Соленое болото


Солончак приливной , солончак или солончак , также известный как прибрежный солончак или приливное болото , представляет собой прибрежную экосистему в верхней прибрежной зоне между сушей и открытой соленой или солоноватой водой , которая регулярно затопляется приливами. Здесь преобладают густые заросли солеустойчивых растений, таких как травы , травы или невысокие кустарники . [1] [2] Эти растения имеют наземное происхождение и необходимы для устойчивости солончаков при улавливании и связывании отложений . Солончаки играют большую роль в водной пищевой сети и доставке питательных веществ в прибрежные воды. Они также поддерживают наземных животных и обеспечивают защиту побережья . [2]
Исторически солончаки подвергались угрозе из-за плохо реализованных методов управления прибрежными зонами , когда земли использовались для нужд человека или загрязнялись сельским хозяйством, расположенным вверх по течению, или другими промышленными прибрежными видами использования. Кроме того, повышение уровня моря, вызванное изменением климата , ставит под угрозу другие болота из-за эрозии и затопления болот, которые в противном случае были бы приливными. [3] [4] Однако недавнее признание как экологами, так и обществом в целом важности соленых болот для биоразнообразия, экологической продуктивности и других экосистемных услуг , таких как связывание углерода , привело к увеличению масштабов восстановления и управления солончаками с 1980-х годов.
Основная информация [ править ]


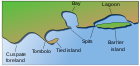
Солончаки встречаются на береговых линиях с низким энергопотреблением в умеренных и высоких широтах. [5] который может быть стабильным, всплывающим или погружающимся в зависимости от того, больше ли осадконакопление , равно или меньше относительного повышения уровня моря ( скорость опускания плюс изменение уровня моря) соответственно. Обычно эти береговые линии состоят из илистых или песчаных отмелей (известных также как приливные отмели или сокращенно илистые отмели ), которые питаются отложениями впадающих рек и ручьев. [6] К ним обычно относятся защищенные места, такие как насыпи, устья рек и подветренная сторона барьерных островов и кос . В тропиках и субтропиках их заменяют мангровые заросли ; территория, отличающаяся от солончака тем, что вместо травянистых растений здесь преобладают солеустойчивые деревья. [1]
Большинство солончаков имеют низкий рельеф с невысокими высотами, но обширную территорию, что делает их чрезвычайно популярными среди населения. [7] Солончаки расположены среди различных форм рельефа в зависимости от их физических и геоморфологических условий. К таким болотным формам рельефа относятся дельтовые болота, устьевые болота, заграждения, открытые берега, заливы и затопленные долинные болота. Дельтовые болота связаны с крупными реками , многие из которых встречаются в Южной Европе, например , Камарг во Франции в дельте Роны или дельта Эбро в Испании. Они также широко распространены в реках дельты Миссисипи в США . [2] В Новой Зеландии большинство солончаков расположено в верховьях эстуариев, в районах с небольшим волновым воздействием и высоким уровнем осадков. [8] Такие болота расположены в региональном парке Аухиту в Окленде , в устье Манавату и в устье Эйвон-Хиткот/Ихутай в Крайстчерче . Забарьерные болота чувствительны к изменению формы барьеров, со стороны суши, от которых они образовались. [2] Они распространены вдоль большей части восточного побережья США и Фризских островов . Большие неглубокие прибрежные заливы могут содержать солончаки, например залив Моркам и Портсмут в Великобритании , а также залив Фанди в Северной Америке. [2]
Солончаки иногда включаются в лагуны, и разница не очень заметна; Венецианская лагуна в Италии , например, состоит из таких видов животных и/или живых организмов, принадлежащих к этой экосистеме. Они оказывают большое влияние на биоразнообразие региона. Экология солончаков включает в себя сложные пищевые сети, которые включают первичных продуцентов (сосудистые растения, макроводоросли, диатомовые водоросли, эпифиты и фитопланктон), первичных потребителей (зоопланктон, макрозоа, моллюски, насекомые) и вторичных потребителей. [9]
Низкая физическая энергия и высокая трава служат убежищем для животных. Многие морские рыбы используют солончаки в качестве мест нагула своих детенышей, прежде чем они переберутся в открытые воды. Птицы могут выращивать птенцов среди высокой травы, потому что болота обеспечивают как убежище от хищников, так и обильные источники пищи, включая рыбу, пойманную в водоемах, насекомых, моллюсков и червей. [10]
Встречается мире во всем
Солончаки в 99 странах (по сути, во всем мире) были нанесены на карту Макоуэном и др. 2017. [11] В общей сложности 5 495 089 гектаров нанесенных на карту солончаков в 43 странах и территориях представлены в многоугольном шейп-файле Географической информационной системы. Эта оценка находится на относительно нижнем уровне предыдущих оценок (2,2–40 млн га). Более позднее исследование консервативно оценило глобальную протяженность солончаков в 90 800 км. 2 (9 080 000 га). [12] Самые обширные солончаки в мире находятся за пределами тропиков, в частности, на низменных, свободных ото льда побережьях, заливах и эстуариях Северной Атлантики, которые хорошо представлены в глобальном наборе полигональных данных. [11]
Формирование [ править ]
Формирование начинается, когда приливные отмели поднимаются над уровнем моря за счет нарастания отложений , а затем скорость и продолжительность приливных наводнений уменьшаются, так что растительность может колонизировать открытую поверхность. [13] Появление размножений пионерных видов, таких как семена или части корневищ , сочетается с созданием подходящих условий для их прорастания и укоренения в процессе колонизации. [14] Когда реки и ручьи достигают низкого уклона приливных отмелей, скорость стока снижается, и взвешенные осадки оседают на приливную плоскую поверхность, чему способствует эффект подпора восходящего прилива. [6] Маты нитчатых сине-зеленых водорослей могут при контакте фиксировать частицы ила и глины на своих липких оболочках. [15] что также может повысить эрозионную устойчивость отложений. [16] Это способствует процессу нарастания отложений, позволяя колонизирующим видам (например, Salicornia размножаться spp.). Эти виды сохраняют осадок, смываемый приливом вокруг своих стеблей и листьев, и образуют низкие илистые холмики, которые в конечном итоге сливаются, образуя террасы осадконакопления, росту которых вверх способствует подповерхностная корневая сеть, связывающая осадки. [17] Как только растительность установится на террасах осадконакопления, дальнейшее улавливание и аккреция наносов может привести к быстрому росту поверхности болота вверх, что приводит к быстрому уменьшению глубины и продолжительности приливных наводнений. В результате на этой территории могут обитать конкурентоспособные виды, предпочитающие возвышенности по отношению к уровню моря, и часто развивается смена растительных сообществ . [13]
наводнения и Приливные зонирование растительности

Прибрежные солончаки можно отличить от наземных мест обитания по ежедневным приливным потокам, которые возникают и постоянно затопляют эту территорию. [1] Это важный процесс доставки наносов, питательных веществ и воды для растений в болото. [7] На возвышенностях в верхней болотной зоне приливный приток гораздо меньше, что приводит к более низким уровням солености . [1] Засоление почв в нижней болотной зоне довольно постоянное из-за ежедневных ежегодных приливов. Однако в верховьях болота проявляется изменчивость солености в результате менее частых наводнений и изменений климата. Осадки могут снизить соленость, а суммарное испарение может увеличить ее уровень в засушливые периоды. [1] В результате возникают микроместа обитания, заселенные разными видами флоры и фауны в зависимости от их физиологических возможностей. Флора солончаков дифференцируется по уровням в зависимости от индивидуальной переносимости растений к солености и уровню грунтовых вод. Растительность, обнаруженная у воды, должна быть способна выдерживать высокие концентрации соли, периодическое погружение в воду и определенное движение воды, в то время как растения, расположенные дальше вглубь болота, могут иногда испытывать засушливые условия с низким содержанием питательных веществ. Было обнаружено, что верхние болотные зоны ограничивают виды из-за конкуренции и отсутствия защиты среды обитания, тогда как нижние болотные зоны определяются способностью растений переносить физиологические стрессы, такие как соленость, погружение в воду и низкий уровень кислорода. [18] [19]

Солончаки Новой Англии подвержены сильному приливно-отливному влиянию и демонстрируют отчетливые закономерности зонирования. [19] На низинных болотистых участках с сильным приливным разливом доминирует монокультура корицы гладкой , Spartina alterniflora зоны соляного сена, Spartina patens , черного камыша, Juncus gerardii и кустарника Iva frutescens соответственно. , затем продвигаясь к суше, наблюдаются [18] Все эти виды имеют разную переносимость, поэтому разные зоны вдоль болота лучше всего подходят для каждого человека.
Видовое разнообразие растений относительно невелико, поскольку флора должна быть толерантна к соли, полному или частичному погружению и бескислородному иловому субстрату. Наиболее распространенными растениями солончаков являются солянка ( Salicornia spp.) и корзинка ( Spartina spp.), Распространенные по всему миру. Зачастую они являются первыми растениями, поселившимися на илистой равнине и начавшими свое экологическое развитие в солончаках. Их побеги поднимают основной поток прилива над поверхностью грязи, а их корни распространяются в субстрат, стабилизируют липкую грязь и переносят в нее кислород, чтобы другие растения также могли прижиться. Такие растения, как морская лаванда ( виды Limonium ), подорожники ( виды Plantago ), а также различные виды осоки и камыша, растут после того, как на иле зарастают виды-первопроходцы .
Солончаки весьма фотосинтетически активны и являются чрезвычайно продуктивной средой обитания. Они служат хранилищами большого количества органического вещества и полны разложения, которое питает широкую пищевую цепь организмов от бактерий до млекопитающих. Многие из галофитных растений, например корда, вообще не поедаются высшими животными, а отмирают и разлагаются, становясь пищей для микроорганизмов, которые, в свою очередь, становятся пищей для рыб и птиц.
