Морской микробиом

Все животные на Земле образуют ассоциации с микроорганизмами, включая простейших, бактерии, археи, грибы и вирусы. В океане взаимоотношения животных и микробов исторически исследовались в системах с одним хозяином-симбионтом. Тем не менее, новые исследования разнообразия морских микроорганизмов, связанных с разнообразными морскими животными -хозяевами, продвигают область исследований, изучающих взаимодействие между животным-хозяином и более многочленным микробиомом . Потенциал микробиомов влиять на здоровье, физиологию, поведение и экологию морских животных может изменить нынешние представления о том, как морские животные адаптируются к изменениям, и особенно к растущим изменениям, связанным с климатом и антропогенным воздействием, которые уже влияют на окружающую среду океана. [1]
В океанах сложно найти эукариотические организмы , которые не живут в тесной связи с микробным партнером. Микробиомы, связанные с хозяином, также влияют на биогеохимический цикл внутри экосистем, оказывая каскадное воздействие на биоразнообразие и экосистемные процессы. В настоящее время изучаются микробиомы разнообразных морских животных: от простых организмов, включая губки и гребневики, до более сложных организмов, таких как асцидии и акулы .
Фон
[ редактировать ]| Часть серии о |
| Микробиомы |
|---|
 |


Среди огромного биологического разнообразия , населяющего мировые океаны, было бы сложно найти эукариотический организм , который не жил бы в тесной связи с микробным партнером . [2] Такие симбиозы , т. е. устойчивые взаимодействия между хозяином и микробом, при которых ни один из партнеров не страдает, а хотя бы один из них получает пользу, распространены повсеместно — от мелких рифов до глубоководных гидротермальных источников . Исследования кораллов , [3] губки , [4] и моллюски [5] [6] [7] выявили некоторые чрезвычайно важные симбиотические роли, которые микробы играют в жизни своих хозяев. Однако эти исследования, как правило, были сосредоточены на небольшом количестве конкретных микробных таксонов. Напротив, большинство хозяев сохраняют группы из многих сотен различных микробов (т. е. микробиом , [8] [9] которые сами по себе могут изменяться на протяжении онтогенеза хозяина и в результате возмущений окружающей среды. [10] [11] [12] Вместо того, чтобы ассоциированные с хозяином микробы функционировали независимо, сложные многосоставные микробиомы оказывают большое влияние на приспособленность и функции своих хозяев. Изучать эти сложные взаимодействия и биологические последствия сложно, но для понимания происхождения и эволюции организмов и популяций, а также структуры и функций сообществ и экосистем необходимо углубить понимание симбиозов в системах хозяин-микробиом. [13] [14] [2]
Существует множество нерешенных вопросов в экологии и эволюции, которые можно было бы решить путем расширения филогенетического и экологического диапазона исследований микробиома, связанного с хозяином, включая все возможные взаимодействия в микробиоме. Имеются убедительные эмпирические данные и новый консенсус в отношении того, что биоразнообразие (т.е. богатство видов и их взаимодействие) повсеместно влияет на функционирование экосистем Земли, включая продуктивность экосистем. [15] [16] Однако эти исследования были сосредоточены почти исключительно на макроорганизмах. Поскольку микробные симбионты являются неотъемлемой частью большинства живых организмов, [11] [17] Необходимо расширить понимание того, как микробные симбионты способствуют продуктивности и адаптивности хозяина. [2]
Основы продуктивных экосистем
[ редактировать ]
Черный кружок: макронуклеус, белый большой кружок: пищевые вакуоли, зеленые кружочки: фототрофы, коричневые кружочки: хемоавтотрофы, желтые овалы: гетеротрофные прокариоты.
Инженеры экосистем , такие как многие виды кораллов, глубоководные мидии и трубчатые черви гидротермальных источников , вносят вклад в первичную продуктивность и создают структурные среды обитания и питательные ресурсы, которые являются основой соответствующих экосистем. [18] Все эти таксоны вступают в мутуалистический пищевой симбиоз с микробами. Существует множество примеров морского пищевого мутуализма, при котором микробы позволяют хозяевам использовать ресурсы или субстраты, которые в противном случае были бы недоступны только хозяину. Такие симбиозы были подробно описаны в редуцированных и бескислородных отложениях (например, моллюски-люциниды , стильбонематидные нематоды и бескишечные олигохеты ) и гидротермальных источниках (например, гигантский трубчатый червь или глубоководные мидии ). [19] Более того, многие основополагающие виды морских макроводорослей являются витаминными ауксотрофами (например, половина из более чем 300 обследованных видов неспособна синтезировать кобаламин ), и их продуктивность зависит от обеспечения их эпифитными бактериями. [20] Рифы часто состоят из каменистых кораллов , одного из самых известных примеров мутуалистического симбиоза, в котором динофлагеллятовые водоросли Symbiodiniaceae снабжают коралл глюкозой , глицерином и аминокислотами , в то время как коралл обеспечивает водорослям защищенную среду и ограничивает соединения (например, виды азота), необходимые для фотосинтеза. Однако это классический пример мутуалистического симбиоза, чувствительного к нарушениям окружающей среды, которые могут нарушить хрупкие взаимодействия между хозяином и микробом. Когда рифы становятся теплыми и эвтрофными, мутуалистические Symbiodiniaceae могут вызывать повреждение клеток хозяина и/или изолировать больше ресурсов для собственного роста, тем самым нанося вред хозяевам и паразитируя на них. [21] [22] Рифовые рыбы , которые ищут дом на коралловых рифах , играют важную роль в восстановлении кораллов после их беспокойства. Бактерии Epulopiscium в кишечнике рыб-хирургов производят ферменты, которые позволяют их хозяевам переваривать сложные полисахариды , позволяя рыбе-хозяину питаться жесткими кожистыми красными и коричневыми макроводорослями. [23] Эта трофическая инновация способствовала диверсификации ниш коралловых рифов среди травоядных животных . Рыбы-хирурги имеют решающее значение для функционирования коралловых рифов Индо-Тихоокеанского региона, поскольку они являются одними из немногих рыб, способных потреблять крупные макроводоросли, которые цветут в результате нарушения экосистемы и подавляют восстановление кораллов. [24] [2]

Наряду с более стандартными примерами пищевого симбиоза у животных, недавние достижения в технологии секвенирования генома привели к открытию многих эндосимбиотических ассоциаций у морских простейших (простейший — это общий термин для обозначения немонофилетической коллекции одноклеточных эукариот , которые не являются грибы или в группе Plantae ) Они иллюстрируют включение эндосимбионтами в простейших-хозяев различных новых биохимических функций, таких как фотосинтез, фиксация и рециркуляция азота, а также метаногенез. [25] Эндосимбиоз у простейших широко распространен и представляет собой важный источник инноваций. Регулярно обнаруживаются ранее непризнанные метаболические инновации морских микробных симбиозов, имеющие экологическое значение. [26] Например, Candidatus Kentron (клада Gammaproteobacteria , обнаруженная в сочетании с инфузориями ) питает своих инфузорных хозяев из рода Kentroporos и перерабатывает ацетат и пропионат , которые являются малоценными клеточными отходами своих хозяев, в биомассу . [27] Другой пример — анаэробная морская инфузория Strombidium purpureum . [28] Инфузория живет в анаэробных условиях и является средой обитания эндосимбиотических пурпурных несерных бактерий , которые содержат как бактериохлорофилл а, так и спириллоксантин. Эндосимбионты фотосинтетически активны; следовательно, этот симбиоз представляет собой эволюционный переход аэробного организма в анаэробный с включением органелл . [2]
Размножение и развитие хозяина
[ редактировать ]| Часть серии обзоров по |
| Морская жизнь |
|---|
 |
Выйдя за рамки пищевого симбиоза, микробные симбионты могут изменять воспроизводство, развитие и рост своих хозяев. Специфические бактериальные штаммы в морских биопленках часто напрямую контролируют привлечение планктонных личинок и размножений, либо подавляя расселение, либо служа сигналом расселения. [29] [30] Например, заселение зооспор зеленой водоросли Ulva кишечной на биопленки специфических бактерий опосредовано их притяжением к кворум-чувствительной молекуле, ацил-гомосерин-лактону, секретируемому бактериями. [31] Классические примеры зависимости развития морского хозяина от микроба включают наблюдение, что культуры водорослей, выращенные изолированно, демонстрируют аномальную морфологию. [32] и последующее открытие соединений, индуцирующих морфогенез, таких как таллузин, секретируемых эпифитными бактериальными симбионтами. [33] Известно также, что бактерии влияют на рост морских растений, макроводорослей и фитопланктона, секретируя фитогормоны, такие как индолуксусная кислота и гормоны цитокининового типа. [34] [35] [36] У морской хоанофлагелляты Salpingoeca Rosetta как многоклеточность, так и размножение запускаются специфическими бактериальными сигналами, что позволяет взглянуть на истоки бактериального контроля над развитием животных (обзор Возницы и Кинга. [37] В свою очередь, выгода для бактерий заключается в том, что они получают физическое пространство для колонизации в определенных точках толщи воды, обычно доступное только для планктонных микробов. Пожалуй, наиболее изученным примером тесного взаимодействия хозяина и микроба, контролирующего развитие животных, являются скопы гавайского бобтейла Euprymna. [38] Он живет в мутуалистическом симбиозе с биолюминесцентными бактериями Aliivibrio fischeri. Бактерии получают от хозяина раствор сахаров и аминокислот и, в свою очередь, обеспечивают биолюминесценцию для противозатенения и предотвращения хищников. [7] Этот мутуализм с микробами обеспечивает кальмарам селективное преимущество во взаимодействиях хищник-жертва. Другой пример беспозвоночных можно найти у трубчатых червей, у которых метаморфоз Hydroides elegans опосредован бактериальным индуктором и передачей сигналов митоген-активируемой протеинкиназы (MAPK) в биопленках. [39] [2]
Биообрастание и собрание микробного сообщества
[ редактировать ]
(C) рифообразующий коралл Stylophora pistillata и (D) микроскопическое изображение клеток Endozoicomonas (зондированных на желтый цвет с помощью гибридизации in situ) в щупальцах хозяина S. pistillata .
