Антозоа
| Антозоа Временной диапазон: Поздний эдиакарский период до недавнего времени | |
|---|---|
 | |
| Обнажение кораллов на Большой Барьерный риф | |
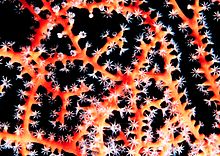 | |
| Горгонария с расширенными полипами | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Книдарийцы |
| Подтип: | Антозоа Эренберг , 1834 г. |
| Подклассы | |
Anthozoa — класс морских беспозвоночных , в который входят морские анемоны , каменистые и мягкие кораллы . Взрослые антозойи почти все прикрепляются к морскому дну, а их личинки могут расселяться в составе планктона . Основной единицей взрослого человека является полип ; он состоит из цилиндрической колонны, увенчанной диском с центральным отверстием, окруженным щупальцами. Морские анемоны в основном одиночные, но большинство кораллов являются колониальными и образуются в результате отпочкования новых полипов от исходной особи-основателя. Колонии укрепляются карбонатом кальция и другими веществами и принимают различную массивную, пластинчатую, кустистую или листовую форму.
Anthozoa включен в тип Cnidaria , который также включает медуз , коробчатых медуз и паразитических Myxozoa и Polypodiozoa . Двумя основными подклассами Anthozoa являются Hexacorallia , представители которых имеют шестикратную симметрию и включают каменистые кораллы, морские анемоны, трубчатые анемоны и зоантиды ; и Octocorallia , которые имеют восьмикратную симметрию и включают мягкие кораллы и горгонарии ( морские загоны , морские веера и морские кнуты ), а также морские анютины глазки . Меньший подкласс Ceriantharia состоит из трубчатых анемонов. Некоторые дополнительные виды также включены как incertae sedis до тех пор, пока не будет установлено их точное таксономическое положение.
Антозои — хищники , ловящие добычу щупальцами. Многие виды восполняют свои энергетические потребности, используя фотосинтезирующие одноклеточные водоросли , живущие в их тканях. Эти виды обитают на мелководье и многие из них строят рифы. У других видов зооксантеллы отсутствуют , и они, не нуждаясь в хорошо освещенных местах, обычно обитают в глубоководных местах.
В отличие от других представителей этого типа, антозойи не имеют в своем развитии стадии медузы . Вместо этого они выпускают сперму и яйцеклетки в воду. После оплодотворения личинки планулы составляют часть планктона. Полностью развившись, личинки оседают на морском дне и прикрепляются к субстрату , претерпевая метаморфозу в полипы. Некоторые антозойные могут также размножаться бесполым путем путем почкования или разрыва на части.
Разнообразие
[ редактировать ]
Название «Антозоа» происходит от греческих слов άνθος ( антос ; « цветок ») и ζώα ( зоа ; «животные»), следовательно, ανθόζωα ( антозоа ) = «цветочные животные», ссылка на цветочный вид их многолетних полипов . . [1]
Антозои являются исключительно морскими и включают морские анемоны , каменистые кораллы , мягкие кораллы , морские загоны , морские вееры и морские анютины глазки . — крупнейший таксон книдарий Anthozoa ; более шести тысяч одиночных и колониальных видов описано . Размер их варьируется от мелких особей менее полсантиметра в поперечнике до крупных колоний диаметром метр и более. Они включают виды с широким спектром цветов и форм, которые создают и улучшают рифовые системы. [2] [3] Хотя рифы и мелководье демонстрируют большое разнообразие видов, на самом деле в глубокой воде обитает больше видов кораллов, чем на мелководье, и многие таксоны в ходе своей эволюционной истории сместились с мелководья на глубоководье и наоборот. [4]
Филогения
[ редактировать ]Anthozoa подразделяется на три подкласса: Octocorallia , Hexacorallia и Ceriantharia , которые образуют монофилетические группы и в целом демонстрируют дифференцирующиеся отражения симметрии структуры полипов для каждого подкласса. Отношения внутри подклассов неразрешены. [5]
| Антозоа | |


Исторически «Ceriantipatharia» считалась отдельным подклассом, но из двух отрядов входящих в него Antipatharia теперь считается частью Hexacorallia, а Ceriantharia теперь считается независимым подклассом. Существующие приказы показаны справа. [5] [6]
К Hexacorallia относятся строители коралловых рифов : каменные кораллы ( Scleractinia ), морские анемоны ( Actiniaria ) и зоантиды ( Zoantharia ). Генетические исследования рибосомальной ДНК показали, что Ceriantharia является монофилетической группой и самым старым, или базальным, отрядом среди них. [7]
Классификация согласно Всемирному регистру морских видов : [8]
- подкласс Hexacorallia
- порядок Актиниария — морские анемоны.
