Морские беспозвоночные

Морские беспозвоночные — это беспозвоночные , обитающие в морской среде обитания . Беспозвоночные — это общий термин , который включает в себя всех животных, кроме позвоночных, представителей типа хордовых . У беспозвоночных отсутствует позвоночный столб , а у некоторых развился панцирь или твердый экзоскелет . Как на суше, так и в воздухе, морские беспозвоночные имеют большое разнообразие строения тела и подразделяются на более чем 30 типов . Они составляют большую часть макроскопической жизни в океанах.
Эволюция [ править ]
| Часть серии обзоров по |
| Морская жизнь |
|---|
 |
Самыми ранними животными были морские беспозвоночные , то есть позвоночные появились позже. Животные – многоклеточные эукариоты , [примечание 1] и отличаются от растений, водорослей и грибов отсутствием клеточных стенок . [1] Морские беспозвоночные — это животные, населяющие морскую среду, помимо позвоночных представителей типа хордовых ; у беспозвоночных отсутствует позвоночный столб . Некоторые из них развили панцирь или твердый экзоскелет .
Самые ранние животные могут принадлежать к роду Dickinsonia . [2] 571–539 миллионов лет назад. [3] Отдельные дикинсонии обычно напоминают двусторонне-симметричный ребристый овал. Они продолжали расти, пока не покрылись осадком или не погибли иным образом. [4] и провели большую часть своей жизни, твердо привязав свои тела к осадку. [5] Их таксономическое сходство в настоящее время неизвестно, но способ их роста соответствует билатеральному сходству. [6]
Помимо дикинсонии , самыми ранними широко признанными окаменелостями животных являются довольно современные на вид книдарии (группа, в которую входят медузы , морские анемоны и гидры ), возможно, датируемые примерно 580 млн лет назад. [7] , Биота Эдиакары процветавшая последние 40 миллионов лет до начала кембрия , [8] были первыми животными длиной более нескольких сантиметров. Подобно Дикинсонии , многие из них были плоскими, со «стеганым» внешним видом, и казались настолько странными, что появилось предложение отнести их к отдельному царству , Вендозоа . [9] Другие, однако, были интерпретированы как ранние моллюски ( Kimberella [10] [11] ), иглокожие ( Arkarua [12] ), и членистоногих ( Сприггина , [13] Парванкорин [14] ). До сих пор ведутся споры о классификации этих образцов, главным образом потому, что диагностические особенности, которые позволяют систематикам классифицировать более поздние организмы, такие как сходство с живыми организмами, обычно отсутствуют у эдиакарцев. Однако мало кто сомневается в том, что кимберелла была, по крайней мере, трехблобластным двусторонним животным, другими словами, животным, значительно более сложным, чем книдарии. [15]
Небольшая ракушечная фауна представляет собой очень смешанную коллекцию окаменелостей, найденных между позднем эдиакарским и средним кембрийским периодами. Самый ранний, Cloudina , демонстрирует признаки успешной защиты от хищников и может указывать на начало эволюционной гонки вооружений . Некоторые крошечные раннекембрийские раковины почти наверняка принадлежали моллюскам, а владельцы некоторых «броневых пластин» — Halkieria и Microdictyon были найдены более полные экземпляры — в конечном итоге были идентифицированы, когда в кембрийских лагерштеттенах , сохранившие мягкотелых животных. [16]
 Дикинсония может быть самым ранним животным. Они появляются в летописи окаменелостей от 571 до 541 миллиона лет назад.
Дикинсония может быть самым ранним животным. Они появляются в летописи окаменелостей от 571 до 541 миллиона лет назад. Кимберелла , ранний моллюск, важный для понимания кембрийского взрыва . Беспозвоночные сгруппированы в разные типы ( планы тела ).
Кимберелла , ранний моллюск, важный для понимания кембрийского взрыва . Беспозвоночные сгруппированы в разные типы ( планы тела ).![Опабиния, вымершее членистоногое стеблевой группы, появилось в среднем кембрии[17]: 124–136.](//upload.wikimedia.org/wikipedia/commons/thumb/2/21/20191108_Opabinia_regalis.png/642px-20191108_Opabinia_regalis.png)


![Опабиния, вымершее членистоногое стеблевой группы, появилось в среднем кембрии[17]: 124–136.](http://upload.wikimedia.org/wikipedia/commons/thumb/2/21/20191108_Opabinia_regalis.png/642px-20191108_Opabinia_regalis.png)
В 1970-х годах уже велись споры о том, было ли появление современных типов «взрывным» или постепенным, но скрытым нехваткой окаменелостей докембрийских животных. [16] Повторный анализ окаменелостей из лагерштетте в сланцах Берджесс повысил интерес к этому вопросу, когда выявил животных, таких как Opabinia , которые не вписывались ни в один известный тип . В то время это было интерпретировано как свидетельство того, что современные типы очень быстро развились в результате кембрийского взрыва и что «странные чудеса» сланцев Бёрджесс показали, что ранний кембрий был уникальным экспериментальным периодом эволюции животных. [17] Более поздние открытия подобных животных и развитие новых теоретических подходов привели к выводу, что многие из «странных чудес» были эволюционными «тетями» или «кузенами» современных групп. [18] — например, что опабиния принадлежала к лобоподам , группе, в которую входят предки членистоногих, и что она, возможно, была тесно связана с современными тихоходками . [19] Тем не менее, до сих пор ведется много споров о том, действительно ли кембрийский взрыв был взрывным, и если да, то как и почему он произошел и почему он кажется уникальным в истории животных. [20]
Классификация [ править ]

Беспозвоночные сгруппированы в разные типы . Неформально типы можно рассматривать как способ группировки организмов в соответствии со строением их тела . [21] [22] : 33 План тела относится к проекту, который описывает форму или морфологию организма, такую как его симметрия , сегментация и расположение его придатков . Идея строения тела возникла у позвоночных , которые были сгруппированы в один тип. Но строение тела позвоночных — лишь одно из многих, а беспозвоночные состоят из многих типов или строений тела. Историю открытия строения тела можно рассматривать как движение от мировоззрения, сосредоточенного на позвоночных, к рассмотрению позвоночных как одного из многих строений тела. первопроходцев Среди зоологов- Линней выделил два строения тела за пределами позвоночных; Кювье назвал троих; у Геккеля было четыре, а у Протиста еще восемь, всего двенадцать. Для сравнения, число типов, признанных современными зоологами, возросло до 35 . [22]
Исторически считалось, что структуры тела быстро изменились во время кембрийского взрыва . [23] но более детальное понимание эволюции животных предполагает постепенное развитие строения тела на протяжении раннего палеозоя и далее. [24] В более общем смысле тип можно определить двумя способами: как описано выше, как группу организмов с определенной степенью морфологического сходства или сходства в развитии ( фенетическое определение) или как группу организмов с определенной степенью эволюционного родства ( филогенетическое определение). определение). [24]
Как на суше, так и в воздухе, беспозвоночные составляют подавляющее большинство всей макроскопической жизни, поскольку позвоночные составляют подтип одного из более чем 30 известных типов животных, что делает этот термин практически бессмысленным для таксономических целей. Морская жизнь беспозвоночных включает следующие группы, некоторые из которых относятся к типам:


