самок Lamellibrachia , по-видимому, привел к уникальной ассоциации с большим двустворчатым моллюском Acesta Bullisi , который живет постоянно прикрепленным к переднему отверстию трубки трубчатого червя и питается периодическим выпуском яиц (Järnegren et al., 2005). Эта тесная связь между двустворчатыми моллюсками и трубчатыми червями была обнаружена в 1984 г. (Boland, 1986), но не объяснена полностью. Практически все половозрелые особи Acesta встречаются у самок, а не у самцов трубчатых червей. Это свидетельство и другие эксперименты Ярнегрена и др. (2005), похоже, разгадали эту загадку. [21]
Сообщалось о темпах роста метанотрофных мидий в местах холодного просачивания (Fisher, 1995). [21] Общие темпы роста оказались относительно высокими. Скорость роста взрослых мидий была такой же, как и у мидий из прибрежной среды при аналогичных температурах. Фишер также обнаружил, что молодые мидии в местах выхода углеводородов сначала растут быстро, но у взрослых скорость роста заметно падает; они очень быстро вырастают до репродуктивных размеров. Как отдельные люди, так и сообщества кажутся очень долгоживущими. Эти метанзависимые мидии имеют строгие химические требования, которые привязывают их к районам наиболее активной утечки в Мексиканском заливе. В результате их быстрых темпов роста реколонизация мидий нарушенного места просачивания может происходить относительно быстро. Есть некоторые свидетельства того, что мидиям также требуется твердый субстрат, и их численность может увеличиться, если на морском дне увеличится количество подходящего субстрата (Fisher, 1995). Два ассоциированных вида всегда обнаруживаются в зарослях мидий — брюхоногие моллюски Bathynerita naticoidea и небольшая креветка Alvinocarid — что позволяет предположить, что эти эндемичные виды обладают превосходными способности к расселению и могут переносить широкий спектр условий (MacDonald, 2002). [21]
В отличие от зарослей мидий, заросли хемосинтезирующих моллюсков могут сохраняться как видимый поверхностный феномен в течение длительного периода без появления новых живых особей из-за низких скоростей растворения и низких скоростей седиментации. Большинство пластов моллюсков, исследованных Пауэллом (1995), были неактивными. Живые особи встречались редко. Пауэлл сообщил, что в течение 50 лет локальное вымирание и повторная колонизация должны быть постепенными и крайне редкими. В отличие от этих неактивных пластов, первое сообщество, обнаруженное в центральной части Мексиканского залива, состояло из многочисленных активно пашущих моллюсков. Изображения, полученные этого сообщества, были использованы для определения соотношения длины/частоты и соотношения живых/мертвых особей, а также пространственных закономерностей (Rosman et al., 1987a). [21]
обширные бактериальные маты из свободноживущих бактерий На всех участках просачивания углеводородов также наблюдаются . Эти бактерии могут конкурировать с основной фауной за источники энергии из сульфидов и метана , а также могут вносить существенный вклад в общее производство (MacDonald, 1998b). Было обнаружено, что белые непигментированные маты представляют собой автотрофные серные бактерии вида Beggiatoa , а оранжевые маты обладают неустановленным нехемосинтетическим метаболизмом (MacDonald, 1998b). [21]
Гетеротрофные виды на участках высачивания представляют собой смесь видов, уникальных для высачиваний (особенно моллюсков и ракообразных беспозвоночных), и тех, которые являются нормальным компонентом окружающей среды. Карни (1993) впервые сообщил о потенциальном дисбалансе, который может возникнуть в результате хронических нарушений. Из-за спорадических моделей пополнения хищники могут получить преимущество, что приведет к истреблению местных популяций зарослей мидий. Очевидно, что системы просачивания действительно взаимодействуют с фоновой фауной, но остаются противоречивые данные относительно того, в какой степени происходит прямое хищничество некоторых конкретных компонентов сообщества, таких как трубчатые черви (MacDonald, 2002). Более удивительные результаты этой недавней работы заключаются в том, почему фоновые виды не используют продукцию сепарации в большей степени, чем кажется очевидным. Фактически, у потребителей, связанных с просачиванием, таких как галатеидные крабы и неритовые брюхоногие моллюски, были изотопные сигнатуры, указывающие на то, что их рацион представлял собой смесь просачивающегося и фонового производства. На некоторых сайтах эндемичные беспозвоночные, которые, как ожидалось, должны были получать большую часть своего рациона, если не весь свой рацион, за счет продукции просачивания, на самом деле потребляли до 50 процентов своего рациона из фонового источника. [21]
В Атлантическом океане
[ редактировать ]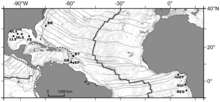
BR – Блейк Ридж подгузник
BT - Барбадосская траншея
ИЛИ – сектора Ореноке
EP - сектор Эль Пилар
NIG - Нигерийский склон
Графический интерфейс – область Guiness
РЭГ – Регабская оспина.
Сообщества холодного просачивания в западной части Атлантического океана также были описаны при нескольких погружениях на грязевые вулканы и диапиры на глубине от 1000 до 5000 м (3300–16 400 футов) в Барбадоса районе аккреционной призмы и в Блейк-Ридж диапире у побережья Северной Каролины . Совсем недавно сообщества сипов были обнаружены в восточной Атлантике, на гигантском скоплении покмарков в Гвинейском заливе возле глубоководного канала Конго , а также на других покмарках на окраине Конго, окраине Габона и окраине Нигерии , а также в Кадисском заливе. . [12]
Впервые о появлении хемосимбиотической биоты на обширных полях грязевых вулканов Кадисского залива было сообщено в 2003 году. [27] Хемосимбиотические двустворчатые моллюски, собранные в грязевых вулканах Кадисского залива, были рассмотрены в 2011 году. [5]
Холодные просачивания известны также из северной части Атлантического океана. [2] даже доходит до Северного Ледовитого океана, у берегов Канады и Норвегии . [14]
Обширные отборы проб фауны были проведены на высоте 400 и 3300 м (1300–10 800 футов) в Атлантическом экваториальном поясе от Мексиканского залива до Гвинейского залива, включая аккреционную призму Барбадоса, диапир Блейк-Ридж, а также в Восточной Атлантике от Окраины Конго и Габона и недавно исследованные окраины Нигерии в ходе Census of Marine Life ChEss проекта . Из 72 таксонов, выявленных на видовом уровне, всего 9 видов или видовых комплексов идентифицированы как амфиатлантические. [12]
На структуру сообщества мегафауны просачивающегося экваториального пояса влияет в первую очередь глубина, а не географическое расстояние. двустворчатых моллюсков Bathymodiolinae (в пределах Mytilidae Наиболее распространены в Атлантике виды или комплексы видов ). Комплекс бумерангов Bathymodiolus обнаружен на откосе Флориды, в диапире Блейк-Ридж, на Барбадосской призме и на участке Регаб в Конго. Комплекс Bathymodiolus childressi также широко распространен вдоль Атлантического экваториального пояса от Мексиканского залива до окраины Нигерии, хотя и не на участках Регаб или Блейк-Ридж. Комменсальный полиноид Branchipolynoeseepensis известен из Мексиканского, Гвинейского заливов и Барбадоса. Другими видами, распространение которых простирается от восточной до западной Атлантики, являются: брюхоногие моллюски Cordesia provannoides , креветка Alvinocaris muricola , галатеиды Munidopsis geyeri и Munidopsis livida и , вероятно, голотурида Chiridota heheva . [12]