- заказать Antipatharia — черный коралл
- отряд Corallimorpharia — кораллиморфы.
- заказать Ругосу †
- отряд Scleractinia — каменистые кораллы.
- отряд Zoantharia — зоантиды
- подкласс Octocorallia
- отряд Alcyonacea — мягкие кораллы и горгонарии.
- отряд Helioporacea — голубые кораллы.
- отряд Pennatulacea — перья, морские перья, морские ручки, морские анютины глазки.
- подкласс Ceriantharia — цериантарии, трубчатые анемоны.
- заказать Penicillaria
- заказать Спирулярию
- Антозоа неопределенного происхождения
- род Аиптасиодес
- заказать Аулопорида †
- род Sarcinula †
Octocorallia включает морские загоны ( Pennatulacea ), мягкие кораллы ( Alcyonacea ) и синие кораллы ( Helioporacea ). Морские кнуты и морские веера, известные как горгонарии , входят в состав Альционацеи и исторически разделялись на отдельные отряды. [6]
Цериантария включает родственные трубчатые анемоны. Трубчатые анемоны или цериантиды очень похожи на морские анемоны, но принадлежат к совершенно другому подклассу антозоев. Они одиночные, живут погребённые в мягких отложениях. Трубчатые анемоны живут и могут проникать в трубки, состоящие из волокнистого материала, состоящего из выделяемой слизи и нитей нематоцистоподобных органелл, известных как птихоцисты. [2]
| Основные таксоны антозоев | |||||
|---|---|---|---|---|---|
| Подкласс | Заказ | Изображение | Пример | Характеристики | Распределение |
| Гексакораллия | Актиниария Морские анемоны |  | Актиностола сп. | Преимущественно крупные одиночные полипы, прикрепленные к твердому субстрату. Часто красочно. Зооксантеллат или азооксантеллат. [2] | По всему миру на мелководье и глубоководье, наибольшее разнообразие наблюдается в тропиках. [2] |
| Гексакораллия | Антипатария Черный коралл |  | Плюмапатес пеннацеа | Густые колонии с тонкими ветвями. Осевой скелет из темных колючих ветвей, усиленный уникальным неколлагеновым белком. Азооксантеллат. [2] | На вертикальных скалах рифов или в глубокой воде. [2] |
| Гексакораллия | Кораллиморфария Кораллиморфы или коралловые анемоны |  | Дискосома сп. | Крупные одиночные полипы, похожие на морские анемоны, но с приземистыми колоннами и большими ротовыми дисками со множеством коротких щупалец. Ловят крупную добычу и некоторые виды зооксантеллят. [2] | На коралловых рифах, преимущественно тропических. [2] |
| Гексакораллия | грубый Вымерший |  | Стереолазма прямой кишки | Вымерший отряд, распространенный в период от среднего ордовика до поздней перми . Одиночные или колониальные, со скелетом из кальцита. Септы развиваются кратно четырем. [9] | Распространён. |
| Гексакораллия | Склерактиния Каменистые кораллы или твердые кораллы |   | Грибы грибы Тубастрея кокцинеа | Одиночные или колониальные кораллы самых разных размеров и форм, каменный скелет которых состоит из арагонита. Септы развиваются в количестве, кратном шести. [9] Зооксантеллат или азооксантеллат. [2] | Мелководные и глубоководные среды обитания по всему миру, наибольшее разнообразие наблюдается в тропических морях. [2] |
| Гексакораллия | Зоантария Зоантиды |  | Коралл «Драконий глаз» Зоантус сп. | Мелкие, преимущественно колониальные виды, соединенные ценсарком или столонами. Твердого скелета нет, но у некоторых есть твердые вещества в мясистой перидерме. [2] | Преимущественно тропические виды, обитающие на рифах. [2] |
| Октокораллия | Альционацея Мягкие кораллы и горгонарии |   | Зимородок пальчатый Грибные кораллы | Колониальные и разнообразные, с полипами, почти полностью погруженными в толстый мясистый ценосарк. Горгонарии имеют роговой скелет. Зооксантеллат или азооксантеллат. [2] | По всему миру, в основном в тропических и субтропических водах, связанных с коралловыми рифами и в глубоком море. [2] |
| Октокораллия | Гелиопорацея Синие кораллы |  | Гелиопора голубая | Октокораллы с массивным скелетом, состоящим из арагонита, выделяемого нижней стороной ценосарка. Зооксантеллат. [10] | Heliopora coerulea — Индо-Тихоокеанский регион; другие виды происходят из Атлантики и Мадагаскара. [11] |