- Акоэла , среди наиболее примитивных двусторонних животных ;
- Annelida , ( полихеты и морские пиявки );
- Брахиоподы , морские животные, имеющие твердые «клапаны» (раковины) на верхней и нижней поверхности;
- Мшанки , также известные как моховые животные или морские маты;
- Chaetognatha , широко известные как черви-стрелы, представляют собой тип хищных морских червей, которые являются основным компонентом планктона;
- Цефалохордовые, представленные в современных океанах ланцетниками ( также известными как амфиоксус);
- Cnidaria , например , медузы , морские анемоны и кораллы ;
- Ракообразные , включая омаров , крабов , креветок , раков , ракообразных , раков-отшельников , креветок-богомолов и веслоногих ракообразных ;
- Гребневики , также известные как гребневики, самые крупные животные, плавающие при помощи ресничек;
- Иглокожие , в том числе морские звезды , хрупкие звезды , морские ежи , песчанки , морские огурцы , криноидеи и морские маргаритки ;
- Эчиура , также известная как ложечные черви;
- Gnathostomulids , тонкие или нитевидные черви, с прозрачным телом, обитающие в песке и иле под мелкими прибрежными водами;
- Gastrotricha , часто называемая волосатой спинкой, встречается в основном интерстициально между частицами осадка;
- Hemichordata , включает желудевых червей , одиночные червеобразные организмы;
- Камптозоа , сидячие водные животные кубковидной формы, с относительно длинными стеблями и «короной» из твердых щупалец, называемые также Entoprocta;
- Киноринчи , сегментированные, без конечностей животные, широко распространенные в иле или песке на всех глубинах, называемые также грязевыми драконами;
- Loricifera , очень мелкие или микроскопические животные, обитающие в морских отложениях, обнаруженные только в 1983 году;
- Моллюски , включая моллюсков , кальмаров , осьминогов , трубачей , наутилусов , каракатиц , голожаберников , морских гребешков , морских улиток , аплакофору , Caudofoveata , моноплакофору , полиплакофору и скафоподу ;
- Myzostomida , таксономическая группа мелких морских червей, паразитирующих на криноидеях или «морских лилиях»;
- Nemertinea , также известная как «ленточные черви» или «хоботковые черви»;
- Orthonectida , небольшой тип малоизвестных паразитов морских беспозвоночных, которые относятся к числу простейших многоклеточных организмов;
- Phoronida , тип морских животных, которые фильтруют пищу с помощью лофофора («короны» из щупалец) и строят вертикальные хитиновые трубки для поддержки и защиты своих мягких тел;
- Плакозоа — маленькие, уплощенные, многоклеточные животные диаметром около 1 миллиметра и простейшие по строению. Они не имеют правильного очертания, хотя нижняя поверхность несколько вогнута, а верхняя всегда приплюснута;
- Porifera (губки), многоклеточные организмы, тела которых полны пор и каналов, позволяющих воде циркулировать через них;
- Priapulida , или черви-пенисы, представляют собой тип морских червей, обитающих в морской грязи. Они названы в честь своего растяжимого колючего хоботка, который у некоторых видов может иметь форму человеческого пениса;
- Пикногониды , также называемые морскими пауками, не имеют отношения к паукам или даже к паукообразным, на которых они похожи;
- Sipunculida , также называемые арахисовыми червями, представляют собой группу, содержащую 144–320 видов (по разным оценкам) двусторонне-симметричных несегментированных морских червей;
- Туниката , также известная как асцидии или морская свинина, представляет собой фильтраторы, прикрепленные к камням или аналогичным подходящим поверхностям на дне океана;
- Некоторые плоские черви классов Turbellaria и Monogenea ;
- Xenoturbella — род двусторонних животных, включающий только два вида морских червеобразных;
- Xiphosura включает большое количество вымерших линий и только четыре современных вида семейства Limulidae, к которым относятся мечехвосты .
Всего членистоногих насчитывается около 1 113 000 описанных современных видов, моллюсков - около 85 000 и хордовых - около 52 000. [25]
Морские губки [ править ]

Губки — животные типа Porifera (современное латинское слово « несущие поры »). [26] ). Это многоклеточные организмы, тела которых полны пор и каналов, позволяющих воде циркулировать через них, и состоят из желеобразного мезоила, зажатого между двумя тонкими слоями клеток . У них есть неспециализированные клетки, которые могут трансформироваться в другие типы и при этом часто мигрируют между основными клеточными слоями и мезохилом. У губок нет нервной , пищеварительной и кровеносной систем . Вместо этого большинство полагаются на поддержание постоянного потока воды через свое тело для получения пищи и кислорода, а также для удаления отходов.
Губки похожи на других животных тем, что они многоклеточные , гетеротрофные , лишены клеточных стенок и производят сперматозоиды . В отличие от других животных, у них отсутствуют настоящие ткани и органы , а также отсутствует симметрия тела . Формы их тел приспособлены для максимальной эффективности потока воды через центральную полость, где она откладывает питательные вещества, и выходит через отверстие, называемое оскулумом . Многие губки имеют внутренний скелет из спонгина и/или спикулы из карбоната кальция или диоксида кремния . Все губки — сидячие водные животные. Хотя существуют пресноводные виды, подавляющее большинство из них являются морскими (солеными) видами, варьирующимися от приливных зон до глубин, превышающих 8800 м (5,5 миль).
В то время как большинство из примерно 5 000–10 000 известных видов питаются бактериями и другими пищевыми частицами в воде, некоторые из них являются хозяевами фотосинтезирующих микроорганизмов в качестве эндосимбионтов , и эти союзы часто производят больше пищи и кислорода, чем потребляют. Некоторые виды губок, обитающие в условиях скудной пищи, превратились в хищников , питающихся в основном мелкими ракообразными . [27]
 Биоразнообразие губок. На этой фотографии четыре вида губок.
Биоразнообразие губок. На этой фотографии четыре вида губок.
 Цветочная корзина Венеры на глубине 2572 метра
Цветочная корзина Венеры на глубине 2572 метра







Линней ошибочно отнес губки к растениям отряда Водоросли . [29] Долгое время после этого губки были отнесены к отдельному подцарству Паразоа (то есть рядом с животными ). [30] Сейчас их классифицируют как парафилетический тип , из которого произошли высшие животные. [31]
Морские книдарии [ править ]

Книдарии (по-гречески крапива ) отличаются наличием стрекательных клеток — специализированных клеток, которые они используют в основном для захвата добычи. Книдарии включают кораллы , морские анемоны , медузы и гидрозои . Они образуют тип, содержащий более 10 000 [32] виды животных , обитающие исключительно в водной (преимущественно морской) среде. Их тела состоят из мезоглеи , неживого желеобразного вещества, зажатого между двумя слоями эпителия , толщиной в основном в одну клетку . У них есть две основные формы тела: плавающие медузы и сидячие полипы , оба из которых радиально симметричны, а рты окружены щупальцами, несущими книдоциты. Обе формы имеют одно отверстие и полость тела, которые используются для пищеварения и дыхания .
Ископаемые книдарии были обнаружены в горных породах, образовавшихся около 580 миллионов лет назад . Окаменелости книдарий, не образующих минерализованных структур, редки. В настоящее время ученые считают, что книдарии, гребневики и билатерии более тесно связаны с известковыми губками, чем с другими губками , и что антозойные особи являются эволюционными «тетями» или «сестрами» других книдарий и наиболее тесно связаны с билатериями.
Книдарии — простейшие животные, у которых клетки организованы в ткани. [33] Звездочка актинии используется в качестве модельного организма в исследованиях. [34] За ним легко ухаживать в лаборатории, и был разработан протокол , позволяющий получать большое количество эмбрионов ежедневно. [35] Существует поразительная степень сходства в сохранении и сложности последовательностей генов между морскими анемонами и позвоночными. [35] В частности, у анемона присутствуют гены, участвующие в формировании головы у позвоночных. [36] [37]
 Морские анемоны распространены в приливных лужах.
Морские анемоны распространены в приливных лужах. Их щупальца жалят и парализуют мелкую рыбу.
Их щупальца жалят и парализуют мелкую рыбу.
 Если остров опускается под воду, рост кораллов может идти в ногу с подъемом воды и образовать атолл.
Если остров опускается под воду, рост кораллов может идти в ногу с подъемом воды и образовать атолл.