(E) атлантический киллифиш и (F) СЭМ- изображение поверхности и чешуи рыбы со стрелками, указывающими на клетки бактериального размера, а более крупные клетки (которые не отмечены) предположительно представляют собой фитопланктон.
(G) прорыв горбатого кита и (H) СЭМ-изображение бактерий, связанных с поверхностью кожи горбатого кита, со стрелками, указывающими две разные морфологии клеток. [1]
Некоторые микробы, ассоциированные с хозяином, производят соединения, которые предотвращают биообрастание и регулируют сборку и поддержание микробиома у многих морских организмов, включая губки, макроводоросли и кораллы. [40] [41] Например, тропические кораллы содержат в своем поверхностном слое слизи разнообразные бактерии, которые производят ингибиторы чувства кворума и другие антибактериальные соединения в качестве защиты от колонизации и заражения потенциальными микробными патогенами. [3] Эпифитные бактерии морских макроводорослей выделяют разнообразный химический арсенал, способный избирательно формировать дальнейшую бактериальную колонизацию и сдерживать заселение морскими беспозвоночными, обрастающими биообрастанием, такими как мшанки. [34] [42] Как и в случае с кораллами, эти разнообразные соединения, секретируемые микробами, включают не только бактерицидные и бактериостатические антибиотики, но также такие соединения, как галогенированные фураноны, циклические дипептиды и имитаторы ацил-гомосерин-лактона, которые нарушают ощущение бактериального кворума и ингибируют образование биопленок. [43] Бактерии, вероятно, способны использовать богатые углеродом экссудаты своих хозяев. [44] [45] Например, в случае гигантской бурой водоросли около 20% первичной продукции выделяется в виде растворенного органического углерода. [45] Хотя эти предыдущие примеры иллюстрируют, как микробиомы могут защищать хозяев от поверхностной колонизации, аналогичный феномен также наблюдался внутри корабельного червя Bankia setacea, у которого симбионты производят борированный антибиотик тартролон , который, как считается, защищает слепую кишку, переваривающую древесину, от бактериальных загрязнений. [46] Производя противомикробные соединения, эти микробы способны защищать свою нишу, не позволяя другим организмам вытеснить их. [2]
Биогеохимический циклизм
[ редактировать ]Микробиомы, связанные с хозяином, также влияют на биогеохимический цикл внутри экосистем, оказывая каскадное воздействие на биоразнообразие и экосистемные процессы. Например, микробные симбионты составляют до 40% биомассы губок-хозяев. [47] С помощью процесса, называемого «губчатой петлей», они преобразуют растворенный органический углерод, выделяемый рифовыми организмами, в твердые частицы органического углерода , которые могут потребляться гетеротрофными организмами. [4] Наряду с мутуализмом кораллов и Symbiodiniaceae, этот симбиоз губок и бактерий помогает объяснить парадокс Дарвина, то есть то, как высокопродуктивные экосистемы коралловых рифов существуют в олиготрофных тропических морях. Некоторые симбионты губок играют значительную роль в круговороте морского фосфора, связывая питательные вещества в виде гранул полифосфата в тканях своего хозяина. [48] и круговорот азота, например, посредством нитрификации, денитрификации и окисления аммиака. [4] [41] ]. Многие бактерии, ассоциированные с макроводорослями, специально адаптированы к расщеплению сложных полисахаридов водорослей (например, фукоидана, порфирана и ламинарина). [49] [50] ) и изменить как качество, так и количество органического углерода, поступающего в экосистему. [44] [51] Эндосимбионты жабр люцинид, окисляющие серу, способствуют первичной продуктивности посредством хемосинтеза и способствуют росту морских трав (важных основных видов) за счет снижения концентрации сульфидов в тропических отложениях. [52] Недавно также было показано, что гаммапротеобактериальные симбионты моллюсков-люцинид и стильбонематидных нематод способны к фиксации азота (геномы бактериальных симбионтов кодируют и экспрессируют гены нитрогеназы, [53] подчеркивая роль симбиотических микробов в круговороте питательных веществ в мелководных морских системах. [2]
Эти примеры демонстрируют важность микробных симбиозов для функционирования океанических экосистем. Понимание симбиозов с таким же уровнем детализации в контексте сложных сообществ (т.е. целых микробиомов) остается актуальным для исследования и, действительно, требует более интегрированной структуры из областей микробиологии, эволюционной биологии, экологии сообществ и океанографии. Отдельные таксоны в микробиоме могут помочь хозяевам противостоять широкому спектру условий окружающей среды, в том числе прогнозируемым в рамках сценариев изменения климата. Далее мы исследуем два разных направления того, как междисциплинарное сотрудничество может продвинуть это направление исследований. [2]
Примеры
[ редактировать ]
В настоящее время изучаются микробиомы разнообразных морских животных, от простых организмов, включая губки. [55] и гребневики [56] к более сложным организмам, таким как морские асцидии [57] и акулы. [58] [1]
Отношения между гавайским бобтейлом и биолюминесцентной бактерией Aliivibrio fischeri являются одними из наиболее изученных симбиотических отношений в море и представляют собой систему выбора для общих исследований симбиоза. Эти отношения позволили понять фундаментальные процессы в симбиозах животных и микробов, и особенно биохимические взаимодействия и передачу сигналов между хозяином и бактерией. [59] [60] [1]
Бескишечный морской -олигохета червь Olavius algarvensis — еще один относительно хорошо изученный морской хозяин микробов. Эти трехсантиметровые черви обитают в неглубоких морских отложениях Средиземного моря. У червей нет рта, пищеварительной или выделительной системы, а вместо этого они питаются с помощью набора внеклеточных бактериальных эндосимбионтов, которые живут за счет скоординированного использования серы, присутствующей в окружающей среде. [61] В этой системе используются некоторые из самых сложных инструментов омики и визуализации. [62] Например, зондирование с несколькими метками улучшило визуализацию микробиома. [63] а транскриптомика и протеомика применялись для изучения взаимодействий хозяина и микробиома, включая передачу энергии между хозяином и микробами. [64] и распознавание консорциумов врожденной иммунной системой червя. [65] Основное преимущество этой системы заключается в том, что она дает возможность изучать взаимодействия хозяина и микробиома с микробным консорциумом с низким разнообразием, а также предлагает ряд геномных ресурсов хозяина и микробов. [62] [66] [1]
Кораллы
[ редактировать ]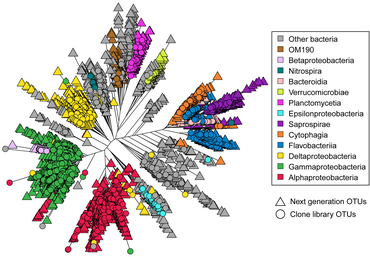
Кораллы являются одним из наиболее распространенных примеров животных-хозяев, чей симбиоз с микроводорослями может превратиться в дисбиоз, что визуально проявляется в обесцвечивании. Коралловые микробиомы изучались в различных исследованиях, которые демонстрируют, как изменения в океанской среде, особенно температура, свет и неорганические питательные вещества, влияют на численность и продуктивность симбионтов микроводорослей, а также на кальцификацию и физиологию хозяина. [68] [69] [1]