| Октокораллия | Пеннатулача Морские ручки, морские перья и морские анютины глазки |  | Птилосаркус гурнейи | Колониальные виды, принимающие перистую, радиальную или булавовидную форму. Главная ось представляет собой одиночный увеличенный и удлиненный полип. Имеет несколько типов специализированных полипов. Азооксантеллат. [10] | По всему миру, от нижнего прилива до 6000 м (20 000 футов) [12] |
| Цериантария | Пенициллярия Трубчатые анемоны |  | Арахнантус сарси | Одиночные особи с двумя кольцами щупалец, живущие в волокнистых трубках в мягких осадках. Отличается от Spirularia анатомией и книдомом. [13] | В мягком осадке по всему миру. [14] |
| Цериантария | Спирулярия Трубчатые анемоны |  | Цериантус нитевидный | Одиночные особи с двумя кольцами щупалец, живущие в фиброзных трубочках. Отличается от Penicillaria анатомией и книдомом. [13] | В мягком осадке по всему миру. [14] |
Анатомия
[ редактировать ]Основная форма тела антозойных животных — полип . Он состоит из трубчатой колонны, увенчанной уплощенной областью, ротовым диском, с центральным ртом; Рот окружает клубок щупалец. У одиночных особей основанием полипа является ножка или педальный диск , который прикрепляется к субстрату , тогда как у колониальных полипов основание соединяется с другими полипами в колонии. [2]

Рот ведет в трубчатую глотку , которая на некоторое расстояние опускается в тело, а затем открывается в кишечнополостную полость, также известную как гастроваскулярная полость , занимающую внутреннюю часть тела. Внутренние напряжения придают рту форму щели, а концы щели ведут в две бороздки на стенке глотки, называемые сифоноглифами . Целентерон разделен на ряд вертикальных перегородок, известных как брыжейки или перегородки. Некоторые из них простираются от стенки тела до глотки и известны как «полные перегородки», тогда как другие не простираются так далеко и являются «неполными». Перегородки также прикрепляются к оральному и педальному дискам. [2]
Стенка тела состоит из эпидермального слоя, студенистого слоя мезоглеи и внутреннего гастродермы ; перегородки представляют собой складки стенки тела и состоят из слоя мезоглеи, зажатого между двумя слоями гастродермы. У некоторых таксонов мышцы сфинктера мезоглеи закрываются над ротовым диском и удерживают полип полностью втянутым. Щупальца содержат отростки кишечнополостного кишечника и имеют на стенках листки продольных мышц. Оральный диск имеет радиальные мышцы в эпидермисе, но большинство мышц позвоночника являются гастродермальными и включают сильные втягивающие мышцы рядом с перегородками. Количество и расположение перегородок, а также расположение этих втягивающих мышц важны для классификации антозоев. [2]
Щупальца вооружены нематоцистами , клетками, содержащими яд, которыми можно стрелять, как гарпуном, чтобы поймать и покорить добычу. Их необходимо заменить после обжига, и этот процесс занимает около сорока восьми часов. У некоторых актиний за пределами щупалец есть круг акрорагов; эти длинные выступы вооружены нематоцистами и действуют как оружие. Другой формой оружия являются аконции с аналогичным вооружением (нитевидные защитные органы), которые можно выдавливать через отверстия в стене колонны. Некоторые каменистые кораллы используют нагруженные нематоцистами «щупальца-чистильщики» в качестве защиты от вторжения других особей. [2]
Многие антозойи являются колониальными и состоят из множества полипов общего происхождения, соединенных живым материалом. Самая простая схема - это когда столон проходит вдоль субстрата в виде двумерной решетки с полипами, отпочковывающимися через определенные промежутки времени. Альтернативно, полипы могут отпочковаться от листа живой ткани, ценосарка , который присоединяется к полипам и закрепляет колонию на субстрате. Ценосарк может состоять из тонкой мембраны, из которой выступают полипы, как у большинства каменистых кораллов, или из толстой мясистой массы, в которую полипы погружены отдельно от ротовых дисков, как у мягких кораллов. [2]
