Криноид
| Криноидеи Временной диапазон: | |
|---|---|
 | |
| Криноид на рифе острова Бату Мончо, Индонезия. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Иглокожие |
| Подтип: | Кринозоа |
| Сорт: | Криноид Миллер , 1821 г. [2] |
| Основные группы | |
| |
Криноидеи — морские беспозвоночные, относящиеся к классу Cinoidea . Морские лилии, которые во взрослой форме остаются прикрепленными к морскому дну стеблем, обычно называются морскими лилиями , а формы без стеблей - перьевыми звездами. [3] [4] или коматулиды , являются членами крупнейшего отряда криноидей , Comatulida . Морские лилии — иглокожие из типа Echinodermata , который также включает морские звезды , хрупкие звезды , морские ежи и морские огурцы . [5] Они живут как на мелководье [6] и на глубине до 9000 метров (30000 футов). [7]
Для взрослых криноидей рот расположен на верхней поверхности. Он окружен питающимися рукавами и связан с U-образной кишкой, при этом анус расположен на ротовом диске возле рта. Хотя основной образец пятикратной симметрии иглокожих можно распознать, у большинства криноидей пять рук подразделяются на десять или более. У них есть перистые перышки, и они широко раскинуты для сбора планктонных частиц из воды. На каком-то этапе своей жизни у большинства криноидей есть короткий стебель, используемый для прикрепления к субстрату , но многие из них живут прикрепленными только в молодом возрасте и становятся свободно плавающими, когда становятся взрослыми.
На сегодняшний день существует всего около 700 видов криноидей. [8] но в прошлом этот класс был гораздо более многочисленным и разнообразным. Некоторые мощные пласты известняков, датируемые периодом от середины до палеозоя юры , почти полностью состоят из разрозненных фрагментов криноидей. [9] [10] [11]
Этимология
[ редактировать ]Название «Crinoidea» происходит от древнегреческого слова κρίνον ( krínon ), «лилия», с суффиксом –oid , означающим «подобный». [12] [13] Морских лилий принято называть морскими лилиями, которые во взрослом состоянии прикрепляются к морскому дну стебельком. [14] а формы без стеблей называются перьевыми звездами. [15] или коматулиды, являющиеся представителями крупнейшего отряда криноидей — Comatulida . [16]
Онлайн-база данных модельных организмов
[ редактировать ]Echinobase — это база данных модельных организмов окрашенного ежа и ряда других иглокожих.
Морфология
[ редактировать ]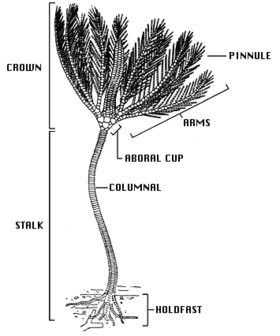
Основная форма тела криноидея — это стебель (отсутствующий у взрослых перьевых звезд) и корона, состоящая из чашеобразного центрального тела, известного как тека, и набора из пяти лучей или рукавов, обычно разветвленных и перистых. Рот ( верхнюю и анус расположены на верхней стороне теки, что делает дорсальную ) поверхность ротовой поверхностью, в отличие от других групп иглокожих, таких как морские ежи , морские звезды и хрупкие звезды , где рот находится на нижней стороне. [17] Многочисленные известковые пластинки составляют основную часть криноидея с небольшим процентом мягких тканей. , Англия, есть пласты известняка, датируемые нижним карбоном Эти косточки хорошо окаменевают, и в окрестностях Клитеро , образовавшиеся почти исключительно из разнообразной фауны окаменелостей криноидей. [18]

Стебель морских лилий состоит из столбика высокопористых косточек, соединенных связочной тканью. Он прикрепляется к субстрату с помощью уплощенной опоры или завитков сочлененных корнеобразных структур, известных как усики . Дальнейшие усики могут располагаться выше по стеблю. У криноидей, прикрепляющихся к твердым поверхностям, усики могут быть крепкими и изогнутыми, напоминая птичьи лапки, но когда криноидеи живут на мягких отложениях, усики могут быть тонкими и стержнеобразными. У молодых перьевых звезд есть стебель, но позже он теряется, и у многих видов сохраняются несколько усиков у основания кроны. Большинство современных криноидей свободно плавают и имеют лишь рудиментарный стебель. У тех глубоководных видов, которые все еще сохраняют стебель, он может достигать 1 м (3 футов) в длину (хотя обычно намного меньше), а известны ископаемые виды со стеблями длиной 20 м (66 футов). [5] самый крупный зарегистрированный криноидей со стеблем длиной 40 м (130 футов). [19]
