Глубоководная рыба

Глубоководные рыбы обитающие в темноте под освещенными солнцем поверхностными водами, то есть ниже эпипелагиали или фотической зоны моря — это рыбы , . Рыба -фонарь , безусловно, самая распространенная глубоководная рыба. Другие глубоководные рыбы включают рыбу-фонарик , акулу-печеньку , щетинистую рыбу , удильщика , рыбу-гадюку и некоторые виды бельдюгов .
Лишь около 2% известных морских видов обитают в пелагической среде. Это означает, что они живут в толще воды , в отличие от бентосных организмов, обитающих на морском дне. [1] Глубоководные организмы обычно обитают в батипелагических (глубина 1 000–4 000 метров (3 281–13 123 футов)) и абиссопелагических (глубина 4 000–6 000 метров (13 123–19 685 футов) зонах. Однако характеристики глубоководных организмов, такие как биолюминесценция, можно увидеть и в мезопелагической (глубиной 200–1000 метров (656–3281 футов)) зоне. Мезопелагическая зона является дисфотической зоной, то есть свет там минимален, но все же измерим. Минимальный слой кислорода существует где-то на глубине от 700 метров (2297 футов) до 1000 метров (3281 фут) в зависимости от места в океане. В этой области также больше всего питательных веществ. Батипелагическая абиссопелагическая и , то есть свет не проникает в зоны афотические эту область океана. Эти зоны составляют около 75% обитаемого пространства океана. [2]
Эпипелагическая зона (глубина 0–200 метров (0–656 футов)) — это область, где свет проникает в воду и происходит фотосинтез. Это также известно как фотозона. Поскольку обычно оно простирается всего на несколько сотен метров под водой, глубокое море, составляющее около 90% объема океана, находится во тьме. Глубокое море также является чрезвычайно агрессивной средой: температура редко превышает 3 °C (37 °F) и опускается до -1,8 °C (29 °F) (за исключением экосистем гидротермальных жерл , температура которых может превышать 350 °F). C или 662 °F), низкий уровень кислорода и давление от 20 до 1000 атм (от 2 до 100 МПа ). [3]
Эволюция
[ редактировать ]Было высказано предположение, что глубоководные экосистемы могли быть негостеприимными для жизни позвоночных до увеличения притока питательных веществ в океан в позднеюрском и раннем меловом периоде после появления покрытосеменных растений на суше, что привело к увеличению жизни абиссальных беспозвоночных. , что позволяет рыбам, в свою очередь, колонизировать эти экосистемы. Однако некоторые современные глубоководные рыбы, такие как голоцефалы, являются потомками гораздо более древних линий, что указывает на то, что могла произойти гораздо более ранняя колонизация глубоководных вод позвоночными, хотя никаких ископаемых свидетельств этого не известно. [4] [5]
Самыми ранними известными находками глубоководных рыб являются следы окаменелостей , связанных с питанием и плавательным поведением, приписываемые неопознанным неотелеостам (относящимся к ихнородам Piscichnus и Undichna ) из раннемеловых (возрастом 130 миллионов лет) сланцев Паломбини в Италии , которые Считается, что он отложился на абиссальной равнине бывшего Пьемонт-Лигурийского океана . До открытия этих окаменелостей не было никаких свидетельств существования глубоководных костных рыб старше 50 миллионов лет в палеогене . [4] [5] Меловое происхождение большинства современных глубоководных рыб было дополнительно подтверждено филогенетическими исследованиями, такими как исследования аулопиформных рыб, которые показывают, что многие глубоководные линии этих групп возникли примерно в это время. [6]
Хотя записи из сланцев Паломбини представляют собой самые ранние находки глубоководных костистых рыб, глубоководных акул, известны также формации, в которых сохранились окаменелости из более позднего мелового периода. К ним относятся формация Нортумберленд в Канаде и отложения аналогичного возраста в Анголе, в которых сохранились окаменелости таксонов, таких как гексанхиды , хламидоселахиды и кошачьи акулы , которые сегодня известны из глубоководных местообитаний, но редки в других формациях того времени. [7] Палеогеновые образования с ископаемыми зубами глубоководных акул известны в Новой Зеландии для среднего палеоцена , а в Дании, Франции, Австрии и Марокко — в эоцене. [8] В море Паратетис до миоцена все еще обитали глубоководные акулы и скаты, которые сохранились в формациях в Венгрии. [9]
В палеогене известны заметные образования, в которых сохранились хорошо сочлененные экземпляры глубоководных костистых рыб. К ним относятся лагерштатте Монте-Солане в раннем эоцене Италии, который сохраняет батипелагическую среду обитания, вероятно, отложенную на глубине 300–600 метров под водой, а также позднеэоценовую формацию Пабде в Иране . Глубоководная среда, сохранившаяся в обеих формациях, проявляется в обилии ископаемых стомиообразных рыб. [10] [11] Известные неогеновые образования, в которых сохранились окаменелости глубоководных костистых рыб, известны из миоцена Италии, Японии и Калифорнии. [12] [13] [14]
Среда
[ редактировать ]
В глубоком океане воды простираются намного ниже эпипелагиали и служат средой обитания самых разных типов пелагических рыб, приспособленных к жизни в этих более глубоких зонах. [15] На глубокой воде морской снег представляет собой непрерывный дождь преимущественно органического детрита, выпадающего из верхних слоев водной толщи. Его происхождение лежит в деятельности продуктивной фотозоны . Морской снег включает мертвый или умирающий планктон , простейших ( диатомовые водоросли ), фекалии, песок, сажу и другую неорганическую пыль. «Снежинки» со временем растут и могут достигать нескольких сантиметров в диаметре, путешествуя неделями, прежде чем достичь дна океана. Однако большинство органических компонентов морского снега потребляются микробами , зоопланктоном и другими животными-фильтраторами в пределах первых 1000 метров (3281 фут) их пути, то есть в пределах эпипелагиали. Таким образом, морской снег можно считать основой глубоководных мезопелагических и донных экосистем : поскольку солнечный свет не может достичь их, глубоководные организмы в значительной степени полагаются на морской снег как на источник энергии. Поскольку на морских глубинах (афотические) нет света, первичных продуцентов не хватает. Таким образом, большинство организмов в батипелагиали полагаются на морской снег из регионов, расположенных выше по вертикальной колонне.
Некоторые глубоководные пелагические группы, такие как семейства фонарей , риджхедов , морских топориков и легких рыб , иногда называют псевдокеаническими , потому что вместо равномерного распределения в открытой воде они встречаются в значительно более высоких количествах вокруг структурных оазисов, особенно подводных гор и над океанами. континентальные склоны . Это явление объясняется также обилием видов добычи, которых также привлекают структуры.