Исследования также показали, что резидентные бактерии, археи и грибы дополнительно способствуют круговороту питательных веществ и органических веществ внутри коралла, а вирусы также, возможно, играют роль в структурировании состава этих членов, что дает один из первых проблесков многообразия. домен морских животных симбиоз. [70] Гаммапротеобактерия Endozoicomonas . становится центральным членом микробиома коралла и ведет гибкий образ жизни [71] [72] Учитывая недавнее массовое обесцвечивание рифов, [73] кораллы, вероятно, останутся полезной и популярной системой для исследования симбиоза и дисбиоза. [1]
Astrangia poculata , северный звездчатый коралл, представляет собой каменистый коралл умеренного пояса , широко встречающийся вдоль восточного побережья Соединенных Штатов. Коралл может жить как с зооксантеллами (водорослевыми симбионтами), так и без них, что делает его идеальным модельным организмом для изучения взаимодействий микробного сообщества, связанных с симбиотическим состоянием. Однако возможность разработки праймеров и зондов для более специфичного воздействия на ключевые микробные группы затруднена из-за отсутствия полноразмерных последовательностей 16S рРНК , поскольку последовательности, полученные с помощью платформы Illumina, имеют недостаточную длину (около 250 пар оснований) для создания праймеры и зонды. [74] В 2019 году Голдсмит и др. продемонстрировали, что секвенирование по Сэнгеру способно воспроизводить биологически значимое разнообразие, обнаруженное с помощью более глубокого секвенирования следующего поколения , а также создавать более длинные последовательности, полезные исследовательскому сообществу для разработки зондов и праймеров (см. Диаграмму справа). [75]
Губки
[ редактировать ]Губки являются обычными представителями разнообразных донных сред обитания в океане, и их численность и способность фильтровать большие объемы морской воды привели к осознанию того, что эти организмы играют решающую роль во влиянии на бентосные и пелагические процессы в океане. [76] Они являются одной из старейших линий животных и имеют относительно простое строение тела, которое обычно ассоциируется с бактериями, архей, водорослями-протистами, грибами и вирусами. [4] Микробиомы губок состоят из специалистов и специалистов широкого профиля, и сложность их микробиома, по-видимому, определяется филогенией хозяина. [77] Исследования показали, что микробиом губок способствует круговороту азота в океанах, особенно за счет окисления аммиака архей и бактерий. [78] [79] Совсем недавно было показано, что микробные симбионты тропических губок производят и запасают гранулы полифосфата. [80] возможно, позволяя хозяину пережить периоды истощения фосфатов в олиготрофной морской среде. [81] Микробиомы некоторых видов губок действительно меняются в структуре сообщества в ответ на изменение условий окружающей среды, включая температуру. [82] и закисление океана, [83] [84] а также синергетический эффект. [85]

Китообразные
[ редактировать ]
Доступ к образцам микробов из кишечника морских млекопитающих ограничен, поскольку большинство видов являются редкими, исчезающими и ныряют на большие глубины. Существуют различные методы отбора проб микробиома кишечника китообразных. Наиболее распространенным является сбор образцов фекалий из окружающей среды и взятие зонда из незагрязненного центра. [88] Кроме того, есть исследования с ректальных мазков и редкие исследования с мертвых или живых животных прямо из кишечника. [89] [90] [91]
Самый внешний эпидермальный слой, то есть кожа, является первым барьером, защищающим человека от внешнего мира, а эпидермальный микробиом на нем считается индикатором не только здоровья животного, но и экологическим индикатором, показывающим состояние окружающей среды. Знание микробиома кожи морских млекопитающих в «нормальных» условиях позволило нам понять, чем эти сообщества отличаются от свободных микробных сообществ, обнаруженных в море, и как они могут меняться в зависимости от абиотических и биотических вариаций, а также « Сообщества различаются между здоровыми и больными людьми» . [92]
Китообразные находятся в опасности, поскольку на них воздействуют многочисленные стрессовые факторы, которые делают их более уязвимыми к различным заболеваниям. Было отмечено, что эти животные демонстрируют высокую восприимчивость к инфекциям дыхательных путей, но об их респираторном микробиоме известно очень мало. Следовательно, отбор проб выдыхаемого воздуха или «вдоха» китообразных может дать оценку состояния здоровья. Удар состоит из смеси микроорганизмов и органического материала , включая липиды , белки и клеточный мусор, образующийся из слизистой оболочки дыхательных путей, которые при попадании в относительно более прохладный наружный воздух конденсируются с образованием видимой массы пара, которая может собрано. Существуют различные методы сбора проб выдыхаемого воздуха, один из последних — использование воздушных дронов. Этот метод обеспечивает более безопасную, тихую и менее инвазивную альтернативу, а зачастую и экономически эффективный вариант мониторинга фауны и флоры. После получения пробы доставляются в лабораторию, и мы приступаем к амплификации и секвенированию микробиоты дыхательных путей. Использование воздушных дронов оказалось более успешным для крупных китообразных из-за низкой скорости плавания и большего размера ударов. [87] [93] [94] [95] [96] [86] [97] [98] [99]
Морские холобионты
[ редактировать ]Кораллы, строящие рифы, представляют собой холобионты, которые включают сам коралл (эукариотические беспозвоночные класса Anthozoa ), фотосинтезирующие динофлагелляты, называемые зооксантеллами ( Symbiodinium ), и связанные с ними бактерии и вирусы. [100] Существуют модели коэволюции коралловых микробных сообществ и филогении кораллов. [101]
![Коралловый холобионт[102]](http://upload.wikimedia.org/wikipedia/commons/thumb/2/26/Trophic_connections_of_the_coral_holobiont_in_the_planktonic_food_web.jpg/271px-Trophic_connections_of_the_coral_holobiont_in_the_planktonic_food_web.jpg)
![Голобионт морских водорослей [103]](http://upload.wikimedia.org/wikipedia/commons/thumb/3/35/Processes_within_the_seagrass_holobiont.webp/225px-Processes_within_the_seagrass_holobiont.webp.png)
![Губка холобионт[41]](http://upload.wikimedia.org/wikipedia/commons/thumb/6/6b/The_sponge_holobiont.webp/347px-The_sponge_holobiont.webp.png)
![Изменение климата и родолитовый холобионт[104]](http://upload.wikimedia.org/wikipedia/commons/thumb/f/f7/Climate_change_stressors_and_rhodolith_holobiont_fitness.webp/474px-Climate_change_stressors_and_rhodolith_holobiont_fitness.webp.png)
Ссылки
[ редактировать ]- ^ Jump up to: а б с д и ж г час Апприл, А. (2017) «Микробиомы морских животных: к пониманию взаимодействия хозяина и микробиома в меняющемся океане». Границы морской науки , 4 : 222. два : 10.3389/fmars.2017.00222 .
 Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 . - ^ Jump up to: а б с д и ж г час я Уилкинс Л.Г., Лерей М., О'Ди А., Юэнь Б., Пейшото Р.С., Перейра Т.Дж. и др. (2019). «Связанные с хозяином микробиомы определяют структуру и функционирование морских экосистем» . ПЛОС Биология . 17 (11): e3000533. дои : 10.1371/journal.pbio.3000533 . ПМК 6874084 . ПМИД 31710600 . Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Jump up to: а б Кредит CJ, Ричи КБ, Пол В.Дж., Теплицкий М (2013). «Микроорганизмы, связанные с кораллами, и их роль в укреплении здоровья кораллов и предотвращении болезней» . Труды Королевского общества B: Биологические науки . 280 (1755). дои : 10.1098/rspb.2012.2328 . ПМЦ 3574386 . ПМИД 23363627 .
- ^ Jump up to: а б с д Вебстер Н.С., Томас Т. (2016). «Губка гологеном» . мБио . 7 (2): e00135-16. дои : 10.1128/mBio.00135-16 . ПМК 4850255 . ПМИД 27103626 .
- ^ Дистел Д.Л., Делонг Э.Ф., Уотербери Дж.Б. (1991). «Филогенетическая характеристика и локализация in situ бактериального симбионта корабельных червей (Teredinidae: Bivalvia) с использованием анализа последовательности 16S рРНК и гибридизации олигодезоксинуклеотидных зондов» . Прикладная и экологическая микробиология . 57 (8): 2376–2382. Бибкод : 1991ApEnM..57.2376D . дои : 10.1128/АЕМ.57.8.2376-2382.1991 . ПМК 183578 . ПМИД 1722662 .
- ^ Рюланд С., Блазжак А., Лотт С., Лой А., Эрсеус С., Дюбилье Н. (2008). «Множественные бактериальные симбионты в двух видах одновременно встречающихся бескишечных червей-олигохет из отложений трав Средиземного моря». Экологическая микробиология . 10 (12): 3404–3416. Бибкод : 2008EnvMi..10.3404R . дои : 10.1111/j.1462-2920.2008.01728.x . ПМИД 18764872 .
- ^ Jump up to: а б Нихолм С.В., Макфолл-Нгай М. (2004). «Веяние: установление симбиоза кальмара и вибриона». Обзоры природы Микробиология . 2 (8): 632–642. дои : 10.1038/nrmicro957 . ПМИД 15263898 . S2CID 21583331 .
- ^ Хаммер Т.Дж., Сандерс Дж.Г., Фирер Н. (2019). «Не всем животным нужен микробиом». Письма FEMS по микробиологии . 366 (10). дои : 10.1093/femsle/fnz117 . ПМИД 31132110 .
- ^ Типтон Л., Дарси Дж.Л., Хинсон Н.А. (2019). «Развивающийся симбиоз: обеспечение перекрестного общения между экологами и учеными-микробиомами» . Границы микробиологии . 10 : 292. дои : 10.3389/fmicb.2019.00292 . ПМК 6391321 . ПМИД 30842763 .
- ^ Апприл А (2017). «Микробиомы морских животных: к пониманию взаимодействия хозяина и микробиома в меняющемся океане» . Границы морской науки . 4 . дои : 10.3389/fmars.2017.00222 . S2CID 9729436 .
- ^ Jump up to: а б Макфолл-Нгай М., Хэдфилд М.Г., Бош Т.С., Кэри Х.В., Домазет-Лошо Т., Дуглас А.Е. и др. (2013). «Животные в бактериальном мире: новый императив для наук о жизни» . Труды Национальной академии наук . 110 (9): 3229–3236. Бибкод : 2013PNAS..110.3229M . дои : 10.1073/pnas.1218525110 . ПМЦ 3587249 . ПМИД 23391737 .
- ^ Вебстер Н.С., Тейлор М.В. (2012). «Морские губки и их микробные симбионты: Любовь и другие взаимоотношения» . Экологическая микробиология . 14 (2): 335–346. Бибкод : 2012EnvMi..14..335W . дои : 10.1111/j.1462-2920.2011.02460.x . ПМИД 21443739 .
- ^ Азам Ф., Уорден АЗ (2004). «ОКЕАНОГРАФИЯ: микробы, молекулы и морские экосистемы». Наука . 303 (5664): 1622–1624. дои : 10.1126/science.1093892 . ПМИД 15016987 . S2CID 10101482 .
- ^ Кавиччиоли Р., Риппл В.Дж., Тиммис К.Н., Азам Ф., Баккен Л.Р., Бэйлис М. и др. (2019). «Предупреждение ученых человечеству: микроорганизмы и изменение климата» . Обзоры природы Микробиология . 17 (9): 569–586. дои : 10.1038/s41579-019-0222-5 . ПМЦ 7136171 . ПМИД 31213707 .
- ^ Даффи Дж. Э., Годвин СМ, Кардинале Б. Дж. (2017). «Воздействие биоразнообразия в дикой природе является обычным явлением и столь же сильным, как и ключевые факторы производительности». Природа . 549 (7671): 261–264. Бибкод : 2017Natur.549..261D . дои : 10.1038/nature23886 . ПМИД 28869964 . S2CID 4459856 .
- ^ Кардинале Б.Дж., Даффи Дж.Э., Гонсалес А., Хупер Д.Ю., Перрингс С., Венаил П. и др. (2012). «Утрата биоразнообразия и ее влияние на человечество» (PDF) . Природа . 486 (7401): 59–67. Бибкод : 2012Natur.486...59C . дои : 10.1038/nature11148 . ПМИД 22678280 . S2CID 4333166 .
- ^ Гулд А.Л., Чжан В., Ламберти Л., Джонс Э.В., Обадиа Б., Корасидис Н. и др. (2018). «Взаимодействие микробиома формирует приспособленность хозяина» . Труды Национальной академии наук . 115 (51): Е11951–Е11960. Бибкод : 2018PNAS..11511951G . дои : 10.1073/pnas.1809349115 . ПМК 6304949 . ПМИД 30510004 .
- ^ Зееманн Дж., Ингст А., Стюарт-Смит Р.Д., Эдгар Г.Дж., Алтьери А.Х. (2018). «Важность губок и мангровых зарослей в поддержке рыбных сообществ деградировавших коралловых рифов в Карибской Панаме» . ПерДж . 6 : е4455. дои : 10.7717/peerj.4455 . ПМЦ 5878927 . ПМИД 29610704 .
- ^ о'Брайен П.А., Вебстер Н.С., Миллер Д.Д., Борн Д.Г. (2019). «Коэволюция хозяина и микроба: применение данных модельных систем к сложным холобионтам морских беспозвоночных» . мБио . 10 (1). дои : 10.1128/mBio.02241-18 . ПМК 6428750 . ПМИД 30723123 .
- ^ Крофт М.Т., Лоуренс А.Д., Ро-Дири Э., Уоррен М.Дж., Смит А.Г. (2005). «Водоросли приобретают витамин B12 посредством симбиотических отношений с бактериями». Природа . 438 (7064): 90–93. Бибкод : 2005Natur.438...90C . дои : 10.1038/nature04056 . ПМИД 16267554 . S2CID 4328049 .
- ^ Куигли К.М., Бэй Л.К., Уиллис Б.Л. (2018). «Использование новых знаний о регулировании сообщества Symbiodinium кораллов для сохранения и восстановления рифов». Серия «Прогресс в области морской экологии» . 600 : 245–253. Бибкод : 2018MEPS..600..245Q . дои : 10.3354/meps12652 . S2CID 90469901 .
- ^ Бейкер Д.М., Фриман С.Дж., Вонг Дж.К., Фогель М.Л., Ноултон Н. (2018). «Изменение климата способствует паразитизму в коралловом симбиозе» . Журнал ISME . 12 (3): 921–930. Бибкод : 2018ISMEJ..12..921B . дои : 10.1038/s41396-018-0046-8 . ПМК 5864192 . ПМИД 29379177 .