Тека пятичленная (имеет пятичастную симметрию) и гомологична телу или диску других иглокожих. Основание теки образовано чашеобразным набором косточек (костных пластинок) — чашечкой , а верхняя поверхность — слабообызвествленным тегменом — перепончатым диском. Тегмен разделен на пять «амбулакральных областей», включая глубокую бороздку, из которой выступают ножки трубки , и пять «интерамбулакральных областей» между ними. Рот находится вблизи центра или на краю надкрылья, а амбулакральные бороздки ведут от основания рук ко рту. Анальное отверстие также расположено на надкрыльях, часто на небольшом приподнятом конусе, в межамбулакральной области. Тека относительно небольшая и содержит органы пищеварения криноидей. [5]
Руки поддерживаются рядом сочленяющихся косточек, подобных тем, что находятся на стебле. Изначально у криноидей было только пять рук, но у большинства современных форм они разделены на две части у II косточки, что дает всего десять рук. У большинства современных видов, особенно у свободноплавающих перьевых звезд, рукава ветвятся еще несколько раз, всего образуя до двухсот ветвей. Будучи сочлененными, руки могут сгибаться. Они поочередно выстланы с обеих сторон более мелкими суставчатыми придатками, известными как «перышки», которые придают им вид перьев. И руки, и перышки имеют трубчатые ножки по краям амбулакральных борозд. Ножки трубки поставляются группами по три штуки разного размера; у них нет присосок, и они используются для удержания и манипулирования частицами пищи. Борозды оснащены ресничками , которые облегчают кормление, перемещая органические частицы вдоль руки и в рот. [5]
 Стебель, тека и руки «настоящего» (стебельчатого) криноидея (семейство Isselicrinidae )
Стебель, тека и руки «настоящего» (стебельчатого) криноидея (семейство Isselicrinidae ) Оксикомантус беннетти (коматулид)
Оксикомантус беннетти (коматулид) Тегмен Lamprometra palmata . Рот расположен в центре пяти питающих желобков, а анальное отверстие — в верхней части колонны.
Тегмен Lamprometra palmata . Рот расположен в центре пяти питающих желобков, а анальное отверстие — в верхней части колонны. Крупный план усиков, которые позволяют коматулидам ходить и прикрепляться.
Крупный план усиков, которые позволяют коматулидам ходить и прикрепляться. Крупный план перьев Tropiometra carinata (с паразитами Myzostoma fuscomaculatum )
Крупный план перьев Tropiometra carinata (с паразитами Myzostoma fuscomaculatum )





Онлайн-база данных модельных организмов
[ редактировать ]Echinobase — это база данных модельных организмов перьевой звезды и ряда других иглокожих.
Биология
[ редактировать ]Кормление
[ редактировать ]
Криноидеи — пассивные питатели взвеси , фильтрующие планктон и мелкие частицы детрита из морской воды, протекающей мимо них, своими перьевыми руками. Руки подняты в форме веера, который держится перпендикулярно течению. Подвижные криноидеи перемещаются, садясь на камни, коралловые вершины или другие возвышенности, чтобы максимизировать свои возможности для кормления. Частицы пищи улавливаются первичными (самыми длинными) ножками трубки, которые полностью выдвинуты и удерживаются прямо на кончиках, образуя сетку для улавливания пищи, в то время как вторичные и третичные ножки трубки участвуют в манипулировании всем, что встречается. [5]
Ножки трубки покрыты липкой слизью , которая задерживает любые попадающие на нее частицы. Как только они поймают частицу пищи, ножки трубки перебрасывают ее в амбулакральную борозду, где реснички продвигают слизь и частицы пищи ко рту. Лепестки по бокам канавки помогают удерживать струю слизи на месте. Общая длина поверхности, улавливающей пищу, может быть очень большой; 56 рук японской морской лилии с руками 24 см (9 дюймов) имеют общую длину 80 м (260 футов), включая перышки. Вообще говоря, криноидеи, живущие в среде с относительно небольшим количеством планктона, имеют более длинные и сильно разветвленные руки, чем те, которые живут в среде, богатой пищей. [5]
Рот опускается в короткий пищевод . Настоящего желудка нет, поэтому пищевод соединяется непосредственно с кишечником , который проходит одной петлей прямо вокруг внутренней части чашечки. Кишечник часто включает многочисленные дивертикулы , некоторые из которых могут быть длинными или разветвленными. Конец кишки открывается в короткую мышечную прямую кишку . Он поднимается к анусу , который выступает из небольшого конического выступа на краю надкрылья. Фекалии формируются в крупные, сцементированные слизистой оболочкой гранулы, которые падают на надкрылья, а затем на субстрат. [5]
Хищничество