Скелет каменистого коралла отряда Scleractinia секретируется эпидермисом нижней части полипа; при этом образуется кораллит , чашеобразная полость из карбоната кальция , в которой находится полип. У колониальных кораллов в результате роста полипа путем почкования образуются новые кораллиты, поверхность скелета которых покрыта слоем ценосарка. Эти колонии принимают различные массивные, ветвящиеся, листовидные и покрытые коркой формы. [15] Мягкие кораллы подкласса Octocorallia также являются колониальными и имеют скелет, образованный из мезоглеальной ткани, часто укрепленный известковыми спикулами или роговым материалом, а некоторые имеют внутри стержнеобразные опоры. [16] Другие антозойные растения, например морские анемоны, голые; в качестве поддержки они опираются на гидростатический скелет. Некоторые из этих видов имеют липкий эпидермис, к которому прилипают песчинки и фрагменты раковин, и зоантиды включают эти вещества в свою мезоглею. [2]
Биология
[ редактировать ]
Большинство антозоев — оппортунистические хищники , ловящие добычу, которая дрейфует в пределах досягаемости их щупалец. Добыча закрепляется с помощью липкой слизи, спироцист (неядовитых клеток-гарпунов) и нематоцист (ядовитых клеток-гарпунов). Затем щупальца изгибаются, чтобы протолкнуть более крупную добычу в рот, в то время как более мелкая добыча размером с планктон перемещается с помощью ресничек к кончику щупалец, которые затем вставляются в рот. Рот может растягиваться, чтобы вместить крупные предметы, а у некоторых видов губы могут расширяться, чтобы помочь поймать добычу. Затем глотка захватывает добычу, которая смешивается со слизью и медленно заглатывается перистальтикой и действием ресничек. Когда пища достигает кишечнополостного, внеклеточное пищеварение начинается за счет выделения нематоцист на основе перегородок и высвобождения ферментов. Частично переваренные фрагменты пищи циркулируют в кишечнополостном ресничках и отсюда поглощаются путем фагоцитоза гастродермальными клетками, выстилающими полость. [2]
Большинство антозоев дополняют свое хищничество, включая в свои ткани определенные одноклеточные фотосинтезирующие организмы, известные как зооксантеллы (или зоохлореллы в некоторых случаях ); многие таким образом удовлетворяют большую часть своих потребностей в питании. В этих симбиотических отношениях зооксантеллы получают выгоду, используя азотистые отходы и углекислый газ, вырабатываемые хозяином, в то время как книдарий приобретает фотосинтетическую способность и увеличивает выработку карбоната кальция, вещества, имеющего большое значение для каменистых кораллов. [17] Наличие зооксантелл не является постоянной связью. При некоторых обстоятельствах симбионты могут быть изгнаны, а их место позже могут занять другие виды. Это также может повлиять на поведение антозоя: он предпочитает селиться в хорошо освещенном месте и конкурирует со своими соседями за свет, позволяющий осуществлять фотосинтез. Если антозой живет в пещере или другом темном месте, симбионт может отсутствовать у вида, который в освещенном солнцем месте обычно получает от него пользу. [18] Антозои, живущие на глубине более 50 м (200 футов), являются азооксантеллами, поскольку для фотосинтеза недостаточно света. [4]

Благодаря продольным, поперечным и радиальным мышцам полипы способны удлиняться и укорачиваться, сгибаться и перекручиваться, надуваться и сдуваться, а также растягивать и сжимать свои щупальца. Большинство полипов вытягиваются, чтобы питаться, и сжимаются, если их потревожить, часто впячивая ротовые диски и щупальца в колонку. Сокращение достигается за счет выкачивания жидкости из кишечнополостного, а рефляция - за счет ее втягивания - задача, выполняемая сифоноглифами в глотке, выстланными бьющимися ресничками. Большинство антозоев прикрепляются к субстрату своими педальными дисками, но некоторые способны отделяться и передвигаться, а другие зарываются в осадок. Движение может представлять собой пассивное дрейф по течению или, в случае морских анемонов, может включать ползание по поверхности на их основании. [2]
Газообмен и выделение осуществляются путем диффузии через щупальца, а также внутренние и внешние стенки тела, чему способствует движение жидкости, переносимой по этим поверхностям ресничками. Сенсорная система состоит из простых нервных сетей в гастродерме и эпидермисе, но специализированных органов чувств нет. [2]