Холодные просачивания были обнаружены и в конусе глубин Амазонки . Сейсмические профили высокого разрешения вблизи края шельфа показывают признаки приповерхностных обвалов и разломов на глубине 20–50 м (66–164 футов) в недрах, а также концентрации (около 500 м) 2 или 5400 кв. футов) метана. Некоторые исследования (например, Исследование шельфа Амазонки — AMASEDS , LEPLAC , REMAC , GLORIA, Ocean Drilling Program ) показывают, что существуют доказательства просачивания газа на склоне веера Амазонки, основанные на распространении отражений от дна (BSR), грязи вулканы, оспины, газ в отложениях и более глубокие залежи углеводородов. В этом регионе картировано наличие метана на относительно небольших глубинах и обширные площади газогидратов. Также сообщалось о газовых трубах , а разведочные скважины обнаружили субкоммерческие скопления газа и выбоины вдоль плоскостей разломов. Точные геологические и геофизические данные о бассейне Фос-ду-Амазонас уже доступны и используются энергетическими компаниями. [28]
Необходимо изучить новые области, такие как потенциальные места просачивания у восточного побережья США и Лаврентийский конус, где хемосинтетические сообщества известны на глубине более 3500 м (11 500 футов), а также более мелкие участки в Гвинейском заливе. будущее. [12] [ нужны разъяснения ]
В Средиземноморье
[ редактировать ]Первым биологическим свидетельством сокращения окружающей среды в Средиземном море стало присутствие Lucinidae и Vesicomyidae, двустворчатых раковин извлеченных из вершины грязевого вулкана Неаполя (
33 ° 43'52 "с.ш. 24 ° 40'52" в.д. / 33,73111 ° с.ш. 24,68111 ° в.д. ; «Неаполь» — это всего лишь название подводной горы. Расположен южнее Крита), расположен на глубине 1900 м на Средиземноморском хребте в зоне субдукции Африканской плиты . За этим последовало описание нового вида двустворчатых моллюсков Lucinidae, Lucinoma kazani , связанного с бактериальными эндосимбионтами . сообщества полихет В юго-восточном Средиземноморье на глубинах 500–800 м также обнаружены и двустворчатых моллюсков, ассоциированные с холодными выходами и карбонатами вблизи Египта и сектора Газа на глубинах 500–800 м, но живая фауна собрана не была. Первые наблюдения in situ обширных живых хемосинтетических сообществ в восточной части Средиземного моря стимулировали сотрудничество биологов, геохимиков и геологов. Во время подводных погружений в различных местообитаниях холодных просачиваний, связанных с карбонатными корками, на глубине 1700–2000 м наблюдались сообщества, состоящие из больших полей мелких двустворчатых моллюсков (мертвых и живых), крупных трубчатых червей сибоглинид , изолированных или образующих плотные скопления, крупных губок и связанной с ними эндемической фауны. глубина м. Впервые были исследованы два поля грязевых вулканов, одно вдоль Средиземноморского хребта, где большинство из них были частично (грязевые вулканы Неаполя, Милана) или полностью (грязевые вулканы Урания, Мейдстоун) затронуты рассолов , а другой — на курганах Анаксимандра к югу от Турции . Последняя область включает крупный грязевой вулкан Амстердам, пострадавший от недавних селей , и меньшие грязевые вулканы Казанский или Кулинский. На грязевых вулканах Амстердама и Казани были отобраны пробы газовых гидратов , а над морским дном зафиксированы высокие уровни метана. несколько провинций глубоководного конуса Нила В последнее время исследовано . К ним относятся очень активное просачивание рассола под названием Кальдера Менес в восточной провинции на высоте от 2500 до 3000 м, оспины в центральной части вдоль средних и нижних склонов, а также грязевые вулканы восточной провинции, а также один в центральной части. верхний склон ( район Северный Алекс ) на глубине 500 м. [29]Во время этих первых исследовательских погружений были отобраны и идентифицированы таксоны-симбионты, аналогичные тем, которые наблюдались на грязевых месторождениях Олимпии и Анаксимандр. Такое сходство неудивительно, поскольку большинство этих таксонов первоначально были описаны при дноуглубительных работах в веере Нила. [29] До пяти видов двустворчатых моллюсков, содержащих бактериальных симбионтов, колонизировали эту богатую метаном и сульфидами среду. Новый вид полихеты Siboglinidae , Lamellibrachia anaximandri , трубчатый червь, колонизирующий холодные выходы от Средиземноморского хребта до глубоководного конуса Нила, был описан только в 2010 году. [29] [30] Кроме того, изучение симбиозов выявило ассоциации с хемоавтотрофными бактериями, окислителями серы у двустворчатых моллюсков Vesicomyidae и Lucinidae и трубчатых червей Siboglinidae, а также выявило исключительное разнообразие бактерий, живущих в симбиозе с мелкими Mytilidae. Средиземноморские выходы, по-видимому, представляют собой богатую среду обитания, характеризующуюся богатством видов мегафауны (например, брюхоногие моллюски ) или исключительными размерами некоторых видов, таких как губки ( Rizaxinellapyrifera ) и крабы ( Chaceon mediterraneus ), по сравнению с их фоновыми аналогами. Это контрастирует с низкой численностью и разнообразием макро- и мегафауны глубокого восточного Средиземноморья . Сообщества сипов в Средиземноморье, которые включают эндемичные хемосинтетические виды и связанную с ними фауну, отличаются от других известных сообществ сипов в мире не только на видовом уровне, но и отсутствием крупных родов двустворчатых моллюсков Calyptogena или Bathymodiolus . Изоляция Средиземного моря просачивается из Атлантического океана после Мессинский кризис привел к развитию уникальных сообществ, которые, вероятно, отличаются по составу и структуре от сообществ Атлантического океана. Дальнейшие экспедиции включали количественный отбор проб местообитаний в разных районах, от Средиземноморского хребта до глубоководного конуса восточного Нила. [29] Холодные просачивания обнаружены в Мраморном море в 2008 году. [31] также выявили сообщества, основанные на хемосинтезе, которые показали значительное сходство с симбионтной фауной холодных выходов Восточного Средиземноморья. [29]
В Индийском океане
[ редактировать ]В Макранском желобе , зоне субдукции вдоль северо-восточной окраины Оманского залива, прилегающей к юго-западному побережью Пакистана и юго-восточному побережью Ирана , сжатие аккреционного клина привело к образованию холодных выходов и грязевых вулканов. [32]
В западной части Тихого океана
[ редактировать ]Самородный алюминий также был обнаружен в холодных просачиваниях на северо-восточном континентальном склоне Южно -Китайского моря и Chen et al. (2011) [33] предложили теорию его происхождения как результат восстановления тетрагидроксоалюмината Al( OH ) 4 − к металлическому алюминию бактериями . [33]
Япония
[ редактировать ]| Холодное просачивание | |
| Гидротермальное жерло | |
| Падение кита |
|
Глубоководные сообщества вокруг Японии в основном исследуются Японским агентством морских наук и технологий о Земле (JAMSTEC). DSV Shinkai 6500 , Kaikō и другие группы обнаружили множество стоянок.