- ^ Нгуги Д.К., Мияке С., Кэхилл М., Вину М., Хакманн Т.Дж., Блом Дж. и др. (2017). «Геномная диверсификация гигантских кишечных симбионтов отражает образ жизни хозяина» . Труды Национальной академии наук . 114 (36): E7592–E7601. Бибкод : 2017PNAS..114E7592N . дои : 10.1073/pnas.1703070114 . ПМЦ 5594648 . ПМИД 28835538 .
- ^ Хоуи А.С., Беллвуд ДР (2009). «Ограниченная функциональная избыточность в системе с высоким разнообразием: один вид доминирует в ключевом экологическом процессе на коралловых рифах». Экосистемы . 12 (8): 1316–1328. Бибкод : 2009Ecosy..12.1316H . дои : 10.1007/s10021-009-9291-z . S2CID 42138428 .
- ^ Новак ЕС, Мелконян М (2010). «Эндосимбиотические ассоциации внутри простейших» . Философские труды Королевского общества B: Биологические науки . 365 (1541): 699–712. дои : 10.1098/rstb.2009.0188 . ПМК 2817226 . ПМИД 20124339 .
- ^ Хиггс Н.Д., Ньютон Дж., Аттрилл М.Дж. (2016). «Карибский промысел лангустов поддерживается трофическими субсидиями от хемосинтетического первичного производства» . Современная биология . 26 (24): 3393–3398. Бибкод : 2016CBio...26.3393H . дои : 10.1016/j.cub.2016.10.034 . hdl : 10026.1/9129 . ПМИД 27939312 . S2CID 14401680 .
- ^ Сеа Б.К., Энтони К.П., Хюттель Б., Заржицкий Дж., Шада фон Боржисковски Л., Эрб Т.Дж. и др. (2019). «Сераокисляющие симбионты без канонических генов автотрофной фиксации CO2» . мБио . 10 (3). дои : 10.1128/mBio.01112-19 . ПМК 6593406 . ПМИД 31239380 .
- ^ Гаст Р.Дж., Сандерс Р.В., Кэрон Д.А. (2009). «Экологические стратегии протистов и их симбиотические отношения с прокариотными микробами». Тенденции в микробиологии . 17 (12): 563–569. дои : 10.1016/j.tim.2009.09.001 . ПМИД 19828317 .
- ^ Хуан Ю, Каллахан С., Хэдфилд М.Г. (2012). «Вербовка в море: бактериальные гены, необходимые для индукции заселения личинок многощетинковых червей» . Научные отчеты . 2 : 228. Бибкод : 2012NatSR...2E.228H . дои : 10.1038/srep00228 . ПМК 3260340 . ПМИД 22355742 . S2CID 14731587 .
- ^ Деланной С., Чавес Б.Д., Изон С.А., Уэбб Х.Э., Беутин Л., Делаваль Дж. и др. (2016). «Пересмотр подхода к тестированию STEC: использование espK и espV для повышения надежности обнаружения энтерогеморрагической Escherichia coli (EHEC) в говядине» . Границы микробиологии . 7 : 1. дои : 10.3389/fmicb.2016.00001 . ПМЦ 4722105 . ПМИД 26834723 .
- ^ Уиллер Г.Л., Тейт К., Тейлор А., Браунли С., Джойнт I (2006). «Ацил-гомосериновые лактоны модулируют скорость заселения зооспор морской водоросли Ulva кишечной с помощью нового хемокинетического механизма» . Растение, клетка и окружающая среда . 29 (4): 608–618. дои : 10.1111/j.1365-3040.2005.01440.x . ПМИД 17080611 .
- ^ Провасоли Л., Пинтнер И.Дж. (1980). «Вызванный бактериями полиморфизм в аксеническом лабораторном штамме Ulva Lactuca (Chlorophyceae)1». Журнал психологии . 16 (2): 196–201. Бибкод : 1980JPcgy..16..196P . дои : 10.1111/j.1529-8817.1980.tb03019.x . S2CID 85817449 .
- ^ Мацуо Ю, Имагава Х, Нисидзава М, Сидзури Ю (2005). «Выделение индуктора морфогенеза водорослей из морской бактерии». Наука . 307 (5715): 1598. doi : 10.1126/science.1105486 . ПМИД 15761147 . S2CID 28850526 .
- ^ Jump up to: а б Гёке Ф., Лабес А., Визе Дж., Имхофф Дж. Ф. (2010). «Химические взаимодействия морских макроводорослей и бактерий» . Серия «Прогресс в области морской экологии» . 409 : 267–299. Бибкод : 2010MEPS..409..267G . дои : 10.3354/meps08607 .
- ^ Амин С.А., Хмело Л.Р., Ван Тол Х.М., Дарем Б.П., Карлсон Л.Т., Хил КР и др. (2015). «Взаимодействие и передача сигналов между космополитическим фитопланктоном и связанными с ним бактериями». Природа . 522 (7554): 98–101. Бибкод : 2015Natur.522...98A . дои : 10.1038/nature14488 . ПМИД 26017307 . S2CID 4462055 .
- ^ Селдран Д., Эспиноса Э., Санчес-Амат А., Марин А. (2012). «Влияние эпибиотических бактерий на рост листьев и эпифиты морской травы Posidonia Oceanica» . Серия «Прогресс в области морской экологии» . 456 : 21–27. Бибкод : 2012MEPS..456...21C . дои : 10.3354/meps09672 .
- ^ Возница А, Король Н (2018). «Уроки простых морских моделей бактериальной регуляции развития эукариот» . Современное мнение в микробиологии . 43 : 108–116. дои : 10.1016/j.mib.2017.12.013 . ПМК 6051772 . ПМИД 29331767 .
- ^ Макфолл-Нгай, MJ (2014). «Важность микробов в развитии животных: уроки симбиоза кальмаров и вибрионов» . Ежегодный обзор микробиологии . 68 : 177–194. doi : 10.1146/annurev-micro-091313-103654 . ПМК 6281398 . ПМИД 24995875 .
- ^ Шикума Н.Дж., Антошечкин И., Медейрос Дж.М., Пилхофер М., Ньюман Д.К. (2016). «Пошаговая метаморфоза трубчатого червя Hydroides elegansis, опосредованная бактериальным индуктором и передачей сигналов МАРК» . Труды Национальной академии наук . 113 (36): 10097–10102. Бибкод : 2016PNAS..11310097S . дои : 10.1073/pnas.1603142113 . ПМК 5018781 . ПМИД 27551098 . S2CID 23501584 .
- ^ Кредитет С.Дж., Карпиноне Э.М., Ричи К.Б., Теплицкий М. (2013). «Характеристика тегакА-зависимой колонизации поверхности и коралловой слизи условно-патогенным коралловым патогеном Serratia marcescensPDL100» . ФЭМС Микробиология Экология . 84 (2): 290–301. Бибкод : 2013FEMME..84..290K . дои : 10.1111/1574-6941.12064 . ПМИД 23278392 .
- ^ Jump up to: а б с Пита Л., Рикс Л., Слаби Б.М., Франке А., Хентшель Ю. (март 2018 г.). «Губка-голобионт в меняющемся океане: от микробов к экосистемам» . Микробиом . 6 (1): 46. дои : 10.1186/s40168-018-0428-1 . ПМЦ 5845141 . ПМИД 29523192 . Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Стейнберг П.Д., Де Нис Р. (2002). «Химическое опосредование колонизации поверхности морских водорослей1». Журнал психологии . 38 (4): 621–629. Бибкод : 2002JPcgy..38..621S . дои : 10.1046/j.1529-8817.2002.02042.x . S2CID 83963124 .
- ^ Добрецов С., Абед Р.М., Теплицкий М. (2013). «Мини-обзор: Ингибирование биообрастания морскими микроорганизмами» . Биологическое обрастание . 29 (4): 423–441. Бибкод : 2013Biofo..29..423D . дои : 10.1080/08927014.2013.776042 . ПМИД 23574279 . S2CID 34459128 .