Гидростатическое давление увеличивается на 1 атм (0,1 МПа) на каждые 10 м (32,8 фута) глубины. [16] Глубоководные организмы испытывают внутри своего тела такое же давление, как и извне, поэтому они не раздавливаются экстремальным давлением. Однако их высокое внутреннее давление приводит к снижению текучести их мембран, поскольку молекулы сжимаются вместе. Текучесть клеточных мембран повышает эффективность биологических функций, прежде всего продукции белков, поэтому организмы приспособились к этому обстоятельству за счет увеличения доли ненасыщенных жирных кислот в липидах клеточных мембран. [17] Помимо различий во внутреннем давлении, у этих организмов сложился иной баланс метаболических реакций, чем у организмов, обитающих в эпипелагиали. Дэвид Уортон, автор книги « Жизнь на пределе: организмы в экстремальных условиях» , отмечает: «Биохимические реакции сопровождаются изменениями объема. Если реакция приводит к увеличению объема, она будет тормозиться давлением, тогда как, если она связана с уменьшение объема, оно будет усилено». [18] Это означает, что их метаболические процессы должны в конечном итоге до некоторой степени уменьшить объем организма.

Большинство рыб, эволюционировавших в этой суровой среде, не способны выжить в лабораторных условиях, а попытки удержать их в неволе привели к их гибели. Глубоководные организмы содержат газонаполненные пространства (вакуоли). Газ сжимается под высоким давлением и расширяется под низким давлением. Известно, что из-за этого эти организмы взрываются, если выходят на поверхность. [18]
Характеристики
[ редактировать ]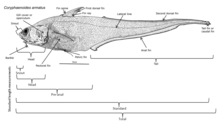

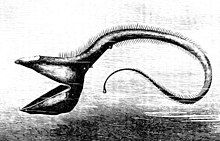

Глубоководные рыбы развили различные приспособления, чтобы выжить в этом регионе. Поскольку многие из этих рыб обитают в регионах, где нет естественного освещения , они не могут полагаться исключительно на свое зрение при обнаружении добычи и партнеров и уклонении от хищников; глубоководные рыбы эволюционировали в соответствии с крайней субфотической областью, в которой они обитают. Многие из этих организмов слепы и полагаются на другие свои чувства, такие как чувствительность к изменениям местного давления и запаха, чтобы поймать пищу и избежать поимки. Те, кто не слеп, имеют большие и чувствительные глаза, которые могут использовать биолюминесцентный свет. Эти глаза могут быть в 100 раз более чувствительными к свету, чем человеческие глаза. Родопсин (Rh1) — это белок, содержащийся в палочковидных клетках глаза, который помогает животным видеть в тусклом свете. В то время как у большинства позвоночных обычно есть один ген опсина Rh1, у некоторых глубоководных рыб имеется несколько генов Rh1, а у одного вида, серебряного колючего плавника ( Diretmus argenteus ), их 38. [20] Такое распространение генов Rh1 может помочь глубоководным рыбам видеть в глубинах океана. Кроме того, чтобы избежать нападения хищников, многие виды имеют темную окраску, чтобы сливаться с окружающей средой. [21]
Многие глубоководные рыбы обладают биолюминесценцией и имеют чрезвычайно большие глаза, приспособленные к темноте. Биолюминесцентные организмы способны биологически производить свет за счет возбуждения молекул люциферина, которые затем производят свет. Этот процесс должен происходить в присутствии кислорода. Эти организмы распространены в мезопелагической области и ниже (200 метров (656 футов) и ниже). Более 50% глубоководных рыб, а также некоторые виды креветок и кальмаров способны к биолюминесценции. Около 80% этих организмов имеют фотофоры – светопродуцирующие железистые клетки, содержащие светящиеся бактерии, окаймленные темной окраской. Некоторые из этих фотофоров содержат линзы, очень похожие на линзы в глазах человека, которые могут усиливать или ослаблять излучение света. Способность производить свет требует всего 1% энергии организма и имеет множество целей: он используется для поиска пищи и привлечения добычи, как у удильщика; претендовать на территорию посредством патрулирования; общаться и находить себе пару, а также отвлекать или временно ослеплять хищников, чтобы сбежать. Кроме того, в мезопелагиали, куда все еще проникает некоторый свет, некоторые организмы маскируются от хищников под ними, освещая свое брюшко так, чтобы оно соответствовало цвету и интенсивности света сверху, чтобы не отбрасывалось тени. Эта тактика известна как контриллюминация. [22]
Жизненный цикл глубоководных рыб может быть исключительно глубоководным, хотя некоторые виды рождаются на мелководье и по мере взросления тонут. Независимо от глубины, на которой находятся яйца и личинки, они обычно пелагические. Этот планктонный, дрейфующий образ жизни требует нейтральной плавучести. Чтобы поддерживать это явление, яйца и личинки часто содержат в плазме капли масла. [23] Когда эти организмы находятся в полностью зрелом состоянии, им необходимы другие приспособления, чтобы сохранять свое положение в толще воды. В общем, плотность воды вызывает подъем — аспект плавучести, который заставляет организмы плавать. Чтобы противодействовать этому, плотность организма должна быть больше, чем плотность окружающей воды. Большинство тканей животных плотнее воды, поэтому им приходится находить равновесие, чтобы плавать. [24] У многих организмов развиваются плавательные пузыри (газовые полости), чтобы оставаться на плаву, но из-за высокого давления окружающей среды у глубоководных рыб этот орган обычно отсутствует. Вместо этого они имеют конструкции, похожие на подводные крылья, для обеспечения гидродинамической подъемной силы. Было также обнаружено, что чем глубже обитает рыба, тем более желеобразной ее плоть и более минимальной костной структурой. Они уменьшают плотность своих тканей за счет высокого содержания жира, уменьшения массы скелета, что достигается за счет уменьшения размера, толщины и содержания минералов, а также накопления воды. [25] делает их медленнее и менее подвижными, чем поверхностные рыбы.
Из-за низкого уровня фотосинтетического света, достигающего глубоководной среды, большинству рыб приходится полагаться на органическое вещество, опускающееся с более высоких уровней, или, в редких случаях, на гидротермальные источники для получения питательных веществ. Это делает глубоководные районы гораздо менее продуктивными , чем более мелководные. Кроме того, животных в пелагической среде мало, и пища попадается нечасто. Из-за этого организмы нуждаются в приспособлениях, позволяющих им выжить. У некоторых есть длинные щупальца, которые помогают им находить добычу или привлекать партнеров в кромешной тьме глубокого океана. В частности, глубоководная рыба-удильщик имеет длинное приспособление, напоминающее удочку, выступающее из ее лица, на конце которого находится биолюминесцентный кусок кожи, который извивается, как червь, чтобы заманить добычу. Некоторым приходится потреблять другую рыбу такого же размера или больше, чем они, и им нужны приспособления, которые помогут эффективно их переваривать. Огромные острые зубы, шарнирные челюсти, непропорционально большие рты и расширяющееся тело — вот лишь некоторые из характеристик, которыми обладают глубоководные рыбы для этой цели. [21] Угорь -глотун — один из примеров организма, обладающего этими характеристиками.