Сообщества метановых сипов в Японии распределены вдоль областей конвергенции плит из-за сопутствующей тектонической активности. Многие выходы были обнаружены в Японском желобе , впадине Нанкай , желобе Рюкю , заливе Сагами , заливе Суруга и Японском море . [35]
Члены сообществ холодных сипов сходны с другими регионами по семейству или роду, например Polycheata, Lamellibrachia, Bivalavia, Solemyidae, Bathymodiolus у Mytilidae, Thyasiridae, Calyptogena у Vesicomyidae и т. д. [34] Многие виды холодных выходов Японии являются эндемиками . [35]
В заливе Кагосима есть выходы метана, называемые «тагири» (кипение). Ламеллибрахия Сацума Там обитает . Глубина этого участка составляет всего 80 м, и это самая мелкая точка, где, как известно, обитают сибоглиниды. L. satsuma можно длительное время держать в аквариуме при температуре 1 атм. В двух аквариумах Японии содержатся и экспонируются L. satsuma . Разрабатывается метод наблюдения для введения его в прозрачную виниловую трубку. [36]

DSV Shinkai 6500 обнаружил сообщества везикомидных моллюсков в Южно- Марианском предгорье . Они зависят от метана, который возникает в серпентините . Другие хемосинтетические сообщества зависели бы от органического вещества углеводородного происхождения в земной коре, но эти сообщества зависят от метана, происходящего из неорганических веществ мантии. [37] [38]
В 2011 году территория вокруг Японского желоба пострадала от землетрясения Тохоку . Есть трещины, утечки метана и бактериальные маты, которые, вероятно, образовались в результате землетрясения. [39] [40]
Новая Зеландия
[ редактировать ]У материкового побережья Новой Зеландии нестабильность края шельфа в некоторых местах усиливается из-за холодных выходов богатых метаном флюидов, которые также поддерживают хемосинтетическую фауну и карбонатные конкреции. [41] [42] [43] [44] Доминирующими животными являются трубчатые черви семейства Siboglinidae и двустворчатые моллюски семейств Vesicomyidae и Mytilidae ( Bathymodiolus ). Многие из его видов кажутся эндемичными. Глубоководное траление нанесло серьезный ущерб сообществам холодного просачивания, и эти экосистемы находятся под угрозой. Холодные просачивания обнаружены на глубине до 2000 м, а топографическая и химическая сложность среды обитания еще не нанесена на карту. [ когда? ] . Масштабы открытия новых видов в этих плохо изученных или неисследованных экосистемах, вероятно, будут высокими. [44] [41]
В восточной части Тихого океана
[ редактировать ]
В глубоководных водах проект COMARGE изучил структуру биоразнообразия вдоль и поперек чилийской окраины с помощью сложных экосистем, таких как выходы метана и зоны минимума кислорода , и сообщил, что такая гетерогенность среды обитания может влиять на структуру биоразнообразия местной фауны. [28] [45] [46] [47] Фауна сипа включает двустворчатых моллюсков семейств Lucinidae , Thyasiridae , Solemyidae ( Acharax sp.) и Vesicomyidae ( Calyptogena Gallardoi ), а также полихет ( Lamellibrachia sp. и два других вида полихет). [46] Кроме того, в этих мягких восстановленных отложениях ниже зоны минимума кислорода у окраины Чили существует разнообразное микробное сообщество, состоящее из множества крупных прокариот (в основном крупных многоклеточных нитчатых «мегабактерий» родов Thioploca и Beggiatoa , а также «макробактерий». (включая разнообразие фенотипов), простейших (инфузории, жгутиконосцы и фораминиферы), а также мелких многоклеточных животных (главным образом нематод и полихет). [28] [48] Галлардо и др. (2007) [48] утверждают, что вероятный хемолитотрофный метаболизм большинства этих мега- и макробактерий предлагает альтернативное объяснение находкам окаменелостей , в частности тем, которые имеют очевидное нелиторальное происхождение, предполагая, что традиционные гипотезы о цианобактериальном происхождении некоторых окаменелостей, возможно, придется пересмотреть. [28]
Холодные просачивания ( оспины ) известны также с глубин 130 м в проливе Геката , Британская Колумбия , Канада. [49] Здесь обнаружена неочевидная фауна (неочевидная также для холодных высачиваний) с такими доминирующими видами: морская улитка Fusitriton oregonensis , анемона Metridium giganteum , корковые губки и двустворчатые моллюски Solemya reidi . [49]
Холодные просачивания с хемосинтетическими сообществами вдоль тихоокеанского побережья США встречаются в каньоне Монтерей , недалеко от залива Монтерей , штат Калифорния, на грязевом вулкане . [50] Встречаются, например, Calyptogena моллюски Calyptogena kimleri и Calyptogena pacifica. [51] и фораминифера Spiroplectammina biformis . [52]
Кроме того, просачивания были обнаружены на шельфе южной Калифорнии во внутренних приграничных районах Калифорнии вдоль нескольких систем разломов, включая разлом Сан-Клементе, [53] Ошибка Сан-Педро, [54] и разлом Сан-Диего . [55] Поток жидкости в выходах вдоль разломов Сан-Педро и Сан-Диего, по-видимому, контролируется локализованными сдерживающими изгибами в разломах. [55]
В Антарктике
[ редактировать ]Первое холодное просачивание из Южного океана было зарегистрировано в 2005 году. [16] Относительно немногие исследования глубоководных вод Антарктики показали наличие глубоководных сред обитания, включая гидротермальные жерла , холодные просачивания и грязевые вулканы . [56] За исключением круизов в рамках Проекта по сохранению глубоководного биоразнообразия антарктического бентоса (ANDEEP), в глубоководных районах мало что было сделано. [56] Есть и другие виды, ожидающие описания. [56]
Обнаружение
[ редактировать ]Благодаря продолжающемуся опыту, особенно на верхнем континентальном склоне Мексиканского залива, успешное предсказание присутствия сообществ трубчатых червей продолжает улучшаться; однако хемосинтетические сообщества не могут быть надежно обнаружены непосредственно с помощью геофизических методов . Выходы углеводородов, которые позволяют существовать хемосинтетическим сообществам ( бассейн Гуаймас ), изменяют геологические характеристики способами, которые можно обнаружить удаленно, но временные рамки одновременного активного просачивания и присутствия живых сообществ всегда неопределенны. Эти известные модификации отложений включают (1) осаждение аутигенного карбоната в форме микроконкреций, конкреций или горных масс; (2) образование газовых гидратов ; (3) изменение состава осадков за счет концентрации остатков твердых хемосинтезирующих организмов (таких как фрагменты и слои раковин); (4) образование межузельных пузырьков газа или углеводородов; и (5) образование впадин или оспин в результате выброса газа. Эти особенности вызывают акустические эффекты, такие как зоны затухания (отсутствие эха), твердое дно (сильное отражающее эхо), яркие пятна (слои с усиленным отражением) или реверберирующие слои (Behrens, 1988; Roberts and Neurauter, 1990). Потенциальные местоположения большинства типов сообществ могут быть определены путем тщательной интерпретации этих различных геофизических модификаций, но на сегодняшний день этот процесс остается несовершенным, и подтверждение существования живых сообществ требует прямых визуальных методов. [21]
Окаменелые записи
[ редактировать ]
Отложения холодного просачивания встречаются на протяжении всей фанерозойской геологической летописи, особенно в позднемезозое и кайнозое . Яркие примеры можно найти в перми Тибета. [57] меловой период Хоккайдо , [58] Палеоген Хонсю , [59] неоген Северной Италии , [60] и плейстоцен Калифорнии . [61] Эти ископаемые холодные просачивания характеризуются холмообразным рельефом (там, где он сохранился), крупнокристаллическими карбонатами и обилием моллюсков и брахиопод .