роль приливных Захват осадков, аккреция и ручьев

Факторы и процессы, влияющие на скорость и пространственное распределение нарастания наносов в пределах солончака, многочисленны. Отложение осадков может происходить, когда болотные виды обеспечивают поверхность, к которой прилипают отложения, с последующим осаждением на поверхность болота, когда осадки отслаиваются во время отлива. [13] Количество наносов, прилипших к видам солончаков, зависит от типа болотных видов, близости вида к источнику наносов, количества растительной биомассы и высоты над уровнем моря. [20] Например, при исследовании приливных болот восточного острова Чонгмин и острова Цзюдуаньша в устье реки Янцзы , Китай, количество отложений, прилипших к видам Spartina alterniflora , Phragmites australis и Scirpus mariqueter, уменьшалось по мере удаления от самых высоких уровней. концентраций взвешенных отложений (обнаруженных на краю болота, граничащего с приливными ручьями или илистыми отмелями); снизился у тех видов, которые обитали на самых высоких высотах, где наблюдались наименьшая частота и глубина приливных наводнений; и увеличивалась с увеличением биомассы растений. Spartina alterniflora , к которой прилипло больше всего отложений, может составлять> 10% от общего накопления отложений на поверхности болота в результате этого процесса. [20]
Виды солончаков также способствуют накоплению отложений, уменьшая скорость течения и способствуя осаждению отложений из взвеси. [13] Скорость течения может быть уменьшена, поскольку стебли высоких болотных видов вызывают гидравлическое сопротивление, что сводит к минимуму повторное взвешивание отложений и способствует осаждению. [21] Было показано, что измеренные концентрации взвешенных отложений в толще воды уменьшаются от открытой воды или приливных ручьев, прилегающих к краю болота, к внутренней части болота. [20] [21] [22] вероятно, в результате непосредственного выселения на поверхность болота под влиянием болотного полога. [21] [22]
Наводнению и отложению наносов на поверхности болота также способствуют приливные ручьи. [22] которые являются общей чертой солончаков. [6] [13] [17] [22] [23] Их типично древовидная и извилистая форма обеспечивает приливу пути для подъема и затопления поверхности болота, а также для отвода воды. [17] и они могут способствовать отложению большего количества отложений, чем солончаки, граничащие с открытым океаном. [23] Отложение отложений коррелирует с размером отложений: более крупные отложения будут откладываться на возвышенностях (ближе к ручью), чем более мелкие отложения (дальше от ручья). Размер отложений также часто коррелирует с конкретными микроэлементами металлов, и, таким образом, приливные ручьи могут влиять на распределение и концентрацию металлов в солончаках, что, в свою очередь, влияет на биоту. [24] Однако солончакам не нужны приливные ручьи, чтобы облегчить поток наносов по их поверхности. [21] хотя солончаки с такой морфологией, по-видимому, редко изучаются.
Важное значение имеет возвышение болотных видов; те виды, которые обитают на более низких высотах, испытывают более длительные и частые приливные наводнения и, следовательно, имеют возможность отложения большего количества наносов. [20] [25] Виды, обитающие на возвышенностях, могут извлечь выгоду из большей вероятности затопления во время самых высоких приливов, когда увеличенная глубина воды и поверхностные потоки болот могут проникать во внутреннюю часть болота. [22]
Человеческое воздействие [ править ]

Побережье является очень привлекательным природным объектом для человека благодаря своей красоте, ресурсам и доступности. По оценкам, по состоянию на 2002 год более половины населения мира проживало в пределах 60 км от прибрежной береговой линии. [2] делая береговые линии весьма уязвимыми для воздействия человека в результате повседневной деятельности, которая оказывает давление на окружающую природную среду. В прошлом солончаки воспринимались как прибрежные «пустыни», что приводило к значительным потерям и изменениям этих экосистем в результате освоения земель для сельского хозяйства, городского развития, добычи соли и отдыха. [7] [26] [27] Косвенные последствия деятельности человека, такие как нагрузка азотом, также играют важную роль в районе солончаков. Солончаки могут пострадать от вымирания в верховьях и вымирания в низинах. По оценкам исследования, опубликованного в 2022 году, 22% потерь солончаков в период с 1999 по 2019 годы были вызваны прямыми человеческими факторами, определяемыми как наблюдаемые действия, происходящие в месте обнаруженных изменений, такие как преобразование в аквакультуру, сельское хозяйство, освоение прибрежных зон или другие физические действия. структуры. [12] Кроме того, 30% прироста солончаков за тот же период также было обусловлено прямыми факторами, такими как мероприятия по восстановлению или модификации прибрежных зон для содействия обмену приливов и отливов. [12]
Мелиорация земель [ править ]
Рекультивация земель для сельского хозяйства путем преобразования болот в возвышенности исторически была обычной практикой. [7] Дамбы часто строились, чтобы учесть этот сдвиг в изменении земель и обеспечить защиту от наводнений вглубь страны. В последнее время приливные отмели также были освоены. [28] На протяжении веков домашний скот, такой как овцы и крупный рогатый скот, пасся на очень плодородных солончаковых землях. [1] [29] Мелиорация земель для сельского хозяйства привела ко многим изменениям, таким как сдвиги в структуре растительности, отложениям осадков, засоленности, расходу воды, утрате биоразнообразия и высокому поступлению питательных веществ. Было предпринято множество попыток искоренить эти проблемы, например, в Новой Зеландии кордграсс Spartina anglica был завезен из Англии в устье реки Манавату в 1913 году, чтобы попытаться вернуть земли устья для ведения сельского хозяйства. [8] Сдвиг структуры от голой приливной равнины к пастбищам произошел в результате увеличения отложений, и кордграсс распространился на другие устья вокруг Новой Зеландии. Местные растения и животные боролись за выживание, поскольку чужаки вытесняли их. В настоящее время предпринимаются усилия по удалению этих видов кордграсса, поскольку ущерб постепенно осознается.
В устье реки Блит в Саффолке на востоке Англии мелиорации среднего устья (болота Анхель и Балкэмп), которые были заброшены в 1940-х годах, были заменены приливными отмелями с уплотненными почвами, оставшимися в результате сельскохозяйственного использования, покрытыми тонким слоем ила. За последние 60–75 лет произошла небольшая колонизация растительности, что объясняется сочетанием высоты поверхности, слишком низкой для развития видов-первопроходцев, и плохого дренажа из уплотненных сельскохозяйственных почв, действующих как водоупор . [30] Наземные почвы такого типа должны перейти от пресной к соленой поровой воде за счет изменения химического состава и структуры почвы, сопровождаемого свежим отложением устьевых отложений, прежде чем сможет утвердиться солончаковая растительность. [14] Структуру растительности, видовое богатство и состав растительных сообществ солончаков, естественным образом восстановившихся на мелиорированных сельскохозяйственных землях, можно сравнить с соседними эталонными солончаками, чтобы оценить успех регенерации болот. [31]
Сельское хозяйство по течению вверх
Обработка земель выше по течению от солончаков может привести к увеличению поступления ила и повышению скорости нарастания первичных отложений на приливных отмелях, так что виды-первопроходцы могут распространиться дальше на отмели и быстро расти вверх от уровня приливного затопления. В результате болотные поверхности в этом режиме могут иметь обширный обрыв на краю моря. [32] В устье острова Плам, штат Массачусетс (США), стратиграфические керны показали, что в течение 18 и 19 веков болото распространилось по сублиторальной и илистой среде, увеличившись в площади с 6 км2. 2 до 9 км 2 после того, как европейские поселенцы вырубили леса вверх по течению и увеличили скорость поступления наносов. [33]
Городское развитие азота нагрузка и
Преобразование болотистых местностей в возвышенности для сельского хозяйства в прошлом столетии было омрачено преобразованием для городского развития. Прибрежные города по всему миру вторглись на бывшие солончаки, а в США рост городов стал искать места для захоронения отходов в солончаках. Загрязнение эстуариев органическими, неорганическими и токсичными веществами в результате городского развития или индустриализации является мировой проблемой. [28] а отложения на солончаках могут уносить это загрязнение, оказывая токсическое воздействие на виды флоры и фауны. [32] Городское развитие солончаков замедлилось примерно с 1970 года из-за растущего осознания экологическими группами того, что они предоставляют полезные экосистемные услуги . [7] Это высокопродуктивные экосистемы , и когда чистая продуктивность измеряется в граммах −2 тот −1 с ними могут сравниться только тропические леса. [28] Кроме того, они могут помочь уменьшить волновую эрозию морских дамб, предназначенных для защиты низменных участков суши от волновой эрозии. [14]
Отмена натурализации границ солончаков со стороны суши в результате вторжения городов или промышленности может иметь негативные последствия. В устье реки Эйвон-Хиткот/Ихутай, Новая Зеландия, численность видов и физические свойства окружающих окраин были тесно связаны, и было обнаружено, что большая часть солончаков обитает вдоль территорий с естественными окраинами в Эйвоне/Отакаро и Опавахо/ Выходы рек Хиткот; и наоборот, искусственные окраины содержали мало болотной растительности и ограничивали отступление к суше. [34] Остальные болота, окружающие эти городские районы, также находятся под огромным давлением со стороны человеческого населения, поскольку антропогенное обогащение азотом проникает в эти места обитания. Нагрузка азота в результате использования человеком косвенно влияет на солончаки, вызывая изменения в структуре растительности и инвазию чужеродных видов. [18]
Такие антропогенные воздействия, как сточные воды, городские стоки, сельскохозяйственные и промышленные отходы, попадают в болота из близлежащих источников. Солончаки ограничены азотом [18] [35] а с увеличением уровня питательных веществ, поступающих в систему в результате антропогенного воздействия , виды растений, связанные с солончаками, реструктурируются за счет изменения конкуренции. [7] Например, на солончаках Новой Англии наблюдается сдвиг в структуре растительности: S. alterniflora распространяется из нижних болот, где они преимущественно обитают, в верхнюю болотную зону. [18] Кроме того, на тех же болотах тростник Phragmites australis вторгся в ареал, распространившись на более низкие болота и став доминирующим видом. P. australis — агрессивный галофит, который может в больших количествах проникать на нарушенные территории, вытесняя местные растения. [7] [36] [37] Эта потеря биоразнообразия наблюдается не только у флоры, но и у многих животных, таких как насекомые и птицы, поскольку изменяются их среда обитания и пищевые ресурсы.