Рыбы в различных пелагических и глубоководных донных зонах имеют физическое строение и поведение, заметно отличающееся друг от друга. Группы сосуществующих видов в каждой зоне, похоже, действуют схожим образом, например, небольшие мезопелагические вертикально мигрирующие питающиеся планктоном, батипелагические удильщики и глубоководные донные гремуучие хвосты . [26]
Виды скатов с колючими плавниками редки среди глубоководных рыб, что позволяет предположить, что глубоководные рыбы являются древними и настолько хорошо адаптированы к окружающей среде, что вторжения более современных рыб оказались безуспешными. [27] Немногие лучевые плавники существуют в основном у Beryciformes и Lampriformes , которые также являются древними формами. Большинство глубоководных пелагических рыб принадлежат к своим отрядам, что позволяет предположить длительную эволюцию в глубоководной среде. Напротив, глубоководные донные виды относятся к отрядам, включающим множество родственных мелководных рыб. [28]
| Виды по пелагической зоне |
|---|
| Сравнительное строение пелагических рыб. |
|---|
Мезопелагическая рыба
[ редактировать ]




Ниже эпипелагиали условия быстро меняются. На высоте от 200 м до примерно 1000 м свет продолжает тускнеть, пока его почти не станет совсем. Температура падает через термоклин до температуры от 3,9 ° C (39 ° F) до 7,8 ° C (46 ° F). Это сумеречная или мезопелагическая зона. Давление продолжает расти со скоростью один атм (0,1 МПа) каждые 10 м (32,8 фута), в то время как концентрация питательных веществ падает вместе с растворенным кислородом и скоростью циркуляции воды. [15]
Операторы гидролокаторов , использовавшие недавно разработанную гидролокационную технологию во время Второй мировой войны , были озадачены тем, что выглядело ложным морским дном глубиной 300–500 метров (984–1640 футов) днем и меньшей глубиной ночью. Оказалось, что это произошло благодаря миллионам морских организмов, особенно мелких мезопелагических рыб, плавательные пузыри которых отражали сигналы сонара. Эти организмы мигрируют на мелководье в сумерках, чтобы питаться планктоном. Слой глубже, когда луна светит, и может стать мельче, когда облака проходят над луной. Это явление стало известно как глубокий слой рассеяния . [34]
Большинство мезопелагических рыб совершают ежедневные вертикальные миграции , перемещаясь ночью в эпипелагическую зону, часто следуя за подобными миграциями зоопланктона, и возвращаясь на глубину в целях безопасности в течение дня. [15] [35] Эти вертикальные миграции часто происходят на большие вертикальные расстояния и осуществляются с помощью плавательного пузыря . [27] Плавательный пузырь надувается, когда рыба хочет двигаться вверх, а, учитывая высокое давление в мессоплегической зоне, это требует значительных затрат энергии. По мере подъема рыбы давление в плавательном пузыре должно регулироваться, чтобы предотвратить его разрыв. Когда рыба хочет вернуться на глубину, плавательный пузырь сдувается. [36] Некоторые мезопелагические рыбы совершают ежедневные миграции через термоклин , где температура колеблется от 50 ° F (10 ° C) до 69 ° F (20 ° C), таким образом демонстрируя значительную устойчивость к изменению температуры. [37]
У этих рыб мускулистое тело, окостеневшие кости, чешуя, хорошо развитые жабры и центральная нервная система, а также большие сердце и почки. Мезопелагические питатели планктона имеют маленькие рты с тонкими жаберными тычинками , тогда как у рыбоядных есть рот большего размера и более грубые жаберные тычинки. [15]
Мезопелагические рыбы приспособлены к активной жизни в условиях недостаточной освещенности. Большинство из них — зрительные хищники с большими глазами. У некоторых глубоководных рыб глаза трубчатые с большими линзами и только палочковые клетки смотрят вверх. Они обеспечивают бинокулярное зрение и высокую чувствительность к слабым световым сигналам. [15] Эта адаптация дает улучшенное конечное зрение за счет бокового зрения и позволяет хищнику различать кальмаров , каракатиц и более мелкую рыбу, силуэты которых вырисовываются на фоне мрака над ними.
Мезопелагические рыбы обычно не имеют защитных шипов и используют цвет, чтобы маскироваться от других рыб. Засадные хищники темные, черные или красные. Поскольку более длинные красные волны света не достигают морских глубин, красный цвет действует так же, как черный. Мигрирующие формы используют с контрастными оттенками серебристые цвета . На животе у них часто имеются фотофоры, излучающие слабый свет. Для хищника снизу, смотрящего вверх, эта биолюминесценция маскирует силуэт рыбы. Однако у некоторых из этих хищников есть желтые линзы, которые фильтруют окружающий свет (с недостатком красного цвета), оставляя видимой биолюминесценцию. [38]
Коричневырылая рыба-призрак , разновидность бочкоглаза , является единственным известным позвоночным, которое использует зеркало, а не линзу, чтобы сфокусировать изображение в своих глазах. [39] [40]
Отбор проб глубоким тралом показывает, что на долю фонарей приходится до 65% всей биомассы глубоководных рыб . [41] Действительно, рыбы-фонарики являются одними из наиболее широко распространенных, густонаселенных и разнообразных среди всех позвоночных животных , играя важную экологическую роль в качестве добычи для более крупных организмов. По оценкам, глобальная биомасса фонарей составляет 550–660 миллионов тонн , что в несколько раз превышает весь мировой вылов рыбы. Рыбы-фонарики также составляют большую часть биомассы, ответственной за глубокий рассеивающий слой мирового океана. [42]
Большеглазый тунец — эпипелагический/мезопелагический вид, питающийся другой рыбой. Спутниковое мечение показало, что большеглазый тунец в дневное время часто проводит длительные периоды времени, путешествуя глубоко под поверхностью воды, иногда ныряя на глубину до 500 метров (1640 футов). Считается, что эти движения являются ответом на вертикальные миграции организмов-жертв в глубоком слое рассеяния .