- ^ Jump up to: а б Рид Д.С., Карлсон К.А., Хейлвуд Э.Р., Нельсон Дж.К., Харрер С.Л., Рассвайлер А. и др. (2015). «Схемы и меры контроля производства растворенного органического углерода в масштабах рифов гигантскими водорослями M acrocystispyrifera» . Лимнология и океанография . 60 (6): 1996–2008. Бибкод : 2015LimOc..60.1996R . дои : 10.1002/lno.10154 . S2CID 85962482 .
- ^ Jump up to: а б Фоллмерс Дж., Виганд С., Кастер А.К. (2017). «Сравнение и оценка инструментов сборки метагенома с точки зрения микробиолога - не только размер имеет значение!» . ПЛОС ОДИН . 12 (1): e0169662. Бибкод : 2017PLoSO..1269662V . дои : 10.1371/journal.pone.0169662 . ПМЦ 5242441 . ПМИД 28099457 .
- ^ Эльшахави С.И., Триндаде-Сильва А.Е., Ханора А., Хан А.В., Флорес М.С., Виццони В. и др. (2013). «Борированный антибиотик тартролон, вырабатываемый симбиотическими бактериями, разлагающими целлюлозу, в жабрах корабельных червей» . Труды Национальной академии наук . 110 (4): E295–E304. дои : 10.1073/pnas.1213892110 . ПМЦ 3557025 . ПМИД 23288898 .
- ^ Де Гоэй Дж. М., Ван Овелен Д., Вермей М. Дж., Осинга Р., Мидделбург Дж. Дж., Де Гоэй А. Ф. и др. (2013). «Выживание в морской пустыне: губчатая петля сохраняет ресурсы внутри коралловых рифов». Наука . 342 (6154): 108–110. Стартовый код : 2013Sci...342..108D . дои : 10.1126/science.1241981 . ПМИД 24092742 . S2CID 6720678 .
- ^ Колман А.С. (2015). «Губковые симбионты и морской П-цикл» . Труды Национальной академии наук . 112 (14): 4191–4192. Бибкод : 2015PNAS..112.4191C . дои : 10.1073/pnas.1502763112 . ПМЦ 4394276 . ПМИД 25825737 .
- ^ Корцетт Ч., Эльшербини Дж., Чиен Д.М., Хехеманн Дж.Х., Хеншель А., Прехейм С.П. и др. (2018). «Эволюция вегетарианского вибриона: метаболическая специализация Vibrio breoganii к субстратам макроводорослей» . Журнал бактериологии . 200 (15). дои : 10.1128/JB.00020-18 . ПМК 6040190 . ПМИД 29632094 .
- ^ Бенгтссон М.М., Сьётун К., Сторесунд Дж.Э., Овреас Дж. (2011). «Использование источников углерода, полученных из водорослей, бактериями, ассоциированными с поверхностью водорослей» . Водная микробная экология . 62 (2): 191–199. дои : 10.3354/ame01477 . HDL : 1956/4610 .
- ^ Пфистер К.А., Альтабет М.А., Вейгель Б.Л. (2019). «Заросли водорослей и их местное влияние на химический состав морской воды, продуктивность и микробные сообщества». Экология . 100 (10): e02798. Бибкод : 2019Ecol..100E2798P . дои : 10.1002/ecy.2798 . ПМИД 31233610 . S2CID 195355739 .
- ^ Ван дер Хайде Т., Говерс Л.Л., Де Фув Дж., Олфф Х., Ван дер Гест М., Ван Катвейк М.М. и др. (2012). «Трехэтапный симбиоз формирует основу экосистем морских водорослей» . Наука . 336 (6087): 1432–1434. Бибкод : 2012Sci...336.1432V . дои : 10.1126/science.1219973 . hdl : 11370/23625acb-7ec0-4480-98d7-fad737d7d4fe . ПМИД 22700927 . S2CID 27806510 .
- ^ Петерсен Дж.М., Кемпер А., Грубер-Водика Х., Кардини У., Ван дер Гест М., Кляйнер М. и др. (2017). «Хемосинтезирующие симбионты морских беспозвоночных животных способны к азотфиксации» . Природная микробиология . 2 (1): 16195. doi : 10.1038/nmicrobiol.2016.195 . ПМК 6872982 . ПМИД 27775707 .
- ^ Пейшото, Р.С., Росадо, П.М., Лейте, DCDA, Росадо, А.С. и Борн, Д.Г. (2017) «Полезные микроорганизмы для кораллов (BMC): предлагаемые механизмы здоровья и устойчивости кораллов». Границы микробиологии , 8 : 341. два : 10.3389/fmicb.2017.00341 .
- ^ Вебстер, Н.С., Негри, А.П., Ботте, Э.С., Лаффи, П.В., Флорес, Ф., Нунан, С., Шмидт, К. и Утик, С. (2016) «Микробы коралловых рифов, связанные с хозяином, реагируют на кумулятивные давление потепления и закисления океана». Научные отчеты , 6 : 19324. дои : 10.1038/srep19324 .
- ^ Дэниэлс, К. и Брейтбарт, М. (2012) «Бактериальные сообщества, связанные с гребневиками Mnemiopsis leidyi и Beroe ovata ». FEMS Микробиология Экология , 82 (1): 90–101. дои : 10.1111/j.1574-6941.2012.01409.x .
- ^ Блазиак, Л.К., Зиндер, С.Х., Бакли, Д.Х. и Хилл, RT (2014) «Бактериальное разнообразие, связанное с туникой модельной хордовой Ciona кишечной ». Журнал ISME , 8 (2): 309–320. два : 10.1038/ismej.2013.156 .
- ^ Гивенс, К.Э., Рэнсом, Б., Бано, Н. и Холлибо, Дж.Т. (2015) «Сравнение кишечных микробиомов 12 костистых рыб и 3 видов акул». Серия достижений морской экологии , 518 : 209–223. два : 10.3354/meps11034 .
- ^ Макфолл-Нгай, MJ (2000) «Переговоры между животными и бактериями:« дипломатия »симбиоза кальмара и вибриона». Сравнительная биохимия и физиология , Часть A: Молекулярная и интегративная физиология , 126 (4): 471–480. два : 10.1016/S1095-6433(00)00233-6 .
- ^ Макфолл-Нгай, М. (2014) «Разгадка сущности симбиоза: выводы из модели кальмар-вибрион». PLoS Biology , 12 (2): e1001783. дои : 10.1371/journal.pbio.1001783 .
- ^ Дюбилье, Н., Мюлдерс, К., Фердельман, Т., де Бир, Д., Пернталер, А., Кляйн, М., Вагнер, М., Эрсеус, К., Тирманн, Ф., Кригер, Дж. и Жир О. (2001) «Эндосимбиотические сульфатредуцирующие и сульфидокисляющие бактерии у олигохетных червей». Природа , 411 (6835): 298–302. дои : 10.1038/35077067 .
- ^ Jump up to: а б Войк Т., Тилинг Х., Иванова Н.Н., Хантеманн М., Рихтер М., Глекнер Ф.О., Боффелли Д., Андерсон И.Дж., Бэрри К.В., Шапиро Х.Дж. и Сзето Э. ( 2006) «Понимание симбиоза посредством метагеномного анализа микробного консорциума». Природа , 443 (7114): 950–955. дои : 10.1038/nature05192 .
- ^ Шимак, М.П., Кляйнер, М., Ветцель, С., Либеке, М., Дубилье, Н. и Фукс, Б.М. (2016) «MiL-FISH: мультимеченые олигонуклеотиды для флуоресцентной гибридизации in situ улучшают визуализацию бактериальных клеток». Прикладная и экологическая микробиология , 82 (1): 62–70. дои : 10.1128/АЕМ.02776-15 .
- ^ Кляйнер, М., Вентруп, К., Лотт, К., Тилинг, Х., Ветцель, С., Янг, Дж., Чанг, Ю.Дж., Шах, М., Верберкмоэс, Северная Каролина, Заржицкий, Дж. и Фукс , Г. (2012) «Метапротеомика бескишечного морского червя и его симбиотического микробного сообщества раскрывает необычные пути использования углерода и энергии». Труды Национальной академии наук , 109 (19): E1173–E1182. дои : 10.1073/pnas.1121198109 .