Повышение уровня моря [ править ]
Из-за таяния арктических морских льдов и теплового расширения океанов в результате глобального потепления уровень моря начал повышаться. Прогнозируется, что, как и в случае со всеми береговыми линиями, такое повышение уровня воды негативно повлияет на солончаки, затопив и размыв их. [38] [10] Повышение уровня моря приводит к увеличению количества зон открытой воды в солончаках. Эти зоны вызывают эрозию по краям, что приводит к дальнейшей эрозии болота в открытую воду, пока все болото не распадется. [39]
Хотя солончаки подвержены угрозам, связанным с повышением уровня моря, они также представляют собой чрезвычайно динамичную прибрежную экосистему. На самом деле солончаки могут справиться с повышением уровня моря: к 2100 году средний уровень моря может увеличиться на 0,6–1,1 метра. [40] Болота подвержены как эрозии, так и аккреции, что играет роль в так называемой биогеоморфической обратной связи. [41] Растительность солончаков захватывает осадки и остается в системе, что, в свою очередь, позволяет растениям лучше расти и, таким образом, растения лучше улавливают осадки и накапливают больше органических веществ. Эта петля положительной обратной связи потенциально позволяет уровням дна солончаков идти в ногу со скоростью повышения уровня моря. [40] Однако эта обратная связь также зависит от других факторов, таких как продуктивность растительности, поступление наносов, проседание земель, накопление биомассы, а также сила и частота штормов. [40] В исследовании, опубликованном Ü. СН Лучший в 2018 году, [40] они обнаружили, что биоаккумуляция была фактором номер один в способности солончаков справляться с темпами SLR. Устойчивость солончака зависит от того, насколько скорость повышения уровня дна будет выше, чем скорость повышения уровня моря, в противном случае болото будет захвачено и затоплено.
Накопление биомассы можно измерить в виде надземного накопления органической биомассы и подземного неорганического накопления посредством улавливания осадков и осаждения осадков из взвеси. [42] Растительность солончаков способствует увеличению осаждения отложений, поскольку замедляет скорость течения, разрушает турбулентные водовороты и помогает рассеивать энергию волн. Виды болотных растений известны своей толерантностью к повышенному воздействию соли из-за обычного затопления болот. Такие растения называются галофитами. Галофиты являются важной частью биоразнообразия солончаков и их способности адаптироваться к повышенному уровню моря. При повышенном уровне моря растительность солончаков, вероятно, будет более подвержена более частым наводнениям, и она должна быть адаптируемой или толерантной к последующему повышению уровня солености и анаэробным условиям. Существует общий предел высоты (над уровнем моря) для выживания этих растений: любое место ниже оптимальной линии приведет к бескислородным почвам из-за постоянного затопления, а слишком высокое значение выше этой линии будет означать опасный уровень засоления почвы из-за высокой скорости суммарного испарения в результате уменьшения погружения под воду. [42] Наряду с вертикальным накоплением отложений и биомассы необходимо также учитывать пространство для зарастания болотистых земель. Пространство для размещения — это земля, доступная для накопления дополнительных отложений и болотной растительности для латеральной колонизации. [43] Это боковое пространство размещения часто ограничено антропогенными сооружениями, такими как прибрежные дороги, дамбы и другие формы освоения прибрежных земель. Исследование Лизы М. Шиле, опубликованное в 2014 году, [44] обнаружили, что в диапазоне темпов повышения уровня моря болота с высокой продуктивностью растений были устойчивы к повышению уровня моря, но все они достигли высшей точки, когда для дальнейшего выживания требовалось жилое пространство. Наличие жилых помещений позволяет формировать новые среды обитания среднего и высокого уровня, а болотам избегать полного затопления.
Борьба с комарами [ править ]
Ранее в 20 веке считалось, что осушение солончаков поможет сократить популяцию комаров , таких как Aedes taeniorhynchus , черный солончаковый комар. Во многих местах, особенно на северо-востоке США, жители, а также местные и государственные учреждения вырыли прямолинейные канавы глубоко в болотистые равнины. Конечным результатом, однако, стало истощение среды обитания кильфиса . Рыба-киллифиш является хищником комаров , поэтому потеря среды обитания фактически привела к увеличению популяции комаров и отрицательно сказалась на болотных птицах , которые охотились на рыбу-киллифиша. Эти канавы все еще можно увидеть, несмотря на некоторые попытки их засыпать. [45]
и биотурбация Травоядность крабов

Повышенное поглощение азота болотными видами в листьях может привести к увеличению скорости роста листьев в зависимости от длины и увеличению уровня травоядности крабов. Роющий краб Neohelice granulata часто встречается в солончаках на юго-западе Атлантического океана, где популяции с высокой плотностью можно обнаружить среди популяций болотных видов Spartina densiflora и Sarcocornia perennis . В лагуне Мар-Чикита , к северу от Мар-дель-Плата , Аргентина , численность травоядных Neohelice granulata увеличилась, что, вероятно, является ответом на повышенную питательную ценность листьев на удобренных участках Spartina densiflora по сравнению с неудобренными участками. Независимо от того, были ли участки удобрены или нет, выпас Neohelice granulata также снижал скорость роста листьев по длине летом, одновременно увеличивая скорость их старения по длине . Возможно, этому способствовала повышенная эффективность грибков на ранах, оставленных крабами. [46]
В солончаках Кейп-Код , штат Массачусетс (США), наблюдается вымирание видов Spartina на берегах ручьев . отнес к травоядным (кордграсс), который краб Sesarma reticulatum . На 12 исследованных участках солончаков Кейп-Код на 10–90% берегов ручьев наблюдалось отмирание кордграсса в связи с сильно оголенным субстратом и высокой плотностью крабовых нор. Популяции Sesarma reticulatum растут, возможно, в результате деградации прибрежной пищевой сети в регионе. [47] Голые участки, оставленные в результате интенсивного выпаса кордовой травы Sesarma reticulatum на Кейп-Коде, пригодны для обитания другого роющего краба, Uca pugnax , который, как известно, не потребляет живые макрофиты. Было показано, что интенсивная биотурбация отложений солончаков в результате роющей деятельности этого краба резко снижает успешность прорастания семян Spartina alterniflora и Suaeda maritima и установленную выживаемость рассады либо за счет захоронения или обнажения семян, либо за счет выкорчевывания или захоронения укоренившихся сеянцев. [48] Однако биотурбация крабов может иметь и положительный эффект. В Новой Зеландии грязевой краб Helice crassa получил величественное прозвище «инженер экосистемы» за его способность создавать новые среды обитания и изменять доступ питательных веществ для других видов. Их норы обеспечивают путь для транспортировки растворенного кислорода в воде норы через кислородный осадок стенок норы в окружающий бескислородный осадок, что создает идеальную среду обитания для особых бактерий, циклически вращающих азот. Эти нитратредуцирующие (денитрифицирующие) бактерии быстро поглощают растворенный кислород, поступающий в стенки норы, создавая слой кислородной грязи, который тоньше, чем на поверхности грязи. Это обеспечивает более прямой путь диффузии для экспорта азота (в форме газообразного азота (N 2 )) в смывную приливную воду. [49]
солончаках Микробная в жизнь
Переменная соленость, климат, уровень питательных веществ и анаэробные условия солончаков оказывают сильное селективное давление на населяющие их микроорганизмы. В солончаках микробы играют основную роль в круговороте питательных веществ и биогеохимической переработке. [50] На сегодняшний день не обнаружено существенных изменений микробного сообщества солончаков из-за воздействия человека, но исследования все еще продолжаются. [51] Из-за важной роли микробов в этих средах очень важно понимать различные процессы, происходящие в солончаках, и различных микробных игроков, присутствующих в солончаках. Солончаки обеспечивают среду обитания как для хемо(лито)автотрофов , так и для гетеротрофов и фотоавтотрофов . Эти организмы оказывают разнообразные экологические услуги, такие как восстановление сульфатов , нитрификация , разложение и ризосферы взаимодействие .