![Биолюминесцентный морской орех имеет временный анус[39], который образуется только тогда, когда ему необходимо испражняться.](//upload.wikimedia.org/wikipedia/commons/thumb/f/f0/Sea_walnut%2C_Boston_Aquarium_%28cropped%29.jpg/265px-Sea_walnut%2C_Boston_Aquarium_%28cropped%29.jpg) Биолюминесцентный . морской орех имеет временный анус [39] который образуется только тогда, когда ему нужно испражняться
Биолюминесцентный . морской орех имеет временный анус [39] который образуется только тогда, когда ему нужно испражняться![Turritopsis dohrnii достигает биологического бессмертия, перенося свои клетки обратно в детство[40][41]](//upload.wikimedia.org/wikipedia/commons/b/b0/Turritopsis_dohrnii_%28cropped%29.jpg) Turritopsis dohrnii достигает биологического бессмертия , перенеся свои клетки обратно в детство [40] [41]
Turritopsis dohrnii достигает биологического бессмертия , перенеся свои клетки обратно в детство [40] [41]



![Биолюминесцентный морской орех имеет временный анус[39], который образуется только тогда, когда ему необходимо испражняться.](http://upload.wikimedia.org/wikipedia/commons/thumb/f/f0/Sea_walnut%2C_Boston_Aquarium_%28cropped%29.jpg/265px-Sea_walnut%2C_Boston_Aquarium_%28cropped%29.jpg)
![Turritopsis dohrnii достигает биологического бессмертия, перенося свои клетки обратно в детство[40][41]](http://upload.wikimedia.org/wikipedia/commons/b/b0/Turritopsis_dohrnii_%28cropped%29.jpg)
Морские черви [ править ]
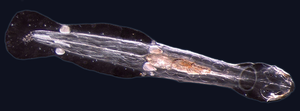
Черви (древнеанглийское слово « змея ») обычно имеют длинные цилиндрические трубчатые тела и не имеют конечностей . Морские черви различаются по размеру от микроскопических до более 1 метра (3,3 фута) в длину для некоторых морских многощетинковых червей ( щетинковых червей ). [42] и до 58 метров (190 футов) для морского немертинского червя ( шнурочного червя ). [43] Некоторые морские черви занимают небольшое количество паразитических ниш, живя внутри тел других животных, в то время как другие живут более свободно в морской среде или роются под землей.
Различные группы морских червей связаны лишь отдаленно, поэтому они встречаются в нескольких разных типах, таких как Annelida (сегментированные черви), Chaetognatha (черви-стрелы), Hemichordata и Phoronida (подковообразные черви). Многие из этих червей имеют специальные щупальца, используемые для обмена кислорода и углекислого газа, а также могут использоваться для размножения. Некоторые морские черви являются трубчатыми , например, гигантский трубчатый червь , который живет в водах вблизи подводных вулканов и может выдерживать температуру до 90 градусов по Цельсию .
Platyhelminthes (плоские черви) образуют другой тип червей, который включает класс Cestoda паразитических ленточных червей . Морской ленточный червь Polygonoporus giganticus , обитающий в кишечнике кашалотов , может вырасти до более чем 30 м (100 футов). [44] [45]
Нематоды (круглые черви) представляют собой еще один тип червей с трубчатой пищеварительной системой и отверстиями на обоих концах. [46] [47] Описано более 25 000 видов нематод. [48] [49] из них более половины являются паразитическими. По оценкам, еще миллион остается неописанным. [50] Они повсеместно распространены в морской, пресноводной и наземной среде, где часто превосходят по численности других животных как по численности, так и по численности видов. Земли Они встречаются во всех частях литосферы , от вершин гор до дна океанических впадин . [51] По подсчетам они составляют 90% всех животных на дне океана . [52] Их численное преобладание, часто превышающее миллион особей на квадратный метр и составляющее около 80% всех отдельных животных на Земле, разнообразие жизненных циклов и присутствие на различных трофических уровнях указывают на важную роль во многих экосистемах. [53]
 Гигантские трубчатые черви скапливаются вокруг гидротермальных источников
Гигантские трубчатые черви скапливаются вокруг гидротермальных источников
 Нематоды — это вездесущие псевдоцеломаты , которые могут паразитировать на морских растениях и животных.
Нематоды — это вездесущие псевдоцеломаты , которые могут паразитировать на морских растениях и животных. Мотыль обычно встречается на дне мелких морских вод.
Мотыль обычно встречается на дне мелких морских вод. Боббитовые черви — хищники, устраивающие засады и обитающие на морском дне.
Боббитовые черви — хищники, устраивающие засады и обитающие на морском дне.





Иглокожие [ править ]

Иглокожие (по-гречески « шипистая кожа ») — это тип, в состав которого входят только морские беспозвоночные. Взрослые особи узнаваемы по радиальной симметрии (обычно пятиточечной) и включают морские звезды , морские ежи , песчаные доллары и морские огурцы , а также морские лилии . [ нужна ссылка ] Иглокожие встречаются на любой глубине океана, от приливной зоны до абиссальной зоны . Тип насчитывает около 7000 живых видов . [54] что делает его второй по величине группой вторичноротых (супертип) после хордовых (которые включают позвоночных , таких как птицы , рыбы , млекопитающие и рептилии ).
Иглокожие уникальны среди животных, поскольку имеют двустороннюю симметрию на личиночной стадии и пятикратную симметрию ( пентамеризм , особый тип радиальной симметрии) во взрослом возрасте. [55]
Иглокожие важны как с биологической, так и с геологической точки зрения. есть несколько других группировок, которые были бы так многочисленны С биологической точки зрения, в биотической пустыне глубокого моря , а также в более мелких океанах . Большинство иглокожих способны к регенерации тканей, органов, конечностей, а также размножаются бесполым путем ; в некоторых случаях они могут подвергнуться полной регенерации из одной конечности. С геологической точки зрения ценность иглокожих заключается в их окостеневших скелетах , которые вносят основной вклад во многие известняковые образования и могут дать ценную информацию о геологической среде. Они были наиболее часто используемыми видами в регенеративных исследованиях в 19 и 20 веках. Кроме того, некоторые ученые считают, что радиация иглокожих была ответственна за мезозойскую морскую революцию .
 Иглокожие буквально означает «колючая кожа», как этот арбузный морской еж. иллюстрирует
Иглокожие буквально означает «колючая кожа», как этот арбузный морской еж. иллюстрирует Морские огурцы фильтруют питание планктона и взвешенных веществ.
Морские огурцы фильтруют питание планктона и взвешенных веществ. Бентопелагические трепанги могут отрываться от морского дна и подниматься вверх по толще воды на высоту до 1000 м (3300 футов).
Бентопелагические трепанги могут отрываться от морского дна и подниматься вверх по толще воды на высоту до 1000 м (3300 футов).![Охряная морская звезда была первым краеугольным хищником, которого удалось изучить. Они ограничивают численность мидий, которые могут уничтожить приливные сообщества.[56]](//upload.wikimedia.org/wikipedia/commons/thumb/8/89/Ochre_sea_star.jpg/280px-Ochre_sea_star.jpg) Охряная морская звезда была первым краеугольным хищником, которого удалось изучить. Они ограничивают численность мидий , которые могут уничтожить приливные сообщества. [56]
Охряная морская звезда была первым краеугольным хищником, которого удалось изучить. Они ограничивают численность мидий , которые могут уничтожить приливные сообщества. [56] Красочные морские лилии на мелководье
Красочные морские лилии на мелководье



