Зооксантеллы

Зооксантеллы ( / ˌ z oʊ ə z æ n ˈ θ ɛ l iː / ; sg.zooxanthella . ) — разговорный термин для одноклеточных динофлагеллят , которые способны жить в симбиозе с разнообразными морскими беспозвоночными, включая демогубок , кораллов , медуз и голожаберников . Наиболее известные зооксантеллы относятся к роду Symbiodinium . [1] но некоторые из них известны из рода Amphidinium , а другие таксоны , пока еще не идентифицированные, могут иметь схожее сходство с эндосимбионтами . [2] Настоящая Zooxanthella K.brandt является мутуалистом радиолярии Collozoum inerme (Joh.Müll., 1856). [3] и систематически помещается в Peridiniales. [4] [а] Другая группа одноклеточных эукариот, которые участвуют в сходных эндосимбиотических отношениях как в морской, так и в пресноводной среде обитания, — это зеленые водоросли зоохлореллы . [6]
Зооксантеллы — фотосинтезирующие организмы, содержащие хлорофилл а и хлорофилл с , а также пигменты динофлагеллят перидинин и диадиноксантин . Они обеспечивают желтоватый и коричневатый цвет, типичный для многих видов-хозяев. [2] В течение дня они снабжают своего хозяина органическими углеродными продуктами фотосинтеза, иногда обеспечивая до 90% энергетических потребностей хозяина для метаболизма, роста и размножения. Взамен они получают питательные вещества, углекислый газ и возвышенное положение с доступом к солнечному свету. [7] [8]
Морфология и классификация
[ редактировать ]Зооксантеллы можно сгруппировать в классы Bacillariophyceae , Cryptophyceae , Dinophyceae и Rhodophycaeae , а также к родам Amphidinium , Gymnodinium , Aureodinium , Gyrodinium , Prorocentrum , Scrippsiella , Gloeodinium и, чаще всего, Symbiodinium . [9] Зооксантеллы рода Symbiodinium принадлежат в общей сложности к восьми филогенетическим кладам AH, дифференцированным по ядерной рибосомальной ДНК и ДНК хлоропластов. [10]
Зооксантеллы — автотрофы , содержащие хлоропласты, состоящие из тилакоидов, собранных в группы по три. [9] Пиреноид выступает из каждого хлоропласта и покрыт вместе с хлоропластом толстым крахмалистым покровом. В цитоплазме клетки также существуют липидные вакуоли, кристаллы оксалата кальция , диктиосомы и митохондрии . [9] Клеточная стенка зооксантелл различается по строению у разных видов. Одна структура состоит из внешней мембраны, среднего слоя, компактного с электронами, и тонкого внутреннего слоя. У других видов характеристики этого внутреннего слоя низкой плотности составляют всю структуру клеточной стенки. [9] Под клеточной стенкой находится клеточная мембрана, а под клеточной мембраной находятся текальные пузырьки. [9]
ДНК в клетке существует в виде плотно спрессованных между собой клубков хроматина. [9] Он конденсируется в ядре вместе с атипичным набором гистонов . [11] [12] [13] ДНК содержит рибосомальную РНК (рРНК), которая свернута и имеет морфологию, аналогичную рРНК у археобактерий. Это указывает на то, что РНК важна для упаковки ДНК зооксантелл. [11] Зооксантеллы, помимо всех других динофлагеллят, содержат в своих геномах 5-гидроксиметилурацил и тимидин , в отличие от любого другого эукариотического генома. [11]
История жизни
[ редактировать ]Зооксантеллы чередуют фазы жизни, выраженные в виде цист и подвижных организмов в толще воды. [14] [15] У зооксантелл рода Gymnodinium один из возможных жизненных циклов клетки начинается с незрелой кисты, которая достигает зрелости, а затем снова делится, образуя незрелую кисту. Вырастая в более старую клетку, она становится бесполезной. В жизненном цикле подвижной клетки зооксантелл ее самая молодая стадия известна как зооспорангий, который созревает в зооспору, способную к подвижности. Эта подвижная клетка производит и выпускает гаметы для размножения. [15]