Воздействие на окружающую среду
[ редактировать ]Основными угрозами, с которыми сегодня сталкиваются экосистемы холодного просачивания и их сообщества, являются мусор на морском дне, химические загрязнители и изменение климата. Мусор на морском дне изменяет среду обитания, предоставляя твердый субстрат там, где его раньше не было, или перекрывая отложения, тем самым подавляя газообмен и мешая организмам на дне моря. Исследования морского мусора в Средиземном море включают исследование мусора морского дна на континентальном шельфе, склоне и батиальной равнине. [62] [63] В большинстве исследований на долю пластиковых предметов приходилось большая часть мусора, иногда до 90% и более от общего количества, из-за их повсеместного использования и плохой способности к разложению.
Оружие и бомбы также выбрасываются в море, а их сброс в открытые воды способствует загрязнению морского дна. Еще одной серьезной угрозой для донной фауны является наличие утерянных рыболовных снастей, таких как сети и ярусы, которые способствуют призрачному лову рыбы и могут нанести ущерб хрупким экосистемам, таким как холодноводные кораллы.
химические загрязнители, такие как стойкие органические загрязнители , токсичные металлы (например, Hg, Cd, Pb, Ni), радиоактивные соединения, пестициды, гербициды и фармацевтические препараты. В глубоководных отложениях также накапливаются [64] Топография (например, каньоны) и гидрография (например, каскадные явления) играют важную роль в транспортировке и накоплении этих химических веществ от побережья и шельфа к глубоким бассейнам, влияя на местную фауну. Недавние исследования выявили наличие значительных уровней диоксинов в коммерческой креветке Aristeus антеннатус. [65] и значительные уровни стойких органических загрязнителей у мезопелагических и батипелагических головоногих моллюсков. [66]
Климатические процессы и изменение климата повлияют на частоту и интенсивность каскадных явлений с неизвестным воздействием на донную фауну. Еще один потенциальный эффект изменения климата связан с переносом энергии из поверхностных вод на морское дно. [67] Первичная продукция будет меняться в поверхностных слоях в зависимости от воздействия солнца, температуры воды, значительной стратификации водных масс и других эффектов, и это повлияет на пищевую цепь вплоть до глубокого морского дна, которая будет подвержена различиям в количестве, качестве, и время внесения органических веществ. Поскольку коммерческое рыболовство перемещается в более глубокие воды, все эти последствия будут влиять на сообщества и популяции организмов в холодных просачиваниях и глубоком море в целом.
См. также
[ редактировать ]Ссылки
[ редактировать ]Эта статья включает в себя общественным достоянием работу правительства США, являющуюся , на основе ссылок. [3] [21] и CC-BY-2.5 из ссылок [2] [4] [6] [12] [28] [29] [35] [41] [56] и текст CC-BY-3.0 из ссылки [5]
- ^ (2008). Фудзикура , Кацунори, Окутани , Тадаши Маруяма , Такаши , 978-4-486-01787-5 . п. 20.
- ↑ Перейти обратно: Перейти обратно: а б с д и ж г час я дж к л м н тот п д Ванрейзель, А.; Де Гроот, А.; Голлнер, С.; Брайт, М. (2010). «Экология и биогеография свободноживущих нематод, связанных с хемосинтетической средой глубокого моря: обзор» . ПЛОС ОДИН . 5 (8): e12449. Бибкод : 2010PLoSO...512449V . дои : 10.1371/journal.pone.0012449 . ПМК 2929199 . ПМИД 20805986 .
- ↑ Перейти обратно: Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С Хсинг П.-Ю. (19 октября 2010 г.). «Газовый круг жизни – преемственность в глубоководной экосистеме» . НОАА Исследователь океана | Лофелия II 2010: Нефтяные просачивания и глубокие рифы | 18 октября Журнал. Проверено 25 января 2011 г.
- ↑ Перейти обратно: Перейти обратно: а б с д и ж г час Бернардино, AF; Левин, Луизиана; Тербер, Арканзас; Смит, ЧР (2012). «Сравнительный состав, разнообразие и трофическая экология донной макрофауны жерл, сипов и органических водопадов» . ПЛОС ОДИН . 7 (4): е33515. Бибкод : 2012PLoSO...733515B . дои : 10.1371/journal.pone.0033515 . ПМЦ 3319539 . ПМИД 22496753 .
- ↑ Перейти обратно: Перейти обратно: а б с Оливер, Г.; Родригес, К; Кунья, MR (2011). «Хемосимбиотические двустворчатые моллюски из грязевых вулканов Кадисского залива, северо-восток Атлантики, с описаниями новых видов Solemyidae, Lucinidae и Vesicomyidae» . ZooKeys (113): 1–38. Бибкод : 2011ZooK..113....1O . дои : 10.3897/ZooKeys.113.1402 . ПМК 3187628 . ПМИД 21976991 .
- ↑ Перейти обратно: Перейти обратно: а б Боэций, А (2005). «Взаимодействие микрофауны и макрофауны на морском дне: уроки трубчатого червя» . ПЛОС Биология . 3 (3): е102. дои : 10.1371/journal.pbio.0030102 . ПМЦ 1065708 . ПМИД 15760275 .
- ↑ Перейти обратно: Перейти обратно: а б с д Соммер, С.; Пфанкуче, О.; Линке, П.; Лафф, Р.; Грейнерт, Дж.; Дрюс, М.; Губш, С.; Пипер, М.; Позер, М.; Виргутц, Т. (июнь 2006 г.). «Эффективность донного фильтра: Биологический контроль выбросов растворенного метана из отложений, содержащих неглубокие газогидраты на Гидратном хребте: БИОЛОГИЧЕСКИЙ КОНТРОЛЬ ВЫБРОСОВ РАСТВОРЕННОГО МЕТАНА» . Глобальные биогеохимические циклы . 20 (2): н/д. дои : 10.1029/2004GB002389 . HDL : 1956/1315 . S2CID 54695808 .