[ редактировать ]экземпляры морского ежа Calocidaris micans, обнаруженные вблизи криноидея Endoxocrinus parrae Было показано, что , содержат большое количество частей стебля в своих кишках. Они состоят из сочлененных косточек с мягкими тканями, тогда как местный осадок содержал только разрозненные косточки без мягких тканей. Это делает весьма вероятным, что эти морские ежи являются хищниками криноидей, и что криноидеи убегают, отдавая при этом часть своего стебля. [20]
Различные окаменелости криноидей намекают на возможных доисторических хищников. копролиты как рыб, так и головоногих моллюсков, Были обнаружены содержащие косточки различных криноидей, таких как пелагический криноид Saccocoma , из юрского lagerstatten Solnhofen , [21] в то время как поврежденные стебли криноидей со следами укусов, соответствующими зубным пластинам коккостейд плакодерм , были обнаружены в позднедевонской Польше . [22] Чашечки некоторых криноидей девонского и каменноугольного возраста имеют раковины улитки Platyceras , тесно связанные с ними. [23] У некоторых улитка располагалась над анусом, что позволяет предположить, что Platyceras был копрофагом- комменсалом, в то время как у других животное располагалось непосредственно над скважиной, что предполагает более пагубную связь. [24]
Водная сосудистая система
[ редактировать ]Как и другие иглокожие, криноидеи обладают водной сосудистой системой , которая поддерживает гидравлическое давление в трубчатых ножках. Он не связан с внешней морской водой через мадрепорит , как у других иглокожих, а лишь соединен через большое количество пор с целомом (полостью тела). Основным резервуаром жидкости является кольцевой канал с мышечными стенками, который соединен с целомом каменными каналами, выстланными известковым материалом. на ряд сообщающихся между собой пространств Целом разделен брыжейками . Он окружает внутренние органы диска и имеет ветви внутри стебля и рук, причем более мелкие ветви доходят до перьев. Это сокращение кольцевого канала, которое расширяет ножки трубки. В каждое плечо входят три узкие ветви целома: две с оральной стороны и одна аборальная, и перышки. Под действием ресничек происходит медленный ток жидкости (1 мм в секунду) в этих каналах наружу в ротовых ветвях и внутрь в аборальных, и это является основным средством транспорта питательных веществ и продуктов жизнедеятельности. Сердца и отдельной системы кровообращения нет, но у основания диска находится большой кровеносный сосуд, известный как осевой орган, содержащий несколько тонких трубок с закрытыми концами неизвестного назначения, которые простираются до стебля. [5]
Эти различные заполненные жидкостью пространства, помимо транспортировки питательных веществ по организму, также функционируют как дыхательная и выделительная системы. Кислород поглощается преимущественно через ножки трубки, которые являются наиболее тонкостенными частями тела, при этом дальнейший газообмен происходит на большой площади поверхности рук. Специализированных органов выделения нет, а отходы собираются фагоцитирующими целомоцитами. [5]
Нервная система
[ редактировать ]Нервная система криноида разделена на три части, между которыми имеются многочисленные связи. Ротовая или самая верхняя часть — единственная гомологична нервной системе других иглокожих. Он состоит из центрального нервного кольца, окружающего рот, и радиальных нервов, разветвляющихся на руки, и выполняет сенсорную функцию. Ниже этого находится промежуточное нервное кольцо, от которого отходят лучевые нервы, иннервирующие руки и пиннулы. Эти нервы являются двигательными по своей природе и контролируют мускулатуру трубчатых стоп. Третья часть нервной системы расположена аборально и отвечает за сгибание и движение рук, перьев и усиков. Он сосредоточен на массе нервной ткани возле основания чашечки и обеспечивает по одному нерву каждой руке и нескольким нервам стебля. [5]
Размножение и жизненный цикл
[ редактировать ]Криноидеи не способны к клональному размножению, как некоторые морские звезды и хрупкие звезды , но способны регенерировать утраченные части тела. Руки, оторванные хищниками или поврежденные неблагоприятными условиями окружающей среды, могут вырасти заново, и даже висцеральная масса может регенерировать в течение нескольких недель. Эта регенерация может иметь жизненно важное значение для выживания при нападениях хищных рыб. [5]
Морские лилии раздельнополы : особи бывают мужского или женского пола. У большинства видов гонады расположены в перышках, но у некоторых - в руках. Не все перышки репродуктивны, только те, что расположены ближе всего к макушке. Гаметы . образуются в половых каналах, заключенных в половые целомы Перышки в конечном итоге разрываются, высвобождая сперму и яйцеклетки в окружающую морскую воду. У некоторых родов, таких как Antedon , оплодотворенные яйца прикрепляются к ветвям секретом эпидермальных желез; у других, особенно холодноводных видов из Антарктиды, икра высиживается в специальных мешочках на рукавах или перышках. [5]