Вегетативная фаза
[ редактировать ]Вегетативная фаза жизненного цикла зооксантелл является преобладающей формой существования организма. [14] В этой форме одноклеточный организм имеет тонкую клеточную стенку. В отличие от зооспоры зооксантелла содержит многочисленные хлоропласты. Однако когда клетка продолжает расти, количество хлоропластов уменьшается. [14] Вегетативная клетка либо разделится на две отдельные дочерние клетки, либо перейдет в стадию кисты. [14]
Стадии кисты
[ редактировать ]Наиболее распространенными фазами в истории жизни зооксантелл после вегетативной фазы являются цисты, делящиеся кисты и дегенеративные кисты. [15] Цисты обладают толстой клеточной стенкой, но сохраняют состав цитоплазмы и составляют большинство сгруппированных зооксантелл в тканях хозяина. Эта стадия развития клетки придает хозяину красновато-коричневый оттенок. [15] Делящиеся кисты составляют четверть состава кластеров зооксантелл в тканях хозяина и выражаются в виде клеточных стадий, где две дочерние клетки остаются соединенными, но обладают отдельными клеточными стенками. Дегенеративные цисты присутствуют в кластерах, хотя и редко, и теряют большую часть своей взаимной пользы для хозяина, в котором они проживают, из-за снижения эффективности фотосинтеза. [15] Молодые стадии зооспорангия и подвижные зооспоры, хотя и наблюдаются в жизненных циклах зооксантелл, среди клад встречаются гораздо реже. Зооспора находится в зооспорангии до тех пор, пока клеточная стенка цисты не лопнет. Зооксантеллы подвижны только в том случае, если они возникли из зооспор. [15]
Подвижность
[ редактировать ]Зооксантеллы на стадии зооспор проявляют подвижность в виде движения вперед или вращательного движения. [15] Двигаясь вперед, организм вращается вокруг оси заднего жгутика, одновременно продвигаясь сквозь толщу воды. Зооспора движется по кругу в толще воды, прикрепляясь задним жгутиком к субстрату. [15]
Экология
[ редактировать ]Приобретение эндосимбионтов
[ редактировать ]Зооксантеллы особенно связаны с кораллами , образующими рифы , но они также населяют других беспозвоночных и протистов ; их хозяевами являются многие морские анемоны , медузы , голожаберные моллюски , некоторые двустворчатые моллюски , такие как гигантский моллюск Тридакна , губки и плоские черви , а также некоторые виды радиолярий и фораминифер . [16] В организмах-хозяевах присутствует множество различных видов зооксантелл, каждый из которых обладает собственными адаптивными возможностями и степенью толерантности к различным факторам окружающей среды. [2]
Молодой организм или вновь созданная колония могут приобрести зооксантеллы путем полового размножения или непосредственно из окружающей среды. Яйцо, из которого развилась особь, могло быть уже заражено зооксантеллами во время оплодотворения, либо клетки симбионта могли быть перенесены от матери в период, когда личинка высиживала своего родителя. Альтернативно, новая особь может приобрести зооксантеллы непосредственно из морской воды, в которой динофлагелляты свободно живут на некоторых стадиях своего жизненного цикла. Некоторые каменистые кораллы используют хемотаксис , при этом заражение происходит в результате выделения кораллом химического аттрактанта. Заражение может также произойти после проглатывания инфицированных фекалий хозяина или добычи, в которой уже обитают симбионты. Такое непрямое заражение может привести к заражению нового хозяина видом зооксантеллы, отличным от того, который присутствует у его родителя. [2]
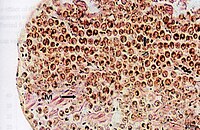 Поперечный разрез мантийной ткани гигантского моллюска, показывающий симбиотических простейших.
Поперечный разрез мантийной ткани гигантского моллюска, показывающий симбиотических простейших.