![Охряная морская звезда была первым краеугольным хищником, которого удалось изучить. Они ограничивают численность мидий, которые могут уничтожить приливные сообщества.[56]](http://upload.wikimedia.org/wikipedia/commons/thumb/8/89/Ochre_sea_star.jpg/280px-Ochre_sea_star.jpg)

Помимо трудно классифицируемого аркаруа ( докембрийского животного с иглокожей пятичленной радиальной симметрией), первые окончательные представители этого типа появились примерно в начале кембрия .
Морские моллюски [ править ]

Моллюски (лат. « мягкие ») образуют тип , насчитывающий около 85 000 современных признанных видов . [57] По количеству видов они являются крупнейшим морским типом, составляющим около 23% всех названных морских организмов . [58] Моллюски имеют более разнообразные формы, чем другие типы беспозвоночных. Они весьма разнообразны не только по размеру и анатомическому строению, но и по поведению и среде обитания. Большинство видов все еще обитает в океанах, от морских побережий до абиссальной зоны , но некоторые составляют значительную часть пресноводной фауны и наземных экосистем .
Тип моллюсков делится на 9—10 таксономических классов , два из которых вымерли . К этим классам относятся брюхоногие моллюски , двустворчатые моллюски и головоногие моллюски , а также другие менее известные, но самобытные классы. Брюхоногих моллюсков с защитным панцирем называют улитками ( морскими улитками ), тогда как брюхоногих моллюсков без защитного панциря называют слизнями ( морскими слизняками ). Брюхоногие моллюски на сегодняшний день являются самыми многочисленными моллюсками с точки зрения классифицированных видов, составляя 80% от общего числа. [25] К двустворчатым моллюскам относятся моллюски , устрицы , моллюски , мидии , морские гребешки и многие другие семейства . Существует около 8000 видов морских двустворчатых моллюсков (включая солоноватоводные и устьевые виды) и около 1200 пресноводных видов. Головоногие включают осьминогов , кальмаров и каракатиц . Они обитают во всех океанах и в неврологическом отношении являются наиболее развитыми из беспозвоночных. [59] Выявлено около 800 ныне живущих видов морских головоногих моллюсков. [60] около 11 000 вымерших таксонов . и описано [61] Полностью пресноводных головоногих не существует. [62]
![Колоссальный кальмар, самый крупный из всех беспозвоночных[63]](//upload.wikimedia.org/wikipedia/commons/thumb/3/37/Calmarcolossal.jpg/147px-Calmarcolossal.jpg) Колоссальный кальмар , самый крупный из всех беспозвоночных [63]
Колоссальный кальмар , самый крупный из всех беспозвоночных [63]![Наутилус — это живое ископаемое, мало изменившееся с тех пор, как он появился 500 миллионов лет назад как одно из первых головоногих моллюсков.[64][65][66]](//upload.wikimedia.org/wikipedia/commons/thumb/d/d5/Nautilus_Palau.JPG/280px-Nautilus_Palau.JPG) Наутилус мало — это живое ископаемое, изменившееся с тех пор, как он появился 500 миллионов лет назад как одно из первых головоногих моллюсков . [64] [65] [66]
Наутилус мало — это живое ископаемое, изменившееся с тех пор, как он появился 500 миллионов лет назад как одно из первых головоногих моллюсков . [64] [65] [66] Морские брюхоногие моллюски — это морские улитки или морские слизни . Этот голожаберный моллюск — морской слизень.
Морские брюхоногие моллюски — это морские улитки или морские слизни . Этот голожаберный моллюск — морской слизень. Морская улитка Syrinx aruanus имеет самый большой панцирь среди всех ныне живущих брюхоногих моллюсков.
Морская улитка Syrinx aruanus имеет самый большой панцирь среди всех ныне живущих брюхоногих моллюсков. У моллюсков обычно есть глаза. Окаймляющий край мантии гребешка , двустворчатого моллюска , может быть свыше 100 простых глаз .
У моллюсков обычно есть глаза. Окаймляющий край мантии гребешка , двустворчатого моллюска , может быть свыше 100 простых глаз . Обыкновенная мидия , еще один двустворчатый моллюск
Обыкновенная мидия , еще один двустворчатый моллюск
![Колоссальный кальмар, самый крупный из всех беспозвоночных[63]](http://upload.wikimedia.org/wikipedia/commons/thumb/3/37/Calmarcolossal.jpg/147px-Calmarcolossal.jpg)
![Наутилус — это живое ископаемое, мало изменившееся с тех пор, как он появился 500 миллионов лет назад как одно из первых головоногих моллюсков.[64][65][66]](http://upload.wikimedia.org/wikipedia/commons/thumb/d/d5/Nautilus_Palau.JPG/280px-Nautilus_Palau.JPG)




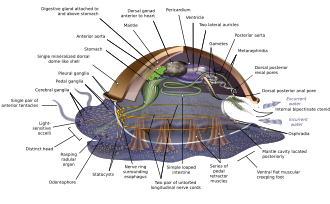
Моллюски имеют настолько разнообразные формы, что многие учебники основывают свои описания анатомии моллюсков на обобщенных или гипотетических предках моллюсков . Этот генерализованный моллюск несегментирован и двусторонне-симметричен , нижняя сторона состоит из одной мускулистой ножки . [67] [68] : 484–628 Помимо этого, у него есть еще три ключевые особенности. Во-первых, у него есть мускулистый покров, называемый мантией, покрывающий его внутренности и содержащий значительную полость, используемую для дыхания и выделения . Раковина , выделяемая мантией, покрывает верхнюю поверхность. [68] Во-вторых (кроме двустворчатых моллюсков) у них есть грубый язык, называемый радулой, используемый для питания. В-третьих, у него есть нервная система , включая сложную пищеварительную систему, использующую микроскопические волоски с мускульной силой, называемые ресничками, для выделения слизи . Генерализованный моллюск имеет две парные нервные цепочки (у двустворчатых моллюсков — три). Мозг у видов , у которых он есть, окружает пищевод . У большинства моллюсков есть глаза , и у всех есть датчики, реагирующие на химические вещества, вибрацию и прикосновение. Простейший тип репродуктивной системы моллюсков основан на внешнем оплодотворении , но встречаются и более сложные варианты. Все они производят яйца , из которых могут выйти трохофоров личинки , более сложные личинки велигеров или миниатюрные взрослые особи. Изображение очень похоже на современных моноплакофоров , и некоторые предполагают, что они могут напоминать очень ранних моллюсков. [67] : 284–291 [67] : 298–300 [69] [70]
Существуют убедительные доказательства появления морских брюхоногих, головоногих и двустворчатых моллюсков в кембрийский период от 538,8 до 485,4 миллионов лет назад . Однако эволюционная история как возникновения моллюсков от предков Lophotrochozoa , так и их диверсификации в известные живые и ископаемые формы до сих пор остается предметом бурных дискуссий среди ученых.
Морские членистоногие [ править ]