- ^ Випплер, Дж., Кляйнер, М., Лотт, К., Грул, А., Абрахам, П.Е., Джанноне, Р.Дж., Янг, Дж.К., Хеттич, Р.Л. и Дубилье, Н. (2016) «Транскриптомные и протеомные взгляды на врожденный иммунитет и приспособления к симбиотическому образу жизни у бескишечного морского червя Olavius algarvensis ». BMC Genomics , 17 (1): 942. дои : 10.1186/s12864-016-3293-y .
- ^ Рюланд, К., Блазежак, А., Лотт, К., Лой, А., Эрсеус, К. и Дюбилье, Н. (2008) «Множественные бактериальные симбионты в двух видах одновременно встречающихся бескишечных червей-олигохет из Средиземного моря». травяные отложения». Экологическая микробиология , 10 (12): 3404–3416. дои : 10.1111/j.1462-2920.2008.01728.x .
- ^ Шарп К.Х., Пратте З.А., Кервин А.Х., Ротьян Р.Д., Стюарт Ф.Дж. (2017). «Структурой микробиома коралла Astrangia poculata умеренного пояса управляет сезон, а не состояние симбионта» . Микробиом . 5 (1): 120. дои : 10.1186/s40168-017-0329-8 . ПМК 5603060 . ПМИД 28915923 .
- ^ Дубинский З. и Джокиел П.Л. (1994) «Соотношение потоков энергии и питательных веществ регулирует симбиоз между зооксантеллами и кораллами». Pacific Science , 48 (3): 313–324.
- ^ Энтони, К.Р., Клайн, Д.И., Диас-Пулидо, Г., Дав, С. и Хоег-Гульдберг, О. (2008) «Закисление океана вызывает обесцвечивание и потерю производительности строителей коралловых рифов». Труды Национальной академии наук , 105 (45): 17442–17446. дои : 10.1073/pnas.0804478105 .
- ^ Борн, Д.Г., Морроу, К.М. и Вебстер, Н.С. (2016) «Изучение кораллового микробиома: поддержка здоровья и устойчивости рифовых экосистем». Ежегодный обзор микробиологии , 70 : 317–340. doi : 10.1146/annurev-micro-102215-095440 .
- ^ Нив, М.Дж., Апприлль, А., Феррье-Пажес, К. и Вулстра, К.Р. (2016) «Разнообразие и функции распространенных симбиотических морских бактерий рода Endozoicomonas ». Прикладная микробиология и биотехнология , 100 (19): 8315–8324. два : 10.1007/s00253-016-7777-0 .
- ^ Нив, М.Дж., Мичелл, К.Т., Аппилл, А. и Вулстра, К.Р. (2017) «Геномы Endozoicomonas обнаруживают функциональную адаптацию и пластичность бактериальных штаммов, симбиотически связанных с различными морскими хозяевами». Научные отчеты , 7 : 40579. дои : 10.1038/srep40579 .
- ^ Хьюз, Т.П., Керри, Дж.Т., Альварес-Норьега, М., Альварес-Ромеро, Дж.Г., Андерсон, К.Д., Бэрд, А.Х., Бэбкок, Р.С., Бегер, М., Беллвуд, Д.Р., Беркельманс, Р. и Бридж, ТК (2017) «Глобальное потепление и периодическое массовое обесцвечивание кораллов». Природа , 543 (7645): 373–377. дои : 10.1038/nature21707 .
- ^ Ученые Геологической службы США публикуют давно читаемые последовательности микробиома кораллов умеренного пояса, предоставляя сообществу ресурсы для разработки зондов и праймеров , Геологическая служба США , 6 марта 2019 г. Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ b Голдсмит Д., a Пратте З., a Келлог С., e Снадер С., h Шарп К. (2019). «Стабильность микробиома коралла Astrangia poculata умеренного пояса отражается в различных методологиях секвенирования» . АИМС Микробиология . 5 (1): 62–76. дои : 10.3934/микробиол.2019.1.62 . ПМК 6646935 . ПМИД 31384703 . Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Белл, Дж. Дж. (2008) «Функциональная роль морских губок». Эстуарий, Наука о прибрежных зонах и шельфе , 79 (3): 341–353. дои : 10.1016/j.ecss.2008.05.002 .
- ^ Томас, Т., Мойтиньо-Сильва, Л., Лурги, М., Бьорк, Дж. Р., Иссон, К., Астудильо-Гарсия, К., Олсон, Дж. Б., Эрвин, П. М., Лопес-Легентиль, С., Лютер , Х. и Чавес-Фоннегра, А. (2016) «Разнообразие, структура и конвергентная эволюция глобального микробиома губок». Природные коммуникации , 7 (1): 1-12. дои : 10.1038/ncomms11870 .
- ^ Байер К., Шмитт С. и Хентшель У. (2008) «Физиология, филогения и данные in situ о бактериальных и архейных нитрификаторах в морской губке Aplysina aerophoba ». Экологическая микробиология , 10 (11): 2942–2955. дои : 10.1111/j.1462-2920.2008.01582.x .
- ^ Радакс Р., Хоффманн Ф., Рапп Х.Т., Лейнингер С. и Шлепер К. (2012) «Археи, окисляющие аммиак, как основные движущие силы нитрификации в холодноводных губках». Экологическая микробиология , 14 (4): 909_923. дои : 10.1111/j.1462-2920.2011.02661.x .
- ^ Чжан Ф., Блазиак, Л.К., Каролин, Дж.О., Пауэлл, Р.Дж., Геддес, К.Д. и Хилл, RT (2015) «Связывание фосфора в форме полифосфата микробными симбионтами в морских губках». Труды Национальной академии наук , 112 (14): 4381–4386. дои : 10.1073/pnas.1423768112 .
- ^ Колман, А.С. (2015) «Губковые симбионты и морской P-цикл». Труды Национальной академии наук , 112 (14): 4191–4192. дои : 10.1073/pnas.1502763112 .
- ^ Симистер Р., Тейлор М.В., Цай П., Фан Л., Брюснер Т.Дж., Кроу М.Л. и Вебстер Н. (2012) «Реакции на термический стресс в бактериальной биосфере губки Большого Барьерного рифа, Rhopaloeides odorabile . Микробиология окружающей среды , 14 (12): 3232–3246. дои : 10.1111/1462-2920.12010 .
- ^ Морроу, К.М., Борн, Д.Г., Хамфри, К., Ботте, Э.С., Лаффи, П., Заневельд, Дж., Утик, С., Фабрициус, К.Э. и Вебстер, Н.С. (2015) «Природные вулканические выходы CO 2 обнаруживают будущие траектории ассоциаций хозяина и микробов в кораллах и губках». Журнал ISME , 9 (4): 894–908. два : 10.1038/ismej.2014.188 .
- ^ Рибес М., Кальво Э., Мовилла Дж., Логарес Р., Кома Р. и Пелейеро К. (2016) «Реструктуризация микробиома губок способствует устойчивости к закислению океана . Отчеты по экологической микробиологии , 8 (4): 536–544. дои : 10.1111/1758-2229.12430 .
- ^ Лессер, М.П., Фиоре, К., Слэттери, М. и Заневельд, Дж. (2016) «Стрессоры изменения климата дестабилизируют микробиом карибской бочковой губки, Xestospongia muta ». Журнал экспериментальной морской биологии и экологии , 475 : 11–18. дои : 10.1016/j.jembe.2015.11.004 .
- ^ Jump up to: а б Асеведо-Уайтхаус К., Роча-Госселин А., Гендрон Д. (апрель 2010 г.). «Новый неинвазивный инструмент для наблюдения за болезнями китов, находящихся на свободном выгуле, и его актуальность для программ сохранения». Охрана животных . 13 (2): 217–225. дои : 10.1111/j.1469-1795.2009.00326.x . S2CID 86518859 .