 У стоп-сигнала свободная челюсть составляет нижняя челюсть четверть длины тела. Челюсть не имеет дна и крепится только шарниром и видоизмененной язычной костью. За большими клыкообразными зубами спереди следует множество мелких зазубренных зубов. [43] [44]
У стоп-сигнала свободная челюсть составляет нижняя челюсть четверть длины тела. Челюсть не имеет дна и крепится только шарниром и видоизмененной язычной костью. За большими клыкообразными зубами спереди следует множество мелких зазубренных зубов. [43] [44]![Светоотражатель также является одной из немногих рыб, которые производят красную биолюминесценцию. Поскольку большая часть их жертв не может воспринимать красный свет, это позволяет им охотиться с помощью практически невидимого луча света.[43]](//upload.wikimedia.org/wikipedia/commons/thumb/b/b2/Malacosteus.JPG/120px-Malacosteus.JPG) Светофорная челюсть также является одной из немногих рыб, которые производят красную биолюминесценцию . Поскольку большая часть их добычи не может воспринимать красный свет, это позволяет ей охотиться с помощью практически невидимого луча света. [43]
Светофорная челюсть также является одной из немногих рыб, которые производят красную биолюминесценцию . Поскольку большая часть их добычи не может воспринимать красный свет, это позволяет ей охотиться с помощью практически невидимого луча света. [43]![Длиннорылая ланцетная рыба. Ланцетные рыбы — хищники-засадники, все время проводят в мезопелагической зоне. Они входят в число крупнейших мезопелагических рыб (до 2 м).[45]](//upload.wikimedia.org/wikipedia/commons/thumb/0/0d/Longnoselancetfish.jpg/120px-Longnoselancetfish.jpg) Длиннорылая ланцетная рыба . Ланцетные рыбы — хищники-засадники, все время проводят в мезопелагической зоне. Это одни из самых крупных мезопелагических рыб (до 2 м). [45]
Длиннорылая ланцетная рыба . Ланцетные рыбы — хищники-засадники, все время проводят в мезопелагической зоне. Это одни из самых крупных мезопелагических рыб (до 2 м). [45] Кинжалозуб парализует других мезопелагических рыб , когда кусает их кинжаловидными зубами. [46]
Кинжалозуб парализует других мезопелагических рыб , когда кусает их кинжаловидными зубами. [46]
![Светоотражатель также является одной из немногих рыб, которые производят красную биолюминесценцию. Поскольку большая часть их жертв не может воспринимать красный свет, это позволяет им охотиться с помощью практически невидимого луча света.[43]](http://upload.wikimedia.org/wikipedia/commons/thumb/b/b2/Malacosteus.JPG/120px-Malacosteus.JPG)
![Длиннорылая ланцетная рыба. Ланцетные рыбы — хищники-засадники, все время проводят в мезопелагической зоне. Они входят в число крупнейших мезопелагических рыб (до 2 м).[45]](http://upload.wikimedia.org/wikipedia/commons/thumb/0/0d/Longnoselancetfish.jpg/120px-Longnoselancetfish.jpg)

Батипелагическая рыба
[ редактировать ]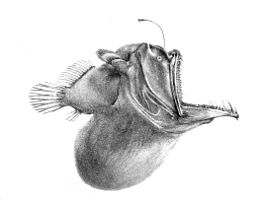



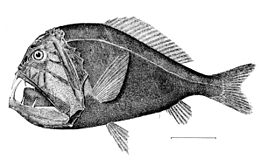
Ниже мезопелагической зоны темно. Это полуночная (или батипелагическая зона ), простирающаяся от 1000 метров (3281 фут) до нижней глубоководной бентической зоны . Если вода исключительно глубокая, пелагическую зону ниже 4000 метров (13 123 фута) иногда называют нижней полночью (или абиссопелагической зоной ). Температура в этой зоне колеблется от 1 до 4 ° C (от 34 до 39 ° F), и она полностью афотична.
Условия в этих зонах в некоторой степени однородны; тьма полная, давление сокрушительное, а температура, уровень питательных веществ и растворенного кислорода низкие. [15]
Батипелагические рыбы обладают особыми приспособлениями, позволяющими справляться с этими условиями: у них медленный метаболизм и неспециализированная диета, они готовы есть все, что попадается под руку. Они предпочитают сидеть и ждать еды, а не тратить энергию на ее поиски. Поведение батипелагических рыб можно противопоставить поведению мезопелагических рыб. Мезопелагические рыбы часто очень подвижны, тогда как батипелагические рыбы почти все являются хищниками, затаившимися в засаде, обычно затрачивающими мало энергии на движение. [53]
Преобладающими батипелагическими рыбами являются щетинорот и удильщик ; клыказуб , гадюка , кинжалозуб и барракудина Также распространены . Эти рыбы маленькие, многие из них имеют длину около 10 сантиметров (3,9 дюйма), но немногие длиннее 25 сантиметров (9,8 дюйма). Большую часть времени они терпеливо ждут в толще воды, пока не появится добыча или ее не заманит их люминофор. То немногое энергии, которое имеется в батипелагической зоне, фильтруется сверху в виде детрита, фекалий и редких беспозвоночных или мезопелагических рыб. [53] Около 20 процентов пищи, происходящей из эпипелагиали, попадает в мезопелагическую зону. [34] но только около 5 процентов фильтруется в батипелагическую зону. [47]
Батипелагические рыбы ведут оседлый образ жизни и приспособлены к выработке минимума энергии в среде обитания с очень небольшим количеством пищи или доступной энергии, даже не солнечного света, а только биолюминесценции. Их тела удлиненные , со слабыми водянистыми мышцами и скелетными структурами. Поскольку большая часть рыбы состоит из воды, на этих глубинах они не сжимаются большим давлением. У них часто есть выдвижные шарнирные челюсти с загнутыми зубами. Они склизкие, без чешуи . Центральная нервная система ограничена боковой линией и обонятельной системой, глаза маленькие и могут не функционировать, а жабры , почки и сердце, а также плавательный пузырь маленькие или отсутствуют. [47] [54] Это те же особенности, которые обнаружены у личинок рыб , что позволяет предположить, что в ходе своей эволюции батипелагические рыбы приобрели эти особенности посредством неотении . Как и в случае с личинками, эти особенности позволяют рыбе оставаться в воде с небольшими затратами энергии. [55]
Несмотря на свой свирепый вид, эти формы в основном представляют собой миниатюрных рыбок со слабыми мышцами и слишком малы, чтобы представлять какую-либо угрозу для человека. Исключением из общего небольшого размера батипелагических рыб является скользеголовка Йокодзуна ( Narcetes shonanmaruae ), описанная в 2021 году, которая является самой крупной из известных полностью батипелагических костистых рыб , ее длина превышает 2,5 метра (8,2 фута). [56]
Плавательные пузыри глубоководных рыб либо отсутствуют, либо почти не функционируют, а батипелагические рыбы обычно не совершают вертикальных миграций. Наполнение мочевых пузырей при таком высоком давлении требует огромных затрат энергии. У некоторых глубоководных рыб есть плавательные пузыри, которые функционируют в молодом возрасте и обитают в верхней эпипелагиали, но увядают или наполняются жиром, когда рыбы спускаются к своей взрослой среде обитания. [57]
Парой известных исключений являются обыкновенный угорь ( Holcomycteronus profundissimus ), добытый на глубине 7160 метров, и грубый абиссальный гренадер ( Coryphaenoides yaquinae ), найденный на глубине 7259 метров. У этих видов все еще есть функциональные плавательные пузыри, что позволяет им сохранять высокую плотность костей и сильные челюсти. [58]
Наиболее важными сенсорными системами обычно являются внутреннее ухо , реагирующее на звук, и боковая линия , реагирующая на изменения давления воды. Обонятельная . система также может быть важна для самцов, которые находят самок по запаху [59] Батипелагические рыбы черные, а иногда и красные, с небольшим количеством фотофоров . Фотофоры обычно используются для соблазнения добычи или привлечения партнера. Поскольку еды очень мало, батипелагические хищники не избирательны в своих пищевых привычках, а хватают все, что оказывается достаточно близко. Они достигают этого за счет большого рта с острыми зубами для захвата крупной добычи и перекрывающихся жаберных тычинок , которые не позволяют проглоченной мелкой добыче ускользнуть. [54]
В этой зоне нелегко найти себе пару. Некоторые виды зависят от биолюминесценции , при этом образцы биолюминесценции уникальны для конкретных видов. Другие являются гермафродитами , что удваивает их шансы на производство яйцеклеток и сперматозоидов при столкновении. [47] Самка удильщика выделяет феромоны , чтобы привлечь крошечных самцов. Когда самец находит ее, он кусает ее и уже не отпускает. Когда самец вида удильщиков Haplophryne mollis кусает кожу самки, он выделяет фермент , который переваривает кожу его рта и ее тела, соединяя пару до точки, где соединяются две системы кровообращения. Затем самец атрофируется, превращаясь в пару гонад . Этот крайний половой диморфизм гарантирует, что, когда самка будет готова к нересту, у нее сразу же появится партнер. [60]
В батипелагиали обитают многие формы, отличные от рыб, такие как кальмары, крупные киты, осьминоги, губки, брахиоподы , морские звезды и морские ежи , но эта зона трудна для обитания рыб.
 Пеликан -угорь использует свой большой рот как сеть, открывая челюсти и плывя к добыче. На кончике хвоста у него есть люминесцентный орган, привлекающий добычу.
Пеликан -угорь использует свой большой рот как сеть, открывая челюсти и плывя к добыче. На кончике хвоста у него есть люминесцентный орган, привлекающий добычу. Черный глотатель с его растяжимым желудком отличается способностью глотать целиком костистых рыб , в десять раз превышающих его массу. [61] [62]
Черный глотатель с его растяжимым желудком отличается способностью глотать целиком костистых рыб , в десять раз превышающих его массу. [61] [62] Самки удильщика Haplophryne mollis прикрепляют к себе самцов, которые атрофировались в пару гонад, для использования, когда самка готова к нересту.
Самки удильщика Haplophryne mollis прикрепляют к себе самцов, которые атрофировались в пару гонад, для использования, когда самка готова к нересту.![Сликхед Ёкодзуна — крупнейшая из известных батипелагических костистых рыб, его длина превышает 250 сантиметров.[56]](//upload.wikimedia.org/wikipedia/commons/thumb/1/1c/Narcetes_shonanmaruae_a_frozen_specimen.jpg/120px-Narcetes_shonanmaruae_a_frozen_specimen.jpg) — Сликхед Ёкодзуна крупнейшая известная батипелагическая костистая рыба, ее длина превышает 250 сантиметров. [56]
— Сликхед Ёкодзуна крупнейшая известная батипелагическая костистая рыба, ее длина превышает 250 сантиметров. [56]


![Сликхед Ёкодзуна — крупнейшая из известных батипелагических костистых рыб, его длина превышает 250 сантиметров.[56]](http://upload.wikimedia.org/wikipedia/commons/thumb/1/1c/Narcetes_shonanmaruae_a_frozen_specimen.jpg/120px-Narcetes_shonanmaruae_a_frozen_specimen.jpg)
Адаптация к высокому давлению
[ редактировать ]По мере того как рыба продвигается глубже в море, вес воды над головой оказывает возрастающее гидростатическое давление на рыбу . Это повышенное давление составляет около одного атм (0,1 МПа) на каждые 10 м (32,8 фута) глубины. Для рыбы на дне батипелагической зоны это давление составляет около 400 атм (40 МПа, или почти 6000 фунтов на квадратный дюйм). [63]
Глубоководные организмы обладают адаптациями на клеточном и физиологическом уровнях, которые позволяют им выживать в условиях большого давления. Отсутствие этих приспособлений ограничивает глубины, на которых могут обитать мелководные виды. Высокий уровень внешнего давления влияет на то, как протекают обменные процессы и биохимические реакции. Равновесие многих химических реакций нарушается давлением, а давление может тормозить процессы, приводящие к увеличению объема. Вода, ключевой компонент многих биологических процессов, очень чувствительна к изменениям объема, главным образом потому, что компоненты клеточной жидкости влияют на структуру воды. Таким образом, ферментативные реакции, вызывающие изменения в организации воды, эффективно изменяют объем системы. [64] Белки, ответственные за катализ реакций, обычно удерживаются вместе слабыми связями , и реакции обычно сопровождаются увеличением объема. [65]
Виды, которые могут переносить такие глубины, претерпели изменения в своей белковой структуре и критериях реакции, чтобы выдерживать давление и выполнять реакции в этих условиях. В условиях высокого давления двухслойные клеточные мембраны теряют текучесть. Глубоководные клеточные мембраны отдают предпочтение бислоям фосфолипидов с более высокой долей ненасыщенных жирных кислот , которые обеспечивают более высокую текучесть, чем их аналоги на уровне моря.
Десять отрядов, тринадцать семейств и около 200 известных видов глубоководных рыб развили студенистый слой под кожей или вокруг позвоночника, который используется для плавучести, быстрого роста и повышения эффективности плавания за счет уменьшения сопротивления. [66]

Глубоководные виды демонстрируют меньшие изменения энтропии и энтальпии по сравнению с организмами поверхностного уровня, поскольку среда с высоким давлением и низкой температурой способствует отрицательным изменениям энтальпии и снижению зависимости от реакций, обусловленных энтропией. Со структурной точки зрения глобулярные белки глубоководных рыб из-за третичной структуры G-актина относительно жесткие по сравнению с белками поверхностных рыб. [64] Тот факт, что белки глубоководных рыб структурно отличаются от белков поверхностных рыб, очевиден из наблюдения, что актин мышечных волокон глубоководных рыб чрезвычайно термоустойчив; наблюдение похожее на то, что наблюдается у ящериц. Эти белки структурно укрепляются за счет модификации связей в третичной структуре белка, что также приводит к повышению уровня термической стабильности. [65] Белки структурно укрепляются, чтобы противостоять давлению за счет модификации связей в третичной структуре. [65] Следовательно, высокий уровень гидростатического давления, аналогичный высоким температурам тела термофильных рептилий пустыни, благоприятствует жестким белковым структурам.
Na+/K+-АТФаза представляет собой фермент липопротеина , который играет важную роль в осморегуляции и находится под сильным влиянием гидростатического давления. Ингибирование Na+/K+-АТФазы происходит за счет увеличения сжатия под действием давления. Ограничивающая скорость стадия реакции Na+/K+-АТФазы вызывает расширение бислоя, окружающего белок, и, следовательно, увеличение объема. Увеличение объема делает реактивность Na+/K+-АТФазы чувствительной к более высокому давлению. Хотя активность Na+/K+-АТФазы на грамм жаберной ткани ниже у глубоководных рыб, Na+/K+-АТФазы глубоководных рыб демонстрируют гораздо более высокую толерантность к гидростатическому давлению по сравнению с их мелководными собратьями. Примером этого является вид C. acrolepis (глубина около 2000 м) и его хадальпелагический аналог C. Armatus (глубина около 4000 метров (13 123 фута)), где Na+/K+-АТФазы C. Armatus гораздо менее чувствительны к давлению. . Такое сопротивление давлению можно объяснить адаптацией белков и липидов. фрагменты Na+/K+-АТФазы. [67]
Рыба-фонарь
[ редактировать ]
Отбор проб с помощью глубоководного траления показывает, что на долю фонарей приходится до 65% всей биомассы глубоководных рыб . [41] Действительно, рыбы-фонарики являются одними из наиболее широко распространенных, густонаселенных и разнообразных среди всех позвоночных животных , играя важную экологическую роль в качестве добычи для более крупных организмов. Имея предполагаемую глобальную биомассу в 550–660 миллионов метрических тонн , что в несколько раз превышает весь мировой вылов рыбы, на фонарей также приходится большая часть биомассы, ответственной за глубокий рассеивающий слой мирового океана. В Южном океане миктофиды служат альтернативой крилю пищевого ресурса для таких хищников, как кальмары и королевские пингвины . Хотя эта рыба многочисленна и плодовита, в настоящее время существует лишь несколько коммерческих промыслов фонаря: они включают ограниченные операции у берегов Южной Африки , в субантарктическом регионе и в Оманском заливе .
Вымирающие виды
[ редактировать ]Исследование канадских учёных 2006 года выявило, что пять видов глубоководных рыб — синий хек , колючий угорь — находятся на грани исчезновения из-за смещения коммерческого рыболовства с континентальных шельфов на склоны континентальных шельфов , на глубины. 1600 метров (5249 футов). Медленное размножение этих рыб – они достигают половой зрелости примерно в том же возрасте, что и люди – является одной из основных причин того, что они не могут оправиться от чрезмерного вылова . [68]
См. также
[ редактировать ]- Перепись морской жизни
- Глубокая океанская вода
- Глубокое море
- Глубоководные сообщества
- Демерсальная рыба
- Пелагическая рыба
Цитаты
[ редактировать ]- ^ Трухильо и Турман 2011 , стр. 354.
- ^ Трухильо и Турман 2011 , стр. 365.
- ^ Трухильо и Турман 2011 , стр. 457, 460.
- ^ Перейти обратно: а б Бокон, Андреа; Ферретти, Анналиса; Фиорони, Кьяра; Пандольфи, Лука; Серпальи, Генри; Пиччинини, Армандо; де Карвалью, Шарль Нето; Качао, Марио; Линли, Томас; Мунис, Фернандо; из Беларуси Заин; Джеймисон, Алан; Русский Джироламо; Геррини, Филип; Феррандо, Сара (12 сентября 2023 г.). «Самые ранние свидетельства существования глубоководных позвоночных » Труды Национальной академии наук . 120 (37): e2306164120. Бибкод : 2023PNAS..12006164B . дои : 10.1073/pnas.2306164120 . ISSN 0027-8424 . ПМК 10500276 . ПМИД 37669391 .
- ^ Перейти обратно: а б Сотрудники, Новости (19 сентября 2023 г.). «Окаменелости возрастом 130 миллионов лет раскрывают самые ранние свидетельства существования глубоководных позвоночных | Sci.News» . Sci.News: Последние новости науки . Проверено 30 сентября 2023 г.
{{cite web}}:|first=имеет общее имя ( справка ) - ^ Дэвис, Мэтью П.; Филитц, Кристофер (1 декабря 2010 г.). «Оценка времени расхождения рыб-ящериц и их союзников (Euteleostei: Aulopiformes) и сроков глубоководной адаптации» . Молекулярная филогенетика и эволюция . 57 (3): 1194–1208. Бибкод : 2010МОЛПЭ..57.1194Д . дои : 10.1016/j.ympev.2010.09.003 . ISSN 1055-7903 . ПМИД 20854916 .
- ^ Каппетта, Анри; Моррисон, Курт; Аднет, Сильвен (3 августа 2021 г.). «Фауна акул из кампана острова Хорнби, Британская Колумбия, Канада: взгляд на разнообразие меловых глубоководных комплексов» . Историческая биология . 33 (8): 1121–1182. Бибкод : 2021HBio...33.1121C . дои : 10.1080/08912963.2019.1681421 . ISSN 0891-2963 . S2CID 212878837 .
- ^ «Вестник Том 72 – 2023» . Датская геологическая ассоциация (на датском языке). 27 января 2023 г. doi : 10.37570/bgsd-2023-72-06 . Проверено 8 марта 2024 г.
- ^ Сабо, Мартон; Кочиш, Ласло; Тот, Эмоке; Сабо, Питер; Немет, Тамаш; Себе, Кристина (2022). Кэвин, Лайонел (ред.). «Хондрихтианы (Holocephali, Squalomorphii и Batomorphii) остаются из бадена южной Венгрии (Текерес, горы Мечек): первые глубоководные хрящевые рыбы из среднего миоцена Центрального Паратетиса» . Статьи по палеонтологии . 8 (6). Бибкод : 2022PPal....8E1471S . дои : 10.1002/spp2.1471 . ISSN 2056-2799 .
- ^ Джусберти, Лука; Банников, Александр; Босколо Галаццо, Флавия; Форначари, Элиана; Фрилинг, Йост; Лучани, Валерия; Папаццони, Чезаре Андреа; Роги, Гвидо; Схоутен, Стефан; Слуйс, Аппи; Боселлини, Франческа Р.; Зорзин, Роберто (1 июня 2014 г.). «Новый ископаемый лагерштетте из нижнего эоцена гор Лессини (северная Италия): междисциплинарный подход» . Палеогеография, Палеоклиматология, Палеоэкология . 403 : 1–15. Бибкод : 2014PPP...403....1G . дои : 10.1016/j.palaeo.2014.03.012 . hdl : 1874/309076 . ISSN 0031-0182 . S2CID 128416509 .
- ^ Банников А.Ф.; Эребакан, И.Г. (1 октября 2023 г.). «Об эволюции некоторых групп морских костистых рыб в кайнозое Тетиса и Паратетиса» . Палеонтологический журнал . 57 (5): 475–490. Бибкод : 2023PalJ...57..475B . дои : 10.1134/S0031030123050015 . ISSN 1555-6174 . S2CID 262543874 .
- ^ Карневале, Джорджио (2008). «Миниатюрный глубоководный топорик (Teleostei: Stomiiformes) из миоцена Италии» . Геологический журнал . 145 (1): 73–84. Бибкод : 2008GeoM..145...73C . дои : 10.1017/S0016756807003937 . ISSN 1469-5081 . S2CID 128491684 .
- ^ Цучия, Юки; Шваржанс, Вернер; Ох, Фумио; Уджихара, Ацуши (6 февраля 2024 г.). «Отолиты глубоководных рыб из нижнемиоценовых формаций Ои и Катада, группа Ичиси, префектура Миэ, центральная Япония» . Историческая биология : 1–24. дои : 10.1080/08912963.2023.2301408 . ISSN 0891-2963 . S2CID 267540826 .
- ^ Карневале, Джорджо; Питч, Теодор В. (12 июня 2009 г.). «Глубоководные удильщики рода Acentrophryne (Teleostei, Ceratioidei, Linophrynidae) в миоцене Калифорнии» . Журнал палеонтологии позвоночных . 29 (2): 372–378. Бибкод : 2009JVPal..29..372C . дои : 10.1671/039.029.0232 . ISSN 0272-4634 . S2CID 86776685 .
- ^ Перейти обратно: а б с д и ж Мойл и Чех 2004 , стр. 585.
- ^ Уортон 2002 , стр. 198.
- ^ Уортон 2002 , стр. 199, 201–202.
- ^ Перейти обратно: а б Уортон 2002 , стр. 199.
- ^ Компаньо, LJV (1984). Акулы мира: аннотированный и иллюстрированный каталог известных на сегодняшний день видов акул . Продовольственная и сельскохозяйственная организация ООН. стр. 14–15. ISBN 92-5-101384-5 .
- ^ Мусилова, Зузана; Кортези, Фабио; Матчинер, Майкл; Дэвис, Уэйн; Патель, Джагдиш; Стиб, Сара; де Бюссероль, Фанни; Мальмстрем, Мартин; Торресен, Оле; Браун, Селеста; Маунтфорд, Джессика; Ханель, Рейнхольд; Стенкамп, Дебора; Якобсен, Кьетилль; Карлтон, Карен; Джентофт, Сиссель; Маршалл, Джастин; Зальцбургер, Вальтер (2019). «Зрение с использованием нескольких различных опсинов палочек у глубоководных рыб» . Наука . 364 (6440). Американская ассоциация содействия развитию науки: 588–592. Бибкод : 2019Sci...364..588M . дои : 10.1126/science.aav4632 . ПМК 6628886 . ПМИД 31073066 .
- ^ Перейти обратно: а б Трухильо и Турман 2011 , стр. 415.
- ^ Трухильо и Турман 2011 , стр. 414–5.
- ^ Рэндалл и Фаррелл 1997 , стр. 217.
- ^ Рэндалл и Фаррелл 1997 , стр. 195.
- ^ Рэндалл и Фаррелл 1997 , стр. 196, 225.
- ^ Мойл и Чех 2004 , стр. 591.
- ^ Перейти обратно: а б Хедрих Р.Л. (1996) «Глубоководные рыбы: эволюция и адаптация в крупнейших жизненных пространствах Земли» , Журнал биологии рыб , 49 (sA): 40–53.
- ^ Мойл и Чех 2004 , стр. 586.
- ^ Перейти обратно: а б с Мойл и Чех 2004 , стр. 571.
- ^ Фрёзе, Райнер; Поли, Дэниел (ред.) (2009). « Argyropelecus aculeatus » в FishBase . Версия за август 2009 года.
- ^ Фрёзе, Райнер; Поли, Дэниел (ред.) (2009). « Dissostichus mawsoni » в FishBase . Версия за август 2009 года.
- ↑ Тайна глубоководных рыб с трубчатыми глазами и прозрачной головой раскрыта ScienceDaily , 24 февраля 2009 г.
- ^ Фрёзе, Райнер; Поли, Дэниел (ред.) (2010). « Гигантура чуни » в FishBase . Версия за октябрь 2010 г.
- ^ Перейти обратно: а б Райан П., «Глубоководные существа: Мезопелагическая зона» , Те Ара – Энциклопедия Новой Зеландии . Обновлено 21 сентября 2007 г.
- ^ Боун и Мур 2008, с. 38.
- ^ Дуглас Э.Л.; Фридл В.А.; Пиквелл Г.В. (1976). «Рыбы в зонах кислородного минимума: особенности оксигенации крови» . Наука . 191 (4230): 957–959. Бибкод : 1976Sci...191..957D . дои : 10.1126/science.1251208 . ПМИД 1251208 .
- ^ Мойл и Чех 2004 , стр. 590.
- ^ Мунц WRA (1976). «О желтых линзах у мезопелагических животных». Морская биологическая ассоциация Великобритании . 56 (4): 963–976. Бибкод : 1976JMBUK..56..963M . дои : 10.1017/S0025315400021019 . S2CID 86353657 .
- ^ Вагнер, HJ; Дуглас, Р.Х.; Фрэнк, ТМ; Робертс, Северо-Запад; Партридж, Дж. К. (2009). «Новый глаз позвоночных, использующий как преломляющую, так и отражающую оптику» . Современная биология . 19 (2): 108–114. Бибкод : 2009CBio...19..108W . дои : 10.1016/j.cub.2008.11.061 . ПМИД 19110427 . S2CID 18680315 .
- ↑ Смит, Л. (8 января 2009 г.). «Рыба с четырьмя глазами может видеть сквозь глубокий морской мрак» . Таймс онлайн . Times Newspapers Ltd. Проверено 14 марта 2009 г.
- ^ Перейти обратно: а б Пакстон и Эшмейер 1998 , стр. 127–8.
- ^ Р. Корнехо; Р. Коппельманн; Т. Саттон. «Разнообразие и экология глубоководных рыб в придонном пограничном слое» . Архивировано из оригинала 1 июня 2013 года . Проверено 2 марта 2013 г.
- ^ Перейти обратно: а б Кенали, CP (2007). «Пересмотр рода Stoplight Loosejaw Malacosteus (Teleostei: Stomiidae: Malacosteinae) с описанием нового вида из умеренного южного полушария и Индийского океана». Копейя . 2007 (4): 886–900. doi : 10.1643/0045-8511(2007)7[886:ROTSLG]2.0.CO;2 . S2CID 1038874 .
- ^ Саттон, Т.Т. (2005). «Трофическая экология глубоководной рыбы Malacosteus niger (Pisces: Stomiidae): загадочная экология питания для создания уникальной зрительной системы?» . Глубоководные исследования. Часть I: Статьи океанографических исследований . 52 (11): 2065–2076. Бибкод : 2005DSRI...52.2065S . дои : 10.1016/j.dsr.2005.06.011 .
- ^ Мойл и Чех 2004 , стр. 336.
- ^ Фрёзе, Райнер; Поли, Дэниел (ред.) (2010). « Анотоптерус фараон » в FishBase . Версия за апрель 2010 года.
- ^ Перейти обратно: а б с д Райан П. «Глубоководные существа: Батипелагическая зона» Те Ара — Энциклопедия Новой Зеландии . Обновлено 21 сентября 2007 г.
- ^ Перейти обратно: а б Фрёзе, Райнер и Дэниел Поли, ред. (2009). «Гоностома» в FishBase . Версия за август 2009 года.
- ^ «Объединяем знания и людей более 10 лет» . Архивировано из оригинала 9 июля 2012 года.
- ^ «Ученые разгадали загадку: три рыбы одинаковы» . 22 января 2009 года . Проверено 22 января 2009 г.
- ^ Фрёзе, Райнер; Поли, Дэниел (ред.) (2010). « Chauliodus sloani » в FishBase . Версия за апрель 2010 года.
- ^ Фрёзе, Райнер; Поли, Дэниел (ред.) (2009). « Anoplogaster cornuta » в FishBase . Версия за август 2009 года.
- ^ Перейти обратно: а б Мойл и Чех 2004 , стр. 594.
- ^ Перейти обратно: а б Мойл и Чех 2004 , стр. 587.
- ^ Маршалл (1984) «Прогенетические тенденции глубоководных рыб», стр. 91-101 в Поттсе Г.В. и Вуттоне Р.Дж. (ред.) (1984) Размножение рыб: стратегии и тактика Рыболовного общества Британских островов.
- ^ Перейти обратно: а б Фудзивара, Ёсихиро; Цучида, Синдзи; Кавато, Масару; Масуда, Котохиро; Сакагути, Сакико Оруи; Садо, Тецуя; Мия, Масаки; Ёсида, Такао (1 июля 2022 г.). «Обнаружение крупнейшей глубоководной эндемической костистой рыбы на глубине более 2000 м посредством сочетания метабаркодирования эДНК и наблюдений с помощью камеры с наживкой» . Границы морской науки . 9 . дои : 10.3389/fmars.2022.945758 . ISSN 2296-7745 .
- ^ Хорн М.Х. (1970). «Плавательный пузырь как ювенильный орган строматоидных рыб» . Бревиора . 359 : 1–9.
- ^ Наблюдение in situ за рыбой-макруридой на высоте 7259 м в Японском желобе: плавучесть плавательного пузыря на экстремальной глубине.
- ^ Джампер Г.Я., Баир Р.К. (1991). «Расположение по обонянию: модель и применение к проблеме спаривания глубоководной рыбы-топорика Argyropelecus hemigymnus ». Американский натуралист . 138 (6): 1431–58. дои : 10.1086/285295 . JSTOR 2462555 . S2CID 84386858 .
- ^ Теодор В. Питч (1975). «Преждевременный половой паразитизм у глубоководных цератиоидных удильщиков Cryptopsaras couesi Gill» . Природа . 256 (5512): 38–40. Бибкод : 1975Natur.256...38P . дои : 10.1038/256038a0 . S2CID 4226567 .
- ^ Джордан, DS (1905). Руководство по изучению рыб . Х. Холт и компания.
- ^ Фрёзе, Райнер; Поли, Дэниел (ред.) (2009). « Хиазмодон нигер » в FishBase . Версия за август 2009 года.
- ^ Скотт, Томас Р.; Пауэлл, Джеймс (2018). Вселенная, какая она есть на самом деле: Земля, Пространство, Материя и Время . Издательство Колумбийского университета. Бибкод : 2018uiri.book.....S . дои : 10.7312/scot18494 . ISBN 978-0-231-54576-1 . S2CID 125357269 .
- ^ Перейти обратно: а б Хочачка и Сомеро 1984 .
- ^ Перейти обратно: а б с Приеде 2017 , стр. 87–138.
- ^ Герринджер, Мэн; Дражен, Дж. К.; Линли, Т.Д.; Саммерс, AP; Джеймисон, Эй Джей; Янси, PH (2017). «Распределение, состав и функции студенистых тканей глубоководных рыб» . Королевское общество открытой науки . 4 (12). дои : 10.1098/rsos.171063 . ПМК 5750012 . ПМИД 29308245 .
- ^ Сомеро, Дж.Н. (1992). «Адаптация к высокому гидростатическому давлению» . Ежегодный обзор физиологии . 54 (1): 557–577. дои : 10.1146/annurev.ph.54.030192.003013 . ISSN 0066-4278 . ПМИД 1314046 .
- ^ Дивайн Дженнифер А.; Бейкер Криста Д.; Хедрих Ричард Л. (2006). «Рыболовство: глубоководные рыбы относятся к категории находящихся под угрозой исчезновения» . Природа . 439 (7072): 29. Бибкод : 2006Natur.439...29D . дои : 10.1038/439029а . ПМИД 16397489 . S2CID 4428618 .
Ссылки
[ редактировать ]- Хочачка, Питер В.; Сомеро, Джордж Н. (1984). Биохимическая адаптация . Издательство Принстонского университета. JSTOR j.ctt7zv9d4 .
- Мойл, ПБ; Чех, Джей-Джей (2004). Рыбы, Введение в ихтиологию (5-е изд.). Бенджамин Каммингс. ISBN 9780131008472 .
- Пакстон, младший; Эшмейер, WN, ред. (1998). Энциклопедия рыб . Сан-Диего: Академическая пресса. ISBN 0125476655 .
- Приеде, Имантс Г., изд. (2017). «Адаптация к глубокому морю». Глубоководные рыбы: биология, разнообразие, экология и рыболовство . Кембридж: Издательство Кембриджского университета. стр. 87–138. дои : 10.1017/9781316018330.004 . ISBN 9781316018330 .
- Рэндалл, Дэвид Дж.; Фаррелл, Энтони Питер (1997). Глубоководные рыбы . Сан-Диего: Академик. ISBN 9780123504401 .
- Трухильо, Алан П.; Турман, Гарольд В. (2011). Основы океанографии (10-е изд.). Бостон: Прентис Холл. ISBN 9780321668127 .
- Уортон, Дэвид (2002). Жизнь на грани: организмы в экстремальных условиях . Кембридж: Издательство Кембриджского университета. п. 198. ИСБН 9780521782128 .
Дальнейшее чтение
[ редактировать ]- Гордон JDM (2001) «Глубоководные рыбы» В: Джон Х. Стил, Стив А. Торп, Карл К. Турекян (ред.) Элементы физической океанографии , страницы 227–233, Academic Press. ISBN 9780123757241 .
- Хоар В.С., Рэндалл DJ и Фаррелл А.П. (ред.) (1997) Глубоководные рыбы , Academic Press. ISBN 9780080585406 .
- Шоттон, Росс (1995) «Глубоководное рыболовство» В: Обзор состояния мировых ресурсов морского рыболовства , Технический документ ФАО по рыболовству 457, ФАО, Рим. ISBN 92-5-105267-0 .
- Тандстад М., Шоттон Р., Сандерс Дж. и Кароччи Ф. (2011) «Глубоководное рыболовство». Архивировано 3 марта 2016 г. в Wayback Machine. В: Обзор состояния мировых ресурсов морского рыболовства , страницы 265–278, ФАО. Технический документ по рыболовству 569, ФАО, Рим. ISBN 978-92-5-107023-9 .
Внешние ссылки
[ редактировать ]| Внешние видео | |
|---|---|
- Глубоководный Бестиарий
- Фотогалерея: Глубоководные существа
- Ускоренный курс глубоководной рыбалки – статьи, факты и изображения глубоководных животных