Антозои обладают огромной способностью к регенерации; утраченные части быстро отрастают заново, и актинию Aiptasia pallida можно подвергнуть вивисекции в лаборатории, а затем вернуть в аквариум, где она заживет. Они способны к различным бесполым способам размножения, включая фрагментацию , продольное и поперечное деление и почкование . [2] Морские анемоны, например, могут ползать по поверхности, оставляя после себя отдельные части педального диска, из которых развиваются новые клональные особи. Виды Anthopleura делятся продольно, разделяясь, в результате чего образуются группы особей с идентичной окраской и рисунком. [19] Поперечное деление встречается реже, но встречается у Anthopleura stellula и Gonactinia prolifera , при этом рудиментарный пучок щупалец появляется на столбике до того, как актиния разрывается на части. [20] Зоантиды способны отпочковывать новых особей. [21]

Большинство кораллов однополые , но некоторые каменистые кораллы гермафродиты . Половые клетки зарождаются в энтодерме и перемещаются в гастродерму, где дифференцируются. По мере созревания они высвобождаются в кишечнополостную и оттуда в открытое море, при этом оплодотворение происходит наружно. [2] Чтобы сделать оплодотворение более вероятным, кораллы выделяют огромное количество гамет , и многие виды синхронизируют их высвобождение в зависимости от времени суток и фазы луны. [22]
Зигота прежде развивается в личинку планулы , которая плавает с помощью ресничек и является частью планктона, некоторое время чем оседать на морском дне и превращаться в ювенильного полипа. Некоторые планулы содержат желточный материал, а другие включают зооксантеллы, и эти адаптации позволяют этим личинкам поддерживать себя и распространяться более широко. [2] Например, планулы каменистого коралла Pocillopora damicornis имеют богатые липидами желтки и остаются жизнеспособными в течение 100 дней, прежде чем им потребуется оседание. [23]
Экология
[ редактировать ]
Коралловые рифы являются одними из самых биоразнообразных мест обитания на Земле, где обитает большое количество видов кораллов, рыб , моллюсков , червей , членистоногих , морских звезд , морских ежей , других беспозвоночных и водорослей . Из-за фотосинтетических потребностей кораллов они встречаются на мелководье и многих из этих окраин суши. [24] Коралловые рифы с трехмерной структурой представляют собой очень продуктивные экосистемы; они обеспечивают пищу своим обитателям, укрытия разных размеров, подходящие для многих организмов, места для насеста, препятствия для крупных хищников и прочные конструкции, на которых можно расти. Они используются в качестве нерестилищ и питомников для многих видов пелагических рыб и влияют на продуктивность океана на многие мили вокруг. [25] Антозои охотятся на животных, меньших их размеров, и сами поедаются такими животными, как рыбы, крабы, ракушки, улитки и морские звезды. Их среда обитания легко нарушается внешними факторами, нарушающими баланс экосистемы. В 1989 году агрессивная морская звезда с терновым венцом ( Acanthaster planci ) вызвала хаос в Американском Самоа , убив 90% кораллов на рифах. [26]
Кораллы, растущие на рифах, называются герматипическими , а кораллы, растущие в других местах, — агерматипическими. Большинство последних являются азооксантеллатами и обитают как на мелководье, так и на глубоководных участках моря. В глубоком море они делят экосистему с мягкими кораллами, многощетинковыми червями , другими червями, ракообразными, моллюсками и губками . В Атлантическом океане холодноводный коралл Lophelia pertusa образует обширные глубоководные рифы, на которых обитают многие другие виды. [27]
Другая фауна, например гидрозоа , мшанки и хрупкие звезды , часто обитает среди ветвей горгонарий и колоний кораллов. [28] Карликовый морской конек не только делает своим домом определенные виды горгонарий, но и очень похож на своего хозяина и поэтому хорошо замаскирован. [29] Некоторые организмы имеют облигатные отношения со своим видом-хозяином. Моллюск Simnialena marferula встречается только на морском кнуте Leptogorgia virgulata , имеет такой же цвет и изолировал свои защитные химические вещества, а голожаберный Tritonia wellsi — еще один облигатный симбионт , его перистые жабры напоминают щупальца полипов. [30]