 Схема радиолярий, содержащих зооксантеллы (з)
Схема радиолярий, содержащих зооксантеллы (з)



Симбиоз с кораллом
[ редактировать ]Зооксантелла в симбиозе с кораллом содержится в вакуолях гастродермальных клеток хозяина и относится к роду Symbiodinium . [17] Зооксантеллы обеспечивают книдарий-хозяев питательными веществами в виде сахаров, глицерина и аминокислот, а взамен получают углекислый газ, фосфаты и соединения азота. [18] [17] Коралл, подвергшийся воздействию стрессоров окружающей среды, может привести к изгнанию зооксантелл из тканей хозяина. Это, в свою очередь, лишает коралл его цвета, в этом явлении, известном как обесцвечивание коралла , когда теперь прозрачные ткани коралла обнажают его белую внутреннюю скелетную структуру. [17] Изменения солености, интенсивности света, температуры, загрязнения, седиментации и болезней могут повлиять на эффективность фотосинтеза зооксантелл или привести к исключению из их мутуалистических отношений. [17]
Физиологические механизмы изгнания эндосимбионтов все еще изучаются, но предполагается, что они включают различные способы отделения зооксантелл или гастродермальных клеток от кораллов-хозяев. [17] Во время обесцвечивания целые гастродермальные клетки, содержащие зооксантеллы, могут покинуть хозяина. В других случаях гастродермальные клетки остаются в тканях хозяина, но зооксантеллы, содержащиеся в вакуолях, могут повреждаться отдельно или физически покидать клетки и проникать в окружающую среду. [17]
Моллюски и зооксантеллы
[ редактировать ]Коралл — не единственный водный организм, на которого влияет обесцвечивание и изгнание зооксантелл; Также было обнаружено, что моллюски подвергаются аналогичному процессу, когда температура становится слишком высокой. [19] Однако моллюски выбрасывают еще живые зооксантеллы и, как было замечено, способны их восстановить. [19] Это имеет положительное значение не только для самих моллюсков, но и для окружающей их экосистемы. [19] Для многих организмов моллюски являются жизненно важной частью пищевой цепи. Мало того, что их самих можно есть, экскременты гигантских моллюсков содержат живые зооксантеллы. [19] Приспособленные кормушки и моллюски используют выделенные зооксантеллы в качестве источника питательных веществ. [19] Потребление зооксантелл особенно важно для моллюсков на стадии велигера, поскольку оно стимулирует рост. [19] Зооксантеллы обнаруживаются не только в экскрементах моллюсков, но и в тканях мантии, где они поглощают аммиак и нитраты. [20] Их также можно найти в глазах моллюсков, таких как Тридакна , где они действуют как линзы. [21] Различные клады зооксантелл оказывают влияние на морфологию моллюсков. [22] Клада E1 зооксантелл, по-видимому, оказывает влияние на более мелкое потомство моллюсков или отдает ему предпочтение по сравнению с моллюсками, несущими другие клады, и все пять клад, по-видимому, необходимы для расселения личинок. [22]
Медузы и зооксантеллы
[ редактировать ]Медузы и зооксантеллы имеют общую историю в научном мире, поскольку Symbiodinium был впервые выращен из медузы Cassiopea, модельного вида медуз. [23] Было замечено, что множество различных типов зооксантелл образуют отношения с медузами во многих различных филогенетических ветвях, и роли, которые они играют, будут меняться на протяжении жизненного цикла медуз. [23] Однако по мере старения медуз разнообразие прикрепляющихся к ним зооксантелл уменьшается, что позволяет предположить, что зооксантеллы конкурируют друг с другом за установление отношений с медузой. [23] Не все медузы вступают в отношения с этими микробами, а те, что по большей части, обитают в тропических и субтропических водах. [23] Изменение климата влияет на отношения между медузами и зооксантеллами немного иначе, чем на кораллы, несмотря на то, что оба они являются частью семейства книдарий. [23] Одно исследование показало, что некоторые виды медуз и их симбиотические зооксантеллы могут до определенного момента обладать некоторой устойчивостью к снижению pH, вызванному изменением климата. [24] Хотя случаи обесцвечивания медуз были зарегистрированы во время экстремальной жары. [23] Хотя причинные факторы, которые обычно влияют на отношения между зооксантеллами и их хозяином, могут не относиться к медузам, интенсивность света применима. [25] Доступность света может повлиять на выработку липидов зооксантелл, которые затем используют медузы. [25] Чтобы максимизировать поглощение света, медузы будут плавать у поверхности и совершать очень специфические миграции. [23] Модели миграции также помогают зооксантеллам получить доступ к определенным питательным веществам. [23] Многие из этих медуз, по-видимому, являются миксотрофными, питаясь как живой добычей, так и используя фототрофию. [23] Возможно, именно это помогает медузам пережить изменение климата и обесцвечивание, поскольку они могут менять методы питания, а не пытаться быстро восстановить утраченные зооксантеллы. [23] Когда дело доходит до взаимоотношений между зооксантеллами и медузами, остается много неизвестных, на которые ученые пытаются ответить. [23]
Ссылки
[ редактировать ]- ^ Брандт предложил Zooxanthella включить один вид — Zooxanthella nutricula . Название рода долгое время считалось проблематичным из-за путаницы с термином «зооксантелла» (единственное число «зооксантелла»), но предложения отказаться от этого названия не увенчались успехом. Согласно малоиспользуемому аргументу Guiry & Anderson 2018, Zooxanthella следует рассматривать как самое раннее родовое название Symbiodinium sensu lato. [5] Гири не цитировал работу Готтшлинга о Peridiniales.