солончаках автотрофы в Хемо(лито )
Хемоавтотрофы , также известные как хемолитоавтотрофы, представляют собой организмы, способные создавать собственную энергию за счет использования неорганических молекул и способны процветать в суровых условиях, таких как глубокие морские жерла или солончаки, поскольку они не зависят от внешних источников органического углерода. для их роста и выживания. [52] [53] Некоторые хемоавтотрофные бактериальные микроорганизмы, обнаруженные в солончаках, включают бетапротеобактерии и гаммапротеобактерии , оба класса включают сульфатредуцирующие бактерии (SRB), сероокисляющие бактерии (SOB) и окисляющие аммиак бактерии (AOB), которые играют решающую роль в круговороте питательных веществ и функционировании экосистемы. . [54]
сульфатредуцирующих разнообразие Численность и хемолитоавтотрофов
Бактериальные хемолитоавтотрофы солончаков включают сульфатредуцирующие бактерии. В этих экосистемах до 50% осадочной реминерализации можно отнести на счет сульфатредукции. [55] Доминирующим классом сульфатредуцирующих бактерий в солончаках, как правило, являются дельтапротеобактерии. [55] Некоторыми примерами дельтапротеобактерий, обитающих в солончаках, являются виды родов Desulfobulbus , Desulfuromonas и Desulfovibrio . [55]
Численность и разнообразие хемолитоавтотрофов солончаков во многом определяется видовым составом экосистемы солончаков. [55] [56] Каждый тип солончаковых растений имеет разную продолжительность вегетационного периода , разную скорость фотосинтеза , и все они теряют разное количество органического вещества в океан, что приводит к разным поступлениям углерода в экосистему. Результаты эксперимента, проведенного на солончаке в устье реки Янцзы в Китае, [55] предположили, что как видовое богатство , так и общая численность сообществ сульфатредуцирующих бактерий увеличились, когда было введено новое растение S. alterniflora с более высоким содержанием углерода в экосистеме. [55] Хотя хемолитотрофы производят собственный углерод, они по-прежнему зависят от поступления углерода из солончаков из-за косвенного влияния, которое он оказывает на количество жизнеспособных доноров электронов , таких как восстановленные соединения серы. [57] Концентрация восстановленных соединений серы, а также других возможных доноров электронов органического вещества увеличивается по мере увеличения разложения (другими организмами). будет доступно больше доноров электронов Следовательно, если экосистема содержит больше разлагающегося органического вещества, как в случае с растениями с высокой скоростью фотосинтеза и размножения, бактериям , и, следовательно, возможно большее восстановление сульфатов. В результате увеличивается численность сульфатредуцирующих бактерий. с высоким уровнем фотосинтеза и большим количеством мусора Было обнаружено, что солончаковое растение S. alterniflora выдерживает высокие концентрации серы в почве, которые обычно были бы несколько токсичными для растений. [58]
Численность хемолитоавтотрофов в солончаках также варьируется во времени в результате некоторой зависимости от поступления органического углерода от растений в экосистеме. Поскольку растения растут больше всего в течение лета и обычно начинают терять биомассу осенью на поздней стадии своего развития, наибольшее количество разлагающихся органических веществ приходится на осень. Таким образом, в сезонном плане численность хемолитотрофов в солончаках максимальна осенью. [55]
Почему сульфатредуцирующие бактерии обитают в солончаках? [ редактировать ]
Солончаки — идеальная среда для сульфатредуцирующих бактерий. Сульфатредуцирующие бактерии, как правило, живут в бескислородных условиях, например, в солончаках, поскольку для производства энергии им требуются восстановленные соединения. [58] Поскольку существует высокая скорость седиментации и большое количество органических веществ , условия осадка обычно являются бескислородными. Однако условия на всей территории солончака (над отложениями) не являются полностью бескислородными, а это означает, что организмы, живущие здесь, должны иметь определенный уровень толерантности к кислороду. Многие хемолитоавтотрофы, живущие вне или на поверхности осадков, также обладают этой характеристикой. [58]
Значение сульфатредуцирующих бактерий [ править ]
Сульфатвосстанавливающие бактерии играют важную роль в переработке питательных веществ и снижении уровня загрязнения нитратами . Поскольку люди добавляют непропорциональное количество нитратов в прибрежные воды, солончаки являются одной из экосистем, где загрязнение нитратами остается проблемой. [59] Обогащение нитратами воды увеличивает денитрификацию , а также микробное разложение и первичную продуктивность . [59] Однако сульфатредуцирующие и окисляющие бактерии играют роль в удалении избытка нитратов из воды и предотвращении эвтрофикации . [59] Поскольку сульфатвосстанавливающие бактерии присутствуют в воде и отложениях , восстановленные молекулы серы обычно присутствуют в изобилии. Эти восстановленные сульфаты затем реагируют с избытком нитратов в воде, восстанавливая нитраты и окисляя восстановленную серу. [59] Прогнозируется, что в результате обогащения человека нитратами численность сероокисляющих бактерий, которые также восстанавливают нитраты, увеличится по сравнению с серовосстанавливающими бактериями. [59]
нитрификаторов в солончаках хемолитоавтотрофных и значение Численность
В солончаках хемолитоавтотрофные нитрифицирующие бактерии также часто обнаруживаются , в том числе бетапротеобактерии окислители аммиака, такие как Nitrosomonas и Nitrosospira . [54] Хотя археи, окисляющие аммиак (AOA), оказались более распространенными, чем бактерии, окисляющие аммоний (AOB), в среде солончаков, преимущественно из группы Crenarchaeota , AOB играют решающую роль и в среде солончаков. [60] [61] Увеличение солености болот имеет тенденцию в пользу АОА, в то время как более высокие уровни кислорода и более низкие соотношения углерода и азота благоприятствуют АОА. [62] Эти АОВ играют важную роль в катализе лимитирующей стадии процесса нитрификации за счет использования монооксигеназы аммония (АМО), получаемой из амоА , для превращения аммония (NH4+) в нитрит (NO2-). [63] В частности, в классе Betaproteobacteria Nitrosomonas aestuarii , Nitrosomonas marina и Nitrosospira ureae широко распространены в среде солончаков; аналогично, в классе Gammaproteobacteria Nitrosococcus spp . являются ключевыми АОБ на болотах. [64]
Численность этих хемолитоавтотрофов варьируется в зависимости от градиента солености , присутствующего в солончаках: Nitrosomonas более распространены в регионах с более низкой соленостью или в пресноводных регионах, в то время как Nitrosospira доминируют в средах с более высокой соленостью. обнаружено, что [54] Кроме того, обилие фиксированного азота в этих средах критически влияет на распределение бетапротеобактерий в солончаках: Nitrosomonas чаще встречаются в средах с высоким содержанием N и C, тогда как Nitrosospira более многочисленны в более низких средах. N и C регионы. [54] Кроме того, такие факторы, как температура, pH, чистая первичная продуктивность и области аноксии, могут ограничивать нитрификацию и, таким образом, критически влиять на распределение нитрификаторов. [65]
Роль нитрификации АОБ в солончаках критически связывает аммиак , образующийся в результате минерализации органических соединений азота, с процессом окисления азота. [66] Кроме того, окисление азота важно для последующего удаления нитратов в газообразный азот, катализируемого денитрификаторами , из болотной среды. [66] Следовательно, АОБ играют косвенную роль в удалении азота в атмосферу. [66]
Фотоавтотрофные бактерии [ править ]
Бактериальное фотоавтотрофное сообщество солончаков представлено преимущественно цианобактериями , пурпурными и зелеными серобактериями . [67]
Цианобактерии в солончаках [ править ]
Цианобактерии являются важными фиксаторами азота в солончаках и обеспечивают азотом такие организмы, как диатомовые водоросли и микроводоросли. [68]
Пурпурные бактерии [ править ]
Кислород подавляет фотосинтез пурпурных бактерий, что делает эстуарии благоприятной средой обитания для них из-за низкого содержания кислорода и высокого уровня света, что оптимизирует их фотосинтез. [69] [70] В бескислородных средах, таких как солончаки, многим микробам приходится использовать сульфат в качестве акцептора электронов во время клеточного дыхания вместо кислорода, производя много сероводорода в качестве побочного продукта. [71] [72] Хотя сероводород токсичен для большинства организмов, пурпурным бактериям он необходим для роста и метаболизирует его до сульфата или серы, тем самым позволяя другим организмам обитать в токсичной среде. [69] Пурпурные бактерии можно далее классифицировать как пурпурные серобактерии или пурпурные несерные бактерии. Пурпурные серобактерии более толерантны к сульфидам и сохраняют образуемую ими серу внутриклеточно, тогда как пурпурные несерные бактерии выделяют всю производимую ими серу.
Зеленые бактерии [ править ]
Зеленые серобактерии ( Chlorobiaceae ) — фотоавтотрофные бактерии, которые используют сульфид и тиосульфат для своего роста, образуя при этом сульфат. [73] Они очень приспособлены к фотосинтезу в условиях низкой освещенности с помощью пигментов бактериохлорофиллов a, c, d и e, которые помогают им поглощать длины волн света, которые другие организмы не могут. Сосуществуя с пурпурными бактериями, они часто занимают меньшие глубины, поскольку менее толерантны к кислороду, но более склонны к фотосинтезу.
Ризосферные микробы [ править ]
Грибы [ править ]
Некоторые микоризные грибы , такие как арбускулярная микориза , широко связаны с солончаковыми растениями и могут даже помогать растениям расти в солончаковой почве, богатой тяжелыми металлами, уменьшая их поглощение растением, хотя точный механизм еще предстоит определить. [74]
Бактерии [ править ]
При исследовании рибосомальной ДНК 16S , обнаруженной в устье реки Янцзы , наиболее распространенными бактериями в ризосфере были протеобактерии, такие как бетапротеобактерии , гаммапротеобактерии , дельтапротеобактерии и эпсилонпротеобактерии . [75] Один из таких широко распространенных видов имел риботип, похожий на риботип животного патогена S. marcescens , и может быть полезен для растений, поскольку бактерии могут расщеплять хитин на доступные углерод и азот для использования растениями. [76] Актинобактерии также были обнаружены в ризосфере растений на прибрежных солончаках и помогают растениям расти, помогая им поглощать больше питательных веществ и выделять противомикробные соединения. В Цзянсу, Китай, актинобактерии из подотрядов Pseudonocardineae , Corynebacterineae , Propionibacterineae , Streptomycineae , Micromonosporineae , Streptosporangineae и Micrococcineae были культивированы и выделены из ризосферной почвы. [77]
в солончаках микробного Активность разложения

Еще одним ключевым процессом в микробных солончаках является активность микробного разложения. Круговороту питательных веществ в солончаках активно способствует местное сообщество бактерий и грибов, участвующих в реминерализации органических веществ. Исследования разложения солончаковой травы Spartina alterniflora показали, что колонизация грибов начинает процесс деградации, который затем завершается бактериальным сообществом. [78] Углерод Spartina alterniflora становится доступным для пищевой сети солончаков в основном через эти бактериальные сообщества, которые затем потребляются бактериоядными организмами . [79] Бактерии ответственны за разложение до 88% лигноцеллюлозного материала в солончаках. [79] Однако было обнаружено, что в зимние месяцы грибковые популяции доминируют над бактериальными популяциями. [80]
Грибы, составляющие сообщество разлагателей солончаков, происходят из типа ascomycota , двумя наиболее распространенными видами являются Phaeosphaeria spartinicola и Mycosphaerella sp. штамм 2. [78] Что касается бактерий, класс альфапротеобактерий является наиболее распространенным классом в среде солончаков, участвующих в разложении. [78] Размножение Phaeosphaeria spartinicola осуществляется через аскоспоры , которые высвобождаются, когда растение-хозяин смачивается во время прилива или дождя. [81]
Восстановление и управление [ править ]

Восприятие заливных солончаков как прибрежной «пустоши» с тех пор изменилось, признав, что они являются одной из самых биологически продуктивных сред обитания на Земле, конкурируя с тропическими лесами . Солончаки имеют экологическое значение, обеспечивая среду обитания для местных мигрирующих рыб и выступая в качестве защищенных мест для кормления и нагула. [27] В настоящее время во многих странах они защищены законодательством, чтобы предотвратить потерю этих экологически важных мест обитания. [82] В Соединенных Штатах и Европе им теперь предоставлен высокий уровень защиты в соответствии с Законом о чистой воде и Директивой о средах обитания соответственно. растущий интерес к восстановлению солончаков посредством управляемого отступления Теперь, когда осознано влияние этих мест обитания и их важность, возник или мелиорации земель. Однако многим азиатским странам, таким как Китай, все еще необходимо признать ценность болот. Учитывая постоянно растущее население и интенсивное развитие вдоль побережья, ценность солончаков, как правило, игнорируется, и земли продолжают осваиваться. [7]
Баккер и др. (1997) [83] предлагает два варианта восстановления солончаков. Первый — отказаться от всякого вмешательства человека и предоставить солончакам возможность завершить свое естественное развитие. Подобные проекты восстановления часто оказываются безуспешными, поскольку растительность имеет тенденцию изо всех сил пытаться вернуться к своей первоначальной структуре, а естественные циклы приливов и отливов смещаются из-за изменений на суше. Второй вариант, предложенный Bakker et al. (1997) [83] заключается в восстановлении разрушенной среды обитания в ее естественном состоянии либо на исходном участке, либо в качестве замены на другом участке. В естественных условиях восстановление может занять 2–10 лет или даже дольше в зависимости от характера и степени нарушения, а также относительной зрелости болота. [82] Болота на первых стадиях развития восстанавливаются быстрее, чем зрелые. [82] поскольку они часто первыми колонизируют землю. Важно отметить, что восстановление часто можно ускорить за счет пересадки местной растительности.