- ^ Jump up to: а б Пиротта В., Смит А., Островски М., Рассел Д., Йонсен И.Д., Греч А. и др. (декабрь 2017 г.). «Экономичный дрон, изготовленный по индивидуальному заказу для оценки здоровья китов» . Границы морской науки . 4 : 425. дои : 10.3389/fmars.2017.00425 .
- ^ Сузуки А, Уэда К, Сегава Т, Сузуки М (июнь 2019 г.). «Фекальная микробиота антильского ламантина в неволе Trichechus manatus manatus». Письма FEMS по микробиологии . 366 (11). дои : 10.1093/femsle/fnz134 . ПМИД 31210263 .
- ^ Сенал Л., Браммер-Роббинс Э., Уормингтон А.М., Блаха Л., Бисези Дж., Ларкин И. и др. (2021). «Состав и функции микробиома водных позвоночных: мелкие организмы, оказывающие большое влияние на здоровье водных животных» . Границы микробиологии . 12 : 567408. doi : 10.3389/fmicb.2021.567408 . ПМЦ 7995652 . ПМИД 33776947 .
- ^ Бик Э.М., Костелло Э.К., Свитцер А.Д., Каллахан Б.Дж., Холмс С.П., Уэллс Р.С. и др. (февраль 2016 г.). «Морские млекопитающие обладают уникальной микробиотой, сформированной морем и в то же время отличной от него» . Природные коммуникации . 7 : 10516. Бибкод : 2016NatCo...710516B . дои : 10.1038/ncomms10516 . ПМЦ 4742810 . ПМИД 26839246 .
- ^ Бай С., Чжан П., Линь М., Линь В., Ян З., Ли С. (май 2021 г.). «Микробное разнообразие и структура желудочно-кишечного тракта двух мельных короткоплавниковых гринд (Globicephala macrorhynchus) и карликового кашалота (Kogia breviceps)» . Интегративная зоология . 16 (3): 324–335. дои : 10.1111/1749-4877.12502 . ПМЦ 9292824 . ПМИД 33174288 . S2CID 226302293 .
- ^ Апприл А., Муни Т.А., Лайман Э., Симперерт А.К., Раппе М.С. (апрель 2011 г.). «Горбатые киты содержат комбинацию специфических и разнообразных кожных бактерий». Отчеты по экологической микробиологии . 3 (2): 223–232. Бибкод : 2011EnvMR...3..223A . дои : 10.1111/j.1758-2229.2010.00213.x . ПМИД 23761254 .
- ^ Апприлл А., Миллер К.А., Мур М.Дж., Дурбан Дж.В., Фернбах Х., Барретт-Леннард Л.Г. (2017). «Обширный основной микробиом китов, пойманных дронами, поддерживает основу для мониторинга здоровья» . mSystems . 2 (5). дои : 10.1128/mSystems.00119-17 . ПМЦ 5634792 . ПМИД 29034331 .
- ^ Вендл С., Феррари Б.С., Томас Т., Славич Э., Чжан Э., Нельсон Т. и др. (август 2019 г.). «Межгодовое сравнение основных таксонов и состава сообщества ударной микробиоты горбатых китов Восточной Австралии» . ФЭМС Микробиология Экология . 95 (8). дои : 10.1093/femsec/fiz102 . ПМИД 31260051 .
- ^ Джонсон В.Р., Торралба М., Фэйр П.А., Боссарт Г.Д., Нельсон К.Е., Моррис П.Дж. (декабрь 2009 г.). «Новое разнообразие бактериальных сообществ, связанных с верхними дыхательными путями афалин». Отчеты по экологической микробиологии . 1 (6): 555–62. Бибкод : 2009EnvMR...1..555J . дои : 10.1111/j.1758-2229.2009.00080.x . ПМИД 23765934 .
- ^ Сентеллеге С., Карраро Л., Гонсалво Дж., Россо М., Эспости Е., Гили С. и др. (2020). «Использование беспилотных летательных аппаратов (БПЛА) для отбора проб микробиома мелких китообразных» . ПЛОС ОДИН . 15 (7): e0235537. Бибкод : 2020PLoSO..1535537C . дои : 10.1371/journal.pone.0235537 . ПМЦ 7332044 . ПМИД 32614926 .
- ^ Раверти С.А., Роудс Л.Д., Забек Э., Эшги А., Кэмерон К.Э., Хансон М.Б. и др. (март 2017 г.). «Респираторный микробиом находящихся под угрозой исчезновения южных косаток и микробиота окружающего микрослоя морской поверхности в восточной части северной части Тихого океана» . Научные отчеты . 7 (1): 394. Бибкод : 2017НатСР...7..394Р . дои : 10.1038/s41598-017-00457-5 . ПМЦ 5428453 . ПМИД 28341851 .
- ^ Лима Н., Роджерс Т., Асеведо-Уайтхаус К., Браун М.В. (февраль 2012 г.). «Временная стабильность и видовая специфичность бактерий, связанных с дыхательной системой афалин». Отчеты по экологической микробиологии . 4 (1): 89–96. Бибкод : 2012EnvMR...4...89L . дои : 10.1111/j.1758-2229.2011.00306.x . ПМИД 23757234 .
- ^ Геохеган Дж.Л., Пиротта В., Харви Э., Смит А., Бухманн Дж.П., Островски М. и др. (июнь 2018 г.). «Вирусологический отбор проб недоступных диких животных с помощью дронов» . Вирусы . 10 (6): 300. дои : 10.3390/v10060300 . ПМК 6024715 . ПМИД 29865228 .
- ^ Ноултон Н., Ровер Ф. (октябрь 2003 г.). «Многовидовые микробные мутуализмы на коралловых рифах: хозяин как среда обитания». Американский натуралист . 162 (4 Приложения): S51–62. дои : 10.1086/378684 . ПМИД 14583857 . S2CID 24127308 .
- ^ Поллок Ф.Дж., МакМиндс Р., Смит С., Борн Д.Г., Уиллис Б.Л., Медина М. и др. (ноябрь 2018 г.). «Бактерии, ассоциированные с кораллами, демонстрируют филосимбиоз и кофилогению» . Природные коммуникации . 9 (1): 4921. Бибкод : 2018NatCo...9.4921P . дои : 10.1038/s41467-018-07275-x . ПМК 6250698 . ПМИД 30467310 .
- ^ Томпсон-младший, Ривера Х.Э., Клосек С.Дж., Медина М. (2014). «Микробы в коралловом холобионте: партнеры в эволюции, развитии и экологических взаимодействиях» . Границы клеточной и инфекционной микробиологии . 4 : 176. дои : 10.3389/fcimb.2014.00176 . ПМК 4286716 . ПМИД 25621279 .
- ^ Угарелли К., Чакрабарти С., Лаас П., Стингл У. (декабрь 2017 г.). «Голобионт морской травы и его микробиом» . Микроорганизмы . 5 (4): 81. doi : 10.3390/microorganisms5040081 . ПМЦ 5748590 . ПМИД 29244764 . Измененный текст был скопирован из этого источника, который доступен по [ https://creativecommons.org/licenses/by/4.0/ Международной лицензии Creative Commons Attribution 4.0.
- ^ Кавальканти Г.С., Шукла П., Моррис М., Рибейро Б., Фоли М., Доан М.П. и др. (сентябрь 2018 г.). «Голобионты родолитов в меняющемся океане: взаимодействие хозяина и микробов обеспечивает устойчивость коралловых водорослей в условиях закисления океана» . БМК Геномика . 19 (1): 701. doi : 10.1186/s12864-018-5064-4 . ПМК 6154897 . ПМИД 30249182 . Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
Дальнейшие ссылки
[ редактировать ]- Сталь, Л.Дж. и Кретойу, М.С. (ред.) (2016) Морской микробиом: неиспользованный источник биоразнообразия и биотехнологического потенциала Springer. ISBN 9783319330006 .
- Морской микробиом и биогеохимические циклы в морских продуктивных районах . Frontiers Media SA 2020. ISBN 978-2-88963-276-3 . OCLC 1291256407 .