Ряд видов актиний являются комменсалами с другими организмами. Некоторые крабы и раки-отшельники ищут морские анемоны и помещают их на свои панцири для защиты, а рыбы, креветки и крабы живут среди щупалец анемоны, получая защиту, находясь в непосредственной близости от стрекательных клеток. Некоторые амфиподы живут внутри кишечнополостного актиния. [31] Несмотря на ядовитые клетки, морские анемоны поедаются рыбами, морскими звездами, червями, морскими пауками и моллюсками. Морской слизень Aeolidia papillosa питается агрегирующей актинией ( Anthopleura Elegantissima ), накапливая нематоцисты для собственной защиты. [31]
Палеонтология
[ редактировать ]Считается, что несколько вымерших отрядов кораллов палеозойской эры (~ 540–252 миллиона лет назад) близки к предкам современных Scleractinia: [32] [33]
- Нумидиафиллида †
- Килбухофиллида †
- Гетерокораллия †
- Ругоса †
- Гелиолитида †
- этажи †
- Котониида †
- Табулоконид †
Все это кораллы и соответствуют временной шкале окаменелостей . Обладая хорошо сохранившимися твердыми известковыми скелетами, они составляют большинство окаменелостей антозоя.
 | |
Хронология основных летописей окаменелостей кораллов и событий от 650 млн лет назад до настоящего времени. [34] [35] | |
Взаимодействие с людьми
[ редактировать ]Коралловым рифам и мелководной морской среде угрожают не только природные явления и повышение температуры моря, но и такие антропогенные проблемы, как загрязнение окружающей среды , отложение осадков и разрушительные методы рыболовства. Загрязнение может быть результатом стока с земель сточных вод, сельскохозяйственной продукции, топлива или химикатов. Они могут напрямую убить или нанести вред морской жизни, или могут способствовать росту водорослей, которые душат местные виды, или вызывать цветение водорослей с далеко идущими последствиями. Разливы нефти в море могут загрязнить рифы, а также повлиять на яйца и личинки морских обитателей, дрейфующие у поверхности. [36]
Кораллы собираются для продажи в аквариумах, и это можно делать, не заботясь о долгосрочном выживании рифа. Рыбалка среди рифов затруднена, а траление наносит большой механический ущерб. В некоторых частях мира взрывчатые вещества используются для вытеснения рыбы из рифов, и для той же цели может использоваться цианид; Обе практики не только убивают без разбора обитателей рифов, но также убивают или повреждают кораллы, иногда подвергая их такому стрессу, что они выбрасывают свои зооксантеллы и обесцвечиваются . [36]
Глубоководным местам обитания кораллов также угрожает деятельность человека, особенно неизбирательное траление. Эти экосистемы мало изучены, но в вечной темноте и низких температурах животные растут и взрослеют медленно, а рыбы, которую стоит поймать, относительно меньше, чем в освещенных солнцем водах наверху. В какой степени глубоководные коралловые рифы обеспечивают безопасную зону нагула молоди рыб, не установлено, но они могут иметь важное значение для многих холодноводных видов. [37]
Ссылки
[ редактировать ]- ^ «Антозоа: Этимология» . Прекрасный словарь . Проверено 25 июня 2017 г.
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С Руперт, Эдвард Э.; Фокс, Ричард, С.; Барнс, Роберт Д. (2004). Зоология беспозвоночных (7-е изд.). Cengage Обучение. стр. 112–148. ISBN 978-81-315-0104-7 .
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Кроутер, Алабама (2011). «Класс Anthozoa Ehrenberg, 1834» (PDF) . В З.-К. Чжан (ред.). Биоразнообразие животных: обзор классификации более высокого уровня и обзор таксономического богатства . Том. 3148. стр. 19–23. дои : 10.11646/zootaxa.3148.1.5 .
{{cite book}}:|journal=игнорируется ( помогите ) - ^ Перейти обратно: а б Вудли, Шерил М.; Даунс, Крейг А.; Брукнер, Эндрю В.; Портер, Джеймс В .; Галлоуэй, Сильвия Б. (2016). Болезни кораллов . Джон Уайли и сыновья. п. 416. ИСБН 978-0-8138-2411-6 .