Оплодотворенные яйца вылупляются и выпускают свободно плавающие личинки желточников . Двусторонне-симметричная личинка имеет бочкообразную форму с кольцами ресничек, идущими по всему телу, и пучком чувствующих волосков на верхнем полюсе. Хотя среди четырех других существующих классов иглокожих существуют как питающиеся (планктотрофные), так и не питающиеся (лецитотрофные) личинки, все современные криноидеи, по-видимому, являются потомками выжившей клады, которая прошла через узкое место после пермского вымирания , потеряв в то время питание на личиночной стадии. [25] Период свободного плавания личинки длится всего несколько дней, прежде чем она оседает на дно и прикрепляется к подстилающей поверхности с помощью клейкой железы на ее нижней стороне. Затем личинка претерпевает длительный период метаморфоз в молодь на стебле , становясь при этом радиально-симметричной. Даже свободно плавающие перьевые звезды проходят через эту стадию, когда взрослая особь в конце концов отрывается от стебля. [5]
Передвижение
[ редактировать ]
Большинство современных криноидей, то есть перьевых звезд, во взрослом состоянии свободно передвигаются и не имеют стебля. Примеры ископаемых криноидей, которые были интерпретированы как свободно плавающие, включают Marsupites , Saccocoma и Uintacrinus . [26] Обычно криноидеи перемещаются в новые места ползком, используя усики в качестве ног. Такое движение может быть вызвано изменением направления течения, необходимостью подняться на возвышение для кормления или агонистическим поведением встреченной особи. [27] Криноидеи также умеют плавать. Они делают это скоординированными, повторяющимися последовательными движениями рук в трех группах. Сначала направление движения вверх, но вскоре становится горизонтальным, перемещаясь со скоростью около 7 см (2,8 дюйма) в секунду, при этом поверхность полости рта находится впереди. Плавание обычно происходит в виде коротких всплесков активности продолжительностью до полминуты, и, по крайней мере, у коматулид Florometra serratissima , происходит только после механической стимуляции или в качестве реакции бегства, вызванной хищником. [27]
В 2005 году был зарегистрирован стебельчатый криноид, ползущий по морскому дну у острова Большой Багама . Хотя было известно, что стебельчатые криноидеи могут двигаться, до этой записи самое быстрое движение, известное для стебельчатых криноидей, составляло 0,6 метра (2 фута) в час. Запись 2005 года показала, что один из них движется по морскому дну с гораздо большей скоростью: от 4 до 5 см (от 1,6 до 2,0 дюйма) в секунду, или от 144 до 180 м (от 472 до 591 фута) в час. [28]
Эволюция
[ редактировать ]Происхождение
[ редактировать ]

Если игнорировать загадочного Echmatocrinus из сланцев Бёрджесс , самые ранние известные однозначные группы криноидей относятся к ордовику , 480 миллионов лет назад. Существуют две конкурирующие гипотезы относительно происхождения группы: традиционная точка зрения предполагает, что криноидеи развились изнутри бластозойных ( эокриноиды и их производные потомки, бластоиды и цистоиды ), тогда как наиболее популярная альтернатива предполагает, что криноидеи рано разделились. из числа эдриоастероидов . [29] Спор трудно разрешить, отчасти потому, что все три предка-кандидата имеют много общих характеристик, включая радиальную симметрию, известковые пластинки и стебельчатое или прямое прикрепление к субстрату. [29]
Разнообразие
[ редактировать ]Иглокожие с минерализованными скелетами вошли в летопись окаменелостей в раннем кембрии (540 млн лет назад), и в течение следующих 100 миллионов лет доминировали криноидеи и бластоиды (также стебельковые фильтраторы). [30] В то время в состав иглокожих входили двадцать таксонов классового ранга, только пять из которых пережили последовавшие за этим массовые вымирания. Долгая и разнообразная геологическая история криноидей демонстрирует, насколько хорошо иглокожие приспособились к фильтрованному питанию. [5]