- ↑ Перейти обратно: Перейти обратно: а б с д Боэций, Антье; Венцхёфер, Франк (сентябрь 2013 г.). «Потребление кислорода на морском дне, вызванное метаном из холодных просачиваний» . Природа Геонауки . 6 (9): 725–734. Бибкод : 2013NatGe...6..725B . дои : 10.1038/ngeo1926 . ISSN 1752-0894 .
- ^ Ховланд, М.; Томсен, Э. (1997). «Холодноводные кораллы - связаны ли они с выходом углеводородов?». Морская геология . 137 (1–2): 159–164. Бибкод : 1997MGeol.137..159H . дои : 10.1016/S0025-3227(96)00086-2 .
- ^ Ховланд М. (2008). Глубоководные коралловые рифы: уникальные очаги биоразнообразия . 8.10. Краткое изложение и повторение гидравлической теории. Спрингер , 278 стр. ISBN 978-1-4020-8461-4 . Страницы 204-205 .
- ^ Полл; Хекер; Коммо; и др. (1984). «Биологические сообщества на откосе Флориды напоминают таксоны гидротермальных жерл». Наука . 226 (4677): 965–967. Бибкод : 1984Sci...226..965P . дои : 10.1126/science.226.4677.965 . ПМИД 17737352 . S2CID 45699993 .
- ↑ Перейти обратно: Перейти обратно: а б с д и ж Олу, К.; Кордес, Э.Э.; Фишер, ЧР; Брукс, Дж. М.; Сибуэт, М.; Дебрюйер, Д. (2010). «Биогеография и потенциальный обмен между фаунами холодного просачивания Атлантического экваториального пояса» . ПЛОС ОДИН . 5 (8): e11967. Бибкод : 2010PLoSO...511967O . дои : 10.1371/journal.pone.0011967 . ПМК 2916822 . ПМИД 20700528 .
- ^ «Гибель антарктического шельфового ледника открывает новую жизнь» . Национальный научный фонд . 2007 . Проверено 14 февраля 2008 г.
- ↑ Перейти обратно: Перейти обратно: а б с д Левин, Луизиана (2005). «Экология холодных осадков: взаимодействие фауны с потоком, химией и микробами». В Гибсоне, штат РН; Р.Дж.А. Аткинсон; JDM Гордон (ред.). Океанография и морская биология: Ежегодный обзор . Том. 43. стр. 1–46. ISBN 9780849335976 .
- ^ «Оазис Пифиас: подводный источник, непохожий ни на один другой» . ООИ Региональная кабельная решетка . Университет Вашингтона. 1 октября 2019 года . Проверено 24 апреля 2023 г.
- ↑ Перейти обратно: Перейти обратно: а б Домак, Э .; Ишман, С.; Левентер, А.; Сильва, С.; Уиллмотт, В. (2005). «Хемотрофная экосистема, обнаруженная под шельфовым ледником Антарктики» . Эос, Труды Американского геофизического союза . 86 (29): 269–276. Бибкод : 2005EOSTr..86..269D . дои : 10.1029/2005EO290001 . S2CID 35944740 .
- ^ Йессен; Пантоха; Гутьеррес; и др. (2011). «Метан в неглубоких холодных выходах на острове Мокко в центре Чили». Исследования континентального шельфа . 31 (6): 574–581. Бибкод : 2011CSR....31..574J . дои : 10.1016/j.csr.2010.12.012 . hdl : 10533/129437 .
- ^ «Красный список - Подводные конструкции, изготовленные из утечек газов» (PDF) . ХЕЛКОМ . 2013 . Проверено 16 июня 2017 г.
- ^ Фудзикура; Кодзима; Тамаки; и др. (1999). «Самое глубокое сообщество, основанное на хемосинтезе, когда-либо обнаруженное в зоне хадал на глубине 7326 м в Японском желобе» . Серия «Прогресс в области морской экологии» . 190 : 17–26. Бибкод : 1999MEPS..190...17F . дои : 10.3354/meps190017 .
- ^ Кэмпбелл, Калифорния; Джей Ди Фармер; Д. Де Марэ (2002). «Древние углеводороды просачиваются из мезозойской конвергентной окраины Калифорнии: карбонатная геохимия, флюиды и палеосреда» . Геофлюиды . 2 (2): 63–94. Бибкод : 2002Gflui...2...63C . дои : 10.1046/j.1468-8123.2002.00022.x .
- ↑ Перейти обратно: Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа аб и объявление но из в ах есть также и аль являюсь а к ап ак с «Продажи нефти и газа в аренду OCS в Мексиканском заливе: 2007–2012 гг. Продажи в западной зоне планирования 204, 207, 210, 215 и 218. Продажи в зоне центрального планирования 205, 206, 208, 213, 216 и 222. Проект воздействия на окружающую среду Заявление, том I: главы 1–8 и приложения» (PDF) . Служба управления минеральными ресурсами Мексиканского залива, регион OCS, Новый Орлеан . Министерство внутренних дел США. Ноябрь 2006 г., стр. 3–27, 3–31. Архивировано из оригинала (PDF) 26 марта 2009 года.
- ^ Робертс, Х.Х. (2001). «Выброс жидкости и газа на континентальном склоне северной части Мексиканского залива: реакция от грязи и минералов» . Гидраты природного газа . Серия геофизических монографий. Том. 124. Американский геофизический союз . стр. 145–161. Бибкод : 2001GMS...124..145R . дои : 10.1029/GM124p0145 . ISBN 9781118668412 . Архивировано из оригинала 28 октября 2012 года . Проверено 26 марта 2012 г.
- ^ Каллендер, В. Рассел; Э. Н. Пауэлл (1999). «Почему древние хемосинтетические просачивания и жерла возникали на более мелководных участках, чем сегодня?». Международный журнал наук о Земле . 88 (3): 377–391. Бибкод : 1999IJEaS..88..377C . дои : 10.1007/s005310050273 . S2CID 140681313 .
- ^ Макдональд, ИК; Гинассо, Нидерланды; Эклесон, С.Г.; Амос, Дж. Ф.; Дакворт, Р.; Сассен, Р.; Брукс, Дж. М. (1993). «Природные нефтяные пятна в Мексиканском заливе, видимые из космоса». Журнал геофизических исследований . 98 (C9): 16351. Бибкод : 1993JGR....9816351M . дои : 10.1029/93JC01289 .
- ^ Макдональд, ИК; Дж. Ф. Рейли младший; МЫ Лучшие; Р. Вкатарамайя; Р. Сассен; Н. С. Гуинассо младший; Дж. Амос (1996). Инвентаризация с помощью дистанционного зондирования активных нефтяных выходов и хемосинтетических сообществ на севере . Американская ассоциация геологов-нефтяников. стр. 27–37. ISBN 978-0-89181-345-3 .
{{cite book}}:|work=игнорируется ( помогите ) - ^ И.Р. Макдональд, изд. (1998). «Стабильность и изменения в хемосинтетических сообществах Мексиканского залива» (PDF) . Министерство внутренних дел США: Исследование OCS MMS 98-0034: Подготовлено Группой геохимических и экологических исследований Техасского университета A&M. Архивировано из оригинала (PDF) 29 декабря 2016 года . Проверено 17 июля 2016 г.