- ^ Перейти обратно: а б Стампар, С.Н.; Маронна, ММ; Китахара, М.В.; Реймер, доктор медицинских наук; Морандини, AC (март 2014 г.). «Быстро развивающаяся митохондриальная ДНК у цериантариев: отражение парафилии гексакораллии?» . ПЛОС ОДИН . 9 (1): е86612. Бибкод : 2014PLoSO...986612S . дои : 10.1371/journal.pone.0086612 . ПМЦ 3903554 . ПМИД 24475157 .
- ^ Перейти обратно: а б Дейли, М.; Брюглер, член парламента; Картрайт, П.; Коллинз, АГ; Доусон, Миннесота; Фаутин, Д.Г.; Франция, Южная Каролина; Макфадден, CS; Опреско, Д.М.; Родригес, Э.; Романо, СЛ; Стейк, JL (2007). «Тип Cnidaria: обзор филогенетических моделей и разнообразия через 300 лет после Линнея» (PDF) . Зоотакса . 1668 : 1–766. дои : 10.5281/zenodo.180149 .
- ^ Чен, Калифорния; ДМ Одорико; М. тен Лохуис; ДЖЕН Верон; DJ Миллер (июнь 1995 г.). «Систематические взаимоотношения внутри Anthozoa (Cnidaria: Anthozoa) с использованием 5'-конца 28S рДНК». Молекулярная филогенетика и эволюция . 4 (2): 175–183. дои : 10.1006/mpev.1995.1017 . ПМИД 7663762 .
- ^ Хоксема, Берт (2013). «Антозоа» . ВОРМС . Всемирный реестр морских видов . Проверено 24 апреля 2015 г.
- ^ Перейти обратно: а б Тейлор, Пол Д.; Льюис, Дэвид Н. (2007). Ископаемые беспозвоночные . Издательство Гарвардского университета. п. 25. ISBN 978-0-674-02574-5 .
- ^ Перейти обратно: а б Барнс, Роберт Д. (1982). Зоология беспозвоночных . Холт-Сондерс Интернэшнл. стр. 168–169. ISBN 978-0-03-056747-6 .
- ^ Фабрициус, Катарина; Олдерслейд, Филип (2001). Мягкие кораллы и морские вееры: всеобъемлющий путеводитель по тропическим мелководным родам центрально-западной части Тихого океана, Индийского океана и Красного моря . Австралийский институт морских наук. п. 6. ISBN 978-0-642-32210-4 .
- ^ Уильямс, GC (2011). «Глобальное разнообразие морских загонов (Cnidaria: Octocorallia: Pennatulacea)» . ПЛОС ОДИН . 6 (7): e22747. Бибкод : 2011PLoSO...622747W . дои : 10.1371/journal.pone.0022747 . ПМК 3146507 . ПМИД 21829500 .
- ^ Перейти обратно: а б Гоффредо, Стефано; Дубинский, Звы (2016). Книдария, прошлое, настоящее и будущее: мир Медузы и ее сестер . Международное издательство Спрингер. п. 66. ИСБН 978-3-319-31305-4 .
- ^ Перейти обратно: а б Фаутен, Дафна Г.; Вестфол, Джейн А.; Картрайт, Полин; Дейли, Мэримеган; Виттенбах, Чарльз Р. (2007). Биология кишечнополостных 2003: Тенденции в исследованиях Cnidaria и Ctenophora . Springer Science & Business Media. п. 261. ИСБН 978-1-4020-2762-8 .
- ^ «Кораллы: как растут каменные кораллы? Какие формы они принимают?» . Национальное управление океанических и атмосферных исследований. 25 марта 2008 года . Проверено 13 июня 2017 г.
- ^ «Знакомство с Октокораллиями» . Музей палеонтологии Калифорнийского университета . Проверено 13 июня 2017 г.
- ^ «Вклад в информационный бюллетень BUFUS, Экскурсия в провинцию Милн-Бэй - Папуа-Новая Гвинея, Мадл и Йип, 2000 г.» . Архивировано из оригинала 11 мая 2020 г. Проверено 23 мая 2011 г.
- ^ Свет, Сол Фелти (2007). Руководство Лайт и Смита: Беспозвоночные приливной зоны от Центральной Калифорнии до Орегона . Издательство Калифорнийского университета. п. 176. ИСБН 978-0-520-23939-5 .