Криноидеи претерпели два периода резкой адаптивной радиации : первый в ордовике (485–444 млн лет назад), а другой в раннем триасе (около 230 млн лет назад). [31] Эта триасовая радиация привела к тому, что формы, обладающие гибкими руками, получили широкое распространение; подвижность , преимущественно ответ на давление хищников, также стала гораздо более распространенной, чем сидячая. [32] Это излучение произошло несколько раньше, чем мезозойская морская революция , возможно, потому, что оно было главным образом вызвано увеличением численности донных хищников, особенно морских ежей. [33] периода последовало избирательное массовое вымирание Затем в конце пермского , в ходе которого вымерли все бластоиды и большинство криноидей. [31] После вымирания в конце пермского периода криноидеи так и не восстановили то морфологическое разнообразие и доминирующее положение, которое они имели в палеозое; они использовали другой набор экологических стратегий, доступных им, нежели те, которые оказались столь успешными в палеозое. [31]
Окаменелости
[ редактировать ]Некоторые ископаемые криноидеи, такие как пентакриниты , по-видимому, жили прикрепленными к плавающим корягам, и часто встречаются целые колонии. Иногда коряги заболачивались и опускались на дно, унося с собой прикрепленные криноидеи. Стебель пентакринитов может достигать длины нескольких метров. Современные родственники пентакринитов живут в плавных течениях, прикрепленных к скалам концом своего стебля. Самый крупный ископаемый криноид из когда-либо зарегистрированных имел стебель длиной 40 м (130 футов). [19]
возрастом 340 миллионов лет ( Миссисипи В 2012 году трое геологов сообщили, что они выделили сложные органические молекулы из окаменелостей нескольких видов криноидей ). Идентифицированные как «напоминающие… ароматические или полиароматические хиноны », это древнейшие молекулы, которые окончательно связаны с конкретными отдельными окаменелостями, поскольку считается, что они были запечатаны внутри пор косточек осажденным кальцитом во время процесса окаменелости. [34]
Окаменелости криноидей, и в частности разрозненные столбчатые криноидеи, могут быть настолько многочисленными, что иногда они служат основными поддерживающими обломками в осадочных породах. [ нужна ссылка ] Породы такой природы называются энкринитами .
Таксономия
[ редактировать ]


Crinoidea была признана отдельной кладой иглокожих с момента определения группы Миллером в 1821 году. [35] Включает в себя множество вымерших отрядов, а также четыре близкородственных ныне живущих отряда ( Comatulida , Cyrtocrinida , Hyocrinida и Isocrinida ), входящих в подгруппу Articulata . Живые членистоногие насчитывают около 540 видов.
- Класс Криноидеи
- † Protocrinoidea ( неопределенное место )
- Подкласс † Камерата
- Орден † Диплобатрида
- Отряд † Монобатрида
- Подкласс Пентакриноидеа
- Парвкласс † Диспарида
- Отряд † Эустенокринида
- Отряд † Менниликринида
- Порядок † Тетрагонокринид
- Отряд † Calceocrinidae .
- Парвкласс Кладида
- Надотряд † Porocrinoidea
- Отряд † Гибокриниды.
- Отряд † Порокринида
- Суперзаказ † Flexibilia
- Отряд † Сагенокринида
- Отряд † Таксокринида
- Магнодер Евкладида
- † Ampelocrinida ( ареал неопределенный )
- Надотряд † Cyathoformes
- Суперотряд Артикулата
- Орден † Энкринида
- Отряд † Холокринида
- Отряд † Миллерикринида
- Заказать † Ровеакринида
- Заказать † Уинтакринид
- Заказать Коматулиду
- Заказать Циртокринида
- Заказать Гиокринида
- Заказать изокриниду
- Надотряд † Porocrinoidea
- Парвкласс † Диспарида
Филогения
[ редактировать ]Филогения . , геологическая история и классификация Crinoidea обсуждались Райтом и др (2017). [36] Эти авторы представили новые классификации, основанные на филогенезе и рангах, основанные на результатах недавнего филогенетического анализа. [35] [37] [38] [39] Их ранговая классификация высших таксонов криноидей (вплоть до отряда), не до конца решенная и с многочисленными группами incertae sedis (неопределенного размещения), проиллюстрирована на кладограмме .
| Криноид |
| |||||||||





В культуре
[ редактировать ]Ископаемые сегменты колонн криноидей, извлеченные из известняка , добытого в карьере Линдисфарна или найденные выброшенными волнами вдоль береговой линии, были пронизаны в ожерелья или четки и стали известны как четки Святого Катберта в средние века . [40] Точно так же на Среднем Западе США окаменелые сегменты колонн криноидей иногда называют индийскими бусинами . [41] Вид криноидей, Eperisocrinus Missouriensis , является окаменелостью штата Миссури . [42] Инопланетяне в фильме « Чужой» были вдохновлены криноидеями. [43]
Галерея ископаемых криноидей
[ редактировать ] Измельченные криноидеи из шамшакской свиты, юра, Иран.
Измельченные криноидеи из шамшакской свиты, юра, Иран. Ископаемое из Германии, демонстрирующее стебель, чашечку и ветви с перышками.