Членистоногие (по-гречески « членистые лапы ») имеют экзоскелет (внешний скелет ), сегментированное тело и суставные придатки (парные придатки). Они образуют тип , который включает насекомых , паукообразных , многоножек и ракообразных . Членистоногие характеризуются членистыми конечностями и кутикулой из хитина , часто минерализованного карбонатом кальция . членистоногих Строение тела состоит из сегментов , каждый из которых имеет пару придатков . Жесткая кутикула тормозит рост, поэтому членистоногие периодически заменяют ее путем линьки . Их универсальность позволила им стать самыми богатыми видами среди всех экологических гильдий в большинстве сред.
Размер морских членистоногих варьируется от микроскопического ракообразного Stygotantulus до японского краба-паука . Первичная внутренняя полость членистоногих представляет собой гемоцель , в котором расположены их внутренние органы и через который циркулирует их гемолимфа — аналог крови ; у них открытая система кровообращения . Как и их внешний вид, внутренние органы членистоногих обычно состоят из повторяющихся сегментов. Их нервная система имеет «лестничную форму»: парные вентральные нервные шнуры проходят через все сегменты и образуют парные ганглии в каждом сегменте . Их головы образуются в результате слияния различного числа сегментов, а мозг образуется в результате слияния ганглиев этих сегментов и окружает пищевод . Дыхательная подтипа и выделительная системы членистоногих различаются в зависимости как от окружающей среды, так и от , к которому они принадлежат.
Их зрение основано на различных комбинациях сложных глаз с пигментными ямками и глазков : у большинства видов глазки могут определять только направление, откуда исходит свет, и сложные глаза являются основным источником информации, но основными глазами пауков являются глазки. которые могут формировать изображения и, в некоторых случаях, поворачиваться, чтобы отслеживать добычу. Членистоногие также обладают широким спектром химических и механических сенсоров, в основном основанных на модификациях множества щетинок (щетинок), которые выступают через их кутикулу. Способы размножения и развития членистоногих разнообразны; все наземные виды используют внутреннее оплодотворение , но часто это происходит путем непрямой передачи спермы через придаток или землю, а не путем прямой инъекции. Все морские виды откладывают яйца и используют либо внутреннее, либо внешнее оплодотворение . Детеныши членистоногих варьируются от миниатюрных взрослых особей до личинок, у которых нет членистых конечностей, и в конечном итоге претерпевают полную метаморфозу , образуя взрослую форму.
![Трилобиты, ныне вымершие, бродили по океанам 270 миллионов лет.[71]](//upload.wikimedia.org/wikipedia/commons/thumb/0/08/Olenoides_typicalis.jpg/143px-Olenoides_typicalis.jpg)
 Мечехвостик , живое ископаемое членистоногое, жившее 450 миллионов лет назад.
Мечехвостик , живое ископаемое членистоногое, жившее 450 миллионов лет назад.
![Трилобиты, ныне вымершие, бродили по океанам 270 миллионов лет.[71]](http://upload.wikimedia.org/wikipedia/commons/thumb/0/08/Olenoides_typicalis.jpg/143px-Olenoides_typicalis.jpg)

- Ракообразные
 Многие ракообразные очень маленькие, как этот крошечный амфипод океана. , и составляют значительную часть зоопланктона
Многие ракообразные очень маленькие, как этот крошечный амфипод океана. , и составляют значительную часть зоопланктона Полосатая креветка-чистильщик — ракообразное, распространенное в тропиках.
Полосатая креветка-чистильщик — ракообразное, распространенное в тропиках.![Тасманский гигантский краб долгоживущий и медленнорастущий, что делает его уязвимым для чрезмерного вылова рыбы.[72]](//upload.wikimedia.org/wikipedia/commons/thumb/2/2c/J_J_Wild_Pseudocarcinus_cropped.jpg/330px-J_J_Wild_Pseudocarcinus_cropped.jpg) Тасманский гигантский краб долгоживущий и медленнорастущий, что делает его уязвимым для чрезмерного вылова рыбы. [72]
Тасманский гигантский краб долгоживущий и медленнорастущий, что делает его уязвимым для чрезмерного вылова рыбы. [72] Японский краб-паук имеет самый длинный размах ног среди членистоногих.
Японский краб-паук имеет самый длинный размах ног среди членистоногих.


![Тасманский гигантский краб долгоживущий и медленнорастущий, что делает его уязвимым для чрезмерного вылова рыбы.[72]](http://upload.wikimedia.org/wikipedia/commons/thumb/2/2c/J_J_Wild_Pseudocarcinus_cropped.jpg/330px-J_J_Wild_Pseudocarcinus_cropped.jpg)

Эволюционное происхождение членистоногих восходит к кембрийскому периоду. Группа обычно считается монофилетической , и многие анализы подтверждают размещение членистоногих с циклонейралиями (или их составляющими кладами) в супертипе Ecdysozoa . Однако в целом базальные взаимоотношения Metazoa еще не вполне выяснены. Точно так же до сих пор активно обсуждаются взаимоотношения между различными группами членистоногих.
Другие типы [ править ]
- Тихоходка , Лобоподия , ( Onychophora )
- Бесчерепные (беспозвоночные) хордовые: Cephalochordate , Tunicata и Haikouella . Эти беспозвоночные являются близкими родственниками позвоночных.
- Бесчерепные хордовые — близкие родственники позвоночных.
 Ланцетник головохордового , небольшая полупрозрачная рыба, похожая на , является ближайшим из ныне живущих беспозвоночных родственников позвоночных. [73] [74]
Ланцетник головохордового , небольшая полупрозрачная рыба, похожая на , является ближайшим из ныне живущих беспозвоночных родственников позвоночных. [73] [74] Морские асцидии флуоресцентного цвета, Rhopalaea crassa . Оболочники могут дать ключ к разгадке происхождения позвоночных (и, следовательно, человека). [75]
Морские асцидии флуоресцентного цвета, Rhopalaea crassa . Оболочники могут дать ключ к разгадке происхождения позвоночных (и, следовательно, человека). [75] Сальповая цепь
Сальповая цепь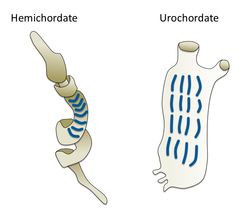 Жаберные щели желудевого червя (слева) и оболочника (справа)
Жаберные щели желудевого червя (слева) и оболочника (справа)




Минералы морской воды [ править ]
Существует ряд морских беспозвоночных, которые используют минералы, присутствующие в море в таких ничтожных количествах, что их невозможно было обнаружить до появления спектроскопии . Ванадий концентрируется некоторыми оболочниками для использования в клетках крови до уровня, в десять миллионов раз превышающего уровень окружающей морской воды. Другие оболочки аналогичным образом концентрируют ниобий и тантал . [67] : 947 Омары содержат медь в своем дыхательном пигменте гемоцианине , несмотря на то, что доля этого металла в морской воде незначительна. [67] : 638 Хотя эти элементы присутствуют в океане в огромных количествах, их добыча человеком неэкономична. [76]
Стеклянные модели [ править ]
- Морские беспозвоночные из стекла Блашка, 1863–1880 гг.