- ^ ЛаЖенесс, Тодд К.; Паркинсон, Джон Эверетт; Габриэльсон, Пол В.; Чон, Хэ Джин; Реймер, Джеймс Дэвис; Вулстра, Кристиан Р.; Сантос, Скотт Р. (20 августа 2018 г.). «Систематический пересмотр Symbiodiniaceae подчеркивает древность и разнообразие коралловых эндосимбионтов» . Современная биология . 28 (16): 2570–2580.e6. дои : 10.1016/j.cub.2018.07.008 . hdl : 10754/630499 . ПМИД 30100341 .
- ^ Перейти обратно: а б с д Биркеланд, Чарльз (1997). Жизнь и смерть коралловых рифов . Springer Science & Business Media. стр. 98–99. ISBN 978-0-412-03541-8 .
- ^ Брандт, К. (1881). «О сосуществовании животных и водорослей». Архив анатомии и физиологии / Физиологический факультет (на немецком языке). 1881 : 570–574.
- ^ Готтшлинг, М.; Маклин, ТИ (2013). «Новый дом для крошечных симбионтов: динофиты, определенные как Zooxanthella, являются Peridiniales и отдаленно связаны с Symbiodinium ». Молекулярная филогенетика и эволюция . 67 (1): 217–222. дои : 10.1016/j.ympev.2013.01.003 . ПМИД 23333735 .
- ^ Майкл Д. Гири; Роберт А. Андерсен (апрель 2018 г.). «Возобновленное подтверждение родового названия Symbiodinium (Dinophyceae, Suessiaceae) и восстановление Zooxanthella K.Brandt» (PDF) . Нотула Альгарум . 58 : 1–5.
- ^ Хук, Кристиан; Манн, Дэвид; Янс, Х.М. (1995). Водоросли: введение в психологию . Издательство Кембриджского университета. п. 278. ИСБН 978-0-521-31687-3 .
- ^ Руперт, Эдвард Э.; Фокс, Ричард С.; Барнс, Роберт Д. (2004). Зоология беспозвоночных, 7-е издание . Cengage Обучение. п. 122. ИСБН 978-81-315-0104-7 .
- ^ Лор, Джейме; Манн, Колин Б.; Уилсон, Уильям Х. (2007). «Характеристика латентной вирусоподобной инфекции симбиотических зооксантелл» . Прикладная и экологическая микробиология . 73 (9): 2976–2981. Бибкод : 2007ApEnM..73.2976L . дои : 10.1128/АЕМ.02449-06 . ЧВК 1892877 . ПМИД 17351090 .
- ^ Перейти обратно: а б с д и ж Уэйкфилд, Тимоти; Фермер, Марк; Кемпф, Стивен (2000). «Пересмотренное описание тонкой структуры in situ рода Symbiodinium Zooxanthellae» . Биологический вестник . 199 (1): 76–84. дои : 10.2307/1542709 . JSTOR 1542709 . ПМИД 10975645 .
- ^ Беркельманс, Рэй; ван Оппен, Мадлен Дж. Х. (2006). «Роль зооксантелл в термоустойчивости кораллов: «самородок надежды» для коралловых рифов в эпоху изменения климата» . Труды Королевского общества B: Биологические науки . 273 (1599): 2305–2312. дои : 10.1098/rspb.2006.3567 . ПМК 1636081 . ПМИД 16928632 .
- ^ Перейти обратно: а б с Стат, Майкл; Картер, Ди; Хоэ-Гульдберг, Уве (2006). «Эволюционная история Symbiodinium и склерактиновых хозяев - симбиоз, разнообразие и влияние изменения климата». Перспективы экологии, эволюции и систематики растений . 8 : 23–43. дои : 10.1016/j.ppees.2006.04.001 .
- ^ Херцог, М.; Сойер-Гобийяр, М. (1981). «Отличительные особенности хроматина динофлагеллят. Отсутствие нуклеосом у примитивного вида Prorocentrum micans ». Европейский журнал клеточной биологии . 23 (2): 295–302. ПМИД 6258920 .
- ^ Риццо, Пи Джей (1981). «Сравнительные аспекты основных белков хроматина динофлагеллят». БиоСистемы . 14 (3–4): 433–443. Бибкод : 1981BiSys..14..433R . дои : 10.1016/0303-2647(81)90048-4 . ПМИД 6175358 .
- ^ Перейти обратно: а б с д Фрейденталь, Гюго (1962). « Symbiodinium gen. nov. и Symbiodinium microadriaticum sp. nov., зооксантелла: таксономия, жизненный цикл и морфология». Журнал протозоологии . 9 (1): 45–52. дои : 10.1111/j.1550-7408.1962.tb02579.x .