Ископаемое из Германии, демонстрирующее стебель, чашечку и ветви с перышками. Окаменелости криноидей возрастом 330 миллионов лет из Айовы
Окаменелости криноидей возрастом 330 миллионов лет из Айовы

 Колонновые криноидеи ( Isocrinus nicoleti ) из среднеюрской формации Кармель в районе Маунт-Кармель-Джанкшен, штат Юта.
Колонновые криноидеи ( Isocrinus nicoleti ) из среднеюрской формации Кармель в районе Маунт-Кармель-Джанкшен, штат Юта. Корневидная фиксация криноидей из верхнего ордовика, юг Огайо.
Корневидная фиксация криноидей из верхнего ордовика, юг Огайо. Внутренняя форма просвета стебля криноидеев (и внешняя форма стебля) из нижнего карбона, штат Огайо.
Внутренняя форма просвета стебля криноидеев (и внешняя форма стебля) из нижнего карбона, штат Огайо. Окаменелости Seirocrinus subsingularis из юрской черносланцевой формации Хольцмаден, Германия.
Окаменелости Seirocrinus subsingularis из юрской черносланцевой формации Хольцмаден, Германия.









Ссылки
[ редактировать ]- ^ «Кринодея – Цифровой атлас древней жизни» .
- ^ Ханссон, Ганс (2012). «Кринодея» . ВОРМС . Всемирный реестр морских видов . Проверено 30 января 2013 г.
- ^ Гордон, ДП (2009). Инвентаризация биоразнообразия Новой Зеландии: 1. Царство животных: Radiata, Lophotrochozoa, Deuterostomia . Крайстчерч: Издательство Кентерберийского университета. п. 373. ИСБН 978-1-877257-72-8 .
- ^ Макфолл-Джонсен, Морган; Ли, Ллойд (12 августа 2023 г.). «Ученые нашли новое морское существо с 20 «руками» и назвали его в честь клубники» . Инсайдер . Архивировано из оригинала 12 августа 2023 года . Проверено 13 августа 2023 г.
- ^ Перейти обратно: а б с д и ж г час я дж к л м н Руперт, Эдвард Э.; Фокс, Ричард С.; Барнс, Роберт Д. (2004). Зоология беспозвоночных: функционально-эволюционный подход (7-е изд.). Бельмонт, Калифорния: Томсон-Брукс/Коул. стр. 917–918 . ISBN 9780030259821 . ОСЛК 53021401 .
- ^ Змарзлы, Д.Л. (1985). «Фауна мелководных криноидей атолла Кваджалейн, Маршалловы острова: экологические наблюдения, сравнения между атоллами и зоогеографическое сходство» . Тихоокеанская наука . 39 : 340–358. hdl : 10125/941 .
- ^ Оджи, Т.; Огава, Ю.; Хантер, А.В. и Китазава, К. (2009). «Обнаружение плотных скоплений черешковых криноидей в желобе Идзу-Огасавара, Япония» . Зоологическая наука . 26 (6): 406–408. дои : 10.2108/zsj.26.406 . ПМИД 19583499 . S2CID 5991969 .
- ^ Размножение и развитие иглокожих и Prochordata.
- ^ Люсия, Ф. Джерри (1962). «Диагенез криноидальных отложений». Журнал SEPM осадочных исследований . 32 : 848–865. дои : 10.1306/74D70D8F-2B21-11D7-8648000102C1865D .
- ^ Блит Кейн, доктор юридических наук (сентябрь 1968 г.). «Аспекты среды осадконакопления и палеоэкологии криноидальных известняков». Шотландский геологический журнал . 4 (3): 191–208. Бибкод : 1968ScJG....4..191B . дои : 10.1144/sjg04030191 . S2CID 219538295 .
- ^ Ях, Рената (апрель 2005 г.). «Штормовые отложения нижнеюрских криноидальных известняков в Крижной толще, Западные Татры, Польша». Фации . 50 (3–4): 561–572. Бибкод : 2005Faci...50..561J . дои : 10.1007/s10347-004-0028-3 . S2CID 128947091 .
- ^ Новый универсальный полный словарь Вебстера. 2-е изд. 1979.
- ^ «криноид» . Интернет-словарь этимологии .
- ^ «Морская лилия» . Британская энциклопедия . Проверено 14 марта 2011 г.
- ^ «Перья звезды» . Британская энциклопедия . Проверено 14 марта 2011 г.
- ^ Аусич, Уильям И.; Мессинг, Чарльз Г. «Кринодея» . Древо Жизни . Проверено 14 марта 2011 г.