См. также [ править ]
- Морская жизнь
- Морская биология
- Морские позвоночные
- Список видов беспозвоночных морских аквариумов
- Виваксия
Примечания [ править ]
- ^ Миксозоа считались исключением, но теперь считаются сильно модифицированными представителями Cnidaria . Хименес-Гури, Ева; Филипп, Эрве; Окамура, Бет; Холланд, Питер WH (6 июля 2007 г.). « Будденброкия — книдарийный червь». Наука . 317 (5834): 116–118. Бибкод : 2007Sci...317..116J . дои : 10.1126/science.1142024 . ISSN 0036-8075 . ПМИД 17615357 . S2CID 5170702 .
Ссылки [ править ]
- ^ Дэвидсон, Майкл В. (26 мая 2005 г.). «Структура животных клеток» . Молекулярные выражения . Таллахасси, Флорида: Университет штата Флорида . Проверено 3 сентября 2008 г.
- ^ Фогель, Гретхен (20 сентября 2018 г.). «Это ископаемое является одним из самых ранних животных в мире, если судить по молекулам жира, сохранившимся в течение полумиллиарда лет» . Наука . АААС . Проверено 21 сентября 2018 г.
- ^ Бобровский, Илья (2018). «Древние стероиды делают эдиакарскую ископаемую дикинсонию одним из самых ранних животных» . Наука . 361 (6408): 1246–1249. Бибкод : 2018Sci...361.1246B . дои : 10.1126/science.aat7228 . hdl : 1885/230014 . ПМИД 30237355 .
- ^ Реталлак, Дж.Дж. (2007). «Рост, распад и уплотнение захоронений дикинсонии, культовой окаменелости Эдиакарского региона» (PDF) . Алчеринга: Австралазийский журнал палеонтологии . 31 (3): 215–240. дои : 10.1080/03115510701484705 . S2CID 17181699 .
- ^ Сперлинг, Эрик; Винтер, Якоб; Пизани, Давиде; Петерсон, Кевин (2008). «Сродство плакозой к дикинсонии и эволюция способов питания многоклеточных животных позднего докембрия» (PDF) . В Кьюсаке, М.; Оуэн, А.; Кларк, Н. (ред.). Программа с тезисами . Ежегодное собрание Палеонтологической ассоциации . Том. 52. Глазго, Великобритания. п. 81.
- ^ Золото, DA; Раннегар, Б.; Гелинг, Дж.Г.; Джейкобс, ДК (2015). «Реконструкция онтогенеза наследственного состояния подтверждает двустороннюю близость к дикинсонии». Эволюция и развитие . 17 (6): 315–397. дои : 10.1111/ede.12168 . ПМИД 26492825 . S2CID 26099557 .
- ^ Цзюнь-Юань Чен; Оливери, Паола; Фэн Гао; и др. (1 августа 2002 г.). «Животный мир докембрия: вероятные формы развития и взрослые книдарии из Юго-Западного Китая» (PDF) . Биология развития . 248 (1): 182–196. дои : 10.1006/dbio.2002.0714 . ISSN 0012-1606 . ПМИД 12142030 . Архивировано из оригинала (PDF) 26 мая 2013 года . Проверено 4 февраля 2015 г.
- ^ Гражданкин, Дима (июнь 2004 г.). «Схемы распространения в эдиакарской биоте: фации против биогеографии и эволюции». Палеобиология . 30 (2): 203–221. doi : 10.1666/0094-8373(2004)030<0203:PODITE>2.0.CO;2 . ISSN 0094-8373 . S2CID 129376371 .
- ^ Зейлахер, Адольф (август 1992 г.). «Вендобионта и Псаммокораллия: утраченные конструкции докембрийской эволюции» . Журнал Геологического общества . 149 (4): 607–613. Бибкод : 1992JGSoc.149..607S . дои : 10.1144/gsjgs.149.4.0607 . ISSN 0016-7649 . S2CID 128681462 . Проверено 4 февраля 2015 г.
- ^ Мартин, Марк В.; Гражданкин Дмитрий В.; Боуринг, Сэмюэл А.; и др. (5 мая 2000 г.). «Возраст неопротерозойского двулатерального тела и следы окаменелостей, Белое море, Россия: значение для эволюции многоклеточных животных». Наука . 288 (5467): 841–845. Бибкод : 2000Sci...288..841M . дои : 10.1126/science.288.5467.841 . ISSN 0036-8075 . ПМИД 10797002 . S2CID 1019572 .
- ^ Федонкин Михаил Александрович; Ваггонер, Бенджамин М. (28 августа 1997 г.). позднего докембрия «Ископаемая кимберелла представляет собой двусторонний организм, похожий на моллюска» . Природа . 388 (6645): 868–871. Бибкод : 1997Natur.388..868F . дои : 10.1038/42242 . ISSN 0028-0836 . S2CID 4395089 .
- ^ Муи, Рич; Дэвид, Бруно (декабрь 1998 г.). «Эволюция внутри причудливого типа: гомологии первых иглокожих» (PDF) . Американский зоолог . 38 (6): 965–974. дои : 10.1093/icb/38.6.965 . ISSN 1540-7063 . Проверено 5 февраля 2015 г.
- ^ Макменамин, Марк А.С. (сентябрь 2003 г.). Сприггина — трилобитоидное экдизозойное животное . Геонаучные горизонты, Сиэтл, 2003 . Рефераты с программами. Том. 35. Боулдер, Колорадо: Геологическое общество Америки. п. 105. OCLC 249088612 . Архивировано из оригинала 12 апреля 2016 года . Проверено 24 ноября 2007 г. Документ № 40-2, представленный на ежегодном собрании Геологического общества Америки в Сиэтле в 2003 г. (2–5 ноября 2003 г.) 2 ноября 2003 г. в Конференц-центре штата Вашингтон .
- ^ Джих-Пай Линь; Гон, Сэмюэл М. III; Гелинг, Джеймс Г.; и др. (2006). « Парванкориноподобное членистоногое из кембрия Южного Китая». Историческая биология: Международный журнал палеобиологии . 18 (1): 33–45. дои : 10.1080/08912960500508689 . ISSN 1029-2381 . S2CID 85821717 .
- ^ Баттерфилд, Николас Дж. (декабрь 2006 г.). «Зацепка некоторых« червей »ствольной группы: ископаемые лофотрохозойи в сланцах Берджесс». Биоэссе . 28 (12): 1161–1166. doi : 10.1002/bies.20507 . ISSN 0265-9247 . ПМИД 17120226 . S2CID 29130876 .
- ^ Jump up to: Перейти обратно: а б Бенгтсон, Стефан (2004). «Ранние скелетные окаменелости». В Липпсе, Джере Х .; Ваггонер, Бенджамин М. (ред.). Неопротерозойско-кембрийские биологические революции: представлено в виде краткого курса палеонтологического общества на ежегодном собрании Геологического общества Америки, Денвер, Колорадо, 6 ноября 2004 г. Документы Палеонтологического общества. Том. 10. Нью-Хейвен, Коннектикут: Служба репрографии и изображений Йельского университета; Палеонтологическое общество . стр. 67–78. OCLC 57481790 .
- ^ Jump up to: Перейти обратно: а б Гулд, Стивен Джей (1990) Замечательная жизнь: сланцы Бёрджесс и природа истории В. В. Нортон. ISBN 9780393307009 .
- ^ Бадд, Грэм Э. (февраль 2003 г.). «Кембрийские окаменелости и происхождение типов» (PDF) . Интегративная и сравнительная биология . 43 (1): 157–165. дои : 10.1093/icb/43.1.157 . ISSN 1557-7023 . ПМИД 21680420 . Проверено 6 февраля 2015 г.