Этот последний подход часто является наиболее практикуемым и, как правило, более успешным, чем естественное восстановление территории самостоятельно. Солончаки в штате Коннектикут в США уже давно являются территорией, которую невозможно засыпать и выкопать. В 1969 году был принят Закон о приливных водно-болотных угодьях, который положил конец этой практике. [37] но, несмотря на принятие закона, система все еще деградировала из-за изменений приливных потоков. Одна из областей Коннектикута — болота на острове Барн. Эти болота были обвалованы дамбами, а затем в 1946–1966 годах засыпаны солью и солоноватыми болотами. [37] В результате болото перешло в пресноводное состояние, и в нем стали доминировать инвазивные виды P. australis , Typha angustifolia и T. latifolia , которые имеют мало экологической связи с этой территорией. [37]
К 1980 году была запущена программа восстановления, которая действует уже более 20 лет. [37] Эта программа направлена на воссоединение болот путем возвращения приливных потоков, а также экологических функций и характеристик болот в исходное состояние. В случае с островом Барн было начато сокращение количества инвазивных видов, восстанавливая приливно-болотную растительность вместе с видами животных, такими как рыбы и насекомые. Этот пример показывает, что для эффективного восстановления систем солончаков необходимо значительное время и усилия. Сроки восстановления солончаков зависят от стадии развития болота, типа и степени нарушения, географического положения, а также факторов экологического и физиологического стресса для связанной с болотами флоры и фауны.
Хотя на восстановление солончаков во всем мире было приложено много усилий, необходимы дальнейшие исследования. Существует множество неудач и проблем, связанных с восстановлением болот, которые требуют тщательного долгосрочного мониторинга. Необходимо понимать и отслеживать информацию обо всех компонентах экосистемы солончаков, начиная с отложений, питательных веществ и приливных влияний и заканчивая моделями поведения и толерантностью видов как флоры, так и фауны. [82] Как только будет достигнуто лучшее понимание этих процессов, и не только на местном, но и в глобальном масштабе, тогда можно будет предпринять более разумные и практические усилия по управлению и восстановлению, чтобы сохранить эти ценные болота и вернуть их в исходное состояние.
Несмотря на то, что люди проживают вдоль береговой линии, всегда будет существовать возможность антропогенного вмешательства, несмотря на количество усилий по восстановлению, которые мы планируем реализовать. Дноуглубительные работы, трубопроводы для морских нефтяных ресурсов, строительство автомагистралей, случайные разливы токсичных веществ или просто небрежность — вот примеры, которые в течение некоторого времени и в будущем будут основными факторами, влияющими на деградацию солончаков. [82]

Помимо восстановления и управления системами солончаков на основе научных принципов, следует использовать возможность информировать общественность об их биологической важности и назначении в качестве естественного буфера для защиты от наводнений. [27] Поскольку солончаки часто расположены рядом с городскими районами, они, вероятно, будут принимать больше посетителей, чем отдаленные водно-болотные угодья . Физически видя болото, люди с большей вероятностью обращают внимание и лучше осведомлены об окружающей среде. Пример участия общественности произошел в Государственном морском заповеднике Фамоза-Слау в Сан-Диего , где группа «друзей» работала более десяти лет, пытаясь предотвратить освоение этого района. [84] В конце концов, участок площадью 5 гектаров (12 акров) был куплен городом, и группа вместе работала над восстановлением территории. Проект включал удаление инвазивных видов и засадку местными видами, а также публичные беседы с другими местными жителями, частые прогулки с птицами и мероприятия по уборке. [84]
Методы исследования [ править ]
Существует широкий спектр и сочетание методологий, используемых для понимания гидрологической динамики солончаков и их способности улавливать и накапливать отложения. Ловушки для отложений часто используются для измерения скорости прироста поверхности болота, когда требуется кратковременное развертывание (например, менее одного месяца). Эти круглые ловушки состоят из предварительно взвешенных фильтров, которые прикрепляются к поверхности болота, затем высушиваются в лаборатории и повторно взвешиваются для определения общего количества отложившихся осадков. [22] [23]
Для долгосрочных исследований (например, более одного года) исследователи могут предпочесть измерять прирост отложений с помощью графиков маркерных горизонтов . Маркерные горизонты состоят из минерала, такого как полевой шпат , который заглублен на известную глубину в субстрате водно-болотных угодий , чтобы регистрировать увеличение количества вышележащего субстрата в течение длительных периодов времени. [25] Чтобы измерить количество отложений, взвешенных в толще воды, вручную или автоматически пробы приливной воды можно пролить через предварительно взвешенные фильтры в лаборатории, а затем высушить, чтобы определить количество осадка на объем воды. [23]
Другой метод оценки концентрации взвешенных отложений заключается в измерении мутности воды с помощью оптических датчиков обратного рассеяния, которые можно калибровать по пробам воды, содержащим известную концентрацию взвешенных отложений, чтобы установить регрессионную зависимость между ними. [20] Высота поверхности болота может быть измерена с помощью стадионной рейки и транзита. [23] электронный теодолит , [22] Кинематическая глобальная система позиционирования в реальном времени, [20] лазерный уровень [25] или электронный дальномер ( тахеометр ). Гидрологическая динамика включает глубину воды, автоматически измеряемую датчиком давления . [22] [23] [25] или с помощью отмеченного деревянного кола, [21] и скорость воды, часто с использованием электромагнитных измерителей тока. [21] [23]
См. также [ править ]
- Байю
- Пляжный луг
- План действий по сохранению биоразнообразия
- Синий углерод
- Водоем
- Сад
- Солоноватое болото
- Калифорнийское прибрежное солончак
- Фен
- Грейт-Марш
- Халлиген (острова соленых лугов)
- Галофит
- Высокое болото
- Лагуна
- Низкое болото
- Мангровое болото
- Морская прибрежная экосистема
- Марш
- Месопотамские болота
- Грязь
- Ocean Data Viewer : содержит набор данных о солончаках по всему миру.
- Выезд
- Растительное сообщество
- Солончак
- Вымирание солончаков
- морская трава
- Морское болото
- Приливное болото
- Пресноводное болото
- водно-болотное угодье
Ссылки [ править ]
- ↑ Перейти обратно: Перейти обратно: а б с д и ж Адам, Пол (1990). Экология солончаков . Нью-Йорк: Издательство Кембриджского университета. ISBN 0-521-24508-7 . OCLC 20217629 .
- ↑ Перейти обратно: Перейти обратно: а б с д и ж Вудрофф, компакт-диск (2002). Берега: форма, процесс и эволюция . Нью-Йорк: Издательство Кембриджского университета. ISBN 0-521-81254-2 . OCLC 48795910 .
- ^ Симас, Т; Нуньес, Япония; Феррейра, JG (март 2001 г.). «Влияние глобального изменения климата на прибрежные солончаки». Экологическое моделирование . 139 (1): 1–15. Бибкод : 2001EcMod.139....1S . дои : 10.1016/S0304-3800(01)00226-5 .
- ^ Агентство по охране окружающей среды (10 апреля 2014 г.). «Изменение климата в прибрежной среде» . Проверено 30 октября 2023 г.
- ^ Аллен, JRL, Пай, К. (1992). Солончаки: морфодинамика , сохранение и инженерное значение. Издательство Кембриджского университета. Кембридж, Великобритания.
- ↑ Перейти обратно: Перейти обратно: а б с Чепмен, виджей (1974). Солончаки и соляные пустыни мира. Филлис Клер Чепмен, Германия.
- ↑ Перейти обратно: Перейти обратно: а б с д и ж г час Бромберг-Гедан К., Силлиман Б.Р. и Бертнесс, доктор медицинских наук (2009). «Века антропогенных изменений в экосистемах солончаков», Annual Review of Marine Science , 1: 117–141.
- ↑ Перейти обратно: Перейти обратно: а б Те Ара - Энциклопедия Новой Зеландии (2005–2010 гг.). «Растения лимана» . Проверено 15 марта 2010 г.