- ^ О'Хара, Тимоти; Бирн, Мария (2017). Австралийские иглокожие: биология, экология и эволюция . Издательство Csiro. стр. 171–180. ISBN 978-1-4863-0763-0 .
- ^ Гесс, Ганс; Бретт, Карлтон Э.; Аусич, Уильям И.; Симмс, Майкл Дж. (2002). Ископаемые криноиды . Издательство Кембриджского университета. стр. 3–5, 45–46. ISBN 978-0-521-52440-7 .
- ^ Перейти обратно: а б Понсонби, Дэвид; Дюссар, Джордж (2005). Анатомия моря . Ванкувер: Книги Raincoast. п. 129. ИСБН 978-0-8118-4633-2 .
- ^ Баумиллер, Томаш К.; Муи, Рич; Мессинг, Чарльз Г. (2008). «Ежи на лугу: палеобиологические и эволюционные последствия хищничества сидароидных криноидей». Палеобиология . 34 (1): 22–34. Бибкод : 2008Pbio...34...22B . дои : 10.1666/07031.1 . JSTOR 20445573 . S2CID 85647638 .
- ^ Гесс, Ганс (2003). «Верхнеюрский Зольнхофен Платтенкальк, Бавария, Германия» . В Бретте, Карлтон Э.; Аусич, Уильям И.; Симмс, Майкл Дж. (ред.). Ископаемые криноиды . Издательство Кембриджского университета. стр. 216–24. ISBN 978-0-521-52440-7 .
- ^ Горжелак, Пшемис Лоу; Ракович, Лукаш; Саламон, Мариуш А.; Шрек, Петр (2011). «Предполагаемые следы укусов плакодермы на девонских криноидеях из Польши». Новогодний ежегодник геологии и палеонтологии - Трактаты . 259 : 105–12. дои : 10.1127/0077-7749/2010/0111 .
- ^ Бретт, Карлтон Э.; Уокер, Салли Э. (2002). «Хищники и хищничество в морской среде палеозоя» (PDF) . Документы Палеонтологического общества . 8 : 93–118. дои : 10.1017/S1089332600001078 . Архивировано из оригинала (PDF) 13 августа 2012 г. Проверено 6 апреля 2014 г.
- ^ Ган, Форест Дж.; Баумиллер, Томаш К. (2003). «Заражение камеральных криноидей среднего девона (живетского периода) платицератидными брюхоногими моллюсками и его влияние на характер их биотического взаимодействия» (PDF) . Летайя . 36 (2): 71–82. Бибкод : 2003Лета..36...71Г . дои : 10.1080/00241160310003072 . hdl : 2027.42/75509 .
- ^ Рафф, РА; Бирн, М. (2006). «Активная эволюционная жизнь личинок иглокожих» . Наследственность . 97 (3): 244–52. дои : 10.1038/sj.hdy.6800866 . ПМИД 16850040 .
- ^ «О криноидеях» . ФоссилЭра . Проверено 15 марта 2019 г.
- ^ Перейти обратно: а б Шоу, Джорджия; Фонтейн, Арканзас (2011). «Передвижение коматулид Florometra serratissima (Echinodermata: Crinoidea) и его адаптивное значение». Канадский журнал зоологии . 68 (5): 942–950. дои : 10.1139/z90-135 .
- ^ Баумиллер, Томаш К.; Мессинг, Чарльз Г. (6 октября 2005 г.). «Ползание стебельчатых криноидей: наблюдения in situ, функциональная морфология и значение для палеозойских таксонов» . Геологическое общество Америки Рефераты с программами . Том. 37. с. 62. Архивировано из оригинала 7 апреля 2014 года . Проверено 6 апреля 2014 г.
- ^ Перейти обратно: а б Генсбург, Томас Э.; Муи, Рич; Посыпь, Джеймс; Дэвид, Бруно; Лефевр, Бертран (2010). «Пельматозойные руки из среднего кембрия Австралии: преодоление разрыва между брахиолами и плечами? Комментарий: моста нет». Летайя . 43 (3): 432–440. Бибкод : 2010Лета..43..432Г . дои : 10.1111/j.1502-3931.2010.00220.x .
- ^ Ваггонер, Бен (16 января 1995 г.). «Иглокожие: летопись окаменелостей» . Знакомство с иглокожими . Музей палеонтологии: Калифорнийский университет в Беркли . Проверено 30 марта 2019 г.
- ^ Перейти обратно: а б с Фут, Майк (1999). «Морфологическое разнообразие эволюционной радиации палеозойских и постпалеозойских криноидей». Палеобиология . 25 (сп1): 1–116. doi : 10.1666/0094-8373(1999)25[1:MDITER]2.0.CO;2 . ISSN 0094-8373 . JSTOR 2666042 . S2CID 85586709 .