- ^ Геллер, Джонатан Б.; Фицджеральд, Лори Дж.; Кинг, Чад Э. (2005). «Деление морских анемонов: интегративные исследования эволюции жизненного цикла1» . Интегративная и сравнительная биология . 45 (4): 615–622. CiteSeerX 10.1.1.315.4480 . дои : 10.1093/icb/45.4.615 . ПМИД 21676808 .
- ^ Гоффредо, Стефано; Дубинский, Звы, ред. (2016). Книдария, прошлое, настоящее и будущее: мир Медузы и ее сестер . Международное издательство Спрингер. п. 240. ИСБН 978-3-319-31305-4 .
- ^ Кэрфут, Том. «Узнайте о морских анемонах и их родственниках: размножение: бесполое» . Одиссея улитки . Архивировано из оригинала 6 июня 2017 года . Проверено 15 июня 2017 г.
- ^ «Кораллы: как размножаются кораллы?» . Национальное управление океанических и атмосферных исследований. 25 марта 2008 года . Проверено 17 июня 2017 г.
- ^ Ричмонд, Р.Х. (1987). «Энергетика, компетентность и распространение личинок планулы коралла Pocillopora damicornis на большие расстояния ». Морская биология . 93 (4): 527–533. дои : 10.1007/BF00392790 . S2CID 84571244 .
- ^ «Кораллы: значение коралловых рифов» . Национальное управление океанических и атмосферных исследований. 25 марта 2008 года . Проверено 17 июня 2017 г.
- ^ Сорокин, Юрий Иванович (2013). Экология коралловых рифов . Springer Science & Business Media. стр. 1–3. ISBN 978-3-642-80046-7 .
- ^ «Кораллы: естественная угроза коралловым рифам» . Национальное управление океанических и атмосферных исследований. 25 марта 2008 года . Проверено 17 июня 2017 г.
- ^ Робертс, Дж. Мюррей (2009). Холодноводные кораллы: биология и геология глубоководных мест обитания кораллов . Издательство Кембриджского университета. п. 152. ИСБН 978-0-521-88485-3 .
- ^ Хейвуд, Мартин; Сью Уэллс (1989). Справочник морских беспозвоночных . Tetra Press:Salamander Books Ltd. с. 208 . ISBN 978-3-89356-033-2 .
- ^ Агбаяни, Эли. «Hippocampus bargibanti, Карликовый морской конёк» . ФишБаза . Проверено 18 июня 2017 г.
- ^ Руперт, Эдвард Э.; Ричард С. Фокс (1988). Приморские животные юго-востока: справочник по обыкновенным мелководным беспозвоночным юго-восточного побережья Атлантического океана . Издательство Университета Южной Каролины. п. 124 . ISBN 9780872495357 .
- ^ Перейти обратно: а б Ван-Прат, М. (1985). Достижения морской биологии . Академическая пресса. стр. 92–93. ISBN 978-0-08-057945-0 .
- ^ Оливер В.А. младший (1996). «Происхождение и взаимоотношения палеозойских коралловых групп и происхождение склерактиний». В Стэнли, GDJ (ред.). Палеобиология и биология кораллов . Том. 1. Колумбус, Огайо: Палеонтологическое общество. стр. 107–134. дои : 10.1017/S1089332600000073 .
{{cite book}}:|journal=игнорируется ( помогите ) - ^ Котрч, Бен (2005). «Антозоа: Подгруппы» . Ископаемые группы . Бристольский университет. Архивировано из оригинала 27 сентября 2009 г. Проверено 10 апреля 2007 г.
- ^ Ваггонер, Бен М. (2000). Смит, Дэвид; Коллинз, Аллен (ред.). «Антозоа: Летопись окаменелостей» . Антозоа . УКМП . Проверено 9 марта 2020 г.
- ^ Оливер, Уильям А. младший (2003). «Кораллы: Таблица 1» . Ископаемые группы . Геологическая служба США . Архивировано из оригинала 9 января 2009 года . Проверено 9 марта 2020 г.
- ^ Перейти обратно: а б «Кораллы: антропогенные угрозы кораллам» . Национальное управление океанических и атмосферных исследований. 25 марта 2008 года . Проверено 13 июня 2017 г.
- ^ Робертс, Дж. Мюррей (2009). Холодноводные кораллы: биология и геология глубоководных мест обитания кораллов . Издательство Кембриджского университета. п. 163. ИСБН 978-0-521-88485-3 .