- ^ Вернберг, Ф.Дж. 1993. Солончаковые процессы: обзор. Экологическая токсикология и химия 12: 2167–2195.
- ↑ Перейти обратно: Перейти обратно: а б Скотт, Д.Б., Дж. Фрейл-Готье и П.Дж. Муди. 2014. Прибрежные водно-болотные угодья мира: геология, экология, распространение и применение . Издательство Кембриджского университета, Нью-Йорк
- ↑ Перейти обратно: Перейти обратно: а б Макоуэн, Крис; Уэтердон, Лорен; Бохове, Ян-Виллем; Салливан, Эмма; Блит, Саймон; Зоклер, Кристоф; Стэнвелл-Смит, Дэймон; Кингстон, Наоми; Мартин, Корин (21 марта 2017 г.). «Глобальная карта солончаков» . Журнал данных по биоразнообразию . 5 (5): е11764. дои : 10.3897/bdj.5.e11764 . ISSN 1314-2828 . ПМК 5515097 . ПМИД 28765720 .
- ↑ Перейти обратно: Перейти обратно: а б с Мюррей, Николас Дж.; Уортингтон, Томас А.; Бантинг, Пит; Дуче, Стефани; Хаггер, Валери; Лавлок, Кэтрин Э.; Лукас, Ричард; Сондерс, Меган И.; Шивс, Маркус; Сполдинг, Марк; Уолтем, Натан Дж.; Лайонс, Митчелл Б. (13 мая 2022 г.). «Картирование с высоким разрешением потерь и прироста приливно-отливных водно-болотных угодий Земли» (PDF) . Наука . 376 (6594): 744–749. Бибкод : 2022Sci...376..744M . дои : 10.1126/science.abm9583 . ПМИД 35549414 . S2CID 248749118 .
- ↑ Перейти обратно: Перейти обратно: а б с д и Петик, Дж. (1984). Введение в прибрежную геоморфологию . Эдвард Арнольд, Лондон.
- ↑ Перейти обратно: Перейти обратно: а б с Бурман Л., Хазелден Дж. и Бурман М. (2002). «Новые солончаки вместо старых – создание солончаков и управление ими». Меняющееся побережье , EUROCAST/EUCC, Прибрежная зона EUROCOAST 2002: Порту, Португалия; 35–45.
- ^ Гинзбург, Р.Н., и Лоуэнстам, HA (1958). «Влияние морских донных сообществ на условия отложения осадков». Геологический журнал , 66: (3), 310–318.
- ^ Аспден, Р.Дж., Варди, С. и Патерсон, DM (2004). Микробная экология солончаков: микробы, донные маты и движение наносов. В Фагерацци С., Марани М. и Блюм Л.К. (редакторы), Экогеоморфология приливных болот (стр. 115–136). Американский геофизический союз, Вашингтон, округ Колумбия.
- ↑ Перейти обратно: Перейти обратно: а б с Берд, Э. (2008). Прибрежная геоморфология: введение . John Wiley & Sons Ltd, Западный Суссекс, Англия.
- ↑ Перейти обратно: Перейти обратно: а б с д и Бертнесс, доктор медицинских наук, Эванчук, П.Дж., Силлиман, Б.Р. (2002). «Антропогенная модификация солончаковых ландшафтов Новой Англии». Труды Национальной академии наук 99 (3): 1395–1398.
- ↑ Перейти обратно: Перейти обратно: а б Рэнд, Т.А. (2000). Распространение семян, пригодность среды обитания и распределение галофитов по приливному градиенту солончаков. Журнал экологии 88 (4): 608–621.
- ↑ Перейти обратно: Перейти обратно: а б с д и ж Ли, Х. и Ян, С.Л. (2000). «Захватывающее воздействие приливной болотной растительности на взвешенные отложения, дельта Янцзы». Журнал прибрежных исследований , 25: (4), 915–924.
- ↑ Перейти обратно: Перейти обратно: а б с д и ж Ши З., Гамильтон Л.Дж. и Волански Э. (2000). «Прикроватные течения и перенос взвешенных наносов в пологах солончаков». Журнал прибрежных исследований , 16: (3), 908–914.
- ↑ Перейти обратно: Перейти обратно: а б с д и ж г час Рид, Д.Д., Спенсер, Т., Мюррей, А.Л., Френч, Дж.Р. и Леонард, Л. (1999). «Отложение наносов на поверхности болот и роль приливных ручьев: последствия для созданных и управляемых прибрежных болот». Журнал охраны побережья , 5: (1), 81–90.
- ↑ Перейти обратно: Перейти обратно: а б с д и ж г Вуд Н. и Хайн AC (2007). «Пространственные тенденции отложения болотных отложений в системе микроприливных ручьев, залив Вакасасса, Флорида». Журнал прибрежных исследований , 23: (4), 823–833.
- ^ Чен, Си; Торрес, Раймонд (21 марта 2012 г.). «Влияние геоморфологии на распределение содержания металлов в отложениях солончаков». Эстуарии и побережья . 35 (4): 1018–1027. Бибкод : 2012EstCo..35.1018C . дои : 10.1007/s12237-012-9494-y . ISSN 1559-2723 . S2CID 129721804 .
- ↑ Перейти обратно: Перейти обратно: а б с д Кахун, Д.Р., Уайт, Д.А. и Линч, Дж.К. (2011). «Динамика заполнения отложений и формирования водно-болотных угодий в активной трещине дельты реки Миссисипи». Геоморфология , 131: 57–68.
- ^ Хинде, HP (1954). «Вертикальное распределение солончаковых фанерогов в зависимости от уровня приливов». Экологические монографии 24 (2): 209–225.
- ↑ Перейти обратно: Перейти обратно: а б с Кинг, С.Э., Лестер, Дж.Н. (1995). «Ценность солончаков как защита от моря». Бюллетень о загрязнении морской среды 30(3): 180–189.
- ↑ Перейти обратно: Перейти обратно: а б с Лонг, С.П. и Мейсон, К.Ф. (1983). Экология солончаков . Blackie & Son Ltd, Глазго.
- ^ Андресен, Х.; Баккер, JP; Бронгерс, М.; Хайдеманн, Б.; Ирмлер, У. (1990). «Долгосрочные изменения солончаковых сообществ в результате выпаса скота» . Растительность . 89 (2): 137–148. дои : 10.1007/BF00032166 . ISSN 0042-3106 . JSTOR 20038672 . S2CID 20754802 .
- ^ Френч, младший и Бернингем, Х. (2003). «Приливное отложение болот и повышение уровня моря: взгляд на устье юго-восточной Англии», Proceedings Coastal Sediments , 1–13.
- ^ Ангус, Г. и Уолтерс, М. (2008). «Естественное восстановление солончаков на ранее освоенных землях». Прикладная наука о растительности , 11: 335–344.
- ↑ Перейти обратно: Перейти обратно: а б Ранвелл, DS (1972). Экология солончаков и песчаных дюн . Chapman and Hall Ltd, Лондон.
- ^ Кирван, М.Л., Мюррей, А.Б., Доннелли, Дж.П. и Корбетт, Д. (2011). «Быстрое расширение водно-болотных угодий во время заселения европейцев и его влияние на выживание болот при современных скоростях доставки наносов». Геологическое общество Америки , 39: (5), 507–510.
- ^ Юпп, К. (2007). Создание физической и биологической основы для восстановления и управления солончаками в устье Эйвон-Хиткот. Крайстчерч, Кентерберийский университет.
- ^ Лангис, Р., Залейко, М., Зедлер, Дж.Б. (1991). «Оценка содержания азота в искусственных и естественных солончаках залива Сан-Диего». Экологические приложения 1 (1): 40–51.
- ^ Чемберс, Р.М., Мейерсон, Л.А., Солтонстолл, К. (1999). «Распространение Phragmites australis на приливные водно-болотные угодья Северной Америки». Водная ботаника 64: 261–273.
- ↑ Перейти обратно: Перейти обратно: а б с д и Уоррен, Р.С., Фелл, П.Е., Розса, Р., Броули, А.Х., Орстед, А.К., Олсон, Э.Т., Свами, В., Ниринг, Вашингтон (2002). «Восстановление солончаков в Коннектикуте: 20 лет науки и управления». Реставрационная экология 10 (3): 497–513.
- ^ Валиэла, Иван; Льорет, Хавьер; Бойер, Тайнан; Майнер, Саймон; Ремсен, Дэвид; Элмстром, Элизабет; Когсуэлл, Шарлотта; Роберт Тилер, Э. (ноябрь 2018 г.). «Переходные прибрежные ландшафты: повышение уровня моря угрожает солончакам». Наука об общей окружающей среде . 640–641: 1148–1156. Бибкод : 2018ScTEn.640.1148V . doi : 10.1016/j.scitotenv.2018.05.235 . hdl : 1912/10488 . ПМИД 30021280 . S2CID 51703514 .
- ^ Ганджу, Нил К.; Дефне, Зафер; Кирван, Мэтью Л.; Фагерацци, Серджио; Д'Альпаос, Андреа; Карниелло, Лука (23 января 2017 г.). «Пространственно-интегративные показатели выявляют скрытую уязвимость микроприливных солончаков» . Природные коммуникации . 8 : 14156. Бибкод : 2017NatCo...814156G . дои : 10.1038/ncomms14156 . ISSN 2041-1723 . ПМК 5264011 . ПМИД 28112167 .
- ↑ Перейти обратно: Перейти обратно: а б с д Бест, Ю. СН; Ван дер Веген, М.; Дейкстра, Дж.; Виллемсен, PWJM; Борсье, БВ; Рулвинк, Дано Дж.А. (2018). «Переживают ли солончаки повышение уровня моря? Моделирование волнового воздействия, морфодинамики и динамики растительности» . Экологическое моделирование и программное обеспечение . 109 : 152–166. Бибкод : 2018EnvMS.109..152B . дои : 10.1016/j.envsoft.2018.08.004 .