- ^ Баумиллер, Томаш К. (2008). «Экологическая морфология криноидей». Ежегодный обзор наук о Земле и планетах . 36 : 221–249. Бибкод : 2008AREPS..36..221B . doi : 10.1146/annurev.earth.36.031207.124116 .
- ^ Баумиллер, ТК; Саламон, Массачусетс; Горзелак, П.; Муи, Р.; Мессинг, К.Г.; Ган, Ф.Дж. (2010). «Постпалеозойская радиация криноидей в ответ на бентосное хищничество предшествовала мезозойской морской революции» . Труды Национальной академии наук . 107 (13): 5893–5896. Бибкод : 2010PNAS..107.5893B . дои : 10.1073/pnas.0914199107 . JSTOR 25665085 . ПМЦ 2851891 . ПМИД 20231453 . ИНИСТ 22572914 .
- ^ О'Мэлли, CE; Аусич, Висконсин; Чин, Ю.-П. (2013). «Выделение и характеристика самых ранних органических молекул, специфичных для таксонов (Миссисипи, Crinoidea)». Геология . 41 (3): 347. Бибкод : 2013Geo....41..347O . дои : 10.1130/G33792.1 . Обратите внимание, что первое предложение статьи на phys.org противоречит самой статье, в которой рассматривается несколько выделений молекул из конкретных ископаемых за последнее десятилетие.
- Пэм Фрост Гордер (19 февраля 2013 г.). «Древние окаменелые морские существа дают древнейшие биомолекулы, выделенные непосредственно из окаменелостей» . Физика.орг .
- ^ Перейти обратно: а б Аусич, Уильям И.; Каммер, Томас В.; Ренберг, Элизабет К.; Райт, Дэвид Ф. (2015). «Ранняя филогения криноидей в кладе пелматозойных» . Палеонтология . 58 (6): 937–952. Бибкод : 2015Palgy..58..937A . дои : 10.1111/пала.12204 .
- ^ Райт, Дэвид Ф.; Аусич, Уильям И.; Коул, Селина Р.; Питер, Марк Э.; Ренберг, Элизабет К. (2017). «Филогенетическая систематика и классификация криноидей (Echinodermata)» . Журнал палеонтологии . 91 (4): 829–846. Бибкод : 2017JPal...91..829W . дои : 10.1017/jpa.2016.142 .
- ^ Райт, Дэвид Ф. (2017). «Байесовская оценка филогении ископаемых и эволюции ранних и средних палеозойских криноидей (Echinodermata)» . Журнал палеонтологии . 91 (4): 799–814. Бибкод : 2017JPal...91..799W . дои : 10.1017/jpa.2016.141 .
- ^ Коул, Селина Р. (2017). «Филогения и морфологическая эволюция ордовикских камер (класс Crinoidea, тип Echinodermata)» . Журнал палеонтологии . 91 (4): 815–828. Бибкод : 2017JPal...91..815C . дои : 10.1017/jpa.2016.137 .
- ^ Роуз, Грег В.; Джермин, Ларс С.; Уилсон, Нерида Г.; Экхаут, Игорь; Лантербек, Дебора; Оджи, Тацуо; Янг, Крейг М.; Браунинг, Тина; Систернас, Паула; Хелген, Лорен Э.; Стаки, Мишель; Мессинг, Чарльз Г. (2013). «Фиксированные, свободные и фиксированные: непостоянная филогения современных Crinoidea (Echinodermata) и их пермско-триасовое происхождение». Молекулярная филогенетика и эволюция . 66 (6): 161–181. Бибкод : 2013МОЛПЭ..66..161Р . дои : 10.1016/j.ympev.2012.09.018 . ПМИД 23063883 .
- ^ Лейн, Н. Гэри; Аусич, Уильям И. (2001). «Легенда о бусах Святого Катберта: палеонтологическая и геологическая перспектива». Фольклор . 112 (1): 65–73. JSTOR 1260865 .
- ^ «Определение неизвестных окаменелостей (по форме)» . Геологическая служба Кентукки / Университет Кентукки . Проверено 21 июня 2009 г.
- ^ «Ископаемое штата Миссури» . Офис государственного секретаря, штат Миссури . Проверено 31 марта 2019 г.
- ^ Брессан, Дэвид (26 апреля 2019 г.). «Окаменелости, вдохновившие «Чужого» » . Форбс . Проверено 1 февраля 2024 г.
Внешние ссылки
[ редактировать ]- Мессинг, Чарльз. «Морская звезда на палке: Знакомство с криноидеями» . Вимео.
- Страницы криноидей Чарльза Мессинга
| Базы данных органов управления : Национальные |
|---|