- ^ Бадд, Грэм Э. (март 1996 г.). «Морфология Opabinia regalis и реконструкция стеблевой группы членистоногих». Летайя . 29 (1): 1–14. дои : 10.1111/j.1502-3931.1996.tb01831.x . ISSN 0024-1164 .
- ^ Маршалл, Чарльз Р. (май 2006 г.). «Объяснение кембрийского «взрыва» животных». Ежегодный обзор наук о Земле и планетах . 34 : 355–384. Бибкод : 2006AREPS..34..355M . doi : 10.1146/annurev.earth.33.031504.103001 . ISSN 1545-4495 . S2CID 85623607 .
- ^ Валентин, Джеймс В. (2004). О происхождении Филы . Чикаго: Издательство Чикагского университета. п. 7. ISBN 978-0-226-84548-7 .
Классификации организмов в иерархических системах использовались в семнадцатом и восемнадцатом веках. Обычно организмы группировались по их морфологическому сходству, как это воспринималось теми ранними исследователями, а затем эти группы группировались по их сходству и так далее, чтобы сформировать иерархию.
- ^ Jump up to: Перейти обратно: а б Валентайн, Джеймс В. (18 июня 2004 г.). О происхождении Филы . Издательство Чикагского университета. ISBN 978-0-226-84548-7 .
- ^ Эрвин, Дуглас; Валентин, Джеймс; Яблонски, Дэвид (1997). «Недавние находки окаменелостей и новое понимание развития животных открывают новый взгляд на загадку массового роста животных в раннем кембрии» . Американский ученый (март – апрель).
- ^ Jump up to: Перейти обратно: а б Бадд, GE; Дженсен, С. (май 2000 г.). «Критическая переоценка летописи окаменелостей двусторонних типов» . Биологические обзоры . 75 (2): 253–295. дои : 10.1111/j.1469-185X.1999.tb00046.x . ПМИД 10881389 . S2CID 39772232 .
- ^ Jump up to: Перейти обратно: а б Подумайте, ВФ; Линдберг, доктор медицинских наук, ред. (2008). Филогения и эволюция моллюсков . Беркли: Издательство Калифорнийского университета. п. 481. ИСБН 978-0-520-25092-5 .
- ^ Porifera (n.) Онлайн-словарь этимологии . Проверено 18 августа 2016 г.
- ^ Васелет Дж., Дюпор Э. (2004). «Захват добычи и переваривание хищной губки Asbestopluma hypogea (Porifera: Demospongiae)». Зооморфология . 123 (4): 179–190. дои : 10.1007/s00435-004-0100-0 . S2CID 24484610 .
- ^ СК «Боб» Брэйн; Энтони Р. Праве; Карл-Хайнц Хоффманн; Энтони Э. Фалик; Андре Бота; Дональд А. Херд; Крейг Старрок; Иэн Янг; Дэниел Дж. Кондон; Стюарт Г. Эллисон (2012). «Первые животные: губкообразные окаменелости возрастом около 760 миллионов лет из Намибии» . Южноафриканский научный журнал . 108 (1/2). дои : 10.4102/sajs.v108i1/2.658 . hdl : 2263/18222 .
- ^ «Губка Линней, 1759 год» . Всемирный реестр морских видов . Проверено 18 июля 2012 г.
- ^ Роуленд, С.М. и Стивенс, Т. (2001). «Археоциаты: история филогенетической интерпретации». Журнал палеонтологии . 75 (6): 1065–1078. doi : 10.1666/0022-3360(2001)075<1065:AAHOPI>2.0.CO;2 . JSTOR 1307076 . S2CID 86211946 .
- ^ Сперлинг, Э.А.; Пизани, Д.; Петерсон, К.Дж. (1 января 2007 г.). «Пориферановая парафилия и ее значение для палеобиологии докембрия» (PDF) . Геологическое общество, Лондон, специальные публикации . 286 (1): 355–368. Бибкод : 2007GSLSP.286..355S . дои : 10.1144/SP286.25 . S2CID 34175521 . Архивировано из оригинала (PDF) 9 мая 2009 года . Проверено 22 августа 2012 г.
- ^ Чжан, З.-Ц. (2011). «Биоразнообразие животных: введение в классификацию более высокого уровня и таксономическое богатство» (PDF) . Зоотакса . 3148 : 7–12. дои : 10.11646/zootaxa.3148.1.3 .
- ^ « Нематостелла vectensis v1.0» . Геномный портал . Калифорнийский университет . Проверено 19 января 2014 г.
- ^ « Нематостелла » . Nematostella.org . Проверено 18 января 2014 г.
- ^ Jump up to: Перейти обратно: а б Генихович, Г. и У. Технау (2009). «Анемон звездочка Nematostella vectensis : модельный организм антозоев для исследований в области сравнительной геномики и функциональной эволюционной биологии развития». Протоколы Колд-Спринг-Харбора . 2009 (9): pdb.emo129. дои : 10.1101/pdb.emo129 . ПМИД 20147257 .
- ^ «Откуда взялась наша голова? Безмозглая актиния проливает новый свет на эволюционное происхождение головы» . Наука Дейли . 12 февраля 2013 года . Проверено 18 января 2014 г.
- ^ Синигалья, К.; и др. (2013). «Ген формирования билатерального паттерна головы шесть3/6 контролирует развитие аборального домена у книдарий» . ПЛОС Биология . 11 (2): e1001488. дои : 10.1371/journal.pbio.1001488 . ПМЦ 3586664 . ПМИД 23483856 .
- ^ Карлескинт Г., Ричард Тернер Р. и Джеймс Смолл Дж. (2012) Введение в морскую биологию Cengage Learning, издание 4, стр. 445. ISBN 9781133364467 .
- ^ Майкл Ле Пейдж (март 2019 г.). «Животное с анусом, который приходит и уходит, может показать, как развивался наш анус» . Новый учёный.
- ^ Бавестрелло, Джорджо; Кристиан Зоммер; Мишель Сара (1992). «Двунаправленное преобразование у Turritopsis nutricula (Hydrozoa)». Морская наука . 56 (2–3): 137–140.
- ^ Пираино, Стефано; Ф. Боэро; Б. Эшбах; В. Шмид (1996). «Обращение жизненного цикла: медузы превращаются в полипы и трансдифференцировка клеток у Turritopsis nutricula (Cnidaria, Hydrozoa)». Биологический бюллетень . 190 (3): 302–312. дои : 10.2307/1543022 . JSTOR 1543022 . ПМИД 29227703 . S2CID 3956265 .
- ^ «Корнуолл – Природа – Червь-суперзвезда» . Би-би-си.
- ^ Марк Карвардин (1995) Книга рекордов Гиннеса животных . Издательство Гиннесс. п. 232.
- ^ «Постоянные паразиты» . Журнал «Тайм» . 8 апреля 1957 года. Архивировано из оригинала 27 июня 2008 года.
- ^ Харгис, Уильям Дж. (1985). «Паразитология и патология морских организмов Мирового океана». НОАА Тех. Представитель .
- ^ «Классификация паразитов животных» . Архивировано из оригинала 6 октября 2017 года . Проверено 5 сентября 2016 г.
- ^ Гарсия, Линн. «Классификация человеческих паразитов, переносчиков и подобных организмов» (PDF) . Медицинский центр Калифорнийского университета в Лос-Анджелесе, Калифорния.