- ^ Баума, Ти Джей; Ван Бельзен, Дж.; Балке, Т.; ван Дален, Дж.; Клаассен, П.; Хартог, AM; Каллаган, ДП; Ху, З.; Стив, MJF; Теммерман, С.; Герман, PMJ (2016). «Краткосрочная динамика илистых отмелей стимулирует долгосрочную циклическую динамику солончаков» . Лимнология и океанография . 61 (2016): 2261–2275. Бибкод : 2016LimOc..61.2261B . дои : 10.1002/lno.10374 . hdl : 10067/1384590151162165141 .
- ↑ Перейти обратно: Перейти обратно: а б Ли, Жуньсян; Ю, Цянь; Ван, Юнвэй; Ван, Чжэн Бин; Гао, Шу; Флемминг, Бург (2018). «Взаимосвязь между продолжительностью наводнения и ростом Spartina alterniflora вдоль побережья Цзянсу, Китай» . Устьевые, прибрежные и шельфовые науки . 213 : 305–313. Бибкод : 2018ECSS..213..305L . дои : 10.1016/j.ecss.2018.08.027 . S2CID 135052098 .
- ^ Шуерх, М.; Спенсер, Т.; Теммерман, С.; Кирван, МЛ; Вольф, К.; Линке, Д.; МакОуэн, CJ; Пикеринг, доктор медицины; Риф, Р.; Вафеидис, АТ; Хинкель, Дж.; Николла, Р.Дж.; Браун, С. (2018). «Будущая реакция прибрежных водно-болотных угодий мира на повышение уровня моря» (PDF) . Природа . 561 (7722): 231–247. Бибкод : 2018Natur.561..231S . дои : 10.1038/s41586-018-0476-5 . ПМИД 30209368 . S2CID 52198604 .
- ^ Шиле, Л.М.; Каллауэй, Джей Си; Моррис, Джей Ти; Штральберг, Д.; Паркер, Вирджиния; Келли, М. (2014). «Оценка роли растительности, отложений и горной среды обитания в устойчивости болот» . ПЛОС ОДИН . 9 (2): е88760. дои : 10.1371/journal.pone.0088760 . ПМЦ 3923833 . ПМИД 24551156 .
- ^ «Восстановление среды обитания Род-Айленда». Архивировано 8 октября 2022 года в Wayback Machine , Университет Род-Айленда:
- ^ Альберти Дж., Себриан Дж., Касарьего А.М., Канепучча А., Эскапа М. и Ирибарн О. (2011). «Влияние обогащения питательными веществами и травоядности крабов на продуктивность солончаков на юго-западе Атлантического океана». Журнал экспериментальной морской биологии и экологии , 405: 99–104.
- ^ Холдридж, К., Бертнесс, доктор медицинских наук и Альтьери, А.Х. (2008). «Роль травоядных крабов в вымирании солончаков Новой Англии». Биология сохранения , 23: (3), 672–679.
- ^ Смит, С.М. и Тиррелл, MC (2012). «Влияние грязевых крабов-скрипачей (Uca pugnax) на пополнение сеянцев галофитов в районах отмирания солончаков Кейп-Код» (Массачусетс, США). Экологические исследования , 27: 233–237.
- ^ Вопель К. и Хэнкок Н. (2005). «Морские экосистемы: больше, чем просто крабовая нора» . Вода и атмосфера , 13: (3), 18–19.
- ^ Яо, Чжиюань; Ду, Шиконг; Лян, Чуньлин; Чжао, Юэджи; Дини-Андреоте, Франциско; Ван, Кай; Чжан, Демин (15 марта 2019 г.). Лю, Шуан-Цзян (ред.). «Сборка бактериального сообщества в типичном устьевом болоте с множественными градиентами окружающей среды» . Прикладная и экологическая микробиология . 85 (6). Бибкод : 2019ApEnM..85E2602Y . дои : 10.1128/АЕМ.02602-18 . ISSN 0099-2240 . ПМК 6414364 . ПМИД 30635381 .
- ^ Боуэн, Дженнифер Л.; Крамп, Байрон С; Диган, Линда А; Хобби, Джон Э. (1 августа 2009 г.). «Бактерии отложений солончаков: их распространение и реакция на внешние питательные вещества» . Журнал ISME . 3 (8): 924–934. Бибкод : 2009ISMEJ...3..924B . дои : 10.1038/ismej.2009.44 . ISSN 1751-7362 . ПМИД 19421233 .
- ^ «5.1B: Хемоавтотрофы и хемогетротрофы» . Свободные тексты по биологии . 8 мая 2017 года . Проверено 4 апреля 2024 г.
- ^ Накагава, Сатоши; Такай, Кен (июль 2008 г.). «Хемоавтотрофы глубоководных жерл: разнообразие, биохимия и экологическое значение: хемоавтотрофия в глубоководных жерлах» . ФЭМС Микробиология Экология . 65 (1): 1–14. дои : 10.1111/j.1574-6941.2008.00502.x . ПМИД 18503548 .
- ↑ Перейти обратно: Перейти обратно: а б с д Бернхард, Энн Э.; Шеффер, Роберта; Гиблин, Энн Э.; Мартон, Джон М.; Робертс, Брайан Дж. (2016). «Динамика численности и состав сообществ окислителей аммиака в солончаках после разлива нефти на глубоководном горизонте» . Границы микробиологии . 7 : 854. дои : 10.3389/fmicb.2016.00854 . ISSN 1664-302X . ПМЦ 4899434 . ПМИД 27375576 .
- ↑ Перейти обратно: Перейти обратно: а б с д и ж г Не, Мин; Ван, Мэн; Ли, Бо (1 декабря 2009 г.). «Влияние вторжения Spartina alterniflora в солончаки на сульфатвосстанавливающие бактерии в устье реки Янцзы, Китай» . Экологическая инженерия . 35 (12): 1804–1808. Бибкод : 2009EcEng..35.1804N . дои : 10.1016/j.ecoleng.2009.08.002 . ISSN 0925-8574 .
- ^ Боуэн, Дженнифер Л.; Крамп, Байрон С; Диган, Линда А; Хобби, Джон Э. (7 мая 2009 г.). «Бактерии отложений солончаков: их распространение и реакция на внешние питательные вещества» . Журнал ISME . 3 (8): 924–934. Бибкод : 2009ISMEJ...3..924B . дои : 10.1038/ismej.2009.44 . ISSN 1751-7362 . ПМИД 19421233 .
- ^ Квон, Ман Джэ; О'Локлин, Эдвард Дж.; Боянов Максим И.; Брюлк, Дженнифер М.; Джонстон, Эрик Р.; Кемнер, Кеннет М.; Антонопулос, Дионисий А. (22 января 2016 г.). «Влияние органических доноров электронов углерода на развитие микробного сообщества в железо- и сульфатредуцирующих условиях» . ПЛОС ОДИН . 11 (1): e0146689. Бибкод : 2016PLoSO..1146689K . дои : 10.1371/journal.pone.0146689 . ISSN 1932-6203 . ПМЦ 4723079 . ПМИД 26800443 .
- ↑ Перейти обратно: Перейти обратно: а б с Чжэн, Ю; Бу, Най-Шун; Лонг, Си-Эн; Сунь, Цзин; Он, Чи-Цюань; Лю, Сяо-Янь; Цуй, Джун; Лю, Дун-Сю; Чен, Сюэ-Пин (1 февраля 2017 г.). «Сульфатредуктор и окислитель серы по-разному реагируют на вторжение Spartina alterniflora в устьевые солончаки Китая» . Экологическая инженерия . 99 : 182–190. Бибкод : 2017EcEng..99..182Z . дои : 10.1016/j.ecoleng.2016.11.031 . ISSN 0925-8574 .
- ↑ Перейти обратно: Перейти обратно: а б с д и Винейс, Джозеф Х; Булсеко, Эшли Н; Боуэн, Дженнифер Л. (2023). «Микробные хемолитоавтотрофы в изобилии обитают в отложениях солончаков после длительного экспериментального обогащения нитратами» . Письма FEMS по микробиологии . 370 . дои : 10.1093/femsle/fnad082 . ISSN 1574-6968 . ПМИД 37541957 .
- ^ Нельсон, Кейтлин А.; Мойн, Николь С.; Бернхард, Энн Э. (15 июня 2009 г.). «Археальное разнообразие и распространенность Crenarchaeota в отложениях солончаков» . Прикладная и экологическая микробиология . 75 (12): 4211–4215. Бибкод : 2009ApEnM..75.4211N . дои : 10.1128/АЕМ.00201-09 . ISSN 0099-2240 . ПМЦ 2698360 . ПМИД 19395565 .
- ^ Мойн, Николь С.; Нельсон, Кейтлин А.; Буш, Александр; Бернхард, Энн Э. (декабрь 2009 г.). «Распространение и разнообразие архейных и бактериальных окислителей аммиака в отложениях солончаков» . Прикладная и экологическая микробиология . 75 (23): 7461–7468. Бибкод : 2009ApEnM..75.7461M . дои : 10.1128/АЕМ.01001-09 . ISSN 0099-2240 . ПМК 2786404 . ПМИД 19801456 .
- ^ Чжан, И; Цай, Цзуконг; Чжан, Цзиньбо; Мюллер, Кристоф (10 июля 2020 г.). «Соотношение C:N не является надежным показателем производства N2O в кислых почвах после 30-дневного искусственного воздействия» . Наука об общей окружающей среде . 725 : 138427. Бибкод : 2020ScTEn.72538427Z . doi : 10.1016/j.scitotenv.2020.138427 . ISSN 0048-9697 . ПМИД 32464751 .
- ^ Доллхопф, Шерри Л.; Хён, Чон Хо; Смит, Эйприл К.; Адамс, Гарольд Дж.; О'Брайен, Шон; Костка, Джоэл Э. (январь 2005 г.). «Количественное определение аммиачно-окисляющих бактерий и факторов, контролирующих нитрификацию в солончаковых отложениях» . Прикладная и экологическая микробиология . 71 (1): 240–246. Бибкод : 2005ApEnM..71..240D . дои : 10.1128/АЕМ.71.1.240-246.2005 . ISSN 0099-2240 . ПМК 544235 . ПМИД 15640193 .