- ^ Пиньейру, LM; Иванов, М.К.; Сауткин А.; Ахаманов Г.; Магальяйнс, В.Х.; Волконская А.; Монтейро, Дж. Х.; Сомоса, Л.; Гарднер, Дж.; Хамуни, Н.; Кунья, MR (2003). «Грязевые вулканы в Кадисском заливе: результаты похода ТТР-10». Морская геология . 195 (1–4): 131–151. Бибкод : 2003МГеол.195..131П . дои : 10.1016/S0025-3227(02)00685-0 .
- ↑ Перейти обратно: Перейти обратно: а б с д и Милославич П.; Кляйн, Э.; Диас, Дж. М.; Эрнандес, CE; Бигатти, Г. (2011). «Морское биоразнообразие на атлантическом и тихоокеанском побережьях Южной Америки: знания и пробелы» . ПЛОС ОДИН . 6 (1): e14631. Бибкод : 2011PLoSO...614631M . дои : 10.1371/journal.pone.0014631 . ПМК 3031619 . ПМИД 21304960 .
- ↑ Перейти обратно: Перейти обратно: а б с д и ж Дановаро, Р.; Компания, JB; Коринальдези, К.; Д'Онгиа, Г.; Галил, Б. (2010). «Глубоководное биоразнообразие в Средиземном море: известное, неизвестное и непознаваемое» . ПЛОС ОДИН . 5 (8): e11832. Бибкод : 2010PLoSO...511832D . дои : 10.1371/journal.pone.0011832 . ПМК 2914020 . ПМИД 20689848 .
- ^ Саутвард Э., Андерсен А., Урдез С. (подано в 2010 г.). « Lamellibrachia anaximandri nsp., новый вестиментиферовый трубчатый червь из Средиземноморья (Annelida)». Зоосистема .
- ^ Зиттер, Т.А. С; Генри, П.; Алоизи, Г.; Делайг, Г.; Чагатай, Миннесота (2008). «Холод просачивается вдоль главного Мраморного разлома в Мраморном море (Турция)» (PDF) . Глубоководные исследования. Часть I: Статьи океанографических исследований . 55 (4): 552–570. Бибкод : 2008DSRI...55..552Z . дои : 10.1016/j.dsr.2008.01.002 .
- ^ Фишер, Д.; Борманн, Г.; Забель, М.; Кастен, С. (апрель 2009 г.): Геохимическое зонирование и характеристики холодных просачиваний вдоль континентальной окраины Макрана у берегов Пакистана, тезисы конференции Генеральной ассамблеи EGU. Проверено 19 ноября 2020 г.
- ↑ Перейти обратно: Перейти обратно: а б Чен, З.; Хуанг, Китай; Чжао, М.; Ян, В.; Чиен, C.-W.; Чен, М.; Ян, Х.; Матияма, Х.; Лин, С. (2011). «Характеристики и возможное происхождение самородного алюминия в отложениях холодного просачивания северо-востока Южно-Китайского моря». Журнал азиатских наук о Земле . 40 (1): 363–370. Бибкод : 2011JAESc..40..363C . дои : 10.1016/j.jseaes.2010.06.006 .
- ↑ Перейти обратно: Перейти обратно: а б Кацунори (2008.ISBN) Фудзикура , (Dai 1-han ed.). Hadano-shi: Tōkai Daigaku Shuppankai. p. 450. . 978-4-486-01787-5 .
- ↑ Перейти обратно: Перейти обратно: а б с Фудзикура, К.; Линдси, Д.; Китазато, Х.; Нисида, С.; Сираяма, Ю. (2010). «Морское биоразнообразие в водах Японии» . ПЛОС ОДИН . 5 (8): е11836. Бибкод : 2010PLoSO...511836F . дои : 10.1371/journal.pone.0011836 . ПМК 2914005 . ПМИД 20689840 .
- ^ Мияке, Хироси; Дзюн ХАСИМОТО; Синдзи ЦУЧИДА (2010). ) в трубке» (PDF) . JAMSTEC Deep Sea Research. «Метод наблюдения за поведением вестиментифского трубчатого червя (Lamellibrachia satsuma Проверено 30 марта 2012 г.
- ^ «Вблизи Бездны Челленджера в Марианской впадине была обнаружена экосистема, которая получает питательные вещества из мантийного материала — результаты работы пилотируемой исследовательской подводной лодки «Синкай 6500» . 7 февраля 2012 года. Архивировано из оригинала 23 сентября 2012 года . Проверено 29 марта 2012 года .
- ^ Охара, Ясухико; Марк К. Рейган; Кацунори Фуджикураб; Хироми Ватанабеб; Кацуёси Мичибаяшид; Теруаки Исииэ; Роберт Дж. Стернф; Игнасио Пуджанаф; Фернандо Мартинезг; Гийом Жирар; Джулия Рибейроф; Мэриджо Бронех; Наоаки Коморид; Масаси Кинод (21 февраля 2012 г.). «Экосистема, содержащая серпентиниты, в Южно-Марианском предгорье» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (8): 2831–2835. Бибкод : 2012PNAS..109.2831O . дои : 10.1073/pnas.1112005109 . ПМЦ 3286937 . ПМИД 22323611 .
- ^ «Изображения, полученные в ходе подводной съемки пилотируемым подводным исследовательским судном «Синкай 6500» в районе эпицентра землетрясения на тихоокеанском побережье Тохоку (Последние новости)» . Японское агентство по морским наукам и технологиям Земли 15 августа 2011 г. Получено . 29 марта 2012 года .
- ^ Кавагуччи, синсук; Iho Hirai; ). Нарушение глубоководной среды, вызванное землетрясением Тохоку M9.0» Научные отчеты . 2 270. Bibcode : 2012NatSR...2E.270K : 10.1038 / srep00270 . : . « doi .
- ↑ Перейти обратно: Перейти обратно: а б с Гордон, ДП; Бомонт, Дж.; МакДиармид, А.; Робертсон, Д.А.; Ахён, С.Т. (2010). «Морское биоразнообразие Аотеароа, Новая Зеландия» . ПЛОС ОДИН . 5 (8): е10905. Бибкод : 2010PLoSO...510905G . дои : 10.1371/journal.pone.0010905 . ПМК 2914018 . ПМИД 20689846 .
- ^ Льюис, КБ; Маршалл, бакалавр (1996). «Просачивающаяся фауна и другие индикаторы обезвоживания, богатого метаном, на сходящихся окраинах Новой Зеландии». Новозеландский журнал геологии и геофизики . 39 (2): 181–200. Бибкод : 1996NZJGG..39..181L . дои : 10.1080/00288306.1996.9514704 .
- ^ Орпин, Арканзас (1997). «Доломитовые трубы как возможное свидетельство выброса береговой жидкости, самая верхняя часть континентального склона Отаго, юг Новой Зеландии». Морская геология . 138 (1–2): 51–67. Бибкод : 1997МГеол.138...51О . дои : 10.1016/S0025-3227(96)00101-6 .
- ↑ Перейти обратно: Перейти обратно: а б Бако, Арканзас; Роуден, А.А.; Левин, Луизиана; Смит, ЧР; Боуден, Д. (2009). «Первоначальная характеристика фаунистических сообществ холодного просачивания на окраине Новой Зеландии». Морская геология . 272 (1–4): 251–259. Бибкод : 2010МГеол.272..251Б . дои : 10.1016/j.margeo.2009.06.015 .
- ^ Селланес, Дж.; Нейра, К.; Кирога, Э.; Тейшидо, Н. (2010). «Разнообразие вдоль и поперек чилийской окраины: континентальный склон, охватывающий градиенты кислорода и бентосные места обитания метана». Морская экология . 31 (1): 111–124. Бибкод : 2010MarEc..31..111S . дои : 10.1111/j.1439-0485.2009.00332.x . hdl : 10261/56612 .
- ↑ Перейти обратно: Перейти обратно: а б Селланес, Дж.; Кирога, Э.; Нейра, К. (2008). «Структура сообщества мегафауны и трофические взаимоотношения в недавно обнаруженном районе метанового просачивания Консепсьон, Чили, ~ 36 ° ю.ш.» . Журнал морских наук ICES . 65 (7): 1102–1111. doi : 10.1093/icesjms/fsn099 .
- ^ Селланес, Дж.; Кирога, Э.; Галлардо, Вирджиния (2004). «Первое прямое свидетельство просачивания метана и связанных с ним хемосинтетических сообществ в батиальной зоне у побережья Чили». Журнал Морской биологической ассоциации Великобритании . 84 (5): 1065–1066. Бибкод : 2004JMBUK..84.1065S . дои : 10.1017/S0025315404010422h . S2CID 85948533 .
- ↑ Перейти обратно: Перейти обратно: а б Галлардо, Вирджиния; Эспиноза, К. (2007). Гувер, Ричард Б; Левин, Гилберт V; Розанов Алексей Юрьевич; Дэвис, Пол К.В. (ред.). «Крупные многоклеточные нитчатые бактерии в зоне минимума кислорода в восточной части южной части Тихого океана: забытая биосфера». Учеб. ШПИОН . Инструменты, методы и миссии астробиологии X. 6694 : 66941H–11. Бибкод : 2007SPIE.6694E..1HG . дои : 10.1117/12.782209 . S2CID 121829940 .
- ↑ Перейти обратно: Перейти обратно: а б Барри, СП; Кук, С.; Конвей, К.В. (2010). «Холодные просачивания и донная среда обитания на тихоокеанской окраине Канады». Исследования континентального шельфа . 31 (2 Дополнение 1): S85–S92. Бибкод : 2011CSR....31S..85V . дои : 10.1016/j.csr.2010.02.013 .
- ^ Лоренсон Т.Д., Квенволден К.А., Хостеттлер Ф.Д., Розенбауэр Р.Дж., Мартин Дж.Б. и Оранж Д.Л. (1999). «Углеводороды, связанные с процессом выброса жидкости в залив Монтерей, Калифорния» . Геологической службы США Тихоокеанский центр прибрежных и морских наук .
- ^ Гоффреди С.К. и Барри Дж.П. (2000). «Факторы, регулирующие продуктивность хемоавтотрофных симбиозов; с акцентом на Calyptogena kilmeri и Calyptogena pacifica ». Плакат, Научно-исследовательский институт аквариумов залива Монтерей . по состоянию на 3 февраля 2011 г. PDF .
- ^ Бернхард, Дж. М.; Бак, КР; Барри, JP (2001). «Биота холодного просачивания залива Монтерей: сообщества, численность и ультраструктура живых фораминифер». Глубоководные исследования. Часть I: Статьи океанографических исследований . 48 (10): 2233–2249. Бибкод : 2001DSRI...48.2233B . дои : 10.1016/S0967-0637(01)00017-6 .
- ^ Лонсдейл, П. (1979). «Глубоководный гидротермальный участок на сдвиговом разломе». Природа . 281 (5732): 531–534. Бибкод : 1979Natur.281..531L . дои : 10.1038/281531a0 . S2CID 4310057 .
- ^ Полл, СК; В. Р. Нормарк; В. Усслер III; Д. У. Кэсс и Р. Китен (2008). «Связь между активной деформацией морского дна, образованием насыпей, а также ростом и накоплением газовых гидратов на морском дне бассейна Санта-Моники, у побережья Калифорнии». Морская геология . 250 (3–4): 258–275. Бибкод : 2008МГеол.250..258П . дои : 10.1016/j.margeo.2008.01.011 .
- ↑ Перейти обратно: Перейти обратно: а б Мэлони, Дж. М.; Групе, Б.М.; Пасулька, АЛ; Доусон, Канзас; Кейс, ДХ; Фридер, Калифорния; Левин, Луизиана; Дрисколл, Северо-Запад (2015). «Границы транспрессионных сегментов в системах сдвиговых разломов на шельфе южной Калифорнии: последствия для вытеснения жидкости и среды обитания холодного просачивания» (PDF) . Письма о геофизических исследованиях . 42 (10): 4080–4088. Бибкод : 2015GeoRL..42.4080M . дои : 10.1002/2015GL063778 .
- ↑ Перейти обратно: Перейти обратно: а б с д Гриффитс, HJ (2010). «Морское биоразнообразие Антарктики – что мы знаем о распространении жизни в Южном океане?» . ПЛОС ОДИН . 5 (8): e11683. Бибкод : 2010PLoSO...511683G . дои : 10.1371/journal.pone.0011683 . ПМК 2914006 . ПМИД 20689841 .
- ^ Лю, Чао; Ань, Сяньинь; Алгео, Томас Дж.; Муннеке, Аксель; Чжан, Юцзе; Чжу, Тунсин (февраль 2021 г.). «Отложения углеводородов в формации Энджи нижней перми, Центральный блок Лхаса, Тибет» . Исследования Гондваны . 90 : 258–272. Бибкод : 2021GondR..90..258L . дои : 10.1016/j.gr.2020.10.017 . S2CID 230530430 . Проверено 15 марта 2023 г.
- ^ Каим, А.; Дженкинс, Р.; Варен, А. (2008). «Прованниды и прованнидоподобные брюхоногие моллюски из позднемеловых холодных просачиваний Хоккайдо (Япония) и летопись окаменелостей прованнид (Gastropoda: Abyssochrysoidea)» . Зоологический журнал Линнеевского общества . 154 (3): 421–436. дои : 10.1111/j.1096-3642.2008.00431.x .
- ^ Аману, Казутака; Дженкинс, Роберт Г.; Сако, Юкио; Охара, Масааки; Киль, Штеффен (1 октября 2013 г.). «Палеогеновое глубоководное сообщество метана из Хонсю, Япония» . Палеогеография, Палеоклиматология, Палеоэкология . 387 : 126–133. Бибкод : 2013PPP...387..126A . дои : 10.1016/j.palaeo.2013.07.015 . Проверено 16 ноября 2022 г.
- ^ Конти, С.; Фиорони, К.; Фонтана, Д. (2017). «Корреляция фаз эволюции карбонатов шельфа с эпизодами выброса жидкости в переднем миоцене, северные Апеннины, Италия». Морская и нефтяная геология . 79 : 351–359. Бибкод : 2017МарPG..79..351C . дои : 10.1016/j.marpetgeo.2016.11.003 . hdl : 11380/1119044 .
- ^ Георгиева Магдалена Н.; Полл, Чарльз К.; Литтл, Криспин Т.С.; Макганн, Мэри; Сахи, Диана; Кондон, Дэниел; Лундстен, Лонни; Пьюси, Джек; Кэсс, Дэвид В.; Вриенгук, Роберт К. (19 марта 2019 г.). «Открытие обширного глубоководного рифа ископаемых серпулид, связанного с холодным просачиванием, бассейн Санта-Моники, Калифорния» . Границы морской науки . 6 : 1–21. дои : 10.3389/fmars.2019.00115 .
- ^ Галил, Б.С.; Голик, А.; Туркай, М. (1995). «Мусор на дне моря: исследование морского дна в Восточном Средиземноморье». Бюллетень о загрязнении морской среды . 30 (1): 22–24. Бибкод : 1995МарПБ..30...22Г . дои : 10.1016/0025-326x(94)00103-g .
- ^ Гальгани, Ф.; Жоне, С.; Кампильо, А.; Генеган, X.; Его, Э. (1995). «Распределение и обилие мусора на континентальном шельфе северо-западной части Средиземного моря». Бюллетень о загрязнении морской среды . 30 (11): 713–717. Бибкод : 1995MarPB..30..713G . дои : 10.1016/0025-326x(95)00055-r .
- ^ Рихтер, Т.О.; де Стигтер, ХК; Бур, В.; Иисус, CC; ван Веринг, TCE (2009). «Распространение природного и антропогенного свинца через подводные каньоны на окраине Португалии». Глубоководные исследования . Часть I. 56 (2): 267–282. Бибкод : 2009DSRI...56..267R . дои : 10.1016/j.dsr.2008.09.006 .
- ^ Ротлант, Г.; Хольгадо, А.Е.; Сарда, Ф.; Абалос, М.; Компания, JB (2006). «Соединения диоксина в глубоководной розовой креветке Aristeus антеннатус (Risso, 1816) по всему Средиземному морю». Глубоководные исследования . Часть I. 53 (12): 1895–1906. Бибкод : 2006DSRI...53.1895R . дои : 10.1016/j.dsr.2006.09.004 .
- ^ Унгер, Массачусетс; Харви, Э.; Вадас, Г.Г.; Веккьоне, М. (2008). «Стойкие загрязнители у девяти видов глубоководных головоногих моллюсков». Бюллетень о загрязнении морской среды . 56 (8): 1486–1512. Бибкод : 2008MarPB..56.1498U . doi : 10.1016/j.marpolbul.2008.04.018 . ПМИД 18501382 .
- ^ Смит, КЛ; Рул, штат Ха; Бетт, Би Джей; Биллет, DSM; Лэмпитт, Р.С. (2009). «Климат, круговорот углерода и глубоководные экосистемы океана» . ПНАС . 106 (46): 19211–19218. Бибкод : 2009PNAS..10619211S . дои : 10.1073/pnas.0908322106 . ПМК 2780780 . ПМИД 19901326 .
Дальнейшее чтение
[ редактировать ]- Брайт, М.; Слива, К.; Риавитц, Луизиана; Николов Н.; Мартинес Арбису, П.; Кордес, Э.Э.; Голлнер, С. (2010). «Эпизоойный многоклеточный мейобентос, связанный с скоплениями трубчатых червей и мидий из холодных просачиваний северной части Мексиканского залива» . Глубоководные исследования. Часть II: Актуальные исследования в океанографии . 57 (21–23): 1982–1989. Бибкод : 2010DSRII..57.1982B . дои : 10.1016/j.dsr2.2010.05.003 . ПМК 2995211 . ПМИД 21264038 .
- немецкий, ЧР; Рамирес-Льодра, Э.; Бейкер, MC; Тайлер, Пенсильвания; Научно-руководящий комитет по шахматам (2011). «Исследование глубоководных хемосинтетических экосистем в ходе переписи морской жизни в течение десятилетия и в последующий период: предлагаемая дорожная карта глубоководных океанов» . ПЛОС ОДИН . 6 (8): e23259. Бибкод : 2011PLoSO...623259G . дои : 10.1371/journal.pone.0023259 . ПМК 3150416 . ПМИД 21829722 .
- Ллойд, КГ; Альберт, Д.Б.; Биддл, Дж. Ф.; Шантон, Япония; Писарро, О.; Теске, А. (2010). «Пространственная структура и активность осадочных микробных сообществ, лежащих в основе мата видов Beggiatoa в просачивании углеводородов в Мексиканском заливе» . ПЛОС ОДИН . 5 (1): е8738. Бибкод : 2010PLoSO...5.8738L . дои : 10.1371/journal.pone.0008738 . ПМК 2806916 . ПМИД 20090951 .
- Метаксас, А.; Келли, штат Нью-Йорк (2010). «Различаются ли поставки и набор личинок в зависимости от хемосинтетической среды глубокого моря?» . ПЛОС ОДИН . 5 (7): e11646. Бибкод : 2010PLoSO...511646M . дои : 10.1371/journal.pone.0011646 . ПМК 2906503 . ПМИД 20657831 .
- Родригес, Э.; Дейли, М. (2010). «Филогенетические связи между глубоководными и хемосинтетическими актиниями: Actinoscyphiidae и Actinostolidae (Actiniaria: Mesomyaria)» . ПЛОС ОДИН . 5 (6): е10958. Бибкод : 2010PLoSO...510958R . дои : 10.1371/journal.pone.0010958 . ПМК 2881040 . ПМИД 20532040 .
- Сибуэт, М.; Олу, К. (1998). «Биогеография, биоразнообразие и зависимость от флюидов глубоководных сообществ холодного просачивания на активных и пассивных окраинах». Глубоководные исследования. Часть II: Актуальные исследования в океанографии . 45 (1–3): 517–567. Бибкод : 1998DSRII..45..517S . дои : 10.1016/S0967-0645(97)00074-X .
- Винн, О.; Гриневич, К; Литтл, CTS; Накрем, ХА (2014). «Бореальная фауна серпулид из волжско-рязанских (последняя юра-ранний мел) шельфовых отложений и выходов углеводородов со Шпицбергена» . Геодиверситас . 36 (4): 527–540. дои : 10.5252/g2014n4a2 . S2CID 129587761 . Проверено 9 января 2014 г.
- Винн, О.; Куприянова Е.К.; Киль, С. (2013). «Серпулиды (Annelida, Polychaeta) от мелового периода до современных выходов углеводородов: экологические и эволюционные закономерности» . Палеогеография, Палеоклиматология, Палеоэкология . 390 : 35–41. Бибкод : 2013PPP...390...35В . дои : 10.1016/j.palaeo.2012.08.003 . Проверено 9 января 2014 г.