- ^ Перейти обратно: а б с д и ж г час Стил, Данбар (1975). «Этапы истории жизни симбиотической зооксантеллы в гранулах, экструдированных ее хозяином Aiptasia tagetes (Германия и Мичиган) (Coelenterata, Anthozoa)» . Биологический вестник . 149 (3): 590–600. дои : 10.2307/1540389 . JSTOR 1540389 . ПМИД 29324193 .
- ^ Тренч, РК (1997). «Разнообразие симбиотических динофлагеллят и эволюция симбиозов микроводорослей и беспозвоночных». В Лессиосе, штат ХА; Макинтайр, И.Г. (ред.). Материалы восьмого Международного симпозиума по коралловым рифам, Панама, 24–29 июня 1996 г. Том. 2. Бальбоа, Панама: Смитсоновский институт тропических исследований. стр. 1275–86. ОСЛК 833272061 .
- ^ Перейти обратно: а б с д и ж Ладриер, Офелия; Компер, Филипп; Деклу, Николь; Вандевалле, Пьер; Пуличек, Матье (2008). «Морфологические изменения зооксантелл у отбеленных книдарий-хозяев». Тетради по морской биологии . 49 (3): 215–227. hdl : 2268/6020 .
- ^ Мускатин, Л; Портер, Джеймс В. (1977). «Рифовые кораллы: мутуалистические симбиозы, адаптированные к среде с низким содержанием питательных веществ». Бионаука . 27 (7): 454–460. дои : 10.2307/1297526 . JSTOR 1297526 .
- ^ Перейти обратно: а б с д и ж Мур, Дэвид (2022), Мур, Дэвид; Хайльвек, Матиас; Петрос, Питер (ред.), «Выращивание гигантских моллюсков в 2021 году: великое будущее для «голубой экономики» тропических островов» , Аквакультура: «Голубой углерод океана соответствует ЦУР ООН» , Серия «Цели устойчивого развития», Cham: Springer International Publishing, стр. 131–153, doi : 10.1007/978-3-030-94846-7_5 , ISBN. 978-3-030-94846-7 , получено 25 апреля 2022 г.
- ^ Фитт, В.; Рис, Т.; Йеллолис, Д. (1995). «Связь между pH и наличием растворенного неорганического азота в симбиозе зооксантеллы и гигантского моллюска» . Лимнология и океанография . 40 (5): 976–982. Бибкод : 1995LimOc..40..976F . дои : 10.4319/LO.1995.40.5.0976 . S2CID 14951608 .
- ^ Фанкбонер, П.В. (1 января 1981 г.). «Сифональные глаза гигантских моллюсков и их связь с соседними зооксантеллами» . Велигер . 23 (3). ОСТИ 6701276 .
- ^ Перейти обратно: а б Лонг, Чао; Чжан, Юэхуань; Ли, Юньцин; Ли, Цзюнь; Чжоу, Цзихуа; Цинь, Яньпин; Ли, Синъю; Ма, Хайтао; Вэй, Цзинькуань; Чжоу, Иньинь; Нур, Зохаиб (2021). «Влияние филотипов Symbiodiniaceae в кладах A – E на потомство двух гигантских моллюсков ( Tridacna squmosa и T. crocea ) на ранних этапах жизни в Южно-Китайском море» . Границы морской науки . 8 : 633761. дои : 10.3389/fmars.2021.633761 .
- ^ Перейти обратно: а б с д и ж г час я дж к Джегри, Николас; Пондавен, Филип; Стибор, Хервиг; Доусон, Майкл Н. (2012). «Обзор разнообразия, особенностей и экологии зооксантелловых медуз» . Морская биология . 166 (11): 147. Бибкод : 2019Вторник.166..147D . дои : 10.1007/s00227-019-3581-6 . S2CID 208553146 .
- ^ Уикс, Челси; Мигер, Шон; Виллинк, Филип; МакКрави, Кеннет В. (сентябрь 2019 г.). «Влияет ли подкисление морской воды на плотность и здоровье зооксантелл у инвазивных перевернутых медуз Cassiopea spp.?» . Биология беспозвоночных . 138 (3). дои : 10.1111/ivb.12255 . ISSN 1077-8306 . S2CID 202017643 .
- ^ Перейти обратно: а б Мортильяро, Дж. М.; Питт, Калифорния; Ли, С.Ю.; Мезиан, Т. (30 сентября 2009 г.). «Интенсивность света влияет на выработку и перемещение жирных кислот зооксантеллами медузы Cassiopea sp» . Журнал экспериментальной морской биологии и экологии . 378 (1): 22–30. дои : 10.1016/j.jembe.2009.07.003 .

| Базы данных органов управления : Национальные |
|---|