- ^ Теббе, Деннис Александр; Гейзер, Симона; Вемхойер, Бернд; Дэниел, Рольф; Шефер, Хендрик; Энгелен, Берт (май 2022 г.). «Сезонная и зональная смена бактериальных сообществ в отложениях солончаков Северного моря» . Микроорганизмы . 10 (5): 859. doi : 10.3390/microorganisms10050859 . ISSN 2076-2607 . ПМК 9146408 . ПМИД 35630305 .
- ^ Пэн, Сюэфэн; Яндо, Эрик; Хильдебранд, Эрика; Дуайер, Кортни; Кирни, Энн; Васьега, Алекс; Валиэла, Иван; Бернхард, Энн (2013). «Дифференциальная реакция архей и бактерий, окисляющих аммиак, на длительное внесение удобрений в солончаке Новой Англии» . Границы микробиологии . 3 : 445. дои : 10.3389/fmicb.2012.00445 . ISSN 1664-302X . ПМЦ 3551258 . ПМИД 23346081 .
- ↑ Перейти обратно: Перейти обратно: а б с Боуэн, Дженнифер Л.; Спивак, Аманда С.; Бернхард, Энн Э.; Фулвейлер, Робинсон В.; Гиблин, Энн Э. (октябрь 2023 г.). «Круговорот азота в солончаках: место, где земля встречается с морем» . Тенденции в микробиологии . дои : 10.1016/j.tim.2023.09.010 . ISSN 0966-842X . ПМИД 37827901 .
- ^ Фрэнкс, Джонатан; Штольц, Джон Ф. (1 октября 2009 г.). «Сообщества плоских слоистых микробных матов» . Обзоры наук о Земле . Микробные маты в летописи ископаемой жизни Земли: геобиология. 96 (3): 163–172. Бибкод : 2009ESRv...96..163F . doi : 10.1016/j.earscirev.2008.10.004 . ISSN 0012-8252 .
- ^ Каррин, Кэролайн А.; Левин, Лиза А.; Талли, Тереза С.; Миченер, Роберт; Тэлли, Дрю (сентябрь 2011 г.). «Роль цианобактерий в пищевых сетях солончаков Южной Калифорнии» . Морская экология . 32 (3): 346–363. Бибкод : 2011MarEc..32..346C . дои : 10.1111/j.1439-0485.2011.00476.x . ISSN 0173-9565 .
- ↑ Перейти обратно: Перейти обратно: а б Мэдиган, Майкл Т.; Юнг, Дебора О. (2009), Хантер, К. Нил; Далдал, Февзи; Турнауэр, Мэрион К.; Битти, Дж. Томас (ред.), «Обзор пурпурных бактерий: систематика, физиология и среда обитания» , «Пурпурные фототрофные бактерии» , Дордрехт: Springer Нидерланды, стр. 1–15, номер документа : 10.1007/978-1-4020. -8815-5_1 , ISBN 978-1-4020-8815-5 , получено 7 апреля 2024 г.
- ^ Министерство торговли США, Национальное управление океанических и атмосферных исследований. «Что такое солончак?» . Oceanservice.noaa.gov . Проверено 7 апреля 2024 г.
- ^ Бродерсен, Каспер Эльгетти; Треватан-Тэкетт, Стейси М.; Нильсен, Дэниел А.; Коннолли, Род М.; Лавлок, Кэтрин Э.; Этвуд, Триша Б.; Макриди, Питер I. (2019). «Потребление кислорода и восстановление сульфатов в прибрежных средах обитания с растительностью: последствия физического нарушения» . Границы морской науки . 6 . дои : 10.3389/fmars.2019.00014 . hdl : 10536/DRO/DU:30117171 . ISSN 2296-7745 .
- ^ Автор (21 марта 2022 г.). «Солончаки — красивые, вонючие и неудобные места для работы» . Офис устойчивого развития . Проверено 7 апреля 2024 г.
{{cite web}}:|last=имеет общее имя ( справка ) - ^ Уитмен, Уильям Б., изд. (14 сентября 2015 г.). Руководство Берги по систематике архей и бактерий (1-е изд.). Уайли. дои : 10.1002/9781118960608.gbm00378 . ISBN 978-1-118-96060-8 .
- ^ Карраско, Л.; Каравака, Ф.; Альварес-Рогель, Дж.; Ролдан, А. (1 июня 2006 г.). «Микробные процессы в ризосферной почве загрязненного тяжелыми металлами средиземноморского солончака: облегчающая роль АМ-грибов» . Хемосфера . 64 (1): 104–111. Бибкод : 2006Chmsp..64..104C . doi : 10.1016/j.chemSphere.2005.11.038 . ISSN 0045-6535 . ПМИД 16403557 .
- ^ Ван, Мэн; Чен, Цзя-Куан; Ли, Бо (1 октября 2007 г.). «Характеристика структуры и разнообразия бактериального сообщества в ризосферных почвах трех растений в быстро меняющихся солончаках с использованием 16S рДНК» . Педосфера . 17 (5): 545–556. Бибкод : 2007Pedos..17..545W . дои : 10.1016/S1002-0160(07)60065-4 . ISSN 1002-0160 .
- ^ Шобаде, Сэмюэл О.; Заботина, Ольга Александровна; Нильсен-Гамильтон, Марит (2024). «Хитиназы, ассоциированные с корнями растений: структура и функции» . Границы в науке о растениях . 15 . дои : 10.3389/fpls.2024.1344142 . ISSN 1664-462X . ПМЦ 10867124 . ПМИД 38362446 .
- ^ Гун, Юань; Бай, Хуан-Луан; Ян, Хуан-Тин; Чжан, Вэнь-Ди; Сюн, Ю-Вэй; Дин, Пэн; Цинь, Шэн (1 сентября 2018 г.). «Филогенетическое разнообразие и исследование свойств актинобактерий, способствующих росту растений, в ризосферах растений прибрежных солончаков из Цзянсу, Китай» . Систематическая и прикладная микробиология . 41 (5): 516–527. дои : 10.1016/j.syapm.2018.06.003 . ISSN 0723-2020 . ПМИД 29934111 .
- ↑ Перейти обратно: Перейти обратно: а б с Бьюкен, Элисон; Ньюэлл, Стивен Ю.; Батлер, Мелисса; Бирс, Эрин Дж.; Холлибо, Джеймс Т.; Моран, Мэри Энн (ноябрь 2003 г.). «Динамика бактериальных и грибковых сообществ на гниющей солончаковой траве» . Прикладная и экологическая микробиология . 69 (11): 6676–6687. Бибкод : 2003ApEnM..69.6676B . дои : 10.1128/АЕМ.69.11.6676-6687.2003 . ISSN 0099-2240 . ПМК 262310 . ПМИД 14602628 .
- ↑ Перейти обратно: Перейти обратно: а б Беннер, Рональд; Моран, Мэри Энн; Ходсон, Роберт Э. (январь 1986 г.). «Биогеохимический круговорот лигноцеллюлозного углерода в морских и пресноводных экосистемах: относительный вклад прокариотов и эукариотов1» . Лимнология и океанография . 31 (1): 89–100. Бибкод : 1986LimOc..31...89B . дои : 10.4319/lo.1986.31.1.0089 . ISSN 0024-3590 .
- ^ Ньюэлл, Стивен Ю.; Портер, Дэвид (2000), Вайнштейн, Майкл П.; Кригер, Дэниел А. (ред.), «Вторичная микробная продукция из побегов солончаковой травы, ее известная и потенциальная судьба» , «Концепции и противоречия в экологии приливных болот » , Дордрехт: Springer Нидерланды, стр. 159–185, doi : 10.1007/0-306-47534-0_9 , ISBN 978-0-306-47534-4 , получено 4 апреля 2024 г.
- ^ Ньюэлл, Стивен Ю.; Васовский, Дженнифер (1995). «Половая продуктивность и весеннее внутриболотное распространение ключевого солончакового микробного вторичного продуцента» . Эстуарии . 18 (1): 241–249. дои : 10.2307/1352634 . ISSN 0160-8347 . JSTOR 1352634 .
- ↑ Перейти обратно: Перейти обратно: а б с д и Брум, Ю.В., Сенека, Э.Д., Вудхаус, WW (1988). «Восстановление приливного болота». Водная ботаника 32: 1–22.
- ↑ Перейти обратно: Перейти обратно: а б Баккер, Дж.П., Эсселинк, П., Ван Дер Валь, Р., Дейкема, К.С. (1997). «Варианты восстановления и управления прибрежными солончаками в Европе», Урбанска, К.М., Уэбб, Н.Р., Эдвардс, П.Дж. (ред.), « Экология восстановления и устойчивое развитие» . Издательство Кембриджского университета, Великобритания. п. 286-322.
- ↑ Перейти обратно: Перейти обратно: а б Каллауэй, Дж. К., Зедлер, Дж. Б. (2004). «Восстановление городских солончаков: уроки южной Калифорнии». Городские экосистемы 7: 107–124.
Дальнейшее чтение [ править ]
- Гедан, Керин Б.; Альтьери, Эндрю Х.; Бертнесс, Марк Д. (2011), «Неопределенное будущее солончаков Новой Англии», Marine Ecology Progress Series , 434 : 229–238, Бибкод : 2011MEPS..434..229G , doi : 10.3354/meps09084 , JSTOR 24875453
Внешние ссылки [ править ]
- Друзья Фамозы Слау
- Ресурс по географии для школ
- Джонсон, Калифорния (2006) «Причина, которую ищут, когда болота превращаются в бесплодные равнины» The Boston Globe
- приливных солончаков Проект восстановления
- Зона изучения морской природы, находящаяся в ведении Департамента охраны природы и водных путей города Хемпстед, расположенная в Оушенсайде, штат Нью-Йорк, США.