- ^ Ходда, М. (2011). «Тип Nematoda Cobb, 1932. В: Чжан, З.-К. (ред.) Биоразнообразие животных: обзор классификации более высокого уровня и обзор таксономического богатства». Зоотакса . 3148 : 63–95. дои : 10.11646/zootaxa.3148.1.11 .
- ^ Чжан, Z (2013). «Биоразнообразие животных: обновление классификации и разнообразия в 2013 году. В: Чжан, З.-К. (ред.) Биоразнообразие животных: план классификации более высокого уровня и исследование таксономического богатства (Дополнения, 2013 г.)» . Зоотакса . 3703 (1): 5–11. дои : 10.11646/zootaxa.3703.1.3 . S2CID 85252974 .
- ^ Лэмбсхед PJD (1993). «Последние достижения в исследованиях биоразнообразия морского бентоса». Океанис . 19 (6): 5–24.
- ^ Боргони Г., Гарсия-Мояно А., Литтауэр Д., Берт В., Бестер А., ван Херден Э., Мёллер С., Эразмус М., Онстотт Т.К. (июнь 2011 г.). «Нематода из земных глубоких недр Южной Африки» . Природа . 474 (7349): 79–82. Бибкод : 2011Natur.474...79B . дои : 10.1038/nature09974 . hdl : 1854/LU-1269676 . ПМИД 21637257 . S2CID 4399763 .
- ^ Дановаро Р., Гамби С., Делл'Анно А., Коринальдези С., Фраскетти С., Ванрёзель А., Винкс М., Гудэй Эй.Дж. (январь 2008 г.). «Экспоненциальное снижение функционирования глубоководных экосистем, связанное с утратой донного биоразнообразия» . Курс. Биол . 18 (1): 1–8. дои : 10.1016/j.cub.2007.11.056 . ПМИД 18164201 . S2CID 15272791 .
- «Исчезновение глубоководных видов может привести к коллапсу океанов, предполагает исследование» . ЭврекАлерт! (Пресс-релиз). 27 декабря 2007 г.
- ^ Платт Х.М. (1994). «предисловие». В Лоренцен С., Лоренцен С.А. (ред.). Филогенетическая систематика свободноживущих нематод . Лондон: Общество Рэев. ISBN 978-0-903874-22-9 .
- ^ «Сеть разнообразия животных - Иглокожие» . Зоологический музей Мичиганского университета . Проверено 26 августа 2012 г.
- ^ Фокс, Ричард. « Астериас форбеси » . Анатомия беспозвоночных онлайн . Ландерский университет . Проверено 14 июня 2014 г.
- ^ Холсингер, К. (2005). Краеугольные виды. Получено 10 мая 2010 г. с сайта «Ключевые виды» . Архивировано из оригинала 30 июня 2010 года . Проверено 12 мая 2010 г.
- ^ Чепмен, AD (2009). Численность живых видов в Австралии и мире, 2-е издание . Австралийское исследование биологических ресурсов, Канберра. Проверено 12 января 2010 г. ISBN 978-0-642-56860-1 (печатный); ISBN 978-0-642-56861-8 (онлайн).
- ^ Хэнкок, Ребекка (2008). «Признание исследований моллюсков» . Австралийский музей. Архивировано из оригинала 30 мая 2009 года . Проверено 9 марта 2009 г.
- ^ Барнс, РСК, Калоу П., Олив, PJW, Голдинг Д.В., Спайсер Дж.И. (2001). Беспозвоночные, синтез (3-е изд.). Великобритания: Blackwell Science.
- ^ «Добро пожаловать в CephBase» . CephBase . Архивировано из оригинала 12 января 2016 года . Проверено 29 января 2016 г.
- ^ Уилбур, Карл М.; Кларк, MR; Труман, ER, ред. (1985), Моллюска , том. 12. Палеонтология и неонтология головоногих моллюсков, Нью-Йорк: Academic Press, ISBN. 0-12-728702-7
- ^ «Есть ли пресноводные головоногие моллюски?» . Австралийская радиовещательная корпорация . 16 января 2013 г.
- ^ Блэк, Ричард (26 апреля 2008 г.). «Колоссальный кальмар из морозилки» . Новости Би-би-си.
- ^ Юэн Каллауэй (2 июня 2008 г.). «Простодушный Наутилус демонстрирует вспышку памяти» . Новый учёный . Проверено 7 марта 2012 г.
- ^ Кэтрин Филлипс (15 июня 2008 г.). «Живые ископаемые воспоминания». Журнал экспериментальной биологии . 211 (12): iii. дои : 10.1242/jeb.020370 . S2CID 84279320 .
- ^ Робин Крук и Дженнифер Бэзил (2008). «Двухфазная кривая памяти у наутилуса с камерами Nautilus pompilius L. (Cephalopoda: Nautiloidea)». Журнал экспериментальной биологии . 211 (12): 1992–1998. дои : 10.1242/jeb.018531 . ПМИД 18515730 . S2CID 6305526 .
- ^ Jump up to: Перейти обратно: а б с д и ж Руперт, Эдвард Э.; Фокс, Ричард С.; Барнс, Роберт Д. (2004). Зоология беспозвоночных, 7-е издание . Cengage Обучение. ISBN 978-81-315-0104-7 .
- ^ Jump up to: Перейти обратно: а б Хейворд, П.Дж. (1996). Справочник морской фауны Северо-Западной Европы . Издательство Оксфордского университета. ISBN 978-0-19-854055-7 .
- ^ Гирибет, Г.; Окусу, А, А.; Линдгрен, Арканзас, Арканзас; Хафф, Юго-Западный, Юго-Западный; Шредль, М, М.; Нисигути, МК, МК (май 2006 г.). «Свидетельства клады, состоящей из моллюсков с серийно повторяющимися структурами: моноплакофоры родственны хитонам» . Труды Национальной академии наук Соединенных Штатов Америки . 103 (20): 7723–7728. Бибкод : 2006PNAS..103.7723G . дои : 10.1073/pnas.0602578103 . ПМЦ 1472512 . ПМИД 16675549 .
- ^ Хили, Дж. М. (2001). «Моллюска». В Андерсоне, Д.Т. (ред.). Зоология беспозвоночных (2-е изд.). Издательство Оксфордского университета. стр. 120–171. ISBN 978-0-19-551368-4 .
- ^ Работаем, да мы. «Узнайте больше о Первой жизни — Первая жизнь Дэвида Аттенборо» . Архивировано из оригинала 26 января 2011 года.
- ^ доктор Карри; ТМ Уорд (2009). Южно-Австралийский гигантский краб ( Pseudocarcinus gigas ) Промысел (PDF) . Южно-Австралийский институт исследований и разработок . Отчет об оценке рыболовства для PIRSA. Архивировано из оригинала (PDF) 28 марта 2012 года . Проверено 9 декабря 2013 г.
- ^ Гевин, В. (2005). «Функциональная геномика расширяет биологический сюжет» . ПЛОС Биология . 3 (6): е219. дои : 10.1371/journal.pbio.0030219 . ПМЦ 1149496 . ПМИД 15941356 .
- ^ Геном ланцетника (амфиоксуса) и происхождение позвоночных Ars Technica , 19 июня 2008 г.
- ^ Лемэр, П. (2011). «Эволюционный перекресток биологии развития: оболочники». Разработка . 138 (11): 2143–2152. дои : 10.1242/dev.048975 . ПМИД 21558365 . S2CID 40452112 .
- ^ Карсон, Рэйчел (1997). Море вокруг нас . Оксфордские книги в мягкой обложке. стр. 190–191. ISBN 978-0-19-506997-